

Abstract
Cross-resistance, and longevity of resistance, induced by the bean leaf beetle, Cerotoma trifurcata, was studied IN the soybean PI 227687 that exhibited induced response in earlier studies. Bean leaf beetle adults and soybean looper, Pseudoplusia includens, larvae were used to induce resistance and to determine beetle feeding preference. Beetles were collected from soybean fields 2 to 5 days prior to the feeding preference test. The level of cross-resistance induced by soybean looper herbivory to subsequent bean leaf beetle feeding was higher when compared to cross-resistance induced by bean leaf beetle herbivory against subsequent feeding by soybean looper. Further, herbivory by the bean leaf beetle also induced resistance against soybean looper feeding. In the longevity study, leaflets from treated plants were collected 5, 10, 12, 14, 16, 20 and 25 days after initiation of feeding. Pairwise comparisons of leaflets from plants treated by bean leaf beetle herbivory with untreated plants revealed that induced responses were highest 14 and lowest 25 days after initiation of feeding. On other sampling days, levels of induced response varied with the sampling day.
Keywords: soybean, induced resistance, longevity, cross-resistance
Introduction
Kogan and Paxton (1983) defined induced resistance as the qualitative and/or quantitative enhancement of a plant's defense mechanisms against pests in response to extrinsic physical or chemical stimuli. From the perspective of herbivore population dynamics, induced resistance is any change in a plant resulting from damage that imparts a negative effect on herbivores (Karban and Baldwin 1997). Insect herbivory has been shown to induce resistance in soybean plants (Lin et al., 1990b, Fischer et al., 1990, Lin and Kogan 1990) against the soybean looper, Pseudoplusia includens and the Mexican bean beetle, Epilachna varivestis.
The bean leaf beetle, Cerotoma trifurcata, is a sporadic pest of soybean in Nebraska, but has increased in importance primarily because of the rapid increase in soybean acreage. In regions where both P. includens and C. trifurcata are present, P. includens attacks plants after C. trifurcata (Hammond et al., 1991). For our purposes we define cross-resistance as resistance to one species of insect induced by another species of insect and vice versa. Therefore, cross-resistance to P. includens due to previous C. trifurcata injury in the form of induced resistance would be beneficial to the plant. In an earlier study by Felton et al. (1994), bean leaf beetle herbivory affected larval growth rates and reduced the suitability of foliage to the corn earworm, Helicoverpa zea, in soybean. Inbar et al., (1998) demonstrated the effectiveness of the chemical injury inducer, Actigard™ (Novartis Crop Protection Inc., Greensboro, NC), in providing cross-resistance to various insect pests and pathogens in the tomato, Lycopersicon esculentum. However, no studies have reported cross-resistance in soybean to one insect species after induction by another insect's herbivory and vice versa.
Earlier studies (Srinivas et al., in press) indicated that soybean induced resistance to C. trifurcata can be caused either by C. trifurcata feeding, or P. includens feeding. Foliar application of Actigard™ was also effective. However, the literature on the longevity or duration of protection offered by induced resistance is very limited. Earlier research in host-pathogen systems documented that protection could be effective for a week or more (McIntyre and Dodds 1979, Chaudary et al., 1983). Recent reports on the duration of induced resistance in host-pathogen systems have been documented in barley (Pelcz and Wolffgang 1986), cotton (Liu et al., 1990) and cucumber (Dalisay and Kuc 1995). Underwood (1998) demonstrated that induced resistance against E. varivestis lasted three days after damage in soybean. Stout et al., (1996b) reported persistence of proteinase inhibitors and oxidative enzymes like polyphenol oxidase, peroxidase and lipoxygenase for at least 21 days after induction in damaged tomato leaflets. Thus far, no studies have been conducted to demonstrate the longevity of bean leaf beetle induced resistance in soybean.
The objectives of this study were to investigate: 1) cross-resistance to P. includens feeding after C. trifurcata herbivory, 2) cross-resistance to C. trifurcata feeding after P. includens herbivory and 3) the longevity of induced resistance due to C. trifurcata feeding in soybean against subsequent C. trifurcata feeding.
Materials and Methods
Seeds of soybean, Glycine max (L.) Merrill, PI 227687 were planted in 15 cm plastic pots, in a sterile soil mixture (consisting of a 3:2:1 mixture of top soil:sand:vermiculite) and grown in a greenhouse maintained at 30±5°C, 70±10% RH, and a14:10 (L:D) photoperiod with supplementary metal halide illumination. The developmental stages of soybean plants were defined as described by Fehr et al., (1971).
Adult been leaf beetle C. trifurcata (Forster) were collected from fields at University of Nebraska-ARDC in Saunders County, NE and from fields at the University of Nebraska East Campus and Havelock farms in Lancaster County, NE. Soybean looper, Pseudoplusia includens (Walker), larvae were obtained from a colony maintained on artificial diet at the USDA-ARS, Southern Field Crops Insect Laboratory at Stoneville, MS.
Cross-Resistance
Treatments of C. trifurcata feeding, P. includens feeding, and untreated (control) plants were applied (one plant per pot) at the V6 (vegetative stage with six nodes) stage of growth in a completely randomized fashion. All the potted plants (including the controls) were covered with cages (radius-7.62 cm and height-60.96 cm) constructed of clear Lexan® FR-60 film (GE Plastics-Polymers, Pittsfield, MA), fitted with vents for aeration and handling of the insects. There were a total of 18 replications in this study and each potted plant was considered an experimental unit. 25–30% defoliation was produced by placing three or four fourth and fifth instar P. includens larvae or six-to-eight C. trifurcata adults, on first five trifoliates for 24–48 h. The apical trifoliates were used for C. trifurcata feeding preference tests 2 weeks after plants were injured.
Longevity of Induced Resistance
In the longevity study, treatments of C. trifurcata feeding and untreated (control) were applied randomly to soybean plants at V6 stage (one plant per pot). All the potted plants (including the control plants) were covered with cages as described above and each caged plant was considered an experimental unit. Five plants each were sampled on 5, 10, 12, 14, 16, 20 and 25 days after herbivory (DAH) and five more from uninjured control plants. There were ten replications in this study. Defoliation (25–30%) by C. trifurcata was produced by placing six-to-eight adults on the first top five trifoliates for 24 h. The sixth trifoliate or the top trifoliates were used for C. trifurcata feeding-preference tests. Only beetles collected within the previous 2 weeks were used for these tests.
Dual-choice tests were conducted to assess C. trifurcata and P. includens feeding preferences on induced plants. Soybean looper and C. trifurcata feeding were treatments in the cross-resistance study and C. trifurcata feeding was the treatment in the longevity study. Six leaflet disks from each pair of experimental plants, assigned as control (C) and treatment (T), were obtained and arranged in an alternating pattern around the bottom of a Petri dish (100 × 15 mm). In these tests, adult C. trifurcata were starved and supplied with only water for 24 h prior to the tests. Four starved adult C. trifurcata were released into each Petri dish, allowed to feed for 4 to 6h, and the remaining leaflet disk area was measured using a LICOR-3000 area meter (LI-COR, Lincoln, NE). In the cross-resistance study, feeding preferences of 5th instar P. includens larvae were determined by allowing larvae that had been pre-starved for 2h before the test to feed for 4 to 6h. Differences in consumed treatment and control leaflet disk area were used to compute the statistical significance of differences in feeding preferences and to calculate the feeding-preference index (PI), where PI = 2T/(T + C) (Kogan and Goeden 1970, Kogan 1972). The PI values ranged from 0 to 2, with PI=1 indicating no feeding preference for either control or treatment disks, PI > 1 indicating preference for treatment disks, and PI < 1 indicating preference for control leaflet disks. Data were analyzed using PROC GLM procedure (SAS Institute 1997) and means were compared using the LSD (Least Significance Difference) computed at α = 0.05.
Results
The acceptability of leaflet disks between treated and untreated plants during feeding was taken as the effect of treatments on C. trifurcata and P. includens feeding. These differences are taken as variations in resistance levels in comparison with constitutive resistance (Lin et al., 1990b).
Cross-Resistance
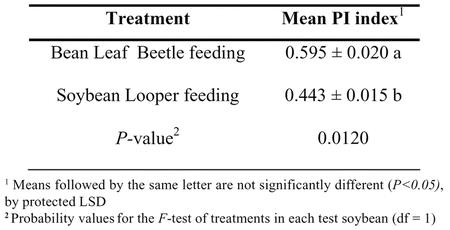
Results from the experiments to demonstrate induced cross-resistance in PI 227687 showed that different levels of cross-resistance were induced by C. trifurcata and P. includens herbivory (Table 1). The initial herbivory of C. trifurcata induced cross-resistance to subsequent P. includens feeding. However, leaflets from plants treated with P. includens feeding induced higher cross-resistance to subsequent C. trifurcata feeding where the mean PI was 0.443. Cross-resistance induced by P. includens feeding against subsequent C. trifurcata feeding, was statistically significant (F Value = 7.05 and Pr > F = 0.0120).
Table 1.
Mean (± SE) feeding preference index (PI) values for soybean looper and bean leaf beetle fed soybean PI 227687 leaflets expressing cross resistance induced by bean leaf beetle and soybean looper feeding, respectively.
Longevity of Induced Resistance
C. trifurcata injury-induced resistance in soybean PI 227687 to subsequent feeding by C. trifurcata varied throughout the duration of experiment (Fig. 1 and Table 2). The induced response was numerically highest at 14 DAH (PI = 0.573) compared to all other sampling days and was not significantly different from 12 and 16 DAH. The feeding preference was lowest at 25 DAH (0.812) and was not significantly different from 5 and 20 DAH. The PI at 10 DAH was not significantly different from all the other sampling days except 14 DAH. The longevity of induced resistance varied between sampling days after treatment (F Value = 4.19 and Pr > F = 0.0040) indicating significant difference in the longevity of the induction response. Since the data have a quantitative variable, regression analysis was done using PROC REG. The data did not fit the linear model (F value = 1.39 and Pr > F = 0.2469), but did fit the quadratic model (F value = 9.469 and Pr > F = 0.0006). This indicates that the induced resistance increased for a certain period of time and then decreased.
Figure 1.
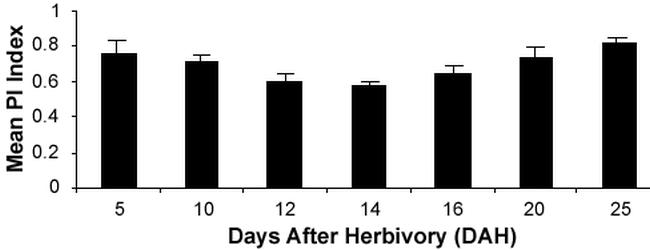
Mean feeding preference (PI) indices (± S.E.) of bean leaf beetles in dual-choice feeding preference tests 5, 10, 12, 14, 16, 20 and 25 days after herbivory on PI 227687 leaflets from control plants and plants exposed to bean leaf beetle feeding. Mean PI values were derived from 10 replicates of each test combination. Means followed by the same letter are not significantly different (p<0.05) by protected LSD.
Table 2.
Mean (± SE) feeding preference index (PI) values for different sampling days of bean leaf beetle fed soybean PI 227687 leaflets expressing induced response by bean leaf beetle feeding.

Discussion
Induced resistance to herbivores, particularly arthropods, has been observed in soybean, tomato, potato, wheat and cotton (Kogan and Paxton 1983, Stout et al., 1996a, Karban and Baldwin 1997). The response of soybean to various biotic and abiotic stresses that induce resistance may involve the production of natural products, phytoalexins or plant pathogenesis-related proteins (Kogan and Paxton 1983, Benhamou and Theriault 1992, Ebel and Cosio 1994). This phenomenon is not well understood, and its persistence in deterring herbivory after elicitation has been documented in only a few insect-plant systems.
Previous studies have documented cross-resistance in other host-pest systems including tomato and potato (Inbar et al., 1998, Singh and Singh 1995). In our experiments, P. includens feeding elicited a higher level of cross-resistance against C. trifurcata feeding than C. trifurcata induction of cross-resistance against P. includens feeding. This is the first report of C. trifurcata-induced cross-resistance to P. includens in soybean. However, Felton et al. (1994) reported the effect of C. trifurcata feeding on corn earworm. Similarly, innoculum containing the pathogen, Phytophthora megasperma induced phytochemical defenses and deterred E. varivestis feeding on soybean (Kogan and Fischer 1991). Inbar et al., (1998) reported cross-resistance in tomato plants where there was a significant reduction of leafminer, Liriomyza spp., larval densities after treatment with Actigard™.
This is the first report of the longevity of C. trifurcata-induced resistance in soybean. Bentur and Kalode (1996) demonstrated induced resistance in rice to the Asian rice gall midge, Orseolia oryzae, to be effective for about 4 weeks after infestation. This research establishes that soybean induced resistance from C. trifurcata feeding is effective for approximately the same duration, although the induction effect began to subside at about 14 days after the first initial feeding by C. trifurcata. In an earlier study by Underwood (1998), the induced response was effective for three days after damage against E. varivestis in soybean. Resistance diminished by 15 days after damage and by 20 days all four genotypes used in the studies exhibited significant induced susceptibility. No such susceptibility was observed in our experiments although the induced response decreased to 0.81 (PI) by 25 DAH. The longevity of induced resistance produced by C. trifurcata was two weeks in our study. Any other type of injury might produce a higher level of induced resistance and protection that lasted longer than two weeks because Lin et al., (1990a, b) demonstrated that the level of induced soybean resistance varied with the type of injury. Thus, if the level of injury is higher and stronger it might offer longer protection.
Induction enhances soybean resistance to insects and pathogens (Lin and Kogan 1990, Dann et al., 1998, Thaler 1999). Induced responses produce volatiles that act as a direct defense against the herbivores (Karban and Baldwin 1997) and as an attractant to parasitoids in the form of indirect defense (Turlings et al. 1995, Dicke and Loon 2000). Moreover, induced responses were also shown to affect the herbivore's feeding preference, growth rate and fecundity that would influence the dynamics of the population (Karban and Baldwin 1997). This may have an indirect effect on population dynamics of natural enemies in the crop ecosystems. The longevity of the induced response offers longer protection against herbivory in pest management. The presence of induced resistance for a longer time extends the possibility of any direct or indirect effects on natural enemies and other non-target arthropods/herbivores. Thus these possible long-term effects of induced resistance should be studied at a tritrophic and possibly multitrophic level of plant-insect interactions.
If induced resistance reduces pest damage to plants, then chemical elicitors such as Actigard™, can be used to induce resistance as a preventive practice and extend the duration of plant protection. However, cross-resistance may act as an important plant protectant in addition to application of chemical elicitors. If a pest population can induce resistance to other pests in the field, a useful management tool could be to monitor the insect population that has the potential of inducing cross-resistance to other pests emerging either at the same time or later. Future research should focus on defining the longevity of soybean induced responses in the field, its effects on the population dynamics of pest insects and also to study the possibilities of extending this longevity using chemical elicitors.
Acknowledgments
We thank Dr. Fred Baxendale and Dr. Thomas E. Hunt for reviewing the manuscript. We also thank Dr. Leon Higley for use of the leaf area meter and Dr. Annaboyan Kessavolu for his help with the statistical analysis. We appreciate the technical support given by Stephanie Wreed during the course of this research project. This is article 13011 of the journal series of the Nebraska Agricultural Research Division, University of Nebraska-Lincoln. Research partially supported by the Interdisciplinary Farm Project of the Agricultural Research and Development Center of the University of Nebraska
Glossary
| Abbreviation: | |
|---|---|
| DAH | days after herbivory |
References
- Bentur JS, Kalode MB. Hypersensitive reaction and induced resistance in rice against the Asian rice gall midge Orseolia oryzae. Entomol. Exp. Appl. 1996;78:77–81. [Google Scholar]
- Benhamou N, Theriault G. Treatment with chitosan enhances resistance of tomato plants to the crown and root rot pathogen Fusarium oxysporum f. sp. radicis-lycopersici. Physiol. Mol. Plant Pathol. 1992;41:33–52. [Google Scholar]
- Chaudary RC, Schwarzbach E, Fischbeck G. Quantative studies of resistance induced by avirulent cultures of Erysiphe graminis f.sp. hordei in barley. Phytopathol. Zeitschrift. 1983;108:80–87. [Google Scholar]
- Dalisay RF, Kuc JA. Persistence of induced resistance and enhanced peroxidase and chitinase activities in cucumber plants. Physiol. Plant Pathol. 1995;47:315–327. [Google Scholar]
- Dann E, Diers B, Byrum J, Hammerschmidt R. Effect of treating soybean with 2,6-dichloroisonicotinic acid (INA) and benzothiadiazole (BTH) on seed yields and the level of disease caused by Sclerotinia sclerotorium in field and greenhouse studies. European J. Plant Pathol. 1998;104:271–278. [Google Scholar]
- Dicke M, van Loon JJA. Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context. Entomol. Exp. Appl. 2000;97:237–249. [Google Scholar]
- Ebel J, Cosio EG. Elicitors of plant defense responses. Int. J. Rev. Cytol. 1994;148:1–36. [Google Scholar]
- Fehr WR, Caviness CE, Burmood DT, Pennington JS. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971;11:929–931. [Google Scholar]
- Felton GW, Summers CB, Mueller AJ. Oxidative responses in soybean foliage to herbivory by bean leaf beetle and three-cornered alfalfa hopper. J. Chem. Ecol. 1994;20(3):639–650. doi: 10.1007/BF02059604. [DOI] [PubMed] [Google Scholar]
- Fischer DC, Kogan M, Paxton J. Effect of glyceollin, a soybean phytoalexin, on feeding by three phytophagous beetles (Coleoptera: Coccinellidae and Chrysomelidae): dose vs. response. Environ. Entomol. 1990;19:1278–1282. [Google Scholar]
- Hammond RB, Higgins RA, Mack TP, Pedigo LP, and Bechinski EJ. 1991 Soybean pest management. In: David Pimentel, editor. CRC Handbook of pest management in agriculture. 3:341–474.CRC press, Boca Raton, FL. [Google Scholar]
- Inbar M, Doodstar H, Sonoda RM, Leibee GL, Mayer RT. Elicitors of plant defensive systems reduce insect densities and disease incidence. J. Chem. Ecol. 1998;24:135–149. [Google Scholar]
- Karban R, Baldwin IT. 1997 Induced responses to herbivory. The University of Chicago Press. Chicago, IL. [Google Scholar]
- Kogan M. Feeding and nutrition of insects associated with soybeans. 2. Soybean resistance and host preferences of the Mexican bean beetle, Epilachna varivestis. Ann. Entomol. Soc. Am. 1972;65:675–683. [Google Scholar]
- Kogan M, Goeden RD. The host plant range of Lema trilineata daturaphila (Coleoptera: Chrysomelidae) Ann. Entomol. Soc. Am. 1970;63:1175–1180. [Google Scholar]
- Kogan M, Fischer DC. 1991 Inducible defenses in soybean against herbivorous insects. In: Tallamy DW, Raupp MJ, editors, Phytochemical induction by herbivores, Pp. 347–378.John Wiley and Sons, Inc. [Google Scholar]
- Kogan M, Paxton J. 1983 Natural inducers of plant resistance to insects. In: Hedin PA, editor. Plant resistance to insects. American Chemical Society Symposium. Series. 208:153–171.American Chemical Society, Washington, DC. [Google Scholar]
- Lin HC, Kogan M. Influence of induced resistance in soybean on the development and nutrition of the soybean looper and the Mexican bean beetle. Entomol. Exp. Appl. 1990;55:131–138. [Google Scholar]
- Lin HC, Kogan M, Endress AG. Influence of ozone on induced resistance in soybean to the Mexican bean beetle (Coleoptera: Coccinellidae) Environ. Entomol. 1990a;19:854–858. [Google Scholar]
- Lin HC, Kogan M, Fischer D. Induced resistance in soybean to the Mexican bean beetle (Coleoptera: Coccinellidae): Comparisons of inducing factors. Environ. Entomol. 1990b;19:1852–1857. [Google Scholar]
- Liu XZ, Chen CY, Wang CJ. Selection of inducer of resistance to cotton Fusarium wilt and the optimal interval duration. Acta Phytopathol. Sinica. 1990;20:123–126. [Google Scholar]
- McIntyre J, Dodds JA. Induction of localized and systemic protection against Phytophthora parasitica var. nicotianae by tobacco mosaic virus of tobacco, hypersensitive to the virus. Physiol. Plant Pathol. 1979;15:321–330. [Google Scholar]
- Pelcz J, Wolffgang H. Duration of induced resistance and susceptibility changes in the host-parasite combination barley/powdery mildew (Hordeum vulgare L./Erysiphe graminis f. sp. hordei March.) Arch. Phytopathol. Plant Prot. E. Ger. 1986;22:459–464. [Google Scholar]
- Singh M, Singh RP. Host dependent cross-protection between PVYN, PVYO, and PVA in potato cultivars and Solanum brachycarpum. Can. J. Plant Pathol. 1995;17:82–86. [Google Scholar]
- SAS Institute. 1997 SAS/STAT user's guide. version 6.12. SAS Institute, Cary, NC. [Google Scholar]
- Srinivas P, Danielson SD, Smith CM, and Foster JE. In press. Induced resistance to bean leaf beetle, Cerotoma trifurcata (Forster) (Coleoptera: Chrysomelidae), in soybean. J. Entomol. Sci. [Google Scholar]
- Stout MJ, Workman J, Duffey SS. Differential induction of tomato foliar proteins by arthropod herbivores. J. Chem. Ecol. 1996a;20:2575–2594. doi: 10.1007/BF02036193. [DOI] [PubMed] [Google Scholar]
- Stout MJ, Workman KV, Workman JS, Duffey SS. Temporal and ontogenic aspects of protein induction in foliage of the tomato, Lycopersicon esculentum. Biochem. Syst. Ecol. 1996b;24(7/8):611–625. [Google Scholar]
- Thaler JS. Induced resistance in agricultural crops: Effects of jasmonic acid on herbivory and yield in tomato plants. Environ. Entomol. 1999;28:30–37. [Google Scholar]
- Turlings TCJ, Loughrin JH, McCall PJ, Röse USR, Lewis WJ, Tumlinson JH. How caterpillar damaged plants protect themselves by attracting parasitic wasps. Proc. Natl. Acad. Sci. USA. 1995;92:4169–4174. doi: 10.1073/pnas.92.10.4169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Underwood NC. The timing of induced resistance and induced susceptibility in the soybean-Mexican bean beetle system. Oecologia. 1998;114:376–381. doi: 10.1007/s004420050460. [DOI] [PubMed] [Google Scholar]