

Abstract
The activity of juvenile hormone esterase (JHE) in feeding fifth instar larvae of Manduca sexta increases gradually with larval weight and rises to a peak after larvae pass the critical weight when juvenile hormone secretion ceases. Starvation of larvae of Manduca sexta (L.) that had exceeded the critical weight inhibited peak levels of JHE, but did not delay entry into the wandering stage when larvae leave the plant in search of a pupation site. This suggests that peak levels of JHE may not be essential for the normal timing of metamorphosis. Starved larvae pupated normally, indicating the peak of JHE was not necessary for a morphologically normal pupation. Treatments of larvae with the selective JHE inhibitor O-ethyl-S-phenyl phosphoramidothiolate (EPPAT) that began immediately after larvae achieved the critical weight (6.0 to 6.5 grams for our strain of Manduca) delayed entry into the wandering stage. By contrast, EPPAT treatment of larvae at weights above 8.0g had no effect on the subsequent timing of the onset of wandering. Therefore, although the normal timing of the onset of wandering does not require peak levels of JHE, it requires low to moderate levels of JHE to be present until larvae reach a weight of about 8.0g.
Keywords: juvenile hormone, juvenile hormone esterase, metamorphosis, Manduca sexta, starvation, critical weight
Introduction
The metamorphosis of insects is controlled by juvenile hormone (JH). In the presence of JH insect tissues maintain their developmental status quo when exposed to ecdysteroids (the molting hormones), whereas in the absence of JH (or when the JH titer is below a tissue-specific threshold), tissues change their commitment to that of the next metamorphic stage upon exposure to ecdysteroids (Riddiford, 1996; Nijhout, 1994). Regulation of the JH titer thus plays a critical role in the control of metamorphosis. Regulation of JH is accomplished by the independent regulation of its synthesis by allatotropic and allatostatic factors, and its metabolism by catabolic enzymes. Juvenile hormone esterase (JHE) plays a key role in the metabolism of JH, and has long been established as a negative regulator of the JH titer (Sparks et al., 1983; Hammock, 1985). The activity of JHE in the hemolymph of the tobacco hornworm, Manduca sexta, rises dramatically during the latter half of the final larval instar. This so-called pre-wandering peak of JHE activity is believed to be essential for normal metamorphosis (Weirich et al., 1973; Sanburg et al., 1974; Vince and Gilbert, 1976; Sparks et al., 1983; Jesudason et al., 1990).
The initiation of metamorphosis in Manduca sexta is stimulated by the disappearance of JH and subsequent secretion of the prothoracicotropic hormone (PTTH) that stimulates the production of ecdysteroids, which induce the wandering stage (when larvae leave the plant and search for a pupation site) and cause the reprogramming of the epidermis from larval commitment to pupal commitment (Truman et al., 1974; Nijhout, 1994). During the last larval instar the secretion of PTTH and ecdysteroids is inhibited by JH (Nijhout and Williams, 1974a,b; Rountree and Bollenbacher, 1986). It is believed that JHE plays a role in the clearance of JH from the hemolymph after secretion of JH ceases (Sparks et al., 1983; Baker et al., 1987). Treatment of M. sexta larvae with the JHE inhibitor O-ethyl-S-phenyl phosphoramidothiolate (EPPAT) early in the final instar significantly delays entry into the wandering stage, presumably by preventing or delaying the degradation of JH which, in turn, continues to inhibit the secretion of PTTH and ecdysteroids (Sparks et al., 1983). Moreover, when larvae are starved early in the final instar, their level of JHE activity is depressed and such larvae experience a delayed entry into the wandering stage (Cymborowski et al., 1982; Sparks et al., 1983). In these experiments larvae were starved before they reached the critical weight, and this is believed to prevent cessation of JH secretion and hence prolong the instar (Cymborowski et al., 1982).
The critical weight defines the time at which the secretion of JH ceases (Nijhout and Williams, 1974a,b). If a larva is starved before it reaches the critical weight, it continues to secrete JH for an abnormally long time and the secretion of PTTH and entry into the wandering stage are correspondingly delayed. By contrast, if a larva is starved after it has reached the critical weight, the timing of the wandering stage and metamorphosis occur at exactly the same time as in larvae that are allowed to continue to feed (Nijhout and Williams, 1974a,b). The critical weight thus marks a transition point after which the time course to metamorphosis is unaffected by further feeding and growth of the larva (Nijhout, 1994). Exogenous JH, applied after a larva reaches the critical weight, can greatly delay PTTH secretion and the onset of wandering (Nijhout and Williams, 1974b). The absence of JH after a larva reaches the critical weight is thus essential not only for a normal metamorphosis, but also for a normal time course to metamorphosis. The pre-wandering peak of JHE activity presumably serves to clear any remaining traces of JH from the hemolymph after a larva reaches its critical weight and has ceased to secrete JH.
Although starvation of a larva before the critical weight severely depresses JHE activity, it is not clear whether this is also true after a larva has reached the critical weight. Insofar as starvation after the critical weight does not affect the timing of entry into the wandering stage, it appears that starvation does not interfere with the normal pattern of JH degradation during that time. To investigate the role of JHE during the period between achieving the critical weight and the onset of wandering, we starved larvae of M. sexta at the critical weight and monitored the subsequent profile of JHE activity. We found that starvation completely eliminated the pre-wandering peak of JHE activity. We also found, however, that a low level of JHE activity during a brief period immediately after the critical weight is essential for a normal time course to PTTH secretion and wandering.
Materials and Methods
Larvae of Manduca sexta were reared on an artificial diet at 27°C on a 16:8 L:D photoperiod. All hemolymph samples used in these experiments were taken between 6 and 12 hours after lights-on. The critical weight of 5th instar larvae in our colony (defined as the weight at or above which starvation no longer affects the normal time course to metamorphosis) was determined by D'Amico et al. (2001) and was found to be 6.0 to 6.5 grams. The final weight of normally-feeding larvae in our colony is 11 to 12 grams.
In order to test the effect of starvation on JHE activity we removed larvae from food when they reached a weight between 6.5 and 7.5 grams, that is, shortly after they passed the critical weight. A single hemolymph sample was taken from each experimental starved larva and used for determination of JHE activity. Control starved larvae were monitored daily and the timing of their entry into the wandering stage was recorded.
Juvenile hormone esterase (JHE) activity in the hemolymph was determined by means of the partition assay described by Hammock and Roe (1985), and expressed as nanomoles of JH-acid produced per minute per ml of hemolymph. 3H juvenile hormone III (717.80 GBq/mmol) was purchased from New England Nuclear. The JHE inhibitor O-ethyl-S-phenyl phosphoramidothiolate (EPPAT) was kindly provided by Dr. Bruce Hammock (University of California at Davis). EPPAT treatment consisted of topical applications of 2 microliters of a solution of 200 nM (in absolute ethanol) (Sparks et al., 1983). Treatments were applied twice daily, beginning at various weights and continuing for two days. All EPPAT-treated larvae continued to feed and grow normally, suggesting the larvae did not experience any toxicity effects from the treatment.
Results
The profile of JHE activity in final (5th) instar larvae is shown in Figure 1. The activity profile is similar to that described previously by Jones et al. (1982) and Jesudason et al. (1990), except that we obtained a higher temporal resolution than these authors by measuring the JHE activity at intervals of 1 gram along the entire growth trajectory of the 5th instar. During the feeding phase, JHE activity increased gradually with larval weight. The JHE activity declined rapidly when larvae purged their gut and entered the wandering stage, and rose again just prior to pupation. The pre-wandering peak of JHE activity was not clearly separated from the gradual rise that occurred with increasing weight.
Figure 1.
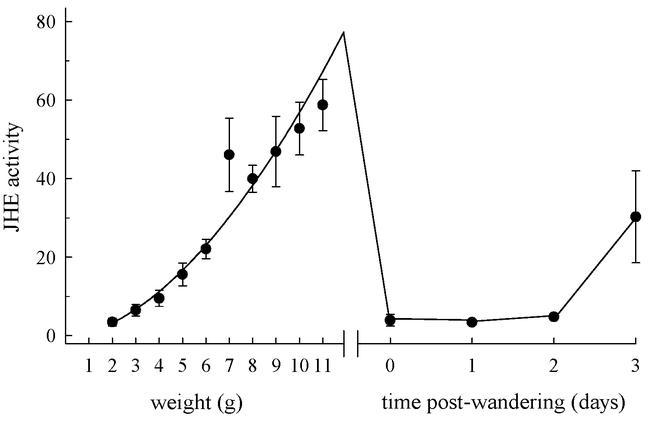
The juvenile hormone esterase (JHE) activity profile of normally feeding last (5th) instar larvae of Manduca sexta. JHE activity was measured as nmoles of JH-acid produced per minute per ml of hemolymph. Each point is the mean of 9 to 11 pre-wandering larvae, or 4 to 6 post-wandering larvae. Vertical bars are standard errors of the mean (which in some cases do not exceed the size of the symbol). The abscissa is a measure of the physiological age of the larva, which is given on two scales: before entry into the wandering stage the larva's physiological age is given by its weight, and after entry into the wandering stage it is given in days. The curve represents the best-fit polynomial regression (2nd degree polynomial, r2 = 0.99), and is extrapolated to animals weighing 12 grams (the maximal weight at which larvae entered the wandering stage).
In order to test the effect of starvation on JHE activity we removed larvae from food when they reached a weight between 6.5 and 7.5 grams, that is, shortly after they passed the critical weight (Figure 2). JHE activity declined rapidly after the initiation of starvation and did not rise to the peak levels characteristic of the pre-wandering phase of the final instar (c.f. Figure 1). The absence of the pre-wandering peak of JHE had no effect on the timing of onset of wandering. Feeding larvae entered the wandering stage an average of 56 ± 2.3 (mean ± standard error) hours after passing 6.5 to 7.5 grams (n = 37), whereas larvae that were starved in this weight range entered the wandering stage an average of 53.4 ± 2.0 hours later (n = 26). Starved larvae metamorphosed into normal pupae, indicating that the pre-wandering peak of JHE activity was not necessary for a morphologically normal pupation. After entry into the wandering stage the level of JHE activity rose in the same way in larvae that had been starved as well as those that had been allowed to feed normally (Figure 2).
Figure 2.
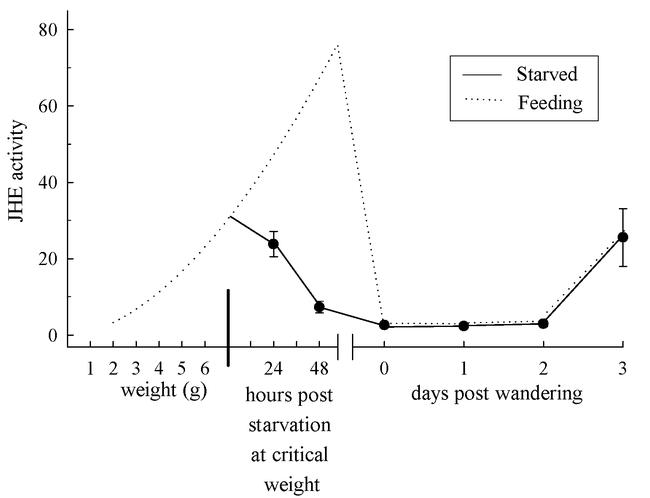
The effect of starvation after the critical period on the JHE activity profile of 5th instar Manduca sexta larvae. JHE activity was measured as nmoles of JH-acid produced per minute per ml of hemolymph. Each point is the mean of 7 to 9 pre-wandering larvae, or 4 to 6 post-wandering larvae. Vertical bars are standard errors of the mean (which in some cases do not exceed the size of the symbol). Larvae were starved at weights of 7 ± 0.5 grams (dark vertical bar), and JHE activity was measured at 24-hour intervals thereafter. The expected JHE activity of normally feeding larvae (from Fig. 1) is indicated by the dotted line.
In view of the fact that JHE activity had already risen to a low but appreciable level by the time larvae reached the critical weight, we investigated whether this low level of JHE activity had a significant physiological consequence. Feeding larvae were treated with twice daily doses of EPPAT, beginning at successive times after they had reached the critical weight. We found that EPPAT treatments that began at weights between 6.0g and 7.5g delayed entry into the wandering stage by 1 to 2 days. By contrast, treatments that began at weights above 8.0g had no effect on the timing of the onset of wandering (probability values are shown in Figure 3). Evidently, inhibiting of JHE during the period immediately after larvae passed the critical weight delayed entry into the wandering stage, suggesting that its activity during this period is important for the normal metabolism of JH.
Figure 3.
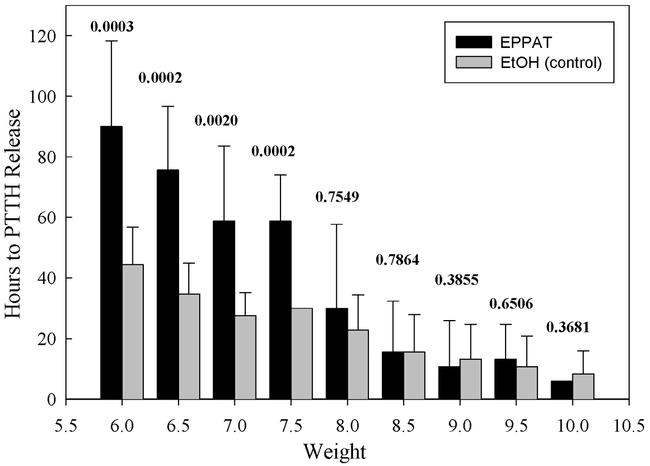
Effect of EPPAT treatment on the timing of entry into the wandering stage of normally feeding Manduca sexta larvae. The abscissa indicates the weight at which EPPAT treatment began. P-values above each set indicate the probability that experimental and control values are identical (Wilcoxon's signed-ranks test; N = 10 for each bar). Vertical lines are standard deviations.
Discussion
The activity of JHE in normally feeding fifth instar larvae of M. sexta increased gradually with larval weight. When larvae were starved soon after reaching the critical weight, the JHE activity in their hemolymph declined rapidly and the pre-wandering peak of JHE failed to occur (Figure 2). In view of the fact that the onset of wandering was not delayed in these larvae, and that they metamorphosed into normal pupae, it appears that the high pre-wandering peak of JHE is not required for the normal timing of metamorphosis, nor for a morphologically normal metamorphosis.
Although starvation inhibits the pre-wandering peak of JHE activity, it has no effect on the post-wandering rise of JHE activity. The post-wandering elevation of JHE activity may be important for metabolizing the brief peak of JH that occurs after entry into the wandering stage (Kiguchi and Riddiford, 1978; Nijhout, 1994). This brief peak of JH is important for the normal metamorphosis of the imaginal disks (Nijhout, 1994), but it is not known whether the reduction or elimination of this post-wandering peak of JHE would affect the normal time course of metamorphosis.
Initial work on the biology of JHE suggested that it might play a key role in the regulation of JH titers by modulating the rate at which this hormone is metabolized (Hammock, 1985). The finding that the peak of JHE activity in the hemolymph of Manduca occurred only after the JH titer had already declined led instead to the suggestion that the primary function of JHE was to “scavenge” remaining traces of JH in preparation for metamorphosis (Baker et al., 1987; De Kort and Granger, 1996). Our results support the view that the pre-wandering peak of JHE has no function related to metamorphosis, because in its absence metamorphosis occurs normally.
Our results suggest, however, that the moderate level of JHE activity at the time the larvae pass the critical weight plays an important role in controlling the timing of entry into the wandering stage and hence the timing of metamorphosis. Evidently, larvae up to a weight of about 8 grams require JHE in order to begin metamorphosis on a normal time schedule. It seems reasonable to conclude that insofar as PTTH secretion by the brain is inhibited by JH during the last larval instar, the primary role of JHE at the critical weight is to enhance the elimination of JH. This function of JHE is evidently exercised before it rises to its normal pre-wandering peak. The function of the high levels of JHE activity during the remainder of the pre-wandering peak remain puzzling. It is possible that it represents a kind of “reserve capacity” to deal with extreme variations in the JH titer that might be more common in some environments than in others. Alternatively, it is possible that JHE has additional, yet undiscovered, functions unrelated the hydrolysis of JH and the phenomena studied here.
Acknowledgments
We thank Julia Bowsher, Laura Grunert, Jeff Marcus, Armin Moczek, Tomalei Vess, and Andy Yang for critical comments on drafts of the manuscript, and Bruce D. Hammock and Tonya Severson for the gift of EPPAT. This research was supported in part by grants from the National Science Foundation and the Howard Hughes Medical Institute's Research Fellows Program.
Glossary
| Abbreviation: | |
|---|---|
| EPPAT | O-ethyl-S-phenyl phosphoramidothiolate, a JH esterase inhibitor |
| JH | juvenile hormone |
| JHE | juvenile hormone esterase |
| PTTH | prothoracicotropic hormone |
References
- Baker FC, Tsai LW, Reuter CC, Schooley DA. In vivo fluctuations of JH, JH acid, and ecdysteroid titer, and JH esterase activity, during development of fifth stadium Manduca sexta. Insect Biochemistry. 1987;17:989–996. [Google Scholar]
- Cymborowski B, Bogus M, Beckage NE, Williams CM, Riddiford LM. Juvenile hormone titres and metabolism during starvation-induced supernumerary larval moulting of the tobacco hornworm, Manduca sexta (L.) Journal of Insect Physiology. 1982;28:129–135. [Google Scholar]
- D'Amico LJ, Davidowitz G, Nijhout HF. The developmental and physiological basis of body size evolution in an insect. Proceedings of the Royal Society, London Series B. 2001;268:1589–1593. doi: 10.1098/rspb.2001.1698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Kort CAD, Granger NA. Regulation of JH titers: The relevance of degradative enzymes and binding proteins. Archives of Insect Biochemistry and Physiology. 1996;33:1–26. [Google Scholar]
- Hammock BD. 1985 Regulation of juvenile hormone titer: Degradation. In: Kerkut GA, Gilbert LI editors. Comprehensive Insect Physiology, Biochemistry, and Pharmacology (Ed.), 7. 431–472.New York, Pergamon Press. [Google Scholar]
- Hammock BD, Roe RM. Analysis of juvenile hormone esterase activity. Methods in Enzymology. 1985;111:487–494. doi: 10.1016/s0076-6879(85)11034-7. [DOI] [PubMed] [Google Scholar]
- Jesudason P, Venkatesh K, Roe RM. Haemolymph juvenile hormone esterase during the life cycle of the tobacco hornworm, Manduca Sexta (L.) Insect Biochemistry. 1990;20:593–604. [Google Scholar]
- Jones D, Jones G, Wing KD, Rudnicka M, Hammock BD. Juvenile hormone esterases of Lepidoptera. Journal of Comparative Physiology. 1982;148:1–10. [Google Scholar]
- Kiguchi K, Riddiford LM. A role of juvenile hormone in pupa development of the tobacco hornworm, Manduca sexta. Journal of Insect Physiology. 1978;24:673–680. [Google Scholar]
- Nijhout HF. 1994 Insect Hormones. Princeton, Princeton University Press. [Google Scholar]
- Nijhout HF, Williams CM. Control of moulting and metamorphosis in the tobacco hornworm, Manduca Sexta (L.): Growth of the Last-Instar Larva and the decision to pupate. Journal of Experimental Biology. 1974a;61:481–491. doi: 10.1242/jeb.61.2.481. [DOI] [PubMed] [Google Scholar]
- Nijhout HF, Williams CM. Control of moulting and metamorphosis in the tobacco hornworm, Manduca Sexta (L.): Cessation of juvenile hormone secretion as a trigger for pupation. Journal of Experimental Biology. 1974b;61:493–501. doi: 10.1242/jeb.61.2.493. [DOI] [PubMed] [Google Scholar]
- Riddiford LM. Juvenile homone: the status quo of its “status quo” action. Archives of Insect Biochemistry and Physiology. 1996;32:271–286. doi: 10.1002/(SICI)1520-6327(1996)32:3/4<271::AID-ARCH2>3.0.CO;2-W. [DOI] [PubMed] [Google Scholar]
- Rountree DB, Bollenbacher WE. The release of prothoracicotropic hormone in the tobacco hornworm, Manduca sexta, is controlled intrinsically by juvenile hormone. Journal of Experimental Biology. 1986;120:41–58. doi: 10.1242/jeb.120.1.41. [DOI] [PubMed] [Google Scholar]
- Sanburg LL, Kramer KJ, Kezdy FJ, Law JH. Juvenile hormone-specific esterases in the haemolymph of the tobacco hornworm, Manduca Sexta. Journal of Insect Physiology. 1974;21:873–887. doi: 10.1016/0022-1910(75)90015-3. [DOI] [PubMed] [Google Scholar]
- Sparks TC, Hammock BD, Riddiford LM. The haemolymph juvenile hormone esterase of Manduca Sexta (L.)-inhibition and regulation. Insect Biochemistry. 1983;13:529–541. [Google Scholar]
- Truman JW, Riddiford LM, Safranek L. Temporal patterns of response to ecdysone and juvenile hormone in the epidermis of the tobacco hornworm, Manduca sexta. Developmental Biology. 1974;39:247–262. doi: 10.1016/0012-1606(74)90238-3. [DOI] [PubMed] [Google Scholar]
- Vince RK, Gilbert LI. Juvenile hormone esterase activity in precisely timed last instar larvae and pharate pupae of Manduca Sexta. Insect Biochemistry. 1976;7:115–120. [Google Scholar]
- Weirich G, Wren J, Siddall JB. Developmental changes of the juvenile hormone Esterase activity in haemolymph of the tobacco hornworm, Manduca sexta. Insect Biochemistry. 1973;3:397–407. [Google Scholar]