

Abstract
The swarm-founding wasp Polybia emaciata is unusual among eusocial Vespidae because it uses mud, rather than wood pulp, as its primary nest construction material. Polybia emaciata nests are more durable than similarly sized paper nests. We tested the hypothesis that the defensive behavior of this wasp may have been modified to take advantage of their strong nests in defense against vertebrate attacks. We simulated vertebrate disturbances by tapping on, and breathing in, P. emaciata nests and similarly sized P. occidentalis paper nests in the same location at the same time. Polybia emaciata responses to disturbance were qualitatively different from those of P. occidentalis. The latter exit the nest and attack, while P. emaciata workers typically fled or entered the nest, attacking only after repeated and extended disturbances. We conclude that durable nest material may permit predator avoidance behavior in P. emaciata. We compare the defensive responses of P. emaciata workers with those of other swarm-founding Vespidae, and discuss several selective forces that could cause the evolution of species variation in nest defense behavior.
Introduction
Among eusocial insects, paper wasps (Hymenoptera: Vespidae: Polistinae) are notable for the diversity of their nest architecture (Jeanne, 1975; Wenzel, 1991). Many swarm-founding polistine wasps, including most species of the Neotropical tribe Epiponini, construct envelopes around their brood combs. Comparative and experimental studies suggest that, among other functions, nest envelopes reduce rates of predation and parasitism on wasp brood (London and Jeanne, 1998; Smith et al., 2001). However, little is known about the adaptive significance of variation in the materials that wasps use to construct nest envelopes (Hansell 1984; Wenzel, 1991; Cole et al., 2001).
Polybia emaciata is unusual among the polistine wasps because it uses mud, rather than wood pulp or plant fibers, as the main raw material for nest construction (Schremmer, 1984). The genus Polybia includes 56 described species; of these, only the other three species in the subgenus Pedothoeca, and one species in the subgenus Furnariana, share the derived trait of mud nest construction (Richards, 1978; Jeanne, 1991; Cooper, 1993). No other swarm-founding polistine wasps are known to build primarily mud nests, although small amounts of inorganic material are sometimes incorporated into the nest paper of other species (Wenzel, 1991). The nest of P. emaciata is otherwise typical of the genus (Jeanne, 1975; Wenzel, 1991), being phragmocyttarus (comprising horizontal layers of comb that are conjoined and enclosed by a continuous envelope) with a single small entrance hole near the bottom of the nest. Numbers of adults in mature colonies are relatively small for the genus, ranging from fewer than 100 up to about 500 adults (Richards, 1978; Chadab, 1979; Strassmann et al., 1992). Polybia emaciata nests are hard and much more durable than most paper nests of similar size and shape (Rau, 1933; Richards, 1978). For example, Skutch (1971) noted that nests of this species in Costa Rica were unlike paper nests in that they persisted for several years. The high durability of P. emaciata construction material suggests that the nest itself may provide greater brood protection than paper nests. If so, we hypothesized that the wasps' defensive responses to vertebrate attacks have changed to take advantage of nest durability. We predicted that the nests' protective properties reduced the need for behavioral defenses by P. emaciata workers, relative to paper-nesting Polybia species with similar colony and nest sizes. We tested this prediction by disturbing P. emaciata colonies, and comparing the workers' responses to those of simultaneously observed paper-nesting P. occidentalis colonies in the same location.
Chadab (1979) surveyed the defensive responses of swarm-founding paper wasps to vertebrate-like disturbances, including tapping on the nest and/or on the supporting vegetation. The most frequently observed response was for dozens to hundreds of workers to rush out onto the nest envelope, sometimes adopting aposematic postures (see also O'Donnell et al., 1997). This “exit-nest” response was seen in the genera Angiopolybia, Charterginus, Leipomeles, Parachartergus, Polybia, Pseudopolybia, and Synoeca, and accords with our observations of what appears to be typical epiponine defensive behavior. Weaker exit-nest responses (either requiring several taps, or involving fewer wasps) were seen in species of Leipomeles, Metapolybia, and Protopolybia. Interspecific variation in mature colony size was not a good predictor of the type of defensive response, as species with colony sizes smaller and larger than P. emaciata performed nest exiting defensive behavior. An attack response often coincides with, or quickly follows, the nest exit response and involves workers flying from the nest, striking the intruder, and attempting to sting (Hermann and Blum, 1981). When paper-using Polybia nests are tapped, some of the workers on the nest surface may immediately perform attacks, while large numbers of workers rush out of the nest entrance and stand in alert posture on the envelope (Jeanne, 1981; Jeanne et al., 1992). Further tapping elicits additional attacks. Breathing on the nest surface or into the nest results in immediate attack by several to dozens of workers. We discuss the possible adaptive significance of deviations from the typical nest exit that have been documented in several paper wasp species, including P. emaciata.
Methods
This study was conducted in and near the town of Gamboa, Panama province, Republic of Panama (09°07′N 79°42′W; 50 m elevation). Observations and manipulations were conducted during the wet season from 1 August to 15 October 1990 and from 3 to 5 July 1998. Surveys of eusocial wasp colonies during these periods indicated that P. emaciata was among the most abundant swarm-founding wasps in terms of nest density (S.O'D., unpublished data). Most of the data presented were collected from four colonies of P. emaciata and five colonies of P. occidentalis that had been moved approximately 0.5 km to 1 km from their natural nesting sites to artificial supports to facilitate observation. All subject colonies were moved into a small area (<0.25 ha), with similar amounts of tree cover (one P. occidentalis colony was additionally sheltered by building eaves). Movement of the nests did not appear to disrupt behavior or colony development. All colonies were in place at least 2 weeks before observations on defense behavior were initiated, and all continued to forage and to raise brood (visible through the nest entrance in the lower combs) throughout the observation period. The P. emaciata observation colonies had nests ranging in size from 9 cm long to 22 cm long. The numbers of adult wasps and colony age probably varied positively with nest size, though we did not measure these variables. We also collected data opportunistically from P. emaciata colonies in situ in 1990 and again in 1998. These nests were located in the Gamboa area on thin tree branches from 1.5 m to 5 m above the ground.
We disturbed P. emaciata and P. occidentalis colonies in an effort to evoke defensive behavior, using stimuli that elicited nest exit and attack responses in Costa Rican P. occidentalis (S.O'D. and R.L.J. personal observation; Jeanne et al., 1992). We disturbed the Polybia nests by first tapping on the side of the outer envelope near the middle of the nest for 30 sec while wearing a full bee suit, and we noted the responses of on-nest and in-nest workers. If there was no attack response to tapping, we blew into the nest entrance for 30 sec. These disturbances were repeated for up to 2 min or until some workers attacked. Each Polybia observation colony was disturbed 10 times over the course of the study period in 1990. At least 24 h elapsed between tests at each nest. The in situ nests of P. emaciata were tested once each in a similar fashion. We used a non-parametric Wilcoxon test for species differences in the probability of immediate attack responses using the four P. emaciata and five P. occidentalis observation colonies. Trials were pooled within colonies and each colony was used as a single data point in the analysis.
Results
All observation and in situ Polybia colonies contained brood (larvae and/or pupae were visible in the lower combs) when they were tested. Based on forager traffic and on the numbers of wasps exiting the nests during attacks, we estimated that each colony contained several hundred adults.
Polybia emaciata only rarely (5 occasions out of 40 trials on the observation nests and 2 occasions out of >15 trials on in situ nests) exhibited a typical Polybia attack response within 30 sec when the nest was tapped. Polybia emaciata workers never rushed out of the nest and onto the nest envelope. In all trials where the wasps did not attack in response to the initial mechanical disturbance (within the first 30 sec), some of the workers on the nest surface flew off the nest and departed; the remainder rapidly entered the nest. After the initial disturbance, the nest surface and the lowest layer of comb (partially visible through the nest entrance, Fig. 1) were apparently deserted by the adult wasps. Breathing into the nest entrance did not elicit rapid (within 10 sec) attack responses. After continued disturbance within a trial (breath and tapping), P. emaciata workers often attacked violently. In approximately 75% of trials, several dozen individuals rapidly exited the nest, took flight, and struck and attempted to sting the observer's bee veil. The P. emaciata nests were not abandoned following these attacks, and workers later returned to the nests.
Figure 1.

Photograph of a mud nest of Polybia emaciata collected July 1998 in Gamboa, Panama. Note the single entrance hole (indicated by the arrow).
In contrast, all 50 nest-tapping trials with P. occidentalis resulted in immediate (within 1 sec) exit-nest and attack responses. The species difference in the per-colony probability of nest exit and attack within 30 sec of disturbance was significant (Fig. 2; Wilcoxon 2-tailed test, Z=−2.60, p<0.01). Breathing in or on P. occidentalis nests always elicited immediate attack responses, therefore, habituation to tapping and breathing stimuli was not evident over the course of the study in P. occidentalis.
Figure 2.
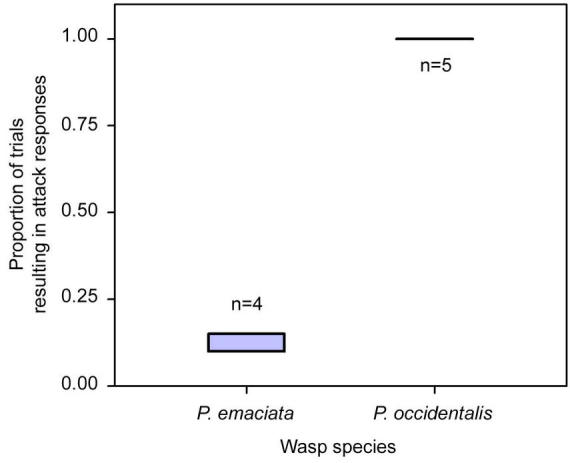
Box plots showing the range of probabilities of response to nest disturbance with attack within 30 sec by Polybia emaciata and P. occidentalis colonies. Samples sizes (numbers of colonies) are given near each box. Each colony was tested 10 times.
The four P. emaciata observation nests suffered no damage while exposed to nearly daily heavy rains for 3 weeks; four of the P. occidentalis observation nests in the same location were visibly damaged by rain during the same period. Even during heavy downpours, the mud nests repelled water much longer than nearby P. occidentalis nests. During one rain shower, the mud nests did not appear waterlogged until 15 min of rainfall had elapsed, while P. occidentalis nests stopped shedding water and became waterlogged after < 5 min. One P. emaciata nest fell from its natural location when its supporting vine snapped. The nest fell 2 m to the ground, with only minor damage to its envelope, and the nest was still occupied by adult wasps when it was collected within 12 h of falling.
Discussion
Rather than exiting and attacking within a few sec of vibrational disturbance, P. emaciata workers usually either departed or fled into the nest interior. Unlike many other paper wasps (S.O'D. and R.L.J., personal observation), P. emaciata failed to respond aggressively to human breath, even when we blew directly into the nest entrance. Polybia emaciata behavioral responses to nest disturbance were unusual among their congeners, and among paper-nesting Epiponini with similar colony sizes (S.O'D and R.L.J. personal observation; Chadab, 1979). However, a few species of paper wasps in other genera exhibit similar responses to human disturbance, which Chadab (1979) labeled ‘retreat behavior’.
Retreat behavior appears to be widespread in P. emaciata. Richards (1978) collected a P. emaciata colony in Brazil, and the wasps did not exit the nest until 1.5 h after it was collected. Residents of Costa Rica from several rural communities reported collecting and moving active P. emaciata colonies without being stung, by placing a finger over the nest entrance and plucking the nest from its attachment point. Chadab (1979) noted that one colony of P. emaciata in Ecuador had “durable” nest paper and exhibited defensive retreat responses like those that we recorded; two other P. emaciata colonies had “fragile” nests and performed exit-nest and attack behavior when jarred. We did not observe qualitative variation in P. emaciata nest durability in Panama (>50 colonies observed; not all were tested for defensive responses).
Predation on swarm-founding wasp nests by bats (Jeanne, 1970a), birds (Skutch, 1971; Windsor, 1976), and primates (Vecht, 1967) has been documented. Vertebrate predation can be a major source of epiponine colony mortality. We hypothesize that the unusual defensive response of P. emaciata is a behavioral adaptation to its use of mud in nest construction, which may make the nest more resistant than paper to entry by vertebrate predators. It appears that the wasps initially rely on the nest itself, rather than on exit and attack behavior, to thwart vertebrate predators. We suggest that this represents a special type of architectural defense (Hermann and Blum, 1981), where the defensive behavior of a species has been modified to exploit the properties of its nest material. However, P. emaciata nests are not impregnable. Skutch (1959) observed a red throated caracara (Daptrius americanus) removing combs from a P. emaciata nest and feeding on the brood in Costa Rica. Our subject nests often responded with attack behavior after extended disturbances, suggesting that attack can be effective for this species. When attack did come it was sudden and massive, suggesting that it was coordinated by an alarm pheromone, as has been demonstrated for P. occidentalis (Jeanne, 1981). Although army ants are among swarm-founding wasps' most frequent predators in wet tropical habitats, preliminary tests on the reaction to Eciton army ants did not suggest that P. emaciata's mud nest is effective in resisting these predators (Chadab, 1979; S.O'D. personal observation).
The use of mud as a nesting material may have evolved in response to predation pressure, particularly by vertebrate enemies. Similar advantages against hornet (Vespa) predation may have favored the evolution of mud construction in hover wasps of the genus Liostenogaster (Hansell 1984; Turillazzi 1991). However, mud construction appears to provide other benefits. Our observations suggest that P. emaciata mud nests were more water repellent, and more resistant to mechanical damage, than similarly sized paper nests. Schremmer (1984) analyzed the water repellent properties of P. emaciata nest material. Even if increases in general durability were the original selective advantage driving the evolution of mud construction, the workers' defensive behavior has apparently been secondarily modified.
There is variability in average colony size and in the details of nest architecture among species within the mud-nesting Polybia subgenus Pedothoeca, and in the independently evolved mud-nester P. furnaria (Richards, 1978; Carpenter et al., 2000). Of special value to our understanding of the evolutionary significance of the mud-nesting habit will be field studies on the defensive behaviors of the other mud-nesting Polybia species. In particular, such studies could establish whether retreat behavior in response to vertebrate disturbance always accompanies the use of mud in nest building. Useful comparisons of defensive behavior could also be made to epiponine paper-nesters with relatively strong nest paper, such as Epipona and Chartergus.
Retreat behavior in response to vertebrate disturbance is not restricted to mud-nesting species of Epiponini. Similar responses to human approach or nest vibration have been observed in paper- and secretion-nesting species (Protopolybia emortualis, Protopolybia exigua, and Protopolybia (formerly Pseudochartergus) fuscatus) and in cavity-nesting Agelaia cajennensis (Jeanne, 1970b; Chadab, 1979; Carpenter et al., 2001). Paper-nesting Polybia jurenei colonies retreated when their nest tree was rubbed, but exited and attacked when the substrate was jarred (S. O'D. personal observation; Rettenmeyer personal communication cited in Chadab, 1979). These patterns suggest that retreat behavior has evolved convergently in several lineages of Epiponini, perhaps in response to particular types of predators.
Acknowledgments
Two anonymous reviewers made helpful comments that improved the paper. We thank Annette Aiello, Frank Joyce, and Donald Windsor for helpful discussions of tropical social wasp behavior and ecology. This work was supported by NSF grants BNS-8517519 to R.L.J. and IBN-9904885 to S.O'D., and by a Smithsonian Tropical Research Institute Short-Term Fellowship to S.O'D.
References
- Carpenter JM, Corbara B, Dejean A. Protopolybia duckei (Du Buysson), a valid species, not a synonym of Protopolybia emortualis (De Saussure) Journal of the New York Entomological Society. 2001;109:179–181. [Google Scholar]
- Carpenter JM, Kojima JI, Wenzel JW. Polybia, paraphyly, and polistine phylogeny. American Museum Novitiates. 2000;3298:1–24. [Google Scholar]
- Chadab R. 1979 Army ant predation on social wasps. Doctoral Dissertation, University of Connecticut, Storrs, CT. [Google Scholar]
- Cole MR, Hansell MH, Seath CJ. A quantitative study of the physical properties of nest paper in three species of Vespine wasps (Hymenoptera, Vespidae) Insectes Sociaux. 2001;48:33–39. [Google Scholar]
- Cooper M. Two new species of Polybia (Hym., Vespidae) with notes on other members of the genus. Entomologists' Monthly Magazine. 1993;129:31–38. [Google Scholar]
- Hansell MH. 1984 Animal Architecture and Building Behaviour. London: Longman. [Google Scholar]
- Hermann HR, Blum MS. 1981 Defensive mechanisms in the social Hymenoptera. In: Hermann HR, editor. Social Insects, vol. II, pp. 78–198.New York: Academic Press. [Google Scholar]
- Jeanne RL. Note on a bat (Phylloderma stenops) preying upon the brood of a social wasp. Journal of Mammalogy. 1970a;51:624–625. [Google Scholar]
- Jeanne RL. Descriptions of the nests of Pseudochartergus fuscatus and Stelopolybia testacea, with a note on a parasite of S. testacea (Hymenoptera, Vespidae) Psyche. 1970b;77:54–69. [Google Scholar]
- Jeanne RL. The adaptiveness of social wasp nest architecture. Quarterly Review of Biology. 1975;50:267–287. [Google Scholar]
- Jeanne RL. Alarm recruitment, attack behavior, and the role of alarm pheromone in Polybia occidentalis (Hymenoptera: Vespidae) Behavioral Ecology and Sociobiology. 1981;9:143–148. [Google Scholar]
- Jeanne RL. 1991 The swarm-founding Polistinae. In: Ross KG, Matthews RW, editors. The Social Biology of Wasps. 191–231.Ithaca: Cornell University Press. [Google Scholar]
- Jeanne RL, Williams NM, Yandell BS. Age polyethism and defense in a tropical social wasp (Hymenoptera: Vespidae) Journal of Insect Behavior. 1992;5:211–227. [Google Scholar]
- London KB, Jeanne RL. Envelopes protect social wasps' nests from phorid infestation (Hymenoptera : Vespidae, Diptera : Phoridae) Journal of the Kansas Entomological Society. 1998;71:175–182. [Google Scholar]
- O'Donnell S, Hunt JH, Jeanne RL. Gaster-flagging during colony defense in Neotropical swarm-founding wasps (Hymenoptera: Vespidae, Epiponini) Journal of the Kansas Entomological Society. 1997;70:175–180. [Google Scholar]
- Rau P. 1933 The Jungle Bees and Wasps of Barro Colorado Island. Kirkwood, MO: Phil Rau. [Google Scholar]
- Richards OW. 1978 The Social Wasps of the Americas. London: British Museum (Natural History). [Google Scholar]
- Schremmer F. 1984 Das Lehmnest der sozialen Faltenwespe Polybia emaciata und Untersuchungen über seine Regenfestigkeit (Hymenoptera, Vespidae, Polybiinae). Sitzungsberichte. Abt. 1, Biologie, Mineralogie, Erdkunde und verwandte Wissenschaften / Österreichische Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse. 193:7–22. [Google Scholar]
- Skutch AF. The scourge of the wasps. Animal Kingdom. 1959;62:8–13. [Google Scholar]
- Skutch AF. 1971 A Naturalist in Costa Rica. Gainseville: Univ. of Florida Press. [Google Scholar]
- Smith A, O'Donnell S, Jeanne RL. Correlated evolution of colony defense and social structure: a comparative analysis in eusocial wasps (Hymenoptera: Vespidae) Evolutionary Ecology Research. 2001;3:331–344. [Google Scholar]
- Strassmann JE, Gastreich KR, Queller DC, Hughes R. Demographic and genetic evidence for cyclical changes in queen number in a Neotropical wasp, Polybia emaciata. American Naturalist. 1992;140:363–372. doi: 10.1086/285417. [DOI] [PubMed] [Google Scholar]
- Turillazzi S. 1991 The Stenogastrinae. In: Ross KG, Matthews RW, editors. The Social Biology of Wasps. 74–98.Ithaca: Cornell University Press. [Google Scholar]
- Vecht J van der. Bouwproblemen van sociale wespen. Verslagen van de gewone vergaderingen der Wis- en Natuurkundige Afdeeling. 1967;76:59–68. [Google Scholar]
- Wenzel JW. 1991 Evolution of nest architecture. In: Ross KG, Matthews RW, editors. The Social Biology of Wasps. 480–519.Ithaca: Cornell University Press. [Google Scholar]
- Windsor DM. Birds as predators on the brood of Polybia wasps (Hymenoptera: Vespidae: Polistinae) in a Costa Rican deciduous forest. Biotropica. 1976;8:111–116. [Google Scholar]