

Abstract
In addition to being a part of the metabolic fatty acid fuel cycle, butyrate is also capable of inducing growth arrest in a variety of normal cell types and senescence-like phenotypes in gynecological cancer cells, inhibiting DNA synthesis and cell growth in colonic tumor cell lines, suppressing hTERT mRNA expression and telomerase activity in human prostate cancer cells, and inducing stem cell differentiation and apoptosis by DNA fragmentation. It regulates gene expression by inhibiting histone deacetylases (HDACs), enhances memory recovery and formation in mice, stimulates neurogenesis in the ischemic brain, promotes osteoblast formation, selectively blocks cell replication in transformed cells (compared to healthy cells), and can prevent and treat diet-induced obesity and insulin resistance in mouse models of obesity, as well as stimulate fetal hemoglobin expression in individuals with hematologic diseases such as the thalassemias and sickle-cell disease, in addition to a multitude of other biochemical effects in vivo. However, efforts to exploit the potential of butyrate in the clinical treatment of cancer and other medical disorders are thwarted by its poor pharmacological properties (short half-life and first-pass hepatic clearance) and the multigram doses needed to achieve therapeutic concentrations in vivo. Herein, we review some of the methods used to overcome these difficulties with an emphasis on HDAC inhibition.
Key words: acylcarnitine, butyrate, butyrylcarnitine, carnitine, histone deacetylase
Introduction
In a seminal article published in 1990, a Japanese group1 reported that (R)-trichostatin A (a fungal antibiotic, Fig. 1) is a potent and specific inhibitor of mammalian histone deacetylase (HDAC) activity in vivo, and that this inhibition strongly induced tumor cell apoptosis, a process that can be mediated epigenetically by regulating the histone function. Since that time, research into the roles HDAC enzymes play in cancer development, progression, and survival has led to the identification of other histone deacetylase inhibitors (HDACi),2–5 and several of these have been evaluated in clinical trials, primarily for the treatment of hematologic malignancies.6–8 HDACi are also being investigated for synergistic cytotoxicity in combination therapies with conventional chemotherapy and targeted agents against solid tumors.9–12 One of the combination strategies fuses the cytotoxic platinum DNA-binding potential of platinum agents and HDAC inhibitory activity into a single molecular entity.13 In 2006, vorinostat (SAHA, Zolinza®, Fig. 1) became the first HDAC inhibitor to be clinically approved by the Food and Drug Administration14 for use in the treatment of cutaneous T-cell lymphoma (a rare form of non-Hodgkin's lymphoma that localizes to the skin). In clinical trials, vorinostat has failed as a monotherapy in the treatment of metastatic breast cancer (and other solid tumors). However, in combination with paclitaxel and bevacizumab as a first-line therapy, the results are more promising.15
FIG. 1.
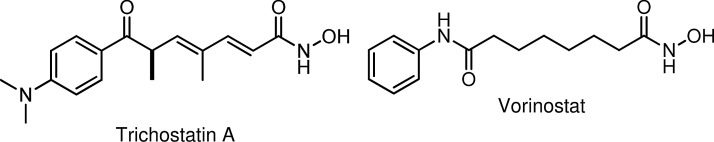
Hydroxamic acid HDACi: trichostatin A and vorinostat. HDACi, histone deacetylase inhibitors.
Mammalian HDAC Isoforms
The mammalian HDAC family of enzymes consists of at least 18 isoforms that are grouped into four principal class types according to their expression patterns and sequence homology relative to the yeast HDAC proteins.16 Class I (HDACs 1, 2, 3, and 8) proteins are found predominantly in the nucleus; Class IIa (HDACs 4, 5, 7, and 9) and Class IIb (HDACs 6 and 10) shuttle between the nucleus and the cytoplasm; Class III enzymes, known as the sirtuins (Sirt1–7), are a silent information regulator (Sir)2 family of proteins17,18 that are NAD+-dependent deacetylases inhibited by nicotinamide;18–20 and Class IV (which contains only HDAC 11) has properties of both Class I and Class II HDACs, and like the Class I and Class IIa/b isoforms, uses a lysine residue to bind to Zn2+ in the active state.3,18,21
In addition to the histone proteins, some nonhistone proteins are also targeted by histone acetyltransferases (referred to as HATs22–24 in keeping with the historical histone relationship) and deacetylases (referred to as HDACs), and as a consequence, acetylation and deacetylation processes figure prominently in mitigating cellular function and activity as well as transcriptional and chromatin-related processes.18,25 Choudhary et al.26 undertook a mass-spectral-based study designed to analyze sites of lysine-specific HDACs (KDACs) and concluded that protein modification by acetylation in vivo could be as common as an event as phosphorylation27–29—a hypothesis predicted a decade earlier by Kouzarides.30 Acetyl is the primary acyl group exchanged by the HATs and HDACs, although propionyl and butyryl may also be substrates.20,31–33 Typically, histone acetylation is associated with the activation of gene transcription, whereas deacetylation is associated with transcriptional repression.34 Similarly, methylation and demethylation are alternative pathways cells utilize to effect epigenetic changes.23,35,36 Strahl and Allis37 suggest that combinatorial sequences of epigenetic changes represent a histone language sometimes referred to as a histone code that proteins could read, write, erase, and modify.38 Importantly, since the energy needed to support epigenetic transformations in a cell is provided by its mitochondria, nuclear and mitochondrial genomic interactions in the cell are coordinated.18
DNA Accessibility
In mammalian cells, the genome is tightly packed into chromatin units called nucleosomes, which consist of ∼147-base-pair segments of DNA wrapped around a core of eight histones (two each of H2A, H2B, H3, and H4).39,40 Electrostatic forces between positive-charged lysine residues on the histone proteins and negative-charged phosphates on the DNA backbone allow the nucleosome to adopt a highly condensed three-dimensional structure that limits access to the DNA segment by transcription factors and other DNA-seeking chemicals. For example, for a segment of DNA to be read by the cell's transcriptional machinery, the DNA must first be made accessible. One means by which this is achieved in the cell is through acetylation by HAT36 of the lysine residues on the histone tails protruding from the nucleosome cores.24,40 This neutralizes the positive charges on the histone tails (relieving electrostatic forces that keep the histone–DNA pair bound closely together) and exposes the DNA.39 Once transcription has been completed, HDAC enzymes remove the N-acetyl groups from the acetylated lysine residues, which restores positive charge to the histone and draws the DNA back into its protected, less-accessible tertiary structure.5
Consequently, inhibiting HDACs can cause gene activation to be initiated or prolonged—an effect that has a therapeutic application across a wide-spectrum of disease phenotypes, as well as in the ex vivo production of stem cells by induced pluripotency (reprogramming),41,42 for use in the emerging field of regenerative therapy.43,44 As a result, much effort is directed toward identifying druggable HDACi with therapeutic potential, particularly of the small-molecule type.7,45–51
n-Butyric Acid
n-Butyric acid, a short-chain naturally occurring fatty acid, is produced in transient amounts during the natural synthesis and breakdown of longer-chain fatty acids in vivo. A significant source in the diet comes from dairy products (such as Greek feta cheese) containing lamb rennet.52,53 However, n-butyric acid (butyrate) is also endogenously made in the human body by anaerobic bacterial fermentation of carbohydrates (derived from dietary fiber) in the colon,54–56 but in addition to being a part of the metabolic fatty acid fuel cycle,57,58 butyrate is also capable of inducing growth arrest in a variety of normal cell types and senescence-like phenotypes in gynecological cancer cells,59,60 inhibiting DNA synthesis and cell growth in colonic tumor cell lines,61–64 suppressing hTERT mRNA expression and telomerase activity in human prostate cancer cells,65 and inducing stem cell differentiation66–71 and apoptosis by DNA fragmentation.72 It regulates gene expression by inhibiting HDACs,73,74 enhances memory recovery and formation in mice,75 stimulates neurogenesis in the ischemic brain,70,76,77 promotes osteoblast formation,78 selectively blocks cell replication in transformed cells (compared to healthy cells),79–81 and can prevent and treat diet-induced obesity and insulin resistance in mouse models of obesity,82 as well as stimulate fetal hemoglobin expression in individuals with hematologic diseases such as the thalassemias and sickle-cell disease,83–85 in addition to a multitude of other biochemical effects in vivo.86–88
However, efforts to exploit the potential of butyrate in the clinical treatment of cancer and other medical disorders are thwarted by its poor pharmacological properties (short half-life and first-pass hepatic clearance) and the multigram doses needed to achieve therapeutic concentrations in vivo.39,79,89–95 Prodrugs of butyric acid96,97 such as Pivanex98–100 and tributyrin58,94,101 (Fig. 2) help mitigate the impediments, but have not been viable as therapeutic agents. Nonetheless, the arginine salt of butyrate has been utilized successfully in clinical studies for the therapy of sickle-cell disease,102,103 thalassemia,104 Epstein-Barr Virus-related malignancies,6 and chronic, non-healing wounds,105 clearly demonstrating the proof of principle for butyrate-based epigenetic therapeutic approaches to these diseases.
FIG. 2.
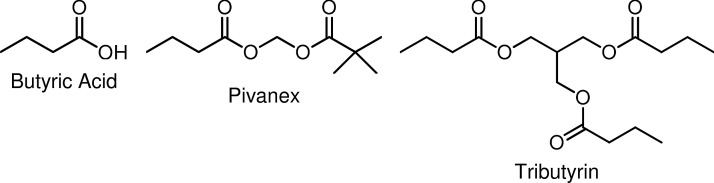
Butyrate HDACi: butyric acid, Pivanex, and tributyrin.
Short-Chain Fatty Acids
Although not as potent as butyric acid, other short-chain fatty acids (SCFAs) like valproic acid and 4-phenylbutyrate (Fig. 3) also have HDAC inhibitory activity.106,107 However, a structure–activity relationship study carried out by Gilbert et al.108 found that nonbranching SCFAs having three to five carbon atoms in length (Fig. 4) are the best inhibitors of HDACs. According to Gilbert et al.108 branching decreases the inhibition of HDACs relative to that observed with n-propionate, n-butyrate, or n-pentanoate. Among the branched-chain acids, valproate exhibited the greatest activity with 2,2-dimethylbutyric acid109,110 and 2-ethylbutyric acid exhibiting only half the activity of valproate. The observations are consistent with the Lu et al.111,112 model of the active site of HDACs being a narrow tube-like pocket, spanning a length equivalent to a straight chain of four to six carbon atoms, with a Zn2+-chelating moiety positioned near the bottom to help facilitate the deacetylation catalysis.113–116 Interestingly, pyruvate, the three-carbon chain end-product of glycolysis, is an endogenous HDAC inhibitor, whereas lactate is not.117 Thus, the tumor-associated cytoplasmic diversion of pyruvate into lactate (the Warburg effect) may serve not only to induce a state of apoptosis resistance by suppressing mitochondrial activity in the tumor cell118 but also as a means for the tumor cell to avoid HDAC inhibition, a potential alternate and suicidal pathway.
FIG. 3.
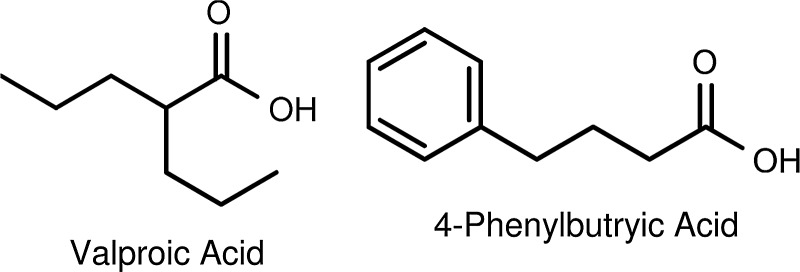
Short-chain fatty acid HDACi: valproic acid and phenylbutyric acid.
FIG. 4.

Variable short-chain fatty acid HDACi.
Cellular Uptake by Transport Proteins
While prodrugs can facilitate cell penetration and help prolong bioavailability, their metabolic cleavage does not necessarily result in therapeutic concentrations of the drug being delivered where needed. Cellular uptake of fatty acids, butyric acid included, is achieved with the aid of specific transporters119,120 and cell surface receptors.120,121 Likewise, cellular entry of carbohydrates is also tightly regulated by carbohydrate-specific transporters and cell surface receptors.120,122,123 The most sensitive clinical test for detecting occult metastases is the enhanced uptake of 2-deoxy-2-[18F]fluoro-d-glucose in vivo by cancer cells compared to normal cells as determined by positron-emission tomography124—implying that glucose (and other natural carbohydrate) scaffolds may be used to carry (cytotoxic) substrates selectively into cancer cells.125–127 This construct was applied to butyrate and other SCFAs,33,79,128–131 for example, Bu4ManNAc and 3,4,6-O-Bu3GlcNAc (Fig. 5).125,130 However, though families of transporters exist for the common dietary sugars such as glucose, they are refractory to modified derivatives (substituted carbohydrates) or analogs, and without assisted cell membrane crossing, cellular uptake of non-natural sugars is limited.122
FIG. 5.
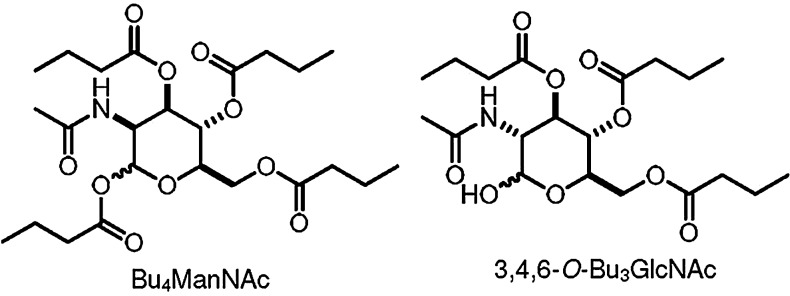
Anticancer sugar-based HDACi butyrates.
Butyryl-l-Carnitine Esters
An alternative delivery method we118,132,133 and others134–138 have found to be an effective transporter of modified SCFA substrates in vivo utilizes the cell's natural carnitine–acylcarnitine transport machinery. More than 100 years ago, Knoop139 published a seminal study on the metabolism of omega-phenyl-substituted fatty acids that he fed to dogs. When the dogs were fed odd-chain-substituted fatty acids, hippuric acid (2-benzamidoacetic acid) was found in their urine, and when fed even-chain-substituted fatty acids, phenylaceturic acid [2-(2-phenylacetamido)acetic acid] was the result.140 To account for these findings, he proposed that the metabolism proceeds by the successive removal of two carbon units via the existence of a mitochondrial fatty acid β-oxidation pathway—a type of carbon oxidation having no reported examples in organic chemistry at the time.140
However, to enter the mitochondrial inner matrix, where β-oxidation takes place, fatty acids (presenting as acylcarnitines)140 are passaged through the mitochondrial membrane via the carnitine acyltransferase pathway.141 Long-chain fatty acids are obligated to be processed this way, but SCFAs can also use this pathway too.142 Thus, as Srinivas et al.135 reported, butyryl-l-carnitine can act as a prodrug for delivering butyrate into cells in vivo, and our own studies (article in preparation) found that butyryl-l-carnitines, PMX™ 550B and PMX 550D (Fig. 6), are more potent HDACi than butyrate itself.
FIG. 6.
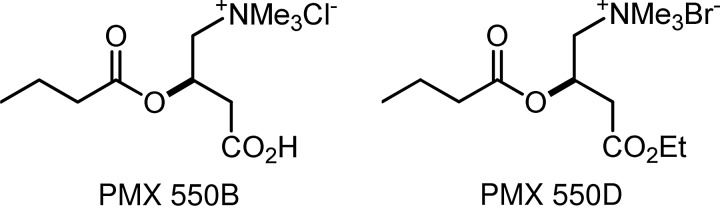
Butyryl-l-carnitines.
Conclusions
Unlike the sugar transporters, which highly discriminate against synthetically modified sugars, the acylcarnitine transporters appear to tolerate a range of acyl-substrate variations118 that can traverse both the plasma118,135,140 and blood–brain barriers.133,134 PMX 550B and PMX 550D are strong butyrate HDACi, and their potential for oral delivery and mitigation of short-half life may lead to more effective epigenetic therapeutics for treating the thalassemias, sickle-cell disease, neurological disorders, and cancer.
Acknowledgments
The authors gratefully acknowledge support from the Foundation for a Cure for Mitochondrial Disease (MitoCure) and from the Karin Grunebaum Cancer Research Foundation, and by the grants: DK-R01-052962, R41 HL-110727, CA-153474, and T32 HL007501 from the National Institutes of Health, and by a grant from the V Foundation.
Authors' Contributions
The article was written through contributions of all authors. All authors have given approval to the final version of the article.
Author Disclosure Statement
K.S. owns shares in PhenoMatriX. No competing financial interests exist.
References
- 1.Yoshida M. Kijima M. Akita M, et al. Potent and specific inhibition of mammalian histone deacetylase both in vivo and in vitro by trichostatin A. J Biol Chem. 1990;265:17174–17179. [PubMed] [Google Scholar]
- 2.Dashwood RH. Ho E. Dietary histone deacetylase inhibitors: From cells to mice to man. Semin Cancer Biol. 2007;17:363–369. doi: 10.1016/j.semcancer.2007.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Balasubramanian S. Verner E. Buggy JJ. Isoform-specific histone deacetylase inhibitors: The next step? Cancer Lett. 2009;280:211–221. doi: 10.1016/j.canlet.2009.02.013. [DOI] [PubMed] [Google Scholar]
- 4.Ma WW. Adjei AA. Novel agents on the horizon for cancer therapy. CA Cancer J Clin. 2009;59:111–137. doi: 10.3322/caac.20003. [DOI] [PubMed] [Google Scholar]
- 5.Newkirk TL. Bowersab AA. Williams RM. Discovery, biological activity, synthesis and potential therapeutic utility of naturally occurring histone deacetylase inhibitors. Nat Prod Rep. 2009;26:1293–1320. doi: 10.1039/b817886k. [DOI] [PubMed] [Google Scholar]
- 6.Perrine SP. Hermine O. Small T, et al. A phase 1/2 trial of arginine butyrate and ganciclovir in patients with Epstein-Barr virus-associated lymphoid malignancies. Blood. 2007;109:2571–2578. doi: 10.1182/blood-2006-01-024703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wang H. Dymock BW. New patented histone deacetylase inhibitors. Expert Opin Ther Patents. 2009;19:1727–1757. doi: 10.1517/13543770903393789. [DOI] [PubMed] [Google Scholar]
- 8.Robey RW. Chakraborty AR. Basseville A, et al. Histone deacetylase inhibitors: Emerging mechanisms of resistance. Mol Pharm. 2011;8:2021–2031. doi: 10.1021/mp200329f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bots M. Johnstone RW. Rational combinations using HDAC inhibitors. Clin Cancer Res. 2009;15:3970–3977. doi: 10.1158/1078-0432.CCR-08-2786. [DOI] [PubMed] [Google Scholar]
- 10.Graham JS. Kaye SB. Brown R. The promises and pitfalls of epigenetic therapies in solid tumours. Eur J Cancer. 2009;45:1129–1136. doi: 10.1016/j.ejca.2009.01.003. [DOI] [PubMed] [Google Scholar]
- 11.Ramalingam SS. Maitland ML. Frankel P, et al. Carboplatin and paclitaxel in combination with either vorinostat or placebo for first-line therapy of advanced non-small-cell lung cancer. J Clin Oncol. 2010;28:56–62. doi: 10.1200/JCO.2009.24.9094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tan J. Cang S. Ma Y, et al. Novel histone deacetylase inhibitors in clinical trials as anti-cancer agents. J Hematol Oncol. 2010;3:5. doi: 10.1186/1756-8722-3-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Griffith D. Morgan MP. Marmion CJ. A novel anti-cancer bifunctional platinum drug candidate with dual DNA binding and histone deacetylase inhibitory activity. Chem Commun. 2009;45:6735–6737. doi: 10.1039/b916715c. [DOI] [PubMed] [Google Scholar]
- 14.Grant S. Easley C. Kirkpatrick P. Vorinostat. Nat Rev Drug Discov. 2007;6:21–22. doi: 10.1038/nrd2227. [DOI] [PubMed] [Google Scholar]
- 15.Fernández Y. Cueva J. Palomo AG, et al. Novel therapeutic approaches to the treatment of metastatic breast cancer. Cancer Treat Rev. 2010;36:33–42. doi: 10.1016/j.ctrv.2009.10.001. [DOI] [PubMed] [Google Scholar]
- 16.Minucci S. Pelicci PG. Histone deacetylase inhibitors and the promise of epigenetic (and more) treatments for cancer. Nat Rev Cancer. 2006;6:38–51. doi: 10.1038/nrc1779. [DOI] [PubMed] [Google Scholar]
- 17.Beher D. Wu J. Cumine S, et al. Resveratrol is not a direct activator of SIRT1 enzyme activity. Chem Biol Drug Des. 2009;74:619–624. doi: 10.1111/j.1747-0285.2009.00901.x. [DOI] [PubMed] [Google Scholar]
- 18.Wallace DC. Fan W. Energetics, epigenetics, mitochondrial genetics. Mitochondrion. 2010;10:12–31. doi: 10.1016/j.mito.2009.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Dai Y. Faller DV. Transcriptional regulation by class III histone deacetylases (sirtuins) Transl Oncogenomics. 2008;3:53–65. doi: 10.4137/tog.s483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Smith BC. Hallows WC. Denu JM. Mechanisms and molecular probes of sirtuins. Chem Biol. 2008;15:1002–1013. doi: 10.1016/j.chembiol.2008.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bäckdahl L. Bushell A. Beck S. Inflammatory signalling as mediator of epigenetic modulation in tissue-specific chronic inflammation. Int J Biochem Cell Biol. 2009;41:176–184. doi: 10.1016/j.biocel.2008.08.023. [DOI] [PubMed] [Google Scholar]
- 22.Manzo F. Tambaro FP. Mai A, et al. Histone acetyltransferase inhibitors and preclinical studies. Expert Opin Ther Patents. 2009;19:761–774. doi: 10.1517/13543770902895727. [DOI] [PubMed] [Google Scholar]
- 23.Munshi A. Shafi G. Aliya N, et al. Histone modifications dictate specific biological readouts. J Genet Genomics. 2009;36:75–88. doi: 10.1016/S1673-8527(08)60094-6. [DOI] [PubMed] [Google Scholar]
- 24.Sleiman SF. Basso M. Mahishi L, et al. Putting the ‘HAT’ back on survival signalling: The promises and challenges of HDAC inhibition in the treatment of neurological conditions. Expert Opin Investig Drugs. 2009;18:573–584. doi: 10.1517/13543780902810345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mellert HS. McMahon SB. Biochemical pathways that regulate acetyltransferase and deacetylase activity in mammalian cells. Trends Biochem Sci. 2009;34:571–578. doi: 10.1016/j.tibs.2009.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Choudhary C. Kumar C. Gnad F, et al. Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science. 2009;325:834–840. doi: 10.1126/science.1175371. [DOI] [PubMed] [Google Scholar]
- 27.Smith KT. Workman JL. Introducing the acetylome. Nat Biotechnol. 2009;27:917–1010. doi: 10.1038/nbt1009-917. [DOI] [PubMed] [Google Scholar]
- 28.Norvell A. McMahon SB. Rise of the rival. Science. 2010;327:964–968. doi: 10.1126/science.1187159. [DOI] [PubMed] [Google Scholar]
- 29.Zhao S. Xu W. Jiang W, et al. Regulation of cellular metabolism by protein lysine acetylation. Science. 2010;327:1000–1004. doi: 10.1126/science.1179689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kouzarides T. Acetylation: A regulatory modification to rival phosphorylation? EMBO J. 2000;19:1176–1179. doi: 10.1093/emboj/19.6.1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chen Y. Sprung R. Tang Y, et al. Lysine propionylation and butyrylation are novel post-translational modifications in histones. Mol Cell Proteomics. 2007;6:812–819. doi: 10.1074/mcp.M700021-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Berndsen CE. Denu JM. Catalysis and substrate selection by histone/protein lysine acetyltransferases. Curr Opin Struct Biol. 2008;18:682–689. doi: 10.1016/j.sbi.2008.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhang K. Chen Y. Zhang Z, et al. Identification and verification of lysine propionylation and butyrylation in yeast core histones using PTMap software. J Proteome Res. 2009;8:900–906. doi: 10.1021/pr8005155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Adachi M. Monteggia LM. Synergistic interactions between histone deacetylase inhibitors and drugs of abuse. Nat Neuropsychopharmacol. 2009;34:2619–2620. doi: 10.1038/npp.2009.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Delage B. Dashwood RH. Dietary manipulation of histone structure and function. Annu Rev Nutr. 2008;28:347–366. doi: 10.1146/annurev.nutr.28.061807.155354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.McGowan PO. Meaney MJ. Szyf M. Diet and the epigenetic (re)programming of phenotypic differences in behavior. Brain Res. 2008;1237:12–24. doi: 10.1016/j.brainres.2008.07.074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Strahl BD. Allis D. The language of covalent histone modifications. Nature. 2000;403:41–45. doi: 10.1038/47412. [DOI] [PubMed] [Google Scholar]
- 38.Perkel JM. Histone code-breakers: The technologies of an epigenetic enigma. Biotechniques. 2010;48:185–191. doi: 10.2144/000113380. [DOI] [PubMed] [Google Scholar]
- 39.Yoo CB. Jones PA. Epigenetic therapy of cancer: Past, present and future. Nat Rev Drug Discov. 2006;5:37–50. doi: 10.1038/nrd1930. [DOI] [PubMed] [Google Scholar]
- 40.Campos EI. Reinberg D. Histones: Annotating chromatin. Annu Rev Genet. 2009;43:559–599. doi: 10.1146/annurev.genet.032608.103928. [DOI] [PubMed] [Google Scholar]
- 41.de Souza M. Primer: Induced pluripotency. Nat Methods. 2010;7:20–21. doi: 10.1038/nmeth.f.293. [DOI] [PubMed] [Google Scholar]
- 42.Eisenstein M. IPSCs: One cell to rule them all? Nat Methods. 2010;7:81–85. [Google Scholar]
- 43.Graf T. Enver T. Forcing cells to change lineages. Nature. 2009;462:587–504. doi: 10.1038/nature08533. [DOI] [PubMed] [Google Scholar]
- 44.Sang Y. Wu M-F. Wagner D. The stem cell—chromatin connection. Semin Cell Dev Biol. 2009;20:1143–1148. doi: 10.1016/j.semcdb.2009.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Miller TA. Witter DJ. Belvedere S. Histone deacetylase inhibitors. J Med Chem. 2003;46:5097–5116. doi: 10.1021/jm0303094. [DOI] [PubMed] [Google Scholar]
- 46.Itoh Y. Suzuki T. Kouketsu A, et al. Design, synthesis, structure-selectivity relationship, and effect on human cancer cells of a novel series of histone deacetylase 6-selective inhibitors. J Med Chem. 2007;50:5425–5438. doi: 10.1021/jm7009217. [DOI] [PubMed] [Google Scholar]
- 47.Mahboobi S. Sellmer A. Höcher H, et al. 2-Aroylindoles and 2-aroylbenzofurans with N-hydroxyacrylamide substructures as a novel series of rationally designed histone deacetylase inhibitors. J Med Chem. 2007;50:4405–4418. doi: 10.1021/jm0703136. [DOI] [PubMed] [Google Scholar]
- 48.Methot JL. Chakravarty PK. Chenard M, et al. Exploration of the internal cavity of histone deacetylase (HDAC) with selective HDAC1/HDAC2 inhibitors (SHI-1:2) Bioorg Med Chem Lett. 2008;18:973–978. doi: 10.1016/j.bmcl.2007.12.031. [DOI] [PubMed] [Google Scholar]
- 49.Kinzel O. Llauger-Bufi L. Pescatore G, et al. Discovery of a potent class I selective ketone histone deacetylase inhibitor with antitumor activity in vivo and optimized pharmacokinetic properties. J Med Chem. 2009;52:3453–3456. doi: 10.1021/jm9004303. [DOI] [PubMed] [Google Scholar]
- 50.Zhang L. Fang H. Xu W. Strategies in developing promising histone deacetylase inhibitors. Med Res Rev. 2010;30:585–602. doi: 10.1002/med.20169. [DOI] [PubMed] [Google Scholar]
- 51.Pontiki E. Hadjipavlou-Litina D. Histone deacetylase inhibitors (HDACIs). Structure–activity relationships: History and new QSAR perspectives. Med Res Rev. 2012;32:1–165. doi: 10.1002/med.20200. [DOI] [PubMed] [Google Scholar]
- 52.Addis M. Piredda G. Pirisi A. The use of lamb rennet paste in traditional sheep milk cheese production. Small Ruminant Res. 2008;79:2–10. [Google Scholar]
- 53.Santillo A. Albenzio M. Quinto M, et al. Probiotic in lamb rennet paste enhances rennet lipolytic activity, and conjugated linoleic acid and linoleic acid content in pecorino cheese. J Dairy Sci. 2009;92:1330–1337. doi: 10.3168/jds.2008-1598. [DOI] [PubMed] [Google Scholar]
- 54.Miller SJ. Cellular and physiological effects of short-chain fatty acids. Mini Rev Med Chem. 2004;4:839–845. doi: 10.2174/1389557043403288. [DOI] [PubMed] [Google Scholar]
- 55.Waldecker M. Kautenburger T. Daumann H, et al. Inhibition of histone-deacetylase activity by short-chain fatty acids and some polyphenol metabolites formed in the colon. J Nutr Biochem. 2008;19:587–593. doi: 10.1016/j.jnutbio.2007.08.002. [DOI] [PubMed] [Google Scholar]
- 56.Scharlau D. Borowicki A. Habermann N, et al. Mechanisms of primary cancer prevention by butyrate and other products formed during gut flora-mediated fermentation of dietary fiber. Mutat Res. 2009;682:39–53. doi: 10.1016/j.mrrev.2009.04.001. [DOI] [PubMed] [Google Scholar]
- 57.Serpa J. Caiado F. Carvalho T, et al. Butyrate-rich colonic microenvironment is a relevant selection factor for metabolically adapted tumor cells. J Biol Chem. 2010;285:39211–39223. doi: 10.1074/jbc.M110.156026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Rajendran P. Williams DE. Ho E, et al. Metabolism as a key to histone deacetylase inhibition. Crit Rev Biochem Mol. 2011;46:181–199. doi: 10.3109/10409238.2011.557713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Terao Y. Nishida J-I. Horiuchi S, et al. Sodium butyrate induces growth arrest and senescence-like phenotypes in gynecological cancer cells. Int J Cancer. 2001;94:257–267. doi: 10.1002/ijc.1448. [DOI] [PubMed] [Google Scholar]
- 60.Rodríguez-Paredes M. Esteller M. Cancer epigenetics reaches mainstream oncology. Nat Med. 2011;17:330–339. doi: 10.1038/nm.2305. [DOI] [PubMed] [Google Scholar]
- 61.Andriamihaja M. Chaumontet C. Tome D, et al. Butyrate metabolism in human colon carcinoma cells: Implications concerning its growth-inhibitory effect. J Cell Physiol. 2009;218:58–65. doi: 10.1002/jcp.21556. [DOI] [PubMed] [Google Scholar]
- 62.Zhang Y. Zhou L. Bao YL, et al. Butyrate induces cell apoptosis through activation of JNK MAP kinase pathway in human colon cancer RKO cells. Chem-Biol Interact. 2010;185:174–181. doi: 10.1016/j.cbi.2010.03.035. [DOI] [PubMed] [Google Scholar]
- 63.Crea F. Nobili S. Paolicchi E, et al. Epigenetics and chemoresistance in colorectal cancer: An opportunity for treatment tailoring and novel therapeutic strategies. Drug Resist Updates. 2011;14:280–296. doi: 10.1016/j.drup.2011.08.001. [DOI] [PubMed] [Google Scholar]
- 64.Rawłuszko AA. Sławek S. Gollogly A, et al. Effect of butyrate on aromatase cytochrome P450 levels in HT29, DLD-1 and LoVo colon cancer cells. Biomed Pharmacother. 2012;66:77–82. doi: 10.1016/j.biopha.2011.12.001. [DOI] [PubMed] [Google Scholar]
- 65.Rahman R. Grundy R. Histone deacetylase inhibition as an anticancer telomerase-targeting strategy. Int J Cancer. 2011;129:2765–2774. doi: 10.1002/ijc.26241. [DOI] [PubMed] [Google Scholar]
- 66.Liang G. Taranova O. Xia K, et al. Butyrate promotes induced pluripotent stem cell generation. J Biol Chem. 2010;285:25516–25521. doi: 10.1074/jbc.M110.142059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mali P. Chou B-K. Yen J, et al. Butyrate greatly enhances derivation of human induced pluripotent stem cells by promoting epigenetic remodeling and the expression of pluripotency-associated genes. Stem Cells. 2010;28:713–720. doi: 10.1002/stem.402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhou M. Li P. Tan L, et al. Differentiation of mouse embryonic stem cells into hepatocytes induced by a combination of cytokines and sodium butyrate. J Cell Biochem. 2010;109:606–614. doi: 10.1002/jcb.22442. [DOI] [PubMed] [Google Scholar]
- 69.Lyssiotis CA. Lairson LL. Boitano AE, et al. Chemical control of stem cell fate and developmental potential. Angew Chem Int Ed. 2011;50:200–242. doi: 10.1002/anie.201004284. [DOI] [PubMed] [Google Scholar]
- 70.Ariff IM. Mitra A. Anirban Basu A. Epigenetic regulation of self-renewal and fate determination in neural stem cells. J Neurosci Res. 2012;90:529–539. doi: 10.1002/jnr.22804. [DOI] [PubMed] [Google Scholar]
- 71.Mali P. Cheng L. Concise review: Human cell engineering: Cellular reprogramming and genome editing. Stem Cells. 2012;30:75–81. doi: 10.1002/stem.735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Roy M-J. Dionne S. Marx G, et al. In vitro studies on the inhibition of colon cancer by butyrate and carnitine. Nutrition. 2009;25:1193–1201. doi: 10.1016/j.nut.2009.04.008. [DOI] [PubMed] [Google Scholar]
- 73.Blouin J-M. Penot G. Collinet M, et al. Butyrate elicits a metabolic switch in human colon cancer cells by targeting the pyruvate dehydrogenase complex. Int J Cancer. 2011;128:2591–2601. doi: 10.1002/ijc.25599. [DOI] [PubMed] [Google Scholar]
- 74.Canani RB. Di Costanzo M. Leone L. The epigenetic effects of butyrate: Potential therapeutic implications for clinical practice. Clin Epigenetics. 2012;4:4. doi: 10.1186/1868-7083-4-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Guan J-S. Haggarty SJ. Giacometti E, et al. HDAC2 negatively regulates memory formation and synaptic plasticity. Nature. 2009;459:55–60. doi: 10.1038/nature07925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Kim HJ. Leeds P. Chuang D-M. The HDAC inhibitor, sodium butyrate, stimulates neurogenesis in the ischemic brain. J Neurochem. 2009;110:1226–1240. doi: 10.1111/j.1471-4159.2009.06212.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Best JD. Carey N. Epigenetic therapies for non-oncology indications. Drug Discov Today. 2010;15:1008–1014. doi: 10.1016/j.drudis.2010.10.006. [DOI] [PubMed] [Google Scholar]
- 78.McGee-Lawrence ME. Westendorf JJ. Histone deacetylases in skeletal development and bone mass maintenance. Gene. 2011;474:1–11. doi: 10.1016/j.gene.2010.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Sampathkumar S-G. Jones MB. Meledeo MA, et al. Targeting glycosylation pathways and the cell cycle: Sugar-dependent activity of butyrate-carbohydrate cancer prodrugs. Chem Biol. 2006;13:1265–1275. doi: 10.1016/j.chembiol.2006.09.016. [DOI] [PubMed] [Google Scholar]
- 80.Pajak B. Orzechowski A. Gajkowska B. Molecular basis of sodium butyrate-dependent proapoptotic activity in cancer cells. Adv Med Sci. 2007;52:83–88. [PubMed] [Google Scholar]
- 81.Lee S. Park J-R. Seo M-S, et al. Histone deacetylase inhibitors decrease proliferation potential and multilineage differentiation capability of human mesenchymal stem cells. Cell Prolif. 2009;42:711–722. doi: 10.1111/j.1365-2184.2009.00633.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Gao Z. Yin J. Zhang J, et al. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes. 2009;58:1509–1517. doi: 10.2337/db08-1637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Pace BS. White GL. Dover GJ, et al. Short-chain fatty acid derivatives induce fetal globin expression and erythropoiesis in vivo. Blood. 2002;100:4640–4648. doi: 10.1182/blood-2002-02-0353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Perrine SP. Fetal globin induction—can it cure β thalassemia? Hematology. 2005;1:38–44. doi: 10.1182/asheducation-2005.1.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Perrine SP. Mankidy R. Boosalis MS, et al. Erythroid Kruppel-like factor (EKLF) is recruited to the γ-globin gene promoter as a co-activator and is required for γ-globin gene induction by short-chain fatty acid derivatives. Eur J Haematol. 2009;82:466–476. doi: 10.1111/j.1600-0609.2009.01234.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Canani RB. Di Costanzo M. Leone L, et al. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J Gastroenterol. 2011;17:1519–1528. doi: 10.3748/wjg.v17.i12.1519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Machado-Vieira R. Ibrahim L. Zarate CA., Jr. Histone deacetylases and mood disorders: Epigenetic programming in gene-environment interactions. CNS Neurosci Ther. 2011;17:699–704. doi: 10.1111/j.1755-5949.2010.00203.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Wang A. Si H. Liu D, et al. Butyrate activates the cAMP-protein kinase A cAMP response element-binding protein signaling pathway in Caco-2 cells. J Nutr. 2012;142:1–6. doi: 10.3945/jn.111.148155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Witt O. Mönkemeyer S. Rönndahl G, et al. Induction of fetal hemoglobin expression by the histone deacetylase inhibitor apicidin. Blood. 2003;101:2001–2007. doi: 10.1182/blood-2002-08-2617. [DOI] [PubMed] [Google Scholar]
- 90.Entin-Meer M. Rephaeli A. Yang X, et al. Butyric acid prodrugs are histone deacetylase inhibitors that show antineoplastic activity and radiosensitizing capacity in the treatment of malignant gliomas. Mol Cancer Ther. 2005;4:1952–1961. doi: 10.1158/1535-7163.MCT-05-0087. [DOI] [PubMed] [Google Scholar]
- 91.Bohacek R. Boosalis MS. McMartin C, et al. Identification of novel small-molecule inducers of fetal hemoglobin using pharmacophore and ‘PSEUDO’ receptor models. Chem Biol Drug Des. 2006;67:318–328. doi: 10.1111/j.1747-0285.2006.00386.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Riester D. Hildmann C. Schwienhorst A. Histone deacetylase inhibitors—turning epigenic mechanisms of gene regulation into tools of therapeutic intervention in malignant and other diseases. Appl Microbiol Biotechnol. 2007;75:499–514. doi: 10.1007/s00253-007-0912-1. [DOI] [PubMed] [Google Scholar]
- 93.Parise RA. Beumera JH. Kangani CO, et al. Liquid chromatography-mass spectrometric assay for quantitation of the short-chain fatty acid, 2,2-dimethylbutyrate (NSC 741804), in rat plasma. J Chromatogr B. 2008;862:168–174. doi: 10.1016/j.jchromb.2007.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Kuroiwa-Trzmielina J. de Conti A. Scolastici C, et al. Chemoprevention of rat hepatocarcinogenesis with histone deacetylase inhibitors: Efficacy of tributyrin, a butyric acid prodrug. Int J Cancer. 2009;124:2520–2527. doi: 10.1002/ijc.24212. [DOI] [PubMed] [Google Scholar]
- 95.Ghosh SK. Perrine SP. Williams RM, et al. Histone deacetylase inhibitors are potent inducers of gene expression in latent EBV and sensitize lymphoma cells to nucleoside antiviral agents. Blood. 2012;119:1008–1017. doi: 10.1182/blood-2011-06-362434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Nudelman A. Gnizi E. Katz Y, et al. Prodrugs of butyric acid. Novel derivatives possessing increased aqueous solubility and potential for treating cancer and blood diseases. Eur J Med. 2001;36:63–74. doi: 10.1016/s0223-5234(00)01199-5. [DOI] [PubMed] [Google Scholar]
- 97.Kessler-Icekson G. Hochhauser E. Sinai T, et al. A histone deacetylase inhibitory prodrug—butyroyloxymethyl diethyl phosphate—protects the heart and cardiomyocytes against ischemia injury. Eur J Pharm Sci. 2012;45:592–599. doi: 10.1016/j.ejps.2011.12.013. [DOI] [PubMed] [Google Scholar]
- 98.Rephaeli A. Entin-Meer M. Angel D, et al. The selectivity and anti-metastatic activity of oral bioavailable butyric acid prodrugs. Invest New Drugs. 2006;24:383–392. doi: 10.1007/s10637-006-6213-1. [DOI] [PubMed] [Google Scholar]
- 99.Chung MC. Ferreira EI. Santos JL, et al. Prodrugs for the treatment of neglected diseases. Molecules. 2008;13:616–677. doi: 10.3390/molecules13030616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Tarasenko N. Nudelman A. Tarasenko I, et al. Histone deacetylase inhibitors: The anticancer, antimetastatic and antiangiogenic activities of AN-7 are superior to those of the clinically tested AN-9 (Pivanex) Clin Exp Metastasis. 2008;25:703–716. doi: 10.1007/s10585-008-9179-x. [DOI] [PubMed] [Google Scholar]
- 101.Mai A. Altucci L. Epi-drugs to fight cancer: From chemistry to cancer treatment, the road ahead. Int J Biochem Cell Biol. 2009;41:199–213. doi: 10.1016/j.biocel.2008.08.020. [DOI] [PubMed] [Google Scholar]
- 102.Perrine SP. Ginder G. Faller DV, et al. A short-term trial of butyrate to stimulate fetal globin gene expression in the β globin gene disorders. N Engl J Med. 1993;328:81–86. doi: 10.1056/NEJM199301143280202. [DOI] [PubMed] [Google Scholar]
- 103.Atweh GF. Dover GJ. Ikuta T, et al. Hematologic responses to pulse butyrate therapy in the beta-hemoglobinopathies. Blood. 1999;93:1790–1797. [PMC free article] [PubMed] [Google Scholar]
- 104.Perrine SP. Castaneda SA. Chui DH, et al. Fetal globin gene inducers: Novel agents and new potential. Ann NY Acad Sci. 2010;1202:158–164. doi: 10.1111/j.1749-6632.2010.05593.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.McMahon L. Tamary H. Askin M, et al. A randomized phase II trial of arginine butyrate with standard local therapy in refractory sickle cell leg ulcers. Br J Haematol. 2010;151:516–524. doi: 10.1111/j.1365-2141.2010.08395.x. [DOI] [PubMed] [Google Scholar]
- 106.Bora-Tatar G. Dayangaç-Erden D. Demir AS, et al. Molecular modifications on carboxylic acid derivatives as potent histone deacetylase inhibitors: Activity and docking studies. Bioorg Med Chem. 2009;17:5219–5228. doi: 10.1016/j.bmc.2009.05.042. [DOI] [PubMed] [Google Scholar]
- 107.Li G-F. Qian T-L. Li G-S, et al. Sodium valproate inhibits MDA-MB-231 breast cancer cell migration by upregulating NM23H1 expression. Genet Mol Res. 2012;11:77–86. doi: 10.4238/2012.January.13.1. [DOI] [PubMed] [Google Scholar]
- 108.Gilbert KM. DeLoose A. Valentine JL, et al. Structure–activity relationship between carboxylic acids and T cell cycle blockade. Life Sci. 2006;78:2159–2165. doi: 10.1016/j.lfs.2005.09.047. [DOI] [PubMed] [Google Scholar]
- 109.Boosalis MS. Bandyopadhyay R. Bresnick EH, et al. Short-chain fatty acid derivatives stimulate cell proliferation and induce STAT-5 activation. Blood. 2001;97:3259–3267. doi: 10.1182/blood.v97.10.3259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Castaneda S. Boosalis MS. Emery D, et al. Enhancement of growth and survival and alterations in Bcl-family proteins in β-thalassemic erythroid progenitors by novel short-chain fatty acid derivatives. Blood Cells Mol Dis. 2005;35:217–226. doi: 10.1016/j.bcmd.2005.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Lu Q. Yang Y-T. Chen C-S, et al. Zn2+-chelating motif-tethered short-chain fatty acids as a novel class of histone deacetylase inhibitors. J Med Chem. 2004;47:467–474. doi: 10.1021/jm0303655. [DOI] [PubMed] [Google Scholar]
- 112.Lu Q. Wang D-S. Chen C-S, et al. Structure-based optimization of phenylbutyrate-derived histone deacetylase inhibitors. J Med Chem. 2005;48:5530–5535. doi: 10.1021/jm0503749. [DOI] [PubMed] [Google Scholar]
- 113.Bieliauskas AV. Weerasinghe SVW. Pflum MKH. Structural requirements of HDAC inhibitors: SAHA analogs functionalized adjacent to the hydroxamic acid. Bioorg Med Chem Lett. 2007;17:2216–2219. doi: 10.1016/j.bmcl.2007.01.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Witter DJ. Harrington P. Wilson KJ, et al. Optimization of biaryl selective HDAC1&2 inhibitors (SHI-1:2) Bioorg Med Chem Lett. 2008;18:726–731. doi: 10.1016/j.bmcl.2007.11.047. [DOI] [PubMed] [Google Scholar]
- 115.Andrianov V. Gailite V. Lola D, et al. Novel amide derivatives as inhibitors of histone deacetylase: Design, synthesis and SAR. Eur J Med Chem. 2009;44:1067–1085. doi: 10.1016/j.ejmech.2008.06.020. [DOI] [PubMed] [Google Scholar]
- 116.Martinet N. Bertrand P. Interpreting clinical assays for histone deacetylase inhibitors. Cancer Manage Res. 2011;3:117–141. doi: 10.2147/CMR.S9661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Thangaraju M. Carswell KN. Prasad PD, et al. Colon cancer cells maintain low levels of pyruvate to avoid cell death caused by inhibition of HDAC1/HDAC3. Biochem J. 2009;417:379–389. doi: 10.1042/BJ20081132. [DOI] [PubMed] [Google Scholar]
- 118.Steliou K. Perrine SP. Faller DV. Lactic acid in cancer and mitochondrial disease. Drug Dev Res. 2009;70:499–511. [Google Scholar]
- 119.Gupta N. Martin PM. Prasad PD, et al. SLC5A8 (SMCT1)-mediated transport of butyrate forms the basis for the tumor suppressive function of the transporter. Life Sci. 2006;78:2419–2425. doi: 10.1016/j.lfs.2005.10.028. [DOI] [PubMed] [Google Scholar]
- 120.Ganapathy V. Thangaraju M. Prasad PD. Nutrient transporters in cancer: Relevance to Warburg hypothesis and beyond. Pharmacol Therapeut. 2009;121:29–40. doi: 10.1016/j.pharmthera.2008.09.005. [DOI] [PubMed] [Google Scholar]
- 121.Brown AJ. Jupe S. Briscoe CP. A family of fatty acid binding receptors. DNA Cell Biol. 2005;24:54–61. doi: 10.1089/dna.2005.24.54. [DOI] [PubMed] [Google Scholar]
- 122.Aich U. Yarema KJ. Non-natural sugar analogues: Chemical probes for metabolic oligosaccharide engineering. In: Fraser-Reid B, editor; Tatsuta K, editor; Thiem J., editor. Glycoscience. Springer-Verlag; Berlin, Germany: 2008. pp. 2133–2190. [Google Scholar]
- 123.Zhang H. Ma Y. Sun X-L. Recent developments in carbohydrate-decorated targeted drug/gene delivery. Med Res Rev. 2010;30:270–289. doi: 10.1002/med.20171. [DOI] [PubMed] [Google Scholar]
- 124.Mertens K. Mees G. Lambert B, et al. In vitro 2-deoxy-2-[18F]fluoro-D-glucose uptake: Practical considerations. Cancer Biother Radiopharm. 2012;27:183–188. doi: 10.1089/cbr.2011.1125. [DOI] [PubMed] [Google Scholar]
- 125.Elmouelhi N. Aich U. Paruchuri VDP, et al. Hexosamine template. A platform for modulating gene expression and for sugar-based drug discovery. J Med Chem. 2009;52:2515–2530. doi: 10.1021/jm801661m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Stallforth P. Lepenies B. Adibekian A, et al. Carbohydrates: A frontier in medicinal chemistry. J Med Chem. 2009;52:5561–5577. doi: 10.1021/jm900819p. [DOI] [PubMed] [Google Scholar]
- 127.Wang Z. Du J. Che P-L, et al. Hexosamine analogs: From metabolic glycoengineering to drug discovery. Curr Opin Chem Biol. 2009;13:565–572. doi: 10.1016/j.cbpa.2009.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Kim EJ. Jones MB. Rhee JK, et al. Establishment of N-acetylmannosamine (ManNAc) analogue-resistant cell lines as improved hosts for sialic acid engineering applications. Biotechnol Prog. 2004;20:1674–1682. doi: 10.1021/bp049841q. [DOI] [PubMed] [Google Scholar]
- 129.Campbell CT. Sampathkumar S-G. Yarema KJ. Metabolic oligosaccharide engineering: Perspectives, applications, and future directions. Mol Biosyst. 2007;3:187–194. doi: 10.1039/b614939c. [DOI] [PubMed] [Google Scholar]
- 130.Campbell CT. Aich U. Weier CA, et al. Targeting pro-invasive oncogenes with short chain fatty acid-hexosamine analogues inhibits the mobility of metastatic MDA-MB-231 breast cancer cells. J Med Chem. 2008;51:8135–8147. doi: 10.1021/jm800873k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Almaraz RT. Aich U. Khanna HS, et al. Metabolic oligosaccharide engineering with N-acyl functionalized ManNAc analogs: Cytotoxicity, metabolic flux, and glycan-display considerations. Biotechnol Bioeng. 2012;109:992–1006. doi: 10.1002/bit.24363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Parameshwaran K. Irwin MH. Steliou K, et al. D-Galactose effectiveness in modeling aging and therapeutic antioxidant treatment in mice. Rejuv Res. 2010;13:729–735. doi: 10.1089/rej.2010.1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Parameshwaran K. Irwin MH. Steliou K, et al. Antioxidant protection of rotenone-induced neuromotor decline, ROS generation and cellular stress in mouse brain. Pharmacol Biochem Behav. 2012;101:487–492. doi: 10.1016/j.pbb.2012.02.011. [DOI] [PubMed] [Google Scholar]
- 134.Nałezcz KA. Miecz D. Berezowski V, et al. Carnitine: Transport and physiological functions in the brain. Mol Aspects Med. 2004;25:551–567. doi: 10.1016/j.mam.2004.06.001. [DOI] [PubMed] [Google Scholar]
- 135.Srinivas SR. Prasad PD. Umapathy NS, et al. Transport of butyryl-L-carnitine, a potential prodrug, via the carnitine transporter OCTN2 and the amino acid transporter ATB0,+ Am J Physiol Gastrointest Liver Physiol. 2007;293:G1046–G1053. doi: 10.1152/ajpgi.00233.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Gong X-w. Li J-p. Wu J-f, et al. Synthesis of carnitine benzyl esters as prodrugs. J Chem Res. 2008;2008:327–330. [Google Scholar]
- 137.Piermatti O. Fringuelli F. Pochini L, et al. Synthesis and characterization of carnitine nitro-derivatives. Bioorg Med Chem. 2008;16:1444–1451. doi: 10.1016/j.bmc.2007.10.053. [DOI] [PubMed] [Google Scholar]
- 138.Rosca MG. Lemieux H. Hoppel CL. Mitochondria in the elderly: Is acetylcarnitine a rejuvenator? Adv Drug Deliver Rev. 2009;61:1332–1342. doi: 10.1016/j.addr.2009.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Knoop F. Der abbau aromatischer fettsäuren im tierkörper (The degradation of aromatic fatty acids in the animal body) Beitr Chem Physiol Pathol. 1904;6:150–162. [Google Scholar]
- 140.Houten SM. Wanders RJA. A general introduction to the biochemistry of mitochondrial fatty acid β-oxidation. J Inherit Metab Dis. 2010;33:469–477. doi: 10.1007/s10545-010-9061-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Bieber LL. Carnitine. Annu Rev Biochem. 1988;57:261–283. doi: 10.1146/annurev.bi.57.070188.001401. [DOI] [PubMed] [Google Scholar]
- 142.Bieber LL. Emaus R. Valkner K, et al. Possible functions of short-chain and medium-chain carnitine acyltransferases. Fed Proc. 1982;41:2858–2862. [PubMed] [Google Scholar]