

Abstract Abstract
A new morphologically cryptic species of phaneropterid bush–cricket from the genus Isophya is described from the Eastern Carpathian Mountains: Isophya dochia sp. n. Sound analysis and morphological details are discussed in the paper comparing the new species with several Isophya species having similar morphology and acoustic behavior.
Keywords: Isophya, bioacoustics, taxonomy, Carpathians
Introduction
One of the largest and most enigmatic phaneropterid genera, with 90 species known and 45 species present in Europe, Isophya Brunner von Wattenwyl inhabits Southern and Eastern Europe, Asia Minor and Caucasus up to Kazakhstan, Iran and Iraq (Eades et al. 2012).
Dissimilar to morphological homogeneity in genus Isophya, the specific structure of male acoustic signals shows clear differences in close related species and is used as the most effective tool for identifying and clarifying taxonomic relations (Heller et al. 2004, Chobanov 2009, Orci et al. 2010a, 2010b, Szövényi et al. 2012). Contrary to this general opinion, a recent study on several related Isophya species from Asia Minor suggests that, in some cases, evolutionary changes in song appear slower than in morphology (Sevgili et al. 2012). In this genus, the male–female pair is usually formed during an acoustic duet; females locate the males by phonotaxis and respond to their songs with simple impulses (Orci et al. 2001, Orci 2007, Orci et al. 2010a, 2010b) or complicated click–series (Iorgu 2012) within a species–specific time span.
The Isophya species are very interesting subjects to study: apart from their remarkable behavior, most of the species have restricted distribution areas and present a large number of endemics. For the time being, 16 Isophya species are known to occur in the Carpathian Mountains (Nagy 2005, Heller 2012). Recently, three new species have been described from the Eastern Carpathian Mountains: Isophya sicula Orci, Szövényi and Nagy, 2010; Isophya ciucasi Iorgu and Iorgu, 2010 and Isophya nagyi Szövényi, Puskás and Orci, 2012.
In the summer of 2005, several bush–crickets were collected from the area called “Poliţa cu Ariniş”, close to the subalpine meadows of Ceahlău Mountains. Studying only the morphological characters, they were identified back then as Isophya camptoxypha (Fieber) and up to the summer of 2010 no acoustic data of this geographically isolated population were available. With the first recorded songs, its status had to be changed in a new taxa, perfectly morphologically cryptic: Isophya dochia sp. n.
Material and methods
Audio recordings were taken with an Edirol R–09HR digital recorder (microphone frequency response 20–40000 Hz, sampling rate of 96000 Hz, 24 bit amplitude resolution). In the field, we used an Edirol CS-15R unidirectional external microphone attached to the digital recorder (frequency response 200–17000 Hz). Temporal and spectral sound analyses were performed with the software Audacity 2.0.2.
Song terminology and abbreviations are adapted from Heller et al. 2004, Orci et al. 2005 and Orci et al. 2010a (Figs 1–4; see Appendix 1: Isophya song abbreviations).
Figures 1–5.

Schematics of studied song (1 Isophya dochia sp. n. 2 Isophya camptoxypha 3 Isophya nagyi 4 Isophya harzi) and morphological (5 Isophya dochia sp. n.) characters.
Morphological traits were examined with a stereomicroscope and the following characters were measured for 20 males and 20 females of the new species: body length (BL), head width (HW), head length (HL), pronotum maximum width (PW), pronotum length (PL), left tegmen maximum width (TW), tegmina length (TL), cercus length (CL) and femur length (FL) (Fig. 5).
Photos were taken with Canon EOS 600D DSLR camera and Canon 100 mm 1:1 and Canon MP–E 65 mm 5:1 macro lenses, using photo stacking method for morphological characters. Movements of tegmina during sound production have been video recorded with the same camera, with the external microphone attached.
The distribution area map was drawn using the altitude layer from Jarvis et al. (2008).
Type specimens are preserved in the collections of “Grigore Antipa” National Museum of Natural History, Bucharest, Romania.
Data resources
The data underpinning the analyses reported in this paper are deposited in the Dryad Data Repository at doi: 10.5061/dryad.256qh
Taxonomy
Isophya dochia sp. n.
urn:lsid:zoobank.org:act:E527F61F-59EC-4761-8E23-495306978BB6
http://species-id.net/wiki/Isophya_dochia
Figs 1 6 –26 32 38 44 45, 51, 57, 63, 69 75
Figures 6–11.

Isophya dochia sp. n.: 6 male habitus 7 copula 8 males’ rivalry 9 female habitus 10 male habitus 11 habitat in Ceahlău Mountains, near Dochia cabin (1740 m).
Figures 12–22.

Isophya dochia sp. n.: 12 dorsal view of male head, pronotum and tegmina 13 lateral view of male pronotum and tegmina 14 male cerci 15 male subgenital plate 16 male stridulatory file (SEM photo) 17 dorsal view of female head, pronotum and tegmina 18 lateral view of female pronotum and tegmina 19 female stridulatory bristles (SEM photo) 20 female cerci 21 female subgenital plate 22 ovipositor. Scale 1 mm.
Figures 23–25.
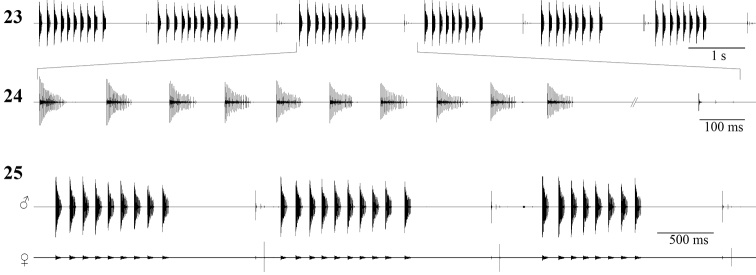
Oscillographic sound analysis in Isophya dochia sp. n., Ceahlău Mountains (24°C): 23 male song, consisting of syllable groups 24 detailed group of syllable 25 male–female mating acoustic duet.
Figures 26–31.
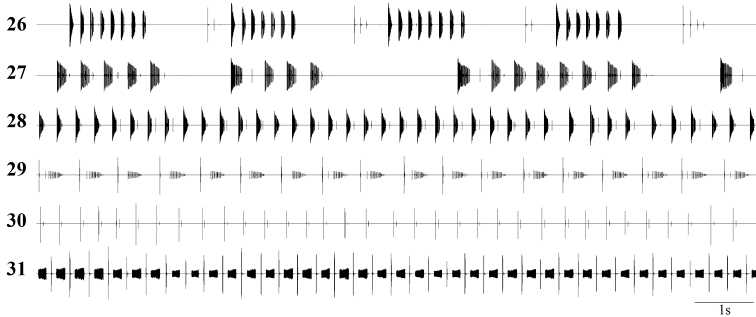
Oscillographic sound analysis: 26 Isophya dochia sp. n., Ceahlău Mountains (24°C) 27 Isophya harzi, Cozia Mountains (25°C) 28 Isophya camptoxypha, Vânători Neamţ(26°C) 29 Isophya ciucasi, Ciucaş Mountains(26°C) 30 Isophya sicula, Harghita Mountains (25°C) 31 Isophya nagyi, Călimani Mountains (25°C).
Figures 32–37.
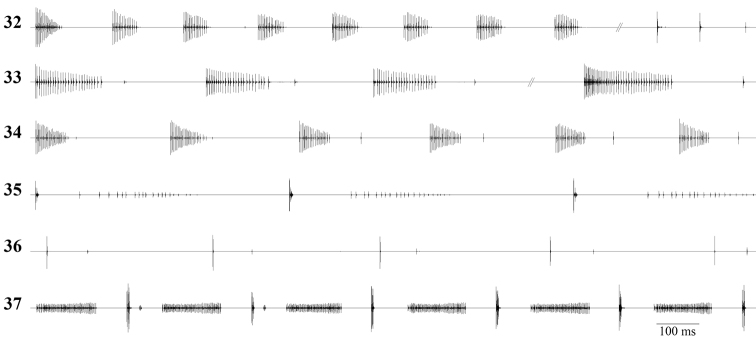
Detailed syllables: 32 Isophya dochia sp. n. 33 Isophya harzi 34 Isophya camptoxypha 35 Isophya ciucasi 36 Isophya sicula 37 Isophya nagyi.
Figures 38–43.

Male–female duet: 38 Isophya dochia sp. n., Ceahlău Mountains (24°C) 39 Isophya harzi, Cozia Mountains (25°C) 40 Isophya camptoxypha, Pleşa(24°C) 41 Isophya ciucasi, Ciucaş Mountains(24°C) 42 Isophya sicula, Harghita Mountains (25°C) 43 Isophya nagyi, Călimani Mountains (25°C).
Figure 44.
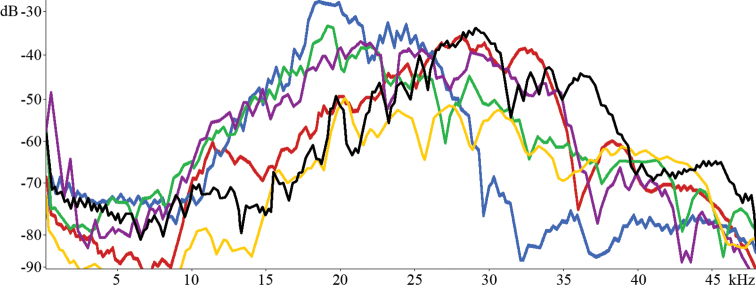
Sound spectrum: black – Isophya dochia sp. n.; blue – Isophya harzi; red – Isophya camptoxypha; green – Isophya ciucasi; yellow – Isophya sicula; purple – Isophya nagyi.
Figures 45–74.

Morphological differences in close–related Isophya species: 45, 51, 57, 63, 69 Isophya dochia sp. n. 46, 52, 58, 64, 70 Isophya harzi, Piatra Craiului Mts. 47, 53, 59, 65, 71 Isophya camptoxypha, Nemira Mts. 48, 54, 60, 66, 72 Isophya ciucasi, Ciucaş Mts. 49, 55, 61, 67, 73 Isophya sicula, Harghita Mts. 50, 56, 62, 68, 74 Isophya nagyi, Călimani Mts. (45–50 male tegmina 51–56 male cerci57–62 ovipositor 63–68 male stridulatory file 69–74 female stridulatory bristles). Scale 1 mm.
Figure 75.

Distribution map of Isophya camptoxypha and allied species in the Romanian Carpathians, based only on acoustic analysis: white – Isophya dochia sp. n. (a); blue – Isophya harzi (b, c); red – Isophya camptoxypha (d, e, f, g, h, i, j, k, l, m, n, o, p, q, r, s, t); green – Isophya ciucasi (u); yellow – Isophya sicula (v, w); purple – Isophya nagyi (x, y, z). (a Ceahlău Mts. b Cozia Mts. c Piatra Craiului Mts. d Rodnei Mts. e Călimani Mts., Pietrosul peak f Mestecăniş g Pietrosul Bistriţei h Rarău i Sadova j Moldoviţa k Pleşa l Vânători Neamţ m Sihla n Nemira Mts. o Penteleu Mts. p Ciuta q Muntele Roşu, Ciucaş Mts. r Bucegi Mts. s Buila–Vânturariţa Mts. t Căpâlna u Ciucaş Mountains v Harghita–Ciceu Mts. w Vânători Neamţ x Călimani Mts., Pietrosul peak y Călimani Mts., Iezerul Călimanului peak z Neagra Şarului).
Type locality.
Romania, Eastern Carpathian Mountains, Ceahlău Mountains.
Type material.
Holotype: male. Original label: “România, Munţii Ceahlău, Poliţa cu Ariniş, 46°57.90'N, 25°56.32'E, alt. 1620 m, 09.08.2005, Leg. Iorgu I. Ş.”
Paratypes: 10 ♂♂ 7 ♀♀, labeled: “România, Munţii Ceahlău, Cabana Dochia, 46°57.94'N, 25°57.02'E, alt. 1740 m, 02.07.2010, Leg. Iorgu I. Ş.”; 6 ♂♂ 5 ♀♀, labeled: “România, Munţii Ceahlău, Piatra Lată, 46°57.73'N, 25°56.93'E, alt. 1720 m, 10.07.2012, Leg. Iorgu I. Ş”, a microSD card containing audio and video records of male song attached; coll. “Grigore Antipa” National Museum of Natural History, Bucharest, Romania.
Audio recordings.
8 ♂♂, 5 July 2010, in laboratory, air temperature 25°C; 17 ♂♂, 10 July 2012, in the field, air temperature 20°C; 5 ♂♂ 5 ♀♀, 12 July 2012, in laboratory, air temperature 24°C (see Appendix 2: Isophya dochia sp n audio and Appendix 3: Isophya dochia sp n video).
Comparative material
(see Appendix: 4 Isophya comparative material).
Description
(Figs 12–22; Table 1). Male. Fastigium verticis slightly tapering frontward, half as wide as scapus, with a dorsal groove (Fig. 12). Head length 1.7 times the pronotum length and head width about 1.3 times the maximum pronotum width. Pronotum saddle shaped from a lateral view, paranota with concave dorsal margins, anterior and ventral borders straight, posterior edge moderately convex. Pronotal disc slightly constricted in the transverse sulcus area, with lateral carinae marginally divergent in mesozona and convex in metazona (Figs 12, 13). Wings as long as or slightly longer than pronotum, with reticulate venation, usually surpass the posterior edge of first abdominal tergite. Cu2 vein length about 3/4 the posterior margin of pronotum; angle between cubital veins about 70°. Speculum large and rectangular. Edge of tegmen at distal end of Cu2 vein forms an obtuse angle of about 130° (Fig. 12). During the quiet wing openings in song production, the white anterior border of tegmen is well visible, but its role has not been established yet (see Appendix 3: Isophya dochia sp n video). Stridulatory file arcuate, 2.2–2.4 mm long, counts 82–89 teeth; distal teeth larger and rarer than proximal ones (Fig. 16). Epiproct about twice as wide as long; cercus slender, narrowing towards tip, slightly curved in its apical fourth, with fine, small hairs; terminal denticle located in middle of cercus apex (Fig. 14). Subgenital plate elongated, narrowed apically, with triangular apical incision (Fig. 15). Hind femur about 4–4.3 times the pronotum length, without ventral spines. Coloration green, densely punctuated with fine, dark green and brown spots. Several males with two dorso–lateral, parallel stripes from pronotum to end of abdomen, red, orange, white, violet or yellow colored. Antennae greenish–brown or reddish–brown, with light brown or green scapus. Compound eyes bicolor: upper part brownish–red and lower part green. A yellowish or white band begins behind the eye and ends at posterolateral angle of wing. Tegmina brown, dark brown or dark red, apically green and costal margin greenish–white or yellowish–white. Cerci brown or reddish–brown, green at base. Ventral side of body yellowish–green. Femora, tibiae and tarsi usually green, brownish or reddish.
Table 1.
Studied morphological characters (see Material and Methods) O – ovipositor.
| HL (mm) | HW (mm) | PL (mm) | PW (mm) | TL (mm) | TW (mm) | BL (mm) | FL (mm) | CL (mm) | O (mm) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| male | Minimum | 1.87 | 3.19 | 3.34 | 3.89 | 3.8 | 3.83 | 21.1 | 14.57 | 2.4 | – |
| Maximum | 1.91 | 3.45 | 4.02 | 4.25 | 4.43 | 4.29 | 24.6 | 16.38 | 2.6 | – | |
| Mean | 1.89 | 3.32 | 3.8 | 4.07 | 4.23 | 4.07 | 22.32 | 15.2 | 2.51 | – | |
| S. D. | 0.02 | 0.1 | 0.24 | 0.14 | 0.23 | 0.16 | 1.12 | 0.61 | 0.08 | – | |
| female | Minimum | 2.79 | 3.64 | 4.02 | 4.14 | 1.68 | – | 22.67 | 14.71 | 1.25 | 8.32 |
| Maximum | 3.07 | 3.98 | 4.58 | 4.66 | 2.39 | – | 26.23 | 16.56 | 1.64 | 8.96 | |
| Mean | 2.9 | 3.8 | 4.31 | 4.37 | 1.92 | – | 24.44 | 15.36 | 1.42 | 8.61 | |
| S. D. | 0.11 | 0.11 | 0.18 | 0.21 | 0.25 | – | 1.31 | 0.66 | 0.15 | 0.21 | |
Female. Fastigium roughly as in male (Fig. 17). Head length 1.7 times the pronotum length and head width about 1.3 times the maximum pronotum width. Pronotum disc marginally enlarged in its posterior part, with straight lateral carinae, paranota as in males (Figs 17, 18). Wing with dense reticulate venation, surpass the posterior margin of first abdominal tergite. Stridulatory bristles located on cubital veins in the inner latero–posterior part of right tegmen (Fig. 19). Cercus short, hairy, conical (Fig. 20). Subgenital plate rounded, narrow, about twice as wide as long (Fig. 21). Ovipositor short, upcurved, 1.9–2.1 times the pronotum length, upper margin with 9–10 denticles and lower margin with 8–9 denticles (Fig. 22). Hind femur 3.5–3.7 times the pronotum length, without ventral spines. Body coloration as in males, wings light brown or green–yellowish, ovipositor green.
Bioacoustics.
Males stridulate at dusk and during the night, rarely during daytime. The tegmino–tegminal stridulation consists of groups of 5–18 syllables (mean±SD: 8.9±3.48, n=30 ♂♂). A group lasts for 1288–4761 ms and successive groups are separated by an interval of 157–326 ms. Groups have a repetition rate of about 10–25 per minute, depending on the number of syllables.
Two types of syllables may be observed in a group: “A” type and “B” type. Both syllable types are produced when the male closes its tegmina. The song pattern may be formulated as “A...AB–A...AB–A...AB–A...AB” and so on, where the unit “A...AB” forms a distinct group of syllables, “...” means a variable number of “A” syllables and “–” means the interval between successive groups of syllables. The “A” type syllable is formed of a compact series of 9–29 impulses (mean±SD: 18.6±6.31), lasting for 30–70 ms (mean±SD: 50.74±12.83). The “B” type syllable is formed of a compact series of 9–15 impulses (mean±SD: 12.36±2.1), lasting for 30–59 ms (mean±SD: 39.3±9.4) and always followed by a series of 1–4 after–clicks at an interval of 621–1655 ms (mean±SD: 1109.1±212.65). The production of the last syllable (“B” type) in each sequence is complex: the male partially closes tegmina, then holds them half–closed for about 621–1655 ms, and finally completely closes the wings with the after–clicks. This late production of after–clicks may function as the trigger element for female acoustic answer. Another possibility is that longer silent gaps may help the male to save up energy or simply listen to its environment, in order to detect possible threats or other singing males (Orci et al. 2010b, Szövényi et al. 2012).
The impulse interval is about 3–6 ms in all syllables and the acoustic signal slowly decreases in amplitude from beginning to end. In a group, the following syllable begins 60–131 ms (mean±SD: 96.28±24.62) later (Figs 23, 24). The carrier wave has the strongest components between 20–40 kHz, with the highest peak at about 29 kHz.
Females find males by phonotaxis and if willing to mate with the singing male (Fig. 7), they produce isolated impulses. In the resulting male–female duet, the female stridulates only after the male after–clicks (ending part of “B” syllable), with a latency of 13–34 ms (mean±SD: 25.66±6.42, n=12 responses from 3 females) (Fig. 25).
Distribution and ecology.
Isophya dochia sp. n. populates mesophytic subalpine meadows at about 1600–1900 m, in Ceahlău Mountain Massif, Eastern Carpathians (Fig. 11). The specimens were collected from leaves of Urtica, Rubus, Veratrum, Rumex, Aconitum, Vaccinium, Hypericum, Stachys, Junniperus etc. Few other bush–crickets and grasshoppers were found occurring simpatrically with the new species: Metrioptera bicolor (Philippi), Metrioptera brachyptera (Linnaeus), Pholidoptera transsylvanica (Fischer), Miramella ebneri (Galvagni), Euthystira brachyptera (Ocskay), Myrmeleotettix maculatus (Thunberg), Chorthippus biguttulus (Linnaeus), Chorthippus parallelus (Zetterstedt) etc. The bush–cricket Isophya dochia sp. n. has the same phenology as other subalpine Isophya species: female lays her eggs isolated in holes bitten in broad leaves of Urtica, Rubus etc. Eggs pass the winter in the litter and larvae hatch in late spring, after the snow melts in the high mountains. Depending on weather, first adults exuviate in late June and live up to August.
Etymology.
A noun in apposition; from the name of Dochia, a Romanian legendary female character based on an earlier deity of land and agriculture from the Dacian pantheon, and that of the eponymous rock in Ceahlău Mountains.
Discussions
(see Appendix 5: Isophya harzi, Appendix 6: Isophya camptoxypha, Appendix 7: Isophya ciucasi, Appendix 8: Isophya sicula, Appendix 9: Isophya nagyi).
Discovery of the bush–cricket Isophya dochia sp. n. is surprising, especially as it is a morphological cryptic species closely related to Isophya camptoxypha. Some other recently described species, Isophya ciucasi, Isophya nagyi and Isophya sicula, show high resemblance in morphology, while Isophya harzi Kis has a similar calling song (Figs 26–37). The song of these six species can be readily distinguished as syllables grouped in short sequences (Isophya dochia sp. n. and Isophya harzi) or arranged in series (Isophya camptoxypha, Isophya ciucasi, Isophya nagyi and Isophya sicula).
Both Isophya dochia sp. n. and Isophya harzi stridulate well defined assemblies of syllables (Figs 26, 27, 32, 33). Two other species have similar song patterns: Isophya posthumoidalis Bazyluk, distributed in Poland, Slovakia and N Romania (Szövényi and Orci 2008) and Isophya beybienkoi Mařan, known so far only from a very small area in SE Slovakia. In all these four species, males produce long sequences in which two types of syllables may be noticed: “A” and “B”, the rhythm of the whole song being a constant repetition that may be formulated as “A...AB–A...AB–A...AB” (in Isophya dochia sp. n. andIsophya harzi), “A...A–B–A...A–B–A...A” (in Isophya posthumoidalis) and “A...A–BA...A–BA...A” (in Isophya beybienkoi). The descriptions of calling songs of Isophya posthumoidalis and Isophya beybienkoi (Orci et al. 2001, Heller et al. 2004, Orci et al. 2010a) suggest that syllable types are very different from Isophya dochia sp. n.: in Isophya posthumoidalis, syllable “A” consists of a compact series of 5–10 impulses, lasting for 12–15 ms, and syllable “B” is a single impulse followed by 1–3 after–clicks, while in Isophya beybienkoi syllable “A” is formed of 5–13 impulses, lasting for 10-32 ms, and syllable “B” consists of a compact series of 2–9 short impulses followed by a longer one and 1–3 after–clicks. Not possessing any personal data on these two species, their songs and morphology were not illustrated in present paper.
In Isophya camptoxypha, the syllable is a short and compact series of impulses, lacking or followed by one (extremely rare 2–7) after–click (Figs 28, 34). Males of Isophya ciucasi stridulate a shorter syllable, but followed by a very high number of after–clicks, usually 10–30 (Figs 29, 35). Isophya sicula produces the shortest known syllables within this genus, consisting of only 1–3 impulses, missing or followed by one (extremely rare 2–5) after–click (Figs 30, 36). Finally, the song of Isophya nagyi is the most interesting, syllables being divided in two or three distinct fragments: the first one is an opening syllable and last two are part of the same closing syllable (Figs 31, 37). During our studies, less than 1% of analyzed syllables contained all three components, males usually producing sounds only when closing the tegmina.
Another interesting feature of the song in some of these species is the similarity of time–windows when after–clicks are produced: usually 50–80 ms after the syllable, but up to 151 ms in Isophya camptoxypha (Heller et al. 2004) and up to 110 ms in Isophya sicula. After–clicks may also follow both types of syllables of Isophya harzi. However, we observed a longer after–click delay (75–202 ms) next to the “B” syllable of the sequence and a shorter delay after the “A” syllables (40–90 ms). The longest after–click delay is found in Isophya dochia sp. n., up to 1655 ms.
A remarkable interspecific variation in this group is the number of teeth on male stridulatory file (Figs 63–68), which may be correlated with the total length and number of impulses in the unit syllable + gap + after–clicks, meaning a total closing stroke of wing. In the species that sing well defined groups of syllables: in Isophya harzi, 98–130 pegs produce a total number of 37–59 impulses lasting for 94–418 ms in the “B” syllable and 23–55 impulses lasting for 75–258 ms in the “A” syllable, while in Isophya dochia sp. n., the 82–89 pegs produce a total number of 9–29 impulses lasting for 30–70 ms in the first syllables from a group (“A” type) and 10–19 impulses lasting for 960–1770 ms in the last syllable (“B” type). In the species that stridulate ungrouped syllables: 65–85 pegs in Isophya ciucasi produce 13–56 impulses lasting for 209–438 ms, 50–80 pegs in Isophya camptoxypha produce 10–39 impulses, total duration of 27–363 ms, 48–60 pegs in Isophya sicula produce a total number of 1–8 impulses lasting for 52–265 ms. In Isophya nagyi, the high number of teeth on the stridulatory file, i.e. 102–109, produce a song of 36–108 impulses lasting for 167–793 ms (Table 2). All these differences in syllable production time are caused by slower or faster species–specific wing movements.
Table 2.
Examined male song characters in close related Isophya species (for abbreviations see Appendix 1: Isophya song abbreviations) n – number of analyzed specimens; t – temperature °C.
| Song type | Species | Song characters | Min. | Max. | Mean | S. D. | n | t |
|---|---|---|---|---|---|---|---|---|
| Grouped syllable sequences | Isophya dochia sp. n. | ASD (ms) | 30 | 70 | 50.74 | 12.83 | 30 | 20–25 |
| NIA | 9 | 29 | 18.6 | 6.31 | ||||
| GS (ms) | 60 | 131 | 96.28 | 24.62 | ||||
| BSD (ms) | 30 | 59 | 39.3 | 9.4 | ||||
| NIB | 9 | 15 | 12.36 | 2.1 | ||||
| DACB (ms) | 621 | 1655 | 1109.1 | 212.65 | ||||
| NACB | 1–4 | |||||||
| BSTD (ms) | 621 | 1770 | 1114.46 | 325.45 | ||||
| SGTD (ms) | 1288 | 4761 | 2633.56 | 792.48 | ||||
| SGD (ms) | 612 | 3028 | 1406.86 | 598.95 | ||||
| GSG (ms) | 157 | 326 | 225.2 | 52.66 | ||||
| Isophya harzi | ASD (ms) | 75 | 168 | 121.64 | 28.36 | 16 | 19–26 | |
| NIA | 23 | 55 | 40.5 | 9.67 | ||||
| DACA (ms) | 40 | 90 | 64 | 16.97 | ||||
| NACA | 0–2 | |||||||
| ASTD (ms) | 75 | 258 | 156.42 | 77.01 | ||||
| GAA (ms) | 122 | 298 | 175.78 | 48.25 | ||||
| GAB (ms) | 367 | 789 | 527.4 | 126.18 | ||||
| BSD (ms) | 94 | 216 | 154.7 | 43.54 | ||||
| NIB | 37 | 59 | 46.5 | 8.38 | ||||
| DACB (ms) | 75 | 202 | 142.3 | 47.19 | ||||
| NACB | 0–2 | |||||||
| BSTD (ms) | 94 | 418 | 223.3 | 114.78 | ||||
| SGTD (ms) | 617 | 6168 | 2254.8 | 1954.04 | ||||
| SGD (ms) | 617 | 6091 | 2126 | 2001.43 | ||||
| GSG (ms) | 191 | 303 | 249.8 | 40.28 | ||||
| Ungrouped syllable sequences | Isophya camptoxypha | SD (ms) | 27 | 90 | 61.76 | 21.37 | 58 | 19–27 |
| NI | 10 | 32 | 19.9 | 7.62 | ||||
| GS (ms) | 110 | 289 | 189 | 50.18 | ||||
| DAC (ms) | 56 | 257 | 112.6 | 70.71 | ||||
| NAC | 0–1 (rarely 2–7) | |||||||
| STD (ms) | 27 | 363 | 77.8 | 97.58 | ||||
| Isophya ciucasi | SD (ms) | 7 | 24 | 15.56 | 5.12 | 34 | 24–28 | |
| NI | 3 | 26 | 15.32 | 6.74 | ||||
| GS (ms) | 134 | 600 | 378.08 | 116.95 | ||||
| DAC (ms) | 52 | 167 | 101.2 | 38.31 | ||||
| NAC | 10–30 | |||||||
| STD (ms) | 209 | 438 | 273.76 | 56.76 | ||||
| Isophya nagyi | SD (ms) | 167 | 793 | 486.28 | 200.49 | 12 | 21–25 | |
| SDC1 (ms) | 9 | 31 | 21 | 8.08 | ||||
| NIC1 | 5 | 18 | 12 | 4.72 | ||||
| GC12 (ms) | 68 | 301 | 187.6 | 101.94 | ||||
| SDC2 (ms) | 127 | 221 | 173.9 | 31.57 | ||||
| NIC2 | 35 | 79 | 57 | 13.5 | ||||
| GC23 (ms) | 39 | 97 | 67.5 | 19.04 | ||||
| SDC3 (ms) | 1 | 35 | 16.4 | 12.33 | ||||
| NIC3 | 1 | 11 | 5.1 | 3.36 | ||||
| GS (ms) | 59 | 235 | 154.26 | 49.59 | ||||
| Isophya sicula | SD (ms) | 1 | 4 | 1.6 | 1.03 | 3 | 25 | |
| NI | 1 | 3 | 1.3 | 0.65 | ||||
| GS (ms) | 190 | 446 | 337.78 | 62.64 | ||||
| DAC (ms) | 51 | 110 | 73.2 | 19.71 | ||||
| NAC | 0–1 (rarely 2–5) | |||||||
| STD (ms) | 52 | 265 | 75.1 | 63.96 |
Female response as male acceptance is formed of isolated impulses, always produced after a particular part of male song, which supposedly acts as trigger. In both Isophya dochia sp. n. and Isophya harzi (n=13 responses from 3 ♀♀), the female replies only after the male’s “B” type syllable. The same behavior was noticed in females from Isophya posthumoidalis and Isophya beybienkoi (Orci et al. 2001, Orci et al. 2010a). Females of Isophya camptoxypha, Isophya ciucasi and Isophya sicula reply right after the male’s syllable main part. In Isophya camptoxypha, female response is produced immediately after the male’s after–click (n=19 responses from 3 ♀♀), while in Isophya ciucasi (n=37 responses from 6 ♀♀) and Isophya sicula (n=10 responses from 1 ♀) its answer is apparently not affected by male after–clicks. Females of Isophya nagyi stridulate only after the last component of the male syllable (n=25 responses from 4 ♀♀) (Figs 38–43).
Spectrographic analysis of sound reveals that in all six species the frequency ranges somewhere within interval 10–40 kHz, the maximum being recorded at about 20–30 kHz (Fig. 44).
In the six related species, males of Isophya harzi and Isophya sicula can be easily separated morphologically from Isophya camptoxypha, Isophya ciucasi, Isophya nagyi and Isophya dochia sp. n. Isophya harzi is more massive, males having shorter wings, marginal angle of tegmina less obtuse (110°), number of stridulatory teeth larger, and females having a longer ovipositor (11–13 mm). Males of Isophya sicula have a narrow left wing, similar with Isophya posthumoidalis and Isophya beybienkoi (Heller et al. 2004). In the other four species, the angle of cubital veins on male wing may be used as a differentiation tool: 80–90° in Isophya ciucasi, 70–80° in Isophya nagyi, 60–70° in Isophya camptoxypha and 70° in Isophya dochia sp. n.The tegmen marginal angle is almost constant in these species (about 120°), but less obtuse in Isophya harzi (110°) (Figs 45–50). In the six species, male cercus morphology has minute variations (Figs 51–56). The ovipositor is relatively similar in length in all species, being longer in Isophya harzi (Figs 57–62) and Isophya beybienkoi (Heller et al. 2004). Female stridulatory area is subject to minor intra– and interspecific variability (Figs 69–74).
Having a look at species distribution in the Romanian Carpathians, Isophya camptoxypha has the widest spread, inhabiting many Carpathian highlands. Isophya harzi is known only from two isolated mountains in the Southern Carpathians and most probably its distribution area is wider in the mountains between Prahova and Olt rivers. At this moment, both Isophya ciucasi and Isophya dochia sp. n. are known to have very restricted distribution areas: the isolated Ciucaş and, respectively, Ceahlău Mountain Massifs. Two species have been recently described from the Transylvanian volcanic mountains, in the Western part of the Eastern Carpathians. Isophya nagyi occurs in Călimani Mountains and Dorna Basin, while Isophya sicula, described from Harghita–Ciceu Mountains, has been recently found in the Moldavian Subcarpathians (Fig. 75). Isophya posthumoidalis was recorded in Romania only in Maramureş Basin (Szövényi and Orci 2008) and Isophya beybienkoi is known only from the area where it was described: Zadiělská planina and Plešivecká planina, Slovenskie Kras, SE Slovakia (Orci et al. 2001, Heller et al. 2004). In order to presume the evolutionary pattern within this group of species, some interesting areas must be taken into account, namely the regions where species live syntopically: Isophya camptoxypha and Isophya nagyi have been found together in W Călimani Mountains, Isophya camptoxypha and Isophya sicula in Moldavian Subcarpathians, Isophya camptoxypha and Isophya ciucasi in Central Ciucaş Mountains.
The Orthoptera species of the Carpathians have been well investigated and morphologically characterized. Yet, Isophya, and particularly Isophya camptoxypha and its allies, remain among the most intriguing and widely distributed bush–crickets in these mountains. Due to Isophya camptoxypha’s high intra– and interpopulational morphological variability, the main tool to separate correctly possible new cryptic species remains the oscillographic analysis of acoustic signals.
Supplementary Material
Acknowledgements
The author is grateful to Klaus–Gerhard Heller for his valuable support with literature, to Elena Iulia Iorgu, Adrian Derscariu, Liviu Moscaliuc and Irinel Popescu, his traveling companions in the Carpathians and to Lucian Fusu for inspiration during the preparation of manuscript. Ceahlău National Park, Rodnei National Park and Călimani National Park Administrations are acknowledged for the excellent work conditions. Advices of anonymous reviewers significantly improved the paper.
Appendix 1
Isophya song abbreviations. (doi: 10.3897/zookeys.254.3892.app1) File format: Microsoft Excel file (xls).
Explanation note: Abbreviations used for calling songs of Isophya males, described in Figures 1-4 and Table 2.
Appendix 2
Isophya dochia sp. n. audio. (doi: 10.3897/zookeys.254.3892.app2) File format: Windows audio file (wav).
Explanation note: Audio recording of calling song of a male Isophya dochia sp. n.
Appendix 3
Isophya dochia sp. n. video. (doi: 10.3897/zookeys.254.3892.app3) File format: Video file (MPEG Layer 4) (mp4).
Explanation note: Video recording of calling song of a male Isophya dochia sp. n.
Appendix 4
Isophya comparative material. (doi: 10.3897/zookeys.254.3892.app4) File format: Microsoft Excel file (xls).
Explanation note: Acoustic analyzed Isophya material in the Romanian Carpathian Mountains.
Appendix 5
Isophya harzi audio. (doi: 10.3897/zookeys.254.3892.app5) File format: Windows audio file (wav).
Explanation note: Audio recording of calling song of a male Isophya harzi.
Appendix 6
Isophya camptoxypha audio. (doi: 10.3897/zookeys.254.3892.app6) File format: Windows audio file (wav).
Explanation note: Audio recording of calling song of a male Isophya camptoxypha.
Appendix 7
Isophya ciucasi audio. (doi: 10.3897/zookeys.254.3892.app7) File format: Windows audio file (wav).
Explanation note: Audio recording of calling song of a male Isophya ciucasi.
Appendix 8
Isophya sicula audio. (doi: 10.3897/zookeys.254.3892.app8) File format: Windows audio file (wav).
Explanation note: audio recording of calling song of a male Isophya sicula.
Appendix 9
Isophya nagyi audio. (doi: 10.3897/zookeys.254.3892.app9) File format: Windows audio file (wav).
Explanation note: Audio recording of calling song of a male Isophya nagyi.
References
- Chobanov DP. (2009) Phylogeny and systematics of the Isophya modesta group (Phaneropteridae). Metaleptea 29 (1): 20–27. http://140.247.119.225/OrthSoc/pdf/met_29_1.pdf [Google Scholar]
- Eades DC, Otte D, Cigliano MM, Braun H. (2012) Orthoptera Species File Online. Version 2.0/4.1. http://orthoptera.speciesfile.org/HomePage.aspx
- Heller K–G. (2012) Fauna Europaea: Orthoptera. Version 2.5. http://www.faunaeur.org/
- Heller K–G, Orci KM, Grein G, Ingrisch S. (2004) The Isophya species of central and western Europe (Orthoptera: Tettigonioidea, Phaneropteridae). Tijdschrift voor Entomologie 147: 237-258. doi: 10.1163/22119434-900000153 [DOI] [Google Scholar]
- Iorgu IŞ. (2012) Bioacoustics of Isophya dobrogensis, a Romanian endemic bush–cricket (Orthoptera: Phaneropteridae). Travaux du Muséum National d’Histoire Naturelle “Grigore Antipa” 55 (1): 51-56. doi: 10.2478/v10191-012-0005-y [DOI] [Google Scholar]
- Jarvis A, Reuter HI, Nelson A, Guevara E. (2008) Hole–filled SRTM for the globe Version 4, available from the CGIAR–CSI SRTM 90m Database. http://srtm.csi.cgiar.org/
- Nagy B. (2005) Orthoptera fauna of the Carpathian Basin – recent status of knowledge and a revised check–list. Entomofauna Carpathica 17: 14-22. [Google Scholar]
- Orci KM. (2007) Female preferences for male song characters in the bush–cricket Isophya camptoxypha (Orthoptera, Tettigonioidea). Journal of Insect Behavior 20 (5): 503-513. doi: 10.1007/s10905-007-9096-x [DOI] [Google Scholar]
- Orci KM, Nagy B, Szövényi G, Rácz I, Varga Z. (2005) A comparative study on the song and morphology of Isophya stysi Čejchan, 1958 and Isophya modestior Brunner von Wattenwyl, 1882 (Orthoptera, Tettigoniidae). Zoologischer Anzeiger 244 (1): 31-42. doi: 10.1016/j.jcz.2004.11.002 http://www.sciencedirect.com/science/article/pii/S004452310500032X [DOI] [Google Scholar]
- Orci KM, Szövényi G, Nagy B. (2001) Description of the song of Isophya beybienkoi (Orthoptera, Tettigonioidea). Biologia (Bratislava) 56: 489-495. [Google Scholar]
- Orci KM, Szövényi G, Nagy B. (2010a) A characterization of the pair forming acoustic signals of Isophya harzi (Orthoptera, Tettigonioidea, Phaneropteridae). Acta Zoologica Academiae Scientiarum Hungaricae 56 (1): 43–53. http://actazool.nhmus.hu/56/1/azh56_1_Orci.pdf [Google Scholar]
- Orci KM, Szövényi G, Nagy B. (2010b) Isophya sicula sp. n. (Orthoptera: Tettigonioidea), a new, morphologically cryptic bush–cricket species from the Eastern Carpathians (Romania) recognized from its peculiar male calling song. Zootaxa 2627: 57–68. http://www.mapress.com/zootaxa/2010/f/zt02627p068.pdf [Google Scholar]
- Sevgili H, Demirsoy A, Çiplak B. (2012) Description and bioacoustics of a new species of the genus Isophya (Orthoptera: Tetigoniidae: Phaneropterinae) from Turkey. Zootaxa 3361: 33–44. http://www.mapress.com/zootaxa/list/2012/3361.html [Google Scholar]
- Szövényi G, Orci KM. (2008) Contributions to the Orthoptera fauna of Maramureş county with the first record of Isophya posthumoidalis Bazyluk 1971 in Romania. Studia Universitatis “Vasile Goldiş”, Seria Ştiinţele Vieţii 19: 235-241. [Google Scholar]
- Szövényi G, Puskás G, Orci KM. (2012) Isophya nagyi, a new phaneropterid bush–cricket (Orthoptera: Tettigonioidea) from the Eastern Carpathians (Caliman Mountains, North Romania). Zootaxa 3521: 67–79. http://www.mapress.com/zootaxa/2012/f/zt03521p079.pdf [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.