

Abstract
DTNBP1 is one of the most established susceptibility genes for schizophrenia, and hippocampal volume reduction is one of the major neuropathological findings in this severe disorder. Consistent with these findings, the encoded protein dysbindin-1 has been shown to be diminished in glutamatergic hippocampal neurons in schizophrenic patients. The aim of this study was to investigate the effects of two single nucleotide polymorphisms of DTNBP1 on grey matter volumes in human subjects using voxel-based morphometry. Seventy-two subjects were included and genotyped with respect to two single nucleotide polymorphisms of DTNBP1 (rs2619522 and rs1018381). All participants underwent structural magnetic resonance imaging (MRI). MRI data were preprocessed and statistically analysed using standard procedures as implemented in SPM5 (Statistical Parametric Mapping), in particular the voxel-based morphometry (VBM) toolbox. We found significant effects of the DTNBP1 SNP rs2619522 bilaterally in the hippocampus as well as in the anterior middle frontal gyrus and the intraparietal cortex. Carriers of the G allele showed significantly higher grey matter volumes in these brain regions than T/T homozygotes. Compatible with previous findings on a role of dysbindin in hippocampal functions as well as in major psychoses, the present study provides first direct in vivo evidence that the DTNBP1 SNP rs2619522 is associated with variation of grey matter volumes bilaterally in the hippocampus.
Keywords: Magnetic resonance imaging, Psychosis, Genetic susceptibility, Genetics, Hippocampus, rs1018381
Introduction
Schizophrenia is a common and severely disabling psychiatric disorder where heritability is of considerable relevance in the aetiology of the disease [9, 10, 43, 53]. One of the most replicated susceptibility genes for schizophrenia is the dysbindin-1 gene (DTNBP1; OMIM *607145) on chromosome 6p22.3. Straub and collaborators first reported an association of DTNBP1 (dystrobrevin-binding protein 1, or dysbindin) with schizophrenia in a family-based association analysis of High-Density Schizophrenia Families in Ireland ([32], Irish Study of High-Density Schizophrenia Families ISHDSF) in 2002. Since then (and even before), numerous investigations have been realized to verify the role of DTNBP1 as a schizophrenia susceptibility gene. While many genetic studies could replicate the association of DTNBP1 with schizophrenia [17, 34, 55, 61, 71, 74, 75, 83, 87], others failed to confirm the finding of Straub and colleagues [50, 51, 67]. A meta-analysis authored by Li & He in 2007 only found a weak association of DTNBP1 with schizophrenia [36].
On the other hand, there is emerging evidence that DTNBP1 might also play a role in the genetics of affective psychosis. Large genetic case–control association studies of European [6, 19], Korean [31, 56], and multiplex Ashkenazi Jewish family samples [14] have shown a linkage of DTNBP1 with bipolar disorder. However, in parallel to the state of research on schizophrenia, there are also negative association studies [58].
So far, more than 20 different SNPs (single nucleotide polymorphisms) have been reported to be significantly associated with schizophrenia and/or bipolar disorder, either individually or within a haplotype [19, 24]. However, there is widespread inconsistency among the associated DTNPB1 haplotypes with a variety of combinations of SNPs and risk alleles reported throughout the literature [82], complicated even more by large genetic differences between major geographic populations [19, 24]. Thus, the question about the veritably underlying DTNBP1 schizophrenia or other psychiatric susceptibility variants still remains [82].
DTNBP1 encodes dysbindin-1 (β-dystrobrevin-binding protein or DTNBP1) [4, 64], a 40- to 50-kDa protein expressed neuronally [4] in many regions of the human brain [66]. DTNBP1 binds both α- and β-dystrobrevin, which are components of the dystrophin glycoprotein complex [4]. This complex is especially concentrated at postsynaptic densities [4] in diverse brain areas [4, 37], but only the β-dystrobrevin isoform is expressed in neurons [4]. By means of binding β-dystrobrevin, DTNBP1 interacts with the dystrophin glycoprotein complex at postsynaptic sites and might thereby alter neuronal synaptic mechanisms. However, independent of β-dystrobrevin and the dystrophin glycoprotein complex, DTNBP1 has also been found to be presynaptically located in glutamatergic neurons in the hippocampus [54, 69]. In this context, dysbindin-1 might play a role in glutamate signal transduction in the hippocampus formation [54].
DTNBP1 is ubiquitously expressed throughout the brain [70, 80] and corresponding to its primary localization to neurons, DTNBP1 mRNA has predominantly been detected in grey matter areas [80]. In the healthy adult brain, DTNBP1 mRNA is most prominently expressed in the frontal cortex (especially in the dorsolateral prefrontal cortex), temporal cortex, hippocampus, caudate, putamen, nucleus accumbens, amygdala, thalamus, and midbrain [80]. Post-mortem brain analyses of patients with schizophrenia showed significantly reduced DTNBP1 mRNA and protein expression in the dorsolateral prefrontal cortex (DLPFC) and midbrain [65, 66, 70] and in the hippocampal formation compared to healthy controls [54, 79]. Weickert et al. [79] reported a decrease in DTNBP1 mRNA levels in schizophrenic patients particularly in the dentate gyrus and CA3 (cornu ammonis 3) of the hippocampus. This decline of mRNA was positively correlated with the expression of other synaptic markers known to be reduced in schizophrenia [79].
Alterations in dopamine and glutamate signal transduction are considered to be the neurochemical basis of the pathophysiology of schizophrenia [3, 12, 20, 47, 57, 73], but also play an important role in other psychiatric conditions [16, 24, 39]. In this context, genetic variation in DTNBP1 might confer the risk of schizophrenia or other psychiatric disorders by mediating effects on synaptic structure and function of glutamatergic neurons, particularly in the hippocampus [48, 66].
Reductions in whole temporal lobe volumes and especially of the medial temporal lobe including the amygdala-hippocampus-complex are a major neuropathological finding in schizophrenia [62, 68, 76]. Most of all, reduced bilateral hippocampal volumes have been reported for chronic schizophrenia patients, but also for first-episode patients with psychosis and for patients with bipolar affective disorder [59, 68, 76]. Another principal neuropathological finding in schizophrenia is a reduced volume of prefrontal cortical regions [7, 22], which has also been described in bipolar patients to a lesser extent [49, 59]. Against this background, the aims of the present study were to directly investigate the effects of at-risk single nucleotide polymorphisms of the schizophrenia/psychiatric susceptibility gene DTNBP1 on regional brain volumes in human subjects using structural MRI. We chose the two single nucleotide polymorphisms rs2619522 and rs1018381, which have been investigated in several genetic studies [14, 17, 19, 38], and even referred to as a set for tagging a high-risk haplotype, reported by van den Oord et al. [75] in the sample of the Irish study on high-density schizophrenia families. We hypothesized that there might be alterations in grey matter volume of the prefrontal cortex and hippocampal regions associated with the at-risk DTNBP1 alleles.
Materials and methods
Subjects
Seventy-one (rs2619255) and seventy-two (rs1018381) subjects of a multi-diagnosis sample including nine healthy controls participated in the study (Table 1). In one case (male, 23 years, obsessive compulsive disorder), genotype analysis failed for SNP rs2619255. Apart from this, subjects were identical in both groups. Of the participating subjects, 22 were diagnosed with schizophrenia, 22 with bipolar affective disorder and 18 (rs2619255), 19 (rs1018381) with obsessive–compulsive disorder according to ICD-10 and DSM-IV criteria. The healthy controls exhibited no past or present psychiatric, neurological or medical disorder and had no positive family history of psychiatric disorders. Exclusion criteria in general were dementia, neurological illness, brain traumas, brain tumours and substance abuse. The patients were recruited from the Department of Psychiatry and Psychotherapy of the Saarland University Hospital between December 2003 and October 2006, and healthy controls were recruited from the hospital staff of the same institution and the local population. Written informed consent was obtained from all subjects prior to their inclusion into the investigation. The study was performed in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki and was approved by the local ethics committee.
Table 1.
Demographic and clinical data of study subjects
| rs2619522 | rs1018381 | |||
|---|---|---|---|---|
| All subjects (N) | 71 | 72 | ||
| Group comparison | GG/GT | TT | TT/CT | CC |
| Group size (N) | 23 (4 GG; 19 GT) | 48 | 10 (0 TT; 10 CT) | 62 |
| Gender (N) | 10 M, 13 F | 26 M, 22 F | 4 M, 5 F | 31 M, 31 F |
| Age (years ± SD) | 35.3 (±11.1) | 36.5 (±11.5) | 34.2 (±10.4) | 36.3 (±11.5) |
| Diagnosis: schizophrenia (N) | 8 | 14 | 4 | 18 |
| Diagnosis: bipolar disorder (N) | 7 | 15 | 2 | 20 |
| Diagnosis: OCD (N) | 4 | 14 | 1 | 18 |
| Control subjects (N) | 4 | 5 | 3 | 6 |
F female, M male, N number, SD standard deviation
The mean age of all participants was 36 ± 11.3 years (rs2619255) and 35.9 ± 11.4 years (rs1018381). Age was ranging from 19 to 65 years. Thirty-five subjects were female, and all subjects were Caucasian (Table 1). The groups were not matched for diagnosis, age, and gender; therefore, these parameters were included into all voxel-based morphometric analysis as covariates of no interest. All participating subjects were scanned in the Saarland University Hospital.
MRI data aquisition
Structural magnetic resonance imaging was performed on a 1.5-T scanner (Siemens, Erlangen, Germany). All participants underwent structural magnetic resonance imaging on the same scanner. A T1-weighted, MPRAGE/echo gradient sequence (TE = 4.42 ms, TR = 1,900 ms, TI = 700 ms, flip angle = 15°, FOV 256 × 256 mm) of 176 consecutive slices was acquired with a voxel size of 1 × 1 × 1 mm3. The magnetic resonance images were realigned in parallel to the anterior commisure—posterior commissure plane.
Voxel-based morphometry and statistical analysis
Images were converted to DICOM format and processed using MATLAB and SPM5 software (Wellcome Department of Imaging Neuroscience Group, London, UK; http://www.fil.ion.ucl.ac.uk/spm). Images were corrected for bias and tissue classified. After segmentation into grey and white matter tissue classes, they were spatially normalized by using the standard SPM5 unified segmentation [2]. The normalized grey matter maps were modulated and finally smoothed with a 12-mm FWHM Gaussian kernel. All preprocessing steps were conducted using standard procedures as implemented in SPM5, in particular the voxel-based morphometry (VBM) toolbox.
Two-sample t tests were performed contrasting different groups according to their genotype. For both SNPs, we compared risk allele-carriers with homozygous non-risk allele-carriers. Confounding variables such as age, gender, and diagnosis were included in all VBM analysis as covariates of no interest to control for variability in these variables. We excluded all voxels with grey matter values of less than 0.05 (absolute threshold masking). One-tailed t contrasts were then generated with p < 0.001 (uncorrected, voxel-level) and extent threshold = 100 voxels.
Genetic analyses
Single nucleotide polymorphism rs2619522 intron (G/T)
DNA was isolated from EDTA-blood samples and extracted according to the salting out procedure [46]. The following primers were used in the polymerase chain reaction (PCR): 5′-TGGGCCAGTGAAGTGAAAAT-3′ (forward) and 5′-TTGCAGCAAAACAGTACTCTCC-3′ (reverse).
The PCR was done in a 50-μl reaction mixture containing 100 ng of genomic DNA, 200 μM of each dNTPs, 10 pmol of each primer, 0.5 units Taq-polymerase, buffer D (KCl 50 mM, Tris–HCl 10 mM, Tween 20 0.025 %, BSA 0.025 mg/ml, and 2.0 mM MgCl2). The PCR had an initial temperature of 94 °C (5 min), followed by 35 cycles of denaturation (94 °C, 30 s), annealing (55.1 °C, 30 s), and extension (72 °C, 30 s). An extension period of 7 min at 72 °C followed the final cycle. The PCRs were done using an ABI GeneAmp®9700 cycler.
PCR products were digested at 55 °C using BseGI (FERMENTAS), separated on 2.0 % agarose gels (ROTI®GAROSE NEEO, Roth, Karlsruhe) and genotyped according to the resulting fragment lengths.
Single nucleotide polymorphism rs1018381 intron (C/T)
DNA was isolated from EDTA-blood samples and extracted according to the salting out procedure [46]. The following primers were used in the polymerase chain reaction (PCR): 5′-TGATTGAGGCTTTGGCTTTT-3′ (forward) and 5′-CCATGAGCATACCACAGCAC-3′ (reverse).
The PCR was done in a 50-μl reaction mixture containing 100 ng of genomic DNA, 200 μM of each dNTPs, 10 pmol of each primer, 0.5 units Taq-polymerase, buffer D (KCl 50 mM, Tris–HCl 10 mM, Tween 20 0.025 %, BSA 0.025 mg/ml, and 2.0 mM MgCl2). The PCR had an initial temperature of 94 °C (5 min), followed by 35 cycles of denaturation (94 °C, 30 s), annealing (55.1 °C, 30 s), and extension (72 °C, 30 s). An extension period of 7 min at 72 °C followed the final cycle. The PCRs were done using an ABI GeneAmp®9700 cycler.
PCR products were digested at 37 °C using NlaIII (NEB, Ipswich, MA, USA), separated on 2.0 % agarose gels (ROTI®GAROSE NEEO, Roth, Karlsruhe) and genotyped according to the resulting fragment lengths.
All genotypes were called by two individuals being blind to the clinical data.
Genotype group classification
Participants were divided into two groups according to their genotype for both at-risk SNPs (rs2619522, rs1018381). For rs2619522 (N = 71), the G (guanine) allele has been defined as the minor allele and has been associated with schizophrenia, schizoaffective disorder [17] and bipolar disorder [6, 19]. For statistical reasons, we contrasted homozygous non-risk allele-carriers T/T (T = thymine; N = 48) with homozygous and heterozygous risk allele-carriers (N = 23; Table 1).
Regarding rs1018381 (N = 72), the T allele is referred to as the minor allele and has been associated with schizophrenia and schizoaffective disorder [17]. In the present study, we contrasted risk allele-carriers (C/T; C = cytosine; N = 10) with homozygous non-risk allele-carriers (C/C; N = 62). No subject was homozygous for the risk allele (T/T) (Table 1).
The DTNBP1 allele distribution did not deviate from Hardy–Weinberg equilibrium for neither SNP (rs2619522: p = 0.2561; rs1018381: p = 1) [81].
Results
Single nucleotide polymorphism rs2619522
We found significant effects of the DTNBP1 SNP rs2619522 on grey matter brain volumes. The major finding was in the left hippocampus with significantly altered grey matter volume associated with the risk allele (G allele; Fig. 1). This result remained significant after false discovery rate (FDR)-correction (p FDR-corr = 0.042) and only scarcely missed the significance level for family wise error (FWE)-correction (p FWE-corr = 0.069).
Fig. 1.

Effects of DTNBP1 SNP rs2619522 on hippocampal grey matter volumes. Carriers of the G allele (risk allele) show significantly higher grey matter volumes in these brain regions than T/T homozygotes. p < 0.001 (uncorrected); extent threshold = 100 voxels
In addition, we found significant effects on a FDR-corrected level contralateral in the right hippocampus (Fig. 1) as well as in anterior prefrontal (Fig. 2) and intraparietal cortices (Fig. 3), in cortical regions of the temporal lobe and bilaterally in the cerebellum.
Fig. 2.
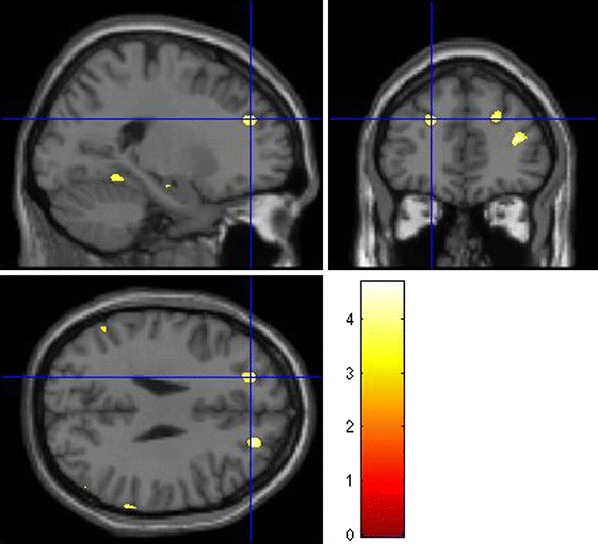
Effects of DTNBP1 SNP rs2619522 on grey matter volumes of anterior prefrontal cortex. Carriers of the G allele (risk allele) exhibit significantly higher grey matter volumes in these brain areas than T/T homozygotes. p < 0.001 (uncorrected); extent threshold = 100 voxels
Fig. 3.
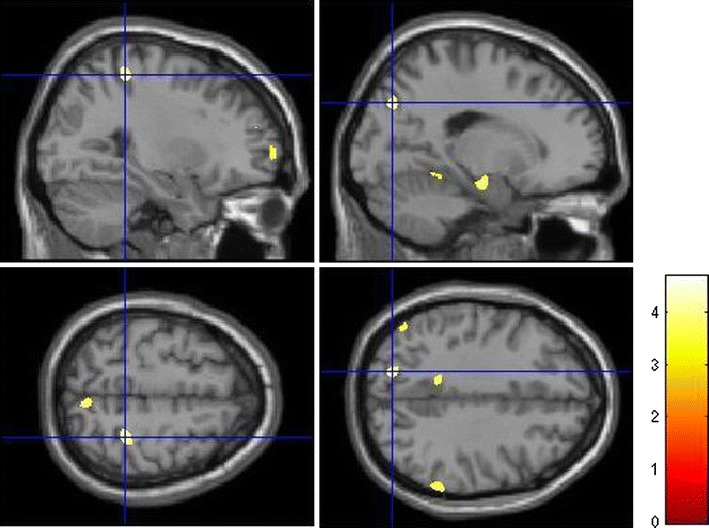
Effects of DTNBP1 SNP rs2619522 on grey matter volumes of parietal cortices. G allele-carriers (risk allele) exhibit significantly higher grey matter volumes in these cortical regions than T/T homozygotes. p < 0.001 (uncorrected); extent threshold = 100 voxels
Carriers of the G allele (risk allele) showed significantly higher grey matter volumes in all these brain regions than T/T homozygotes (Table 2).
Table 2.
DTNBP1 SNP rs2619522 G/G + G/T > T/T
| Anatomical regions | MNI coordinates | Statistical significance (T value) |
|---|---|---|
| Left hippocampus | −12/−13/−18 | 5.25 |
| Right hippocampus | 15/−16/−19 | 4.29 |
| Left lingual gyrus | −20/−48/−11 | 4.17 |
| Left lingual gyrus | −10/−48/−4 | 3.44 |
| Left superior frontal gyrus | −16/53/32 | 3.83 |
| Right superior frontal gyrus | 10/59/25 | 3.86 |
| Left middle frontal gyrus | −29/23/60 | 4.65 |
| −22/42/28 | 4.29 | |
| −32/47/31 | 3.99 | |
| −31/55/9 | 3.98 | |
| −44/47/5 | 3.95 | |
| Right middle frontal gyrus | 22/45/29 | 4.52 |
| 39/42/16 | 4.36 | |
| 31/61/4 | 4.20 | |
| 43/48/0 | 3.81 | |
| 28/60/16 | 3.52 | |
| Right frontopolar gyrus | 9/70/−6 | 3.93 |
| Left inferior frontal gyrus | −57/20/9 | 4.82 |
| Left inferior frontal gyrus, pars opercularis | −43/13/3 | 3.98 |
| Right inferior frontal gyrus | 41/23/5 | 4.36 |
| 51/30/−11 | 4.05 | |
| Left postcentral gyrus | −57/−21/17 | 3.78 |
| Right postcentral gyrus | 67/−20/14 | 3.64 |
| Left pregenual cingulate cortex | −5/54/2 | 3.66 |
| Left anterior cingulate cortex | −3/33/32 | 3.56 |
| (Right) anterior cingulate cortex | 0/45/25 | 4.07 |
| Left posterior cingulate gyrus | −11/−45/38 | 4.00 |
| Right posterior cingulate cortex | 6/−41/51 | 5.13 |
| 5/−55/31 | 3.68 | |
| Left superior temporal gyrus | −46/−8/2 | 4.01 |
| Right superior temporal gyrus | 57/−34/15 | 4.15 |
| Right superior temporal gyrus | 61/−44/8 | 3.97 |
| 70/−27/6 | 3.60 | |
| Left middle temporal gyrus | −58/−66/6 | 3.70 |
| Right middle temporal gyrus | 68/−6/−12 | 4.20 |
| 70/−36/−5 | 4.01 | |
| Left angular gyrus | −48/−69/38 | 4.21 |
| −54/−59/31 | 3.92 | |
| Right angular gyrus | 60/−45/39 | 4.65 |
| Right supramarginal gyrus | 52/−45/25 | 3.72 |
| Left precuneus | −2/−56/50 | 3.56 |
| Right precuneus | 6/−69/56 | 4.29 |
| Left superior occipital gyrus | −17/−77/37 | 4.99 |
| Right middle occipital gyrus | 59/−70/4 | 4.54 |
| Right postcentral gyrus | 55/−22/48 | 5.16 |
| 29/−41/57 | 4.31 | |
| Right precentral gyrus | 10/−7/75 | 4.25 |
| 51/−9/35 | 3.83 | |
| 26/−31/67 | 3.50 | |
| Left cerebellum | −27/−38/−52 | 4.12 |
| Right cerebellum | 16/−57/−10 | 3.71 |
| 23/−92/−29 | 3.70 | |
| Left inferior occipital gyrus | −13/−105/−3 | 3.92 |
p < 0.001 (uncorrected, voxel-level); extent threshold = 100 voxels
Single nucleotide polymorphism rs1018381
There were significant alterations of grey matter volume in association with the risk allele T (genotype C/T) on a FDR-correction level. T-carriers exhibited significantly higher brain volume bilaterally in frontal cortices as well as in the left lingual gyrus, the right thalamus/pulvinar and the left cerebellum (Table 3).
Table 3.
DTNBP1 SNP rs1018381 C/T > C/C
| Anatomical regions | MNI coordinates | Statistical significance (T value) |
|---|---|---|
| Right inferior frontal gyrus | 49/14/22 | 4.08 |
| Left lingual gyrus | −20/−50/−8 | 4.06 |
| Right thalamus/pulvinar | 8/−26/6 | 3.85 |
| Left middle frontal gyrus | −32/52/7 | 3.80 |
| Right middle frontal gyrus | 35/46/14 | 3.75 |
| Left cerebellum | −29/−54/−63 | 3.60 |
p < 0.001 (uncorrected, voxel-level); extent threshold = 100 voxels
Discussion
In the present study, we found variations of grey matter brain volumes independently associated with two different single nucleotide polymorphisms of the DTNBP1 gene, both together characterized as a set for tagging a schizophrenia high-risk haplotype reported by van den Oord et al. [75].
Regarding the SNP rs2619522, we found significantly higher grey matter volumes in several brain regions associated with the risk allele (G allele) [6, 17, 19]. The major finding was in the left hippocampus, which was significant on a FDR-correction level. Contralaterally, the right hippocampus also was significantly enlarged, although on a slightly lower significance level. Besides this main finding, we observed significant volume increases associated with the G allele in frontal and parietal cortices as well as in lateral and medial temporal areas.
Concerning the SNP rs1018381, we also found significantly higher grey matter volumes in association with the reported risk allele (T allele) [17]. Risk allele-carriers exhibited increased brain volumes bilaterally in frontal cortices, in the basal right thalamus/pulvinar and the left lingual gyrus.
Our results therefore complement the current state of research on schizophrenia and affective disorders with manifold findings relating to hippocampal and prefrontal morphology and function in the course of these psychiatric diseases [60, 62]. Schizophrenia has been associated with reduced hippocampal volume in patients as well as in their biologic relatives [5, 21, 28]. In bipolar disorder, findings of either enlarged [30] or reduced hippocampal volumes [13] have been described [60]. In addition, there are also several reports of asymmetric hippocampal volume alterations with emphasis on the left hippocampus in schizophrenia and individuals at ultra-high risk for psychosis [45, 77, 85], corresponding to our findings in association with rs2619522 (Table 2).
Hippocampal function, but also prefrontal and parietal cortical networks underlying higher cognitive functions, has been shown to be altered in schizophrenic and bipolar patients as well as in their healthy family members in several recent fMRI studies [11, 18, 23, 25, 26]. Moreover, cognitive impairment and abnormal executive functioning in schizophrenia and bipolar disorder have been reported in association with DTNBP1 risk genotypes [8, 15], and deviating brain activation in prefrontal, temporal and parietal areas during cognitive and emotional tasks has been described in healthy carriers of DTNBP1 risk alleles [33, 41, 42, 72, 84].
With this in mind, our findings of increased grey matter volumes in association with the DTNBP1 risk alleles of both at-risk SNPs in the above-mentioned brain regions seem to be counterintuitive at first sight, especially with respect to the reported structural brain volume reductions in schizophrenia.
However, the pathophysiology of functional psychosis is still far from being understood in detail. According to the empirically well-supported dopamine hypothesis in schizophrenia [27], productive psychotic states are associated with dopaminergic hyperactivity in subcortical regions [29, 35, 44] and positive symptoms as well as memory deficits have been linked to dysfunctional hippocampal hyperactivity [86]. Along with this, there is a body of evidence of dynamic structural brain alterations over the course of time in schizophrenia [63], which applies to the hippocampal formation in particular [45, 78].
Intriguingly, Nickl-Jockschat and co-workers recently reported reduced white matter fractional anisotropy in the right perihippocampal region in healthy T allele-carriers of DTNBP1 SNP rs1018381 [52]. At the same time, they also found elevated fibre tract integrity in bilateral frontal lobes in proximity to the bilateral medial frontal cortices, corresponding to our findings of higher grey matter volumes in association with the T allele (rs1018381) in these cortical brain areas.
Moreover, increased grey and white matter volumes in association with a schizophrenia risk allele-carrier status have also been described for a Neuregulin-1 at-risk SNP in childhood-onset schizophrenia [1]. It is worth remarking that these risk allele-associated volumetric alterations were found equally in schizophrenia patients and healthy controls at the beginning of that prospective investigation, but differed in terms of structural neurodevelopment in the longitudinal course between groups to the detriment of the schizophrenia risk allele-carriers [1].
In this sense, our findings of increased grey matter volumes in critical cerebral regions associated with the here-reported risk alleles of DTNBP1 might represent an endophenotype of a compromised brain [29], particularly vulnerable for developing further pathophysiological processes leading to clinical psychosis in the context of additional environmental stress. However, in which way and to what extent DTNBP1 risk genotypes exactly modulate brain morphology and function still is a topic of current research.
It is noteworthy though that the present study, despite the relatively small sample size and its diagnostic heterogeneity, could provide such distinct findings on grey matter volumes in association with the two at-risk SNPs of DTNBP1. Given the phenotypic and genetic overlap of schizophrenia with bipolar disorder [40], the present results support existing evidence that genetic variation of DTNBP1 may not only be a risk factor for schizophrenia, but also play a role in a broader context of psychiatric disorders including affective psychoses. Nevertheless, these results are limited by the diversity of the participating subjects and the findings will have to be replicated in a more homogeneous sample and with a higher number of subjects.
To conclude, compatible with previous reports on a role of the dysbindin-1 gene in hippocampal functions as well as in major psychoses, the present study provides first direct in vivo evidence that two risk alleles of DTNBP1 show linkage to altered grey matter volumes in the human brain. The risk alleles of rs2619522 and rs1018381 are significantly associated with variation of grey matter volumes in prefrontal areas and the latter (rs2619522) even with altered grey matter volumes in parietal cortices and bilaterally in the human hippocampus.
Acknowledgments
This work was supported by the Competence Network Schizophrenia. JM is supported by DFG (Deutsche Forschungsgemeinschaft) grants ME 1923/5-1 and ME 1923/5-3.
Conflict of interest
The authors declare that they have no conflict of interest.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Contributor Information
S. Trost, Phone: +49-551-3912289, FAX: +49-551-3912289, Email: s.trost@med.uni-goettingen.de
O. Gruber, Phone: +49-551-3912289, FAX: +49-551-3912289, Email: ogruber@gwdg.de
References
- 1.Addington AM, Gornick MC, Shaw P, Seal J, Gogtay N, Greenstein D, Clasen L, Coffey M, Gochman P, Long R, Rapoport JL. Neuregulin 1 (8p12) and childhood-onset schizophrenia: susceptibility haplotypes for diagnosis and brain developmental trajectories. Mol Psychiatry. 2007;12:195–205. doi: 10.1038/sj.mp.4001906. [DOI] [PubMed] [Google Scholar]
- 2.Ashburner J. A fast diffeomorphic image registration algorithm. Neuroimage. 2007;38:95–113. doi: 10.1016/j.neuroimage.2007.07.007. [DOI] [PubMed] [Google Scholar]
- 3.Benes FM. Emerging principles of altered neural circuitry in schizophrenia. Brain Res. 2000;31:251–269. doi: 10.1016/S0165-0173(99)00041-7. [DOI] [PubMed] [Google Scholar]
- 4.Benson MA, Newey SE, Martin-Rendon E, Hawkes R, Blake DJ. Dysbindin, a novel coiled-coil-containing protein that interacts with the dystrobrevins in muscle and brain. J Biol Chem. 2001;276:24232–24241. doi: 10.1074/jbc.M010418200. [DOI] [PubMed] [Google Scholar]
- 5.Boos HB, Aleman A, Cahn W, Hulshoff Pol H, Kahn RS. Brain volumes in relatives of patients with schizophrenia: a meta-analysis. Arch Gen Psychiatry. 2007;64:297–304. doi: 10.1001/archpsyc.64.3.297. [DOI] [PubMed] [Google Scholar]
- 6.Breen G, Prata D, Osborne S, Munro J, Sinclair M, Li T, Staddon S, Dempster D, Sainz R, Arroyo B, Kerwin RW, St Clair D, Collier D. Association of the dysbindin gene with bipolar affective disorder. Am J Psychiatry. 2006;163:1636–1638. doi: 10.1176/appi.ajp.163.9.1636. [DOI] [PubMed] [Google Scholar]
- 7.Buchanan RW, Vladar K, Barta PE, Pearlson GD. Structural evaluation of the prefrontal cortex in schizophrenia. Am J Psychiatry. 1998;155:1049–1055. doi: 10.1176/ajp.155.8.1049. [DOI] [PubMed] [Google Scholar]
- 8.Burdick KE, Goldberg TE, Funke B, Bates JA, Lencz T, Kucherlapati R, Malhotra AK. DTNBP1 genotype influences cognitive decline in schizophrenia. Schizophr Res. 2007;89:169–172. doi: 10.1016/j.schres.2006.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cannon TD, Thompson PM, van Erp TG, Huttunen M, Lonnqvist J, Kaprio J, Toga AW. Mapping heritability and molecular genetic associations with cortical features using probabilistic brain atlases: methods and applications to schizophrenia. Neuroinformatics. 2006;4:5–19. doi: 10.1385/NI:4:1:5. [DOI] [PubMed] [Google Scholar]
- 10.Cardno AG, Gottesman II. Twin studies of schizophrenia: from bow-and-arrow concordances to star wars Mx and functional genomics. Am J Med Genet. 2000;97:12–17. doi: 10.1002/(SICI)1096-8628(200021)97:1<12::AID-AJMG3>3.0.CO;2-U. [DOI] [PubMed] [Google Scholar]
- 11.Chen CH, Suckling J, Lennox BR, Ooi C, Bullmore ET. A quantitative meta-analysis of fMRI studies in bipolar disorder. Bipolar Disord. 2011;13:1–15. doi: 10.1111/j.1399-5618.2011.00893.x. [DOI] [PubMed] [Google Scholar]
- 12.Creese I, Burt DR, Snyder SH (1976) Dopamine receptor binding predicts clinical and pharmacological potencies of antischizophrenic drugs. Science (New York, NY) 192:481–483 [DOI] [PubMed]
- 13.Doring TM, Kubo TT, Cruz LC, Jr, Juruena MF, Fainberg J, Domingues RC, Gasparetto EL. Evaluation of hippocampal volume based on MR imaging in patients with bipolar affective disorder applying manual and automatic segmentation techniques. J Magn Reson Imaging. 2011;33:565–572. doi: 10.1002/jmri.22473. [DOI] [PubMed] [Google Scholar]
- 14.Fallin MD, Lasseter VK, Avramopoulos D, Nicodemus KK, Wolyniec PS, McGrath JA, Steel G, Nestadt G, Liang KY, Huganir RL, Valle D, Pulver AE. Bipolar I disorder and schizophrenia: a 440-single-nucleotide polymorphism screen of 64 candidate genes among Ashkenazi Jewish case-parent trios. Am J Hum Genet. 2005;77:918–936. doi: 10.1086/497703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Fatjo-Vilas M, Papiol S, Estrada G, Bombin I, Peralta V, Rosa A, Parellada M, Miret S, Martin M, Lazaro L, Campanera S, Munoz MJ, Lera-Miguel S, Arias B, Navarro ME, Castro-Fornieles J, Cuesta MJ, Arango C, Fananas L. Dysbindin-1 gene contributes differentially to early- and adult-onset forms of functional psychosis. Am J Med Genet B Neuropsychiatr Genet. 2010;156B:322–333. doi: 10.1002/ajmg.b.31166. [DOI] [PubMed] [Google Scholar]
- 16.Fountoulakis KN (2012) The possible involvement of NMDA glutamate receptor in the etiopathogenesis of bipolar disorder. Curr Pharm Des 18:1605–1608 [DOI] [PubMed]
- 17.Funke B, Finn CT, Plocik AM, Lake S, DeRosse P, Kane JM, Kucherlapati R, Malhotra AK. Association of the DTNBP1 locus with schizophrenia in a US population. Am J Hum Genet. 2004;75:891–898. doi: 10.1086/425279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fusar-Poli P, Howes O, Bechdolf A, Borgwardt S. Mapping vulnerability to bipolar disorder: a systematic review and meta-analysis of neuroimaging studies. J Psychiatry Neurosci. 2011;37:110061. doi: 10.1503/jpn.110061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gaysina D, Cohen-Woods S, Chow PC, Martucci L, Schosser A, Ball HA, Tozzi F, Perry J, Muglia P, Craig IW, McGuffin P, Farmer A. Association of the dystrobrevin binding protein 1 gene (DTNBP1) in a bipolar case-control study (BACCS) Am J Med Genet B Neuropsychiatr Genet. 2009;150B:836–844. doi: 10.1002/ajmg.b.30906. [DOI] [PubMed] [Google Scholar]
- 20.Goff DC, Coyle JT. The emerging role of glutamate in the pathophysiology and treatment of schizophrenia. Am J Psychiatry. 2001;158:1367–1377. doi: 10.1176/appi.ajp.158.9.1367. [DOI] [PubMed] [Google Scholar]
- 21.Goldman AL, Pezawas L, Mattay VS, Fischl B, Verchinski BA, Zoltick B, Weinberger DR, Meyer-Lindenberg A. Heritability of brain morphology related to schizophrenia: a large-scale automated magnetic resonance imaging segmentation study. Biol Psychiatry. 2008;63:475–483. doi: 10.1016/j.biopsych.2007.06.006. [DOI] [PubMed] [Google Scholar]
- 22.Goldstein JM, Goodman JM, Seidman LJ, Kennedy DN, Makris N, Lee H, Tourville J, Caviness VS, Jr, Faraone SV, Tsuang MT. Cortical abnormalities in schizophrenia identified by structural magnetic resonance imaging. Arch Gen Psychiatry. 1999;56:537–547. doi: 10.1001/archpsyc.56.6.537. [DOI] [PubMed] [Google Scholar]
- 23.Gruber O, Gruber E, Falkai P. Articulatory rehearsal in verbal working memory: a possible neurocognitive endophenotype that differentiates between schizophrenia and schizoaffective disorder. Neurosci Lett. 2006;405:24–28. doi: 10.1016/j.neulet.2006.06.062. [DOI] [PubMed] [Google Scholar]
- 24.Guo AY, Sun J, Riley BP, Thiselton DL, Kendler KS, Zhao Z. The dystrobrevin-binding protein 1 gene: features and networks. Mol Psychiatry. 2009;14:18–29. doi: 10.1038/mp.2008.88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gur RE, Gur RC (2010) Functional magnetic resonance imaging in schizophrenia. Dialog Clin Neurosci 12:333–343 [DOI] [PMC free article] [PubMed]
- 26.Henseler I, Falkai P, Gruber O. A systematic fMRI investigation of the brain systems subserving different working memory components in schizophrenia. Eur J Neurosci. 2009;30:693–702. doi: 10.1111/j.1460-9568.2009.06850.x. [DOI] [PubMed] [Google Scholar]
- 27.Hirvonen J, Hietala J (2011) Dysfunctional brain networks and genetic risk for schizophrenia: specific neurotransmitter systems. CNS Neurosci Ther 17:89–96 [DOI] [PMC free article] [PubMed]
- 28.Ho BC, Magnotta V. Hippocampal volume deficits and shape deformities in young biological relatives of schizophrenia probands. NeuroImage. 2010;49:3385–3393. doi: 10.1016/j.neuroimage.2009.11.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Howes OD, Kapur S. The dopamine hypothesis of schizophrenia: version III—the final common pathway. Schizophr Bull. 2009;35:549–562. doi: 10.1093/schbul/sbp006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Javadapour A, Malhi GS, Ivanovski B, Chen X, Wen W, Sachdev P. Hippocampal volumes in adults with bipolar disorder. J Neuropsychiatr Clin Neurosci. 2010;22:55–62. doi: 10.1176/appi.neuropsych.22.1.55. [DOI] [PubMed] [Google Scholar]
- 31.Joo EJ, Lee KY, Jeong SH, Chang JS, Ahn YM, Koo YJ, Kim YS. Dysbindin gene variants are associated with bipolar I disorder in a Korean population. Neurosci Lett. 2007;418:272–275. doi: 10.1016/j.neulet.2007.03.037. [DOI] [PubMed] [Google Scholar]
- 32.Kendler KS, MacLean CJ, O’Neill FA, Burke J, Murphy B, Duke F, Shinkwin R, Easter SM, Webb BT, Zhang J, Walsh D, Straub RE. Evidence for a schizophrenia vulnerability locus on chromosome 8p in the Irish study of high-density schizophrenia families. Am J Psychiatry. 1996;153:1534–1540. doi: 10.1176/ajp.153.12.1534. [DOI] [PubMed] [Google Scholar]
- 33.Kircher T, Markov V, Krug A, Eggermann T, Zerres K, Nothen MM, Skowronek MH, Rietschel M. Association of the DTNBP1 genotype with cognition and personality traits in healthy subjects. Psychol Med. 2009;39:1657–1665. doi: 10.1017/S0033291709005388. [DOI] [PubMed] [Google Scholar]
- 34.Kirov G, Ivanov D, Williams NM, Preece A, Nikolov I, Milev R, Koleva S, Dimitrova A, Toncheva D, O’Donovan MC, Owen MJ. Strong evidence for association between the dystrobrevin binding protein 1 gene (DTNBP1) and schizophrenia in 488 parent-offspring trios from Bulgaria. Biol Psychiatry. 2004;55:971–975. doi: 10.1016/j.biopsych.2004.01.025. [DOI] [PubMed] [Google Scholar]
- 35.Laruelle M, Abi-Dargham A, Gil R, Kegeles L, Innis R. Increased dopamine transmission in schizophrenia: relationship to illness phases. Biol Psychiatry. 1999;46:56–72. doi: 10.1016/S0006-3223(99)00067-0. [DOI] [PubMed] [Google Scholar]
- 36.Li D, He L. Association study between the dystrobrevin binding protein 1 gene (DTNBP1) and schizophrenia: a meta-analysis. Schizophr Res. 2007;96:112–118. doi: 10.1016/j.schres.2007.05.017. [DOI] [PubMed] [Google Scholar]
- 37.Lidov HG (1996) Dystrophin in the nervous system. Brain Pathol (Zurich, Switzerland) 6:63–77 [DOI] [PubMed]
- 38.Luciano M, Miyajima F, Lind PA, Bates TC, Horan M, Harris SE, Wright MJ, Ollier WE, Hayward C, Pendleton N, Gow AJ, Visscher PM, Starr JM, Deary IJ, Martin NG, Payton A. Variation in the dysbindin gene and normal cognitive function in three independent population samples. Genes Brain Behav. 2009;8:218–227. doi: 10.1111/j.1601-183X.2008.00462.x. [DOI] [PubMed] [Google Scholar]
- 39.Maddock RJ, Buonocore MH (2011) MR spectroscopic studies of the brain in psychiatric disorders. Curr Top Behav Neurosci [Epub ahead of print] [DOI] [PubMed]
- 40.Maier W. Common risk genes for affective and schizophrenic psychoses. Eur Arch Psychiatry Clin Neurosci. 2008;258(Suppl 2):37–40. doi: 10.1007/s00406-008-2008-z. [DOI] [PubMed] [Google Scholar]
- 41.Markov V, Krug A, Krach S, Jansen A, Eggermann T, Zerres K, Stocker T, Shah NJ, Nothen MM, Treutlein J, Rietschel M, Kircher T. Impact of schizophrenia-risk gene dysbindin 1 on brain activation in bilateral middle frontal gyrus during a working memory task in healthy individuals. Hum Brain Mapp. 2010;31:266–275. doi: 10.1002/hbm.20862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Markov V, Krug A, Krach S, Whitney C, Eggermann T, Zerres K, Stocker T, Shah NJ, Nothen MM, Treutlein J, Rietschel M, Kircher T. Genetic variation in schizophrenia-risk-gene dysbindin 1 modulates brain activation in anterior cingulate cortex and right temporal gyrus during language production in healthy individuals. NeuroImage. 2009;47:2016–2022. doi: 10.1016/j.neuroimage.2009.05.067. [DOI] [PubMed] [Google Scholar]
- 43.McGuffin P, Owen MJ, Farmer AE. Genetic basis of schizophrenia. Lancet. 1995;346:678–682. doi: 10.1016/S0140-6736(95)92285-7. [DOI] [PubMed] [Google Scholar]
- 44.Meisenzahl EM, Schmitt GJ, Scheuerecker J, Moller HJ (2007) The role of dopamine for the pathophysiology of schizophrenia. Int Rev Psychiatry (Abingdon, England) 19:337–345 [DOI] [PubMed]
- 45.Meisenzahl EM, Seifert D, Bottlender R, Teipel S, Zetzsche T, Jager M, Koutsouleris N, Schmitt G, Scheuerecker J, Burgermeister B, Hampel H, Rupprecht T, Born C, Reiser M, Moller HJ, Frodl T. Differences in hippocampal volume between major depression and schizophrenia: a comparative neuroimaging study. Eur Arch Psychiatry Clin Neurosci. 2009;260:127–137. doi: 10.1007/s00406-009-0023-3. [DOI] [PubMed] [Google Scholar]
- 46.Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988;16:1215. doi: 10.1093/nar/16.3.1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Mirnics K, Middleton FA, Lewis DA, Levitt P. Analysis of complex brain disorders with gene expression microarrays: schizophrenia as a disease of the synapse. Trends Neurosci. 2001;24:479–486. doi: 10.1016/S0166-2236(00)01862-2. [DOI] [PubMed] [Google Scholar]
- 48.Moghaddam B. Bringing order to the glutamate chaos in schizophrenia. Neuron. 2003;40:881–884. doi: 10.1016/S0896-6273(03)00757-8. [DOI] [PubMed] [Google Scholar]
- 49.Molina V, Galindo G, Cortes B, de Herrera AG, Ledo A, Sanz J, Montes C, Hernandez-Tamames JA. Different gray matter patterns in chronic schizophrenia and chronic bipolar disorder patients identified using voxel-based morphometry. Eur Arch Psychiatry Clin Neurosci. 2010;261:313–322. doi: 10.1007/s00406-010-0183-1. [DOI] [PubMed] [Google Scholar]
- 50.Morris DW, McGhee KA, Schwaiger S, Scully P, Quinn J, Meagher D, Waddington JL, Gill M, Corvin AP. No evidence for association of the dysbindin gene [DTNBP1] with schizophrenia in an Irish population-based study. Schizophr Res. 2003;60:167–172. doi: 10.1016/S0920-9964(02)00527-3. [DOI] [PubMed] [Google Scholar]
- 51.Mutsuddi M, Morris DW, Waggoner SG, Daly MJ, Scolnick EM, Sklar P. Analysis of high-resolution HapMap of DTNBP1 (Dysbindin) suggests no consistency between reported common variant associations and schizophrenia. Am J Hum Genet. 2006;79:903–909. doi: 10.1086/508942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Nickl-Jockschat T, Stocker T, Markov V, Krug A, Huang R, Schneider F, Habel U, Zerres K, Nothen MM, Treutlein J, Rietschel M, Shah NJ, Kircher T. The impact of a Dysbindin schizophrenia susceptibility variant on fiber tract integrity in healthy individuals: a TBSS-based diffusion tensor imaging study. NeuroImage. 2011;60:847–853. doi: 10.1016/j.neuroimage.2011.10.012. [DOI] [PubMed] [Google Scholar]
- 53.Owen MJ, Williams HJ, O’Donovan MC. Schizophrenia genetics: advancing on two fronts. Curr Opin Genet Dev. 2009;19:266–270. doi: 10.1016/j.gde.2009.02.008. [DOI] [PubMed] [Google Scholar]
- 54.Owen MJ, Williams NM, O’Donovan MC. Dysbindin-1 and schizophrenia: from genetics to neuropathology. J Clin Investig. 2004;113:1255–1257. doi: 10.1172/JCI21470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Pae CU, Mandelli L, De Ronchi D, Kim JJ, Jun TY, Patkar AA, Serretti A. Dysbindin gene (DTNBP1) and schizophrenia in Korean population. Eur Arch Psychiatry Clin Neurosci. 2009;259:137–142. doi: 10.1007/s00406-008-0830-y. [DOI] [PubMed] [Google Scholar]
- 56.Pae CU, Serretti A, Mandelli L, Yu HS, Patkar AA, Lee CU, Lee SJ, Jun TY, Lee C, Paik IH, Kim JJ. Effect of 5-haplotype of dysbindin gene (DTNBP1) polymorphisms for the susceptibility to bipolar I disorder. Am J Med Genet B Neuropsychiatr Genet. 2007;144B:701–703. doi: 10.1002/ajmg.b.30439. [DOI] [PubMed] [Google Scholar]
- 57.Papaleo F, Weinberger DR. Dysbindin and Schizophrenia: it’s dopamine and glutamate all over again. Biol Psychiatry. 2011;69:2–4. doi: 10.1016/j.biopsych.2010.10.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Raybould R, Green EK, MacGregor S, Gordon-Smith K, Heron J, Hyde S, Caesar S, Nikolov I, Williams N, Jones L, O’Donovan MC, Owen MJ, Jones I, Kirov G, Craddock N. Bipolar disorder and polymorphisms in the dysbindin gene (DTNBP1) Biol Psychiatry. 2005;57:696–701. doi: 10.1016/j.biopsych.2005.01.018. [DOI] [PubMed] [Google Scholar]
- 59.Rimol LM, Hartberg CB, Nesvag R, Fennema-Notestine C, Hagler DJ, Jr, Pung CJ, Jennings RG, Haukvik UK, Lange E, Nakstad PH, Melle I, Andreassen OA, Dale AM, Agartz I. Cortical thickness and subcortical volumes in schizophrenia and bipolar disorder. Biol Psychiatry. 2010;68:41–50. doi: 10.1016/j.biopsych.2010.03.036. [DOI] [PubMed] [Google Scholar]
- 60.Savitz J, Drevets WC. Bipolar and major depressive disorder: neuroimaging the developmental-degenerative divide. Neurosci Biobehav Rev. 2009;33:699–771. doi: 10.1016/j.neubiorev.2009.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Schwab SG, Knapp M, Mondabon S, Hallmayer J, Borrmann-Hassenbach M, Albus M, Lerer B, Rietschel M, Trixler M, Maier W, Wildenauer DB. Support for association of schizophrenia with genetic variation in the 6p22.3 gene, dysbindin, in sib-pair families with linkage and in an additional sample of triad families. Am J Hum Genet. 2003;72:185–190. doi: 10.1086/345463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Shenton ME, Dickey CC, Frumin M, McCarley RW. A review of MRI findings in schizophrenia. Schizophr Res. 2001;49:1–52. doi: 10.1016/S0920-9964(01)00163-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Shenton ME, Whitford TJ, Kubicki M. Structural neuroimaging in schizophrenia: from methods to insights to treatments. Dialog Clin Neurosci. 2010;12:317–332. doi: 10.31887/DCNS.2010.12.3/mshenton. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sillitoe RV, Benson MA, Blake DJ, Hawkes R. Abnormal dysbindin expression in cerebellar mossy fiber synapses in the mdx mouse model of Duchenne muscular dystrophy. J Neurosci. 2003;23:6576–6585. doi: 10.1523/JNEUROSCI.23-16-06576.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Straub RE (2004) Dysbindin protein is decreased in the dorsolateral prefrontal cortex of schizophrenia patients. Biol Psychiatry 55(Suppl 1):116S
- 66.Straub RE, Jiang Y, MacLean CJ, Ma Y, Webb BT, Myakishev MV, Harris-Kerr C, Wormley B, Sadek H, Kadambi B, Cesare AJ, Gibberman A, Wang X, O’Neill FA, Walsh D, Kendler KS. Genetic variation in the 6p22.3 gene DTNBP1, the human ortholog of the mouse dysbindin gene, is associated with schizophrenia. Am J Hum Genet. 2002;71:337–348. doi: 10.1086/341750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Strohmaier J, Frank J, Wendland JR, Schumacher J, Jamra RA, Treutlein J, Nieratschker V, Breuer R, Mattheisen M, Herms S, Muhleisen TW, Maier W, Nothen MM, Cichon S, Rietschel M, Schulze TG. A reappraisal of the association between Dysbindin (DTNBP1) and schizophrenia in a large combined case-control and family-based sample of German ancestry. Schizophr Res. 2010;118:98–105. doi: 10.1016/j.schres.2009.12.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Suddath RL, Christison GW, Torrey EF, Casanova MF, Weinberger DR. Anatomical abnormalities in the brains of monozygotic twins discordant for schizophrenia. N Engl J Med. 1990;322:789–794. doi: 10.1056/NEJM199003223221201. [DOI] [PubMed] [Google Scholar]
- 69.Talbot K, Eidem WL, Tinsley CL, Benson MA, Thompson EW, Smith RJ, Hahn CG, Siegel SJ, Trojanowski JQ, Gur RE, Blake DJ, Arnold SE. Dysbindin-1 is reduced in intrinsic, glutamatergic terminals of the hippocampal formation in schizophrenia. J Clin Investig. 2004;113:1353–1363. doi: 10.1172/JCI20425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Tang J, LeGros RP, Louneva N, Yeh L, Cohen JW, Hahn CG, Blake DJ, Arnold SE, Talbot K. Dysbindin-1 in dorsolateral prefrontal cortex of schizophrenia cases is reduced in an isoform-specific manner unrelated to dysbindin-1 mRNA expression. Hum Mol Genet. 2009;18:3851–3863. doi: 10.1093/hmg/ddp329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Tang JX, Zhou J, Fan JB, Li XW, Shi YY, Gu NF, Feng GY, Xing YL, Shi JG, He L. Family-based association study of DTNBP1 in 6p22.3 and schizophrenia. Mol Psychiatry. 2003;8:717–718. doi: 10.1038/sj.mp.4001287. [DOI] [PubMed] [Google Scholar]
- 72.Thimm M, Krug A, Kellermann T, Markov V, Krach S, Jansen A, Zerres K, Eggermann T, Stocker T, Shah NJ, Nothen MM, Rietschel M, Kircher T. The effects of a DTNBP1 gene variant on attention networks: an fMRI study. Behav Brain Funct. 2010;6:54. doi: 10.1186/1744-9081-6-54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Tsai G, Coyle JT. Glutamatergic mechanisms in schizophrenia. Annu Rev Pharmacol Toxicol. 2002;42:165–179. doi: 10.1146/annurev.pharmtox.42.082701.160735. [DOI] [PubMed] [Google Scholar]
- 74.Van Den Bogaert A, Schumacher J, Schulze TG, Otte AC, Ohlraun S, Kovalenko S, Becker T, Freudenberg J, Jonsson EG, Mattila-Evenden M, Sedvall GC, Czerski PM, Kapelski P, Hauser J, Maier W, Rietschel M, Propping P, Nothen MM, Cichon S. The DTNBP1 (dysbindin) gene contributes to schizophrenia, depending on family history of the disease. Am J Hum Genet. 2003;73:1438–1443. doi: 10.1086/379928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.van den Oord EJ, Sullivan PF, Jiang Y, Walsh D, O’Neill FA, Kendler KS, Riley BP. Identification of a high-risk haplotype for the dystrobrevin binding protein 1 (DTNBP1) gene in the Irish study of high-density schizophrenia families. Mol Psychiatry. 2003;8:499–510. doi: 10.1038/sj.mp.4001263. [DOI] [PubMed] [Google Scholar]
- 76.Velakoulis D, Pantelis C, McGorry PD, Dudgeon P, Brewer W, Cook M, Desmond P, Bridle N, Tierney P, Murrie V, Singh B, Copolov D. Hippocampal volume in first-episode psychoses and chronic schizophrenia: a high-resolution magnetic resonance imaging study. Arch Gen Psychiatry. 1999;56:133–141. doi: 10.1001/archpsyc.56.2.133. [DOI] [PubMed] [Google Scholar]
- 77.Velakoulis D, Wood SJ, Wong MT, McGorry PD, Yung A, Phillips L, Smith D, Brewer W, Proffitt T, Desmond P, Pantelis C. Hippocampal and amygdala volumes according to psychosis stage and diagnosis: a magnetic resonance imaging study of chronic schizophrenia, first-episode psychosis, and ultra-high-risk individuals. Arch Gen Psychiatry. 2006;63:139–149. doi: 10.1001/archpsyc.63.2.139. [DOI] [PubMed] [Google Scholar]
- 78.Wang L, Mamah D, Harms MP, Karnik M, Price JL, Gado MH, Thompson PA, Barch DM, Miller MI, Csernansky JG. Progressive deformation of deep brain nuclei and hippocampal-amygdala formation in schizophrenia. Biol Psychiatry. 2008;64:1060–1068. doi: 10.1016/j.biopsych.2008.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Weickert CS, Rothmond DA, Hyde TM, Kleinman JE, Straub RE. Reduced DTNBP1 (dysbindin-1) mRNA in the hippocampal formation of schizophrenia patients. Schizophr Res. 2008;98:105–110. doi: 10.1016/j.schres.2007.05.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Weickert CS, Straub RE, McClintock BW, Matsumoto M, Hashimoto R, Hyde TM, Herman MM, Weinberger DR, Kleinman JE. Human dysbindin (DTNBP1) gene expression in normal brain and in schizophrenic prefrontal cortex and midbrain. Arch Gen Psychiatry. 2004;61:544–555. doi: 10.1001/archpsyc.61.6.544. [DOI] [PubMed] [Google Scholar]
- 81.Wigginton JE, Cutler DJ, Abecasis GR. A note on exact tests of Hardy-Weinberg equilibrium. Am J Hum Genet. 2005;76:887–893. doi: 10.1086/429864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Williams NM, O’Donovan MC, Owen MJ. Is the dysbindin gene (DTNBP1) a susceptibility gene for schizophrenia? Schizophr Bull. 2005;31:800–805. doi: 10.1093/schbul/sbi061. [DOI] [PubMed] [Google Scholar]
- 83.Williams NM, Preece A, Morris DW, Spurlock G, Bray NJ, Stephens M, Norton N, Williams H, Clement M, Dwyer S, Curran C, Wilkinson J, Moskvina V, Waddington JL, Gill M, Corvin AP, Zammit S, Kirov G, Owen MJ, O’Donovan MC. Identification in 2 independent samples of a novel schizophrenia risk haplotype of the dystrobrevin binding protein gene (DTNBP1) Arch Gen Psychiatry. 2004;61:336–344. doi: 10.1001/archpsyc.61.4.336. [DOI] [PubMed] [Google Scholar]
- 84.Wolf C, Jackson MC, Kissling C, Thome J, Linden DE. Dysbindin-1 genotype effects on emotional working memory. Mol Psychiatry. 2011;16:145–155. doi: 10.1038/mp.2009.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wood SJ, Kennedy D, Phillips LJ, Seal ML, Yucel M, Nelson B, Yung AR, Jackson G, McGorry PD, Velakoulis D, Pantelis C. Hippocampal pathology in individuals at ultra-high risk for psychosis: a multi-modal magnetic resonance study. NeuroImage. 2010;52:62–68. doi: 10.1016/j.neuroimage.2010.04.012. [DOI] [PubMed] [Google Scholar]
- 86.Zierhut K, Bogerts B, Schott B, Fenker D, Walter M, Albrecht D, Steiner J, Schutze H, Northoff G, Duzel E, Schiltz K. The role of hippocampus dysfunction in deficient memory encoding and positive symptoms in schizophrenia. Psychiatry Res. 2010;183:187–194. doi: 10.1016/j.pscychresns.2010.03.007. [DOI] [PubMed] [Google Scholar]
- 87.Zuo L, Luo X, Kranzler HR, Lu L, Rosenheck RA, Cramer J, van Kammen DP, Erdos J, Charney DS, Krystal J, Gelernter J. Association study of DTNBP1 with schizophrenia in a US sample. Psychiatr Genet. 2009;19:292–304. doi: 10.1097/YPG.0b013e32832a50bc. [DOI] [PMC free article] [PubMed] [Google Scholar]