

Abstract
Interactions between cells and their surroundings are important for proper function and homeostasis in a multicellular organism. These interactions can either be established between the cells and molecules in their extracellular milieu, but also involve interactions between cells. In all these situations, proteins in the plasma membranes are critically involved to relay information obtained from the exterior of the cell. The cell surface glycoprotein CD47 (integrin-associated protein (IAP)) was first identified as an important regulator of integrin function, but later also was shown to function in ways that do not necessarily involve integrins. Ligation of CD47 can induce intracellular signaling resulting in cell activation or cell death depending on the exact context. By binding to another cell surface glycoprotein, signal regulatory protein alpha (SIRPα), CD47 can regulate the function of cells in the monocyte/macrophage lineage. In this spotlight paper, several functions of CD47 will be reviewed, although some functions may be more briefly mentioned. Focus will be on the ways CD47 regulates hematopoietic cells and functions such as CD47 signaling, induction of apoptosis, and regulation of phagocytosis or cell-cell fusion.
1. Structure and Expression of CD47 in the Plasma Membrane
CD47 (originally named integrin-associated protein (IAP)) is a cell surface protein of the immunoglobulin (Ig) superfamily, which is heavily glycosylated and expressed by virtually all cells in the body [1]. CD47 was first recognized as a 50 kDa protein associated and copurified with the α v β 3 integrin in placenta and neutrophil granulocytes and later shown to have the capacity to regulate integrin function and the responsiveness of leukocytes to RGD-containing extracellular matrix proteins [1–4]. Soon after this integrin-associated protein was cloned, it was shown to be identical to the erythrocyte cell surface antigen CD47 [5]. The fact that CD47 is also expressed by cells like erythrocytes, that do not express integrins, indicates that it can be more appropriate to refer to this protein as CD47 than using its original name integrin-associated protein (IAP). The protein is fairly well conserved between species and has about 60–70% similarity in the amino acid sequence when comparing human CD47 with that of mouse, rat, and bovine CD47. CD47 has also been shown to be identical to the OA-3/OVTL3 antigen highly expressed on most ovarian carcinomas [6, 7]. It also shows homology to a protein family of variola and vaccinia viruses [1, 5, 6], the significance of which is still unclear. CD47 consists of an extracellular IgV domain, a five times transmembrane-spanning domain, and a short alternatively spliced cytoplasmic tail. In both humans and mice, the cytoplasmic tail can be found as four different splice isoforms ranging from 4 to 36 amino acids, showing different tissue expression patterns. The 16 amino acid form 2, which is by far the predominant isoform, is expressed in all cells of hematopoietic origin, as well as in endothelial and epithelial cells [8]. In contrast, the 36 amino acid form 4 is expressed primarily in neurons, intestine, and testis [8]. Expression of the 4 amino acid form 1 is found in epithelial and endothelial cells, while the expression pattern of the 23 amino acid form 3 resembles that of form 4 [8]. Despite a study showing that CD47 form 3 and 4 could be associated with memory retention in rats, and that form 2 is predominating in astrocytes [9], little is known as to what these splice variants mean in terms of possible difference in the functionality of the protein. A new twist in the understanding of CD47 comes from a recent finding that CD47 can be expressed as a proteoglycan with a molecular weight of >250 kDa, having both heparin and chondroitin sulfate glycosaminoglycan (GAG) chains [10]. This form of CD47 was found to be expressed in both the human Jurkat T cell line and in murine primary T cells, as well as in human umbilical vein endothelial cells (HUVECs), murine lung endothelial cells, and in human smooth muscle cells [10]. Functionally, the GAG chains at CD47 Ser64 were found to be crucial to inhibit T cell receptor signaling following the ligation of CD47 by its ligand thrombospondin-1 [10].
2. Intracellular Signaling following CD47 Interactions with Integrins, Thrombospondin (TSP), or Signal Regulatory Proteins (SIRPs)
2.1. Interaction with Integrins
Early studies of CD47 were based on the use of monoclonal antibodies (mAbs) raised against the CD47 protein purified from placenta [2, 11], showing a role of CD47 in mediating an enhanced IgG-mediated phagocytosis response in the presence of RGD-containing ligands, such as fibronectin, fibrinogen, vitronectin, or collagen type IV [2, 11]. The same mAbs were also found to block neutrophil transendothelial migration stimulated by interleukin 8 (IL-8) or the bacterial peptide N-formyl-methionyl-leucyl-phenylalanine (f-Met-Leu-Phe) and to inhibit neutrophil migration across tumor-necrosis-factor-α- (TNFα-) stimulated endothelial cells, where CD47 on both the neutrophils and the endothelial cells was found to be important [12]. Generation of other anti-CD47mAbs, raised against epithelial membrane preparations, showed that CD47 is present at the basolateral membrane of epithelial cell monolayers, that mAbs blocking CD47 on either neutrophils or the epithelial cells delay neutrophil trans-epithelial migration [13], and that efficient neutrophil chemotaxis correlates with an increased neutrophil cell surface expression of CD47 [14]. The basement membrane protein entactin, which contains an RGD sequence, was also found to stimulate neutrophil adhesion and chemotaxis in a CD47-dependent manner in vitro [15]. Generation of CD47-deficient mice further proved the importance of this protein in regulating neutrophil inflammatory responses, by showing an increased sensitivity to bacterial infection due to a delayed neutrophil accumulation in bacterial peritonitis [4]. CD47-deficient neutrophils also show a strongly impaired RGD-stimulated neutrophil adhesion, phagocytosis, and respiratory burst [4]. For α v β 3 integrin-mediated cellular responses to the extracellular matrix protein vitronectin, CD47 was found to be required for α v β 3-mediated binding to vitronectin-coated beads, but not α v β 3-mediated adhesion to vitronectin-coated surfaces [3]. In addition to its original association with α v β 3 integrins, CD47 has also been shown to interact with and regulate the integrins α 2 β 1 and αIIb β 3 on platelets [16, 17], the α 2 β 1 integrin on smooth muscle cells [18], the α 4 β 1 integrin on sickle red blood cells and B lymphocytes [19, 20], the α 6 β 1 integrin in microglia [21], and the α 5 integrin in chondrocytes [22] (Figure 1).
Figure 1.
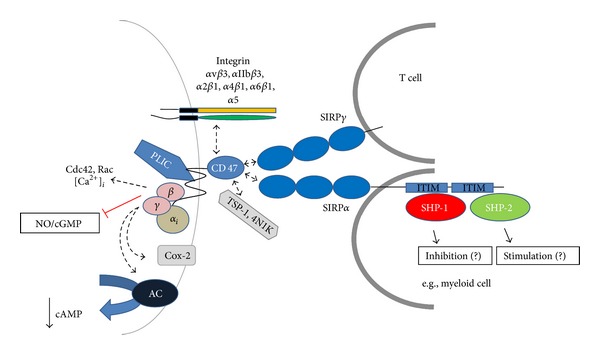
Interactions of CD47 in cis and trans. CD47 interacts in cis with integrins, in trans with SIRPs, and can also bind the soluble protein TSP-1. The figure summarizes intracellular signaling events associated with CD47 upon binding to its interaction partners.
2.2. Interaction with Thrombospondin
Thrombospondin-1 (TSP-1) is the prototypic member of the thrombospondin family of extracellular matrix glycoproteins, which are implicated in regulating cell motility, proliferation, and differentiation [23]. The extracellular IgV domain of CD47 was found to be a receptor for the C-terminal cell-binding domain (CBD) of TSP-1, since the expression of CD47 in otherwise CD47-deficient cells promotes adhesion to TSP-1 or its CBD, and a functional blocking mAb against CD47 can block endothelial cell chemotaxis against TSP-1 or the CD47 binding CBD-peptide 4N1K [24]. It was later shown that TSP-1, its CBD, or the 4N1K peptide stimulates α v β 3 integrin-mediated cell spreading on vitronectin in a CD47-dependent manner [24]. In platelets, TSP-1, the TSP-1 CBD, or 4N1K activates the platelet αIIb β 3 integrin and induces platelet spreading on fibrinogen, platelet aggregation, and increased focal adhesion kinase (FAK) tyrosine phosphorylation, which are all dependent on interaction between CD47 and integrin αIIb β 3 [16]. Furthermore, CD47 was found to mediate a synergistic effect of soluble type I collagen and TSP-1 or 4N1K, which enhance α 2 β 1 integrin-mediated platelet activation or vascular smooth muscle cell chemotaxis [17, 18]. Early experiments also suggested that CD47 regulates TSP-1-induced cell spreading or platelet activation by affecting signal transduction in a pertussis toxin-sensitive way via a heterotrimeric Gi protein [16, 24]. CD47 was later shown to functionally associate with heterotrimeric Gi, to suppress cAMP levels, and mediate the inhibition of ERK in platelets and smooth muscle cells [25]. More recently, CD47 ligated by TSP-1 was found to inhibit nitric oxide (NO) signaling in vascular cells [26] and to oppose NO/cGMP-mediated inhibition of integrin activation to facilitate platelet aggregation [27].
In addition to signaling through Gi proteins, CD47 has been shown to signal via the β 3 integrin cytoplasmic tail [28]. Although it is not entirely clear how much these two signaling pathways overlap, accumulation of CD47/β 3-integrin-complexes in cholesterol-rich lipid rafts, which appears to depend on both the CD47 IgV domain, the multiple transmembrane-spanning domain, and a long range disulfide bond between Cys33 in the IgV domain and Cys263 in the transmembrane domain [29], engage in Gi signaling [30]. CD47 has also been shown to be involved in the regulation of intracellular Ca2+ ([Ca2+]i), exemplified by its regulation of an integrin-dependent increase in [Ca2+]i in endothelial cells [31] and tumor cells [32], and that CD47 synergizes with T cell receptor-stimulated elevations of [Ca2+]i in T lymphocytes [33, 34]. The cytoplasmic tails of the form 2 and form 4 splice variants of CD47 were found to bind to the cytosolic ubiquitin-related proteins PLIC-1 and PLIC-2 (PLIC, proteins linking IAP to cytoskeleton) [35]. In B lymphocytes, CD47 was suggested to synergistically promote cell migration trough Cdc42 [20]. A role of Cdc42 and Rac has also been suggested in CD47-dependent regulation of neuronal development and neurite formation [36, 37]. Adhesion of intestinal epithelial cells to collagen I induces the association of CD47 with α 2 integrins, and CD47 is necessary for collagen I-induced cyclooxygenase-2 (Cox-2) expression and epithelial cell migration, which is mediated by Gα i3 [38] (Figure 1).
2.3. Interaction with SIRP Proteins
SIRP proteins belong to the Ig family of cell surface glycoproteins, where the first member identified was SIRPα (also known as SHPS-1, CD172a, BIT, MFR, or P84) [39–44]. SIRPα is highly expressed in myeloid cells and neurons, but also in endothelial cells and fibroblasts, and has three extracellular Ig-like domains, one distal IgV-like domain, and two membrane proximal IgC-like domains [41, 42]. In addition, an alternatively spliced form having only one IgV domain has also been reported [45]. In its intracellular tail, SIRPα has two immunoreceptor tyrosine-based inhibitory motifs (ITIMs), which when tyrosine phosphorylated can bind the Src homology 2 (SH2) domain-containing protein-tyrosine phosphatases SHP-1 and SHP-2 [42]. Additional cytoplasmic binding partners for SIRPα are the adaptor molecules Src kinase-associated protein of 55 kDa homolog/SKAP2 (SKAP55hom/R), Fyn-binding protein/SLP-76-associated phosphoprotein of 130 kDa (FYB/SLAP-130), and the tyrosine kinase PYK2 [46]. SIRPα is also a substrate for the kinase activity of the insulin, EGF, and bPDGF receptors, and the overexpression of SIRPα in fibroblasts decreases proliferation and other downstream events in response to insulin, EGF, and bPDGF [42]. Since SIRPα is also constitutively associated with the M-CSF receptor c-fms, SIRPα overexpression partially reverses the v-fms phenotype [42]. Two other family members have also been identified, SIRPβ (also known as CD172b) [42, 47] and SIRPγ (also known as CD172g or SIRPβ2) [48], whose extracellular Ig-like domains are similar to that of SIRPα. However, the cytoplasmatic regions of SIRPβ and SIRPγ are different from that of SIRPα. SIRPβ has a very short cytoplasmatic tail with no signaling motifs. Instead, the transmembrane region contains a positively charged lysine residue, which can bind the immunoreceptor-tyrosine-based-activating-motif- (ITAM-) carrying adaptor protein DNAX activation protein 12 (DAP12/KARAP) [49, 50]. SIRPγ has no recognizable signaling motif or capability to interact with cytoplasmic signaling molecules and is therefore unlikely to generate intracellular signals [51]. CD47 has been shown to be a ligand for SIRPα [52, 53] and SIRPγ [54, 55], but does not bind SIRPβ [47]. The CD47/SIRPα interaction regulates not only a multitude of intercellular interactions in many body systems, such as the immune system where it regulates lymphocyte homeostasis [56, 57], dendritic cell (DC) maturation and activation [58], proper localization of certain DC subsets in secondary lymphoid organs [59–61], and cellular transmigration [62, 63], but also regulates cells of the nervous system (reviewed in [64, 65]). An interaction between these two proteins also plays an important role in bone remodeling [66, 67]. Cellular responses regulated by the CD47/SIRPα interaction are many times dependent on a bidirectional signaling through both receptors [51, 64, 65] (Figure 1). The finding that CD47 on host cells can function as a “marker of self” and regulate phagocytosis by binding to SIRPα [68] will be further described in a subsequent section. The interaction between CD47 and SIRPα has proven to be very specific species, as shown by the relatively weak binding of CD47 from mouse, rat, or cow to human SIRPα [69, 70]. In addition, the glycosylation of CD47 or SIRPα does not seem to be necessary for their interaction [70], but the level of N-glycosylation of SIRPα has, however, an impact on the interaction such that over glycosylation reduces the binding of CD47 [71]. The long range disulfide bond between Cys33 in the CD47 IgV domain and Cys263 in the transmembrane domain is also important to establish an orientation of the CD47 IgV domain that enhances its binding to SIRPα [29].
3. CD47-Induced Apoptosis
Ligation of CD47 by anti-CD47 mAbs was found to induce apoptosis in a number of different cell types. This phenomenon was first described in Jurkat T cells, in anti-CD3 activated but not in resting primary T cells, and in B-cell chronic lymphocytic leukemia (B-CLL) cells [72, 73]. CD47-induced apoptosis can be induced by several different mAbs; however, while some of these (e.g., Ad22, 1F7, or MABL) show potent apoptosis induction in suspension [73–76], others (e.g., B6H12 and 2D3) need to be immobilized to a surface to promote cell death [72, 77]. Of the two SIRP-family members known to bind the CD47 IgV domain (SIRPα and SIRPγ), SIRPα as a soluble Fc-fusion protein does not induce CD47-dependent apoptosis [78], while SIRPα or SIRPγ bound onto the surface of beads induces apoptosis through CD47 in Jurkat T cells and the myelomonocytic cell line U937 [54]. In addition, TSP-1 or the CD47-binding TSP-1 CBD-peptide 4N1K also induces CD47-dependent apoptosis [74, 75, 78, 79]. Indeed, mice deficient in CD47 or TSP-1 sustain oxazolone-induced inflammation significantly longer than wild-type mice due to a deficiency in T cell apoptosis [79]. This form of cell death was initially described to be characterized by cell shrinkage, reduction in mitochondrial transmembrane potential, and exposure of phosphatidylserine (PS) on the cell surface, but to be independent of Fas (CD95) or TNF receptor signaling. Neither does CD47-induced cell death include DNA fragmentation nor other nuclear features associated with “classical” apoptosis, and it is independent of cysteinyl aspartate protease (caspase) activation [72, 73] (Figure 2). Furthermore, inhibitors of actin polymerization or mitochondrial electron transfer prevent CD47-induced PS exposure [77]. In support of a role for the actin cytoskeleton, peripheral blood mononuclear cells (PBMCs) from Wiskott-Aldrich syndrome (WAS) patients, where mutations in the WAS protein (WASP) results in defective Cdc42-induced regulation of the actin cytoskeleton [80], are resistant to CD47-induced apoptosis [77]. Although the mitochondrial transmembrane potential is affected in CD47-induced apoptosis, it does not involve the mitochondrial release of cytochrome c or apoptosis-inducing factor (AIF), but does involve the production of reactive oxygen species (ROS) [81]. In Jurkat T cells, it was shown that the inhibition of Giα signaling with pertussis toxin can counteract CD47-induced apoptosis, that ligation of CD47 reduces intracellular cAMP levels, and that cAMP elevating agents prevents apoptosis by CD47 ligands [74]. This signaling pathway, which likely also involves reduced signaling through protein kinase A (PKA), is not only described in T cells, but also in several breast cancer cell lines [75]. In the latter cell type, it was shown that epidermal growth factor can inhibit the CD47 death pathway via protein kinase C (PKC) [75]. A yeast two-hybrid screen, where CD47 was used as bait, identified the pro-apoptotic Bcl-2 family member Bcl-2-homology-3- (BH3-) only protein 19 kDa interacting protein-3 (BNIP3) [78]. In T cells, BNIP3 was found to physically associate with CD47, which prevents its degradation in proteasomes and sensitizes T cells to CD47-induced apoptosis [78, 79]. Ligation of CD47 induces the translocation of BNIP3 to mitochondria, and attenuation of BNIP3 activity inhibits CD47-induced apoptosis [78], which together suggests that BNIP3 is crucial as a mediator of this cell death pathway. Moreover, BNIP3 gene expression was found to be increased and regulated by hypoxia-inducible factor-1α (HIF-1α) following the ligation of CD47 by single-chain fragments of an anti-CD47 mAb which kills B-CLL cell lines both in vitro and in vivo, where the knockdown of HIF-1α represses CD47-induced cell death [82] (Figure 2). The finding that a Jurkat T cell clone lacking CD47 is resistant to Fas- (CD95-) induced apoptosis, but that expression of CD47 restores the sensitivity to Fas-ligation, suggested that CD47 can augment Fas-induced apoptosis via a mechanism that requires neither CD47 signaling nor its association with lipid rafts [83]. In fact, the lack of CD47 impairs important proapoptotic events downstream of Fas, such as loss of mitochondrial membrane potential, cytochrome c release, caspase activation, poly(ADP-ribose) polymerase (PARP) cleavage, and DNA fragmentation [83]. This function of CD47 is likely also important in primary cells, since T cells from CD47-deficient mice are protected from Fas-induced apoptosis [83].
Figure 2.
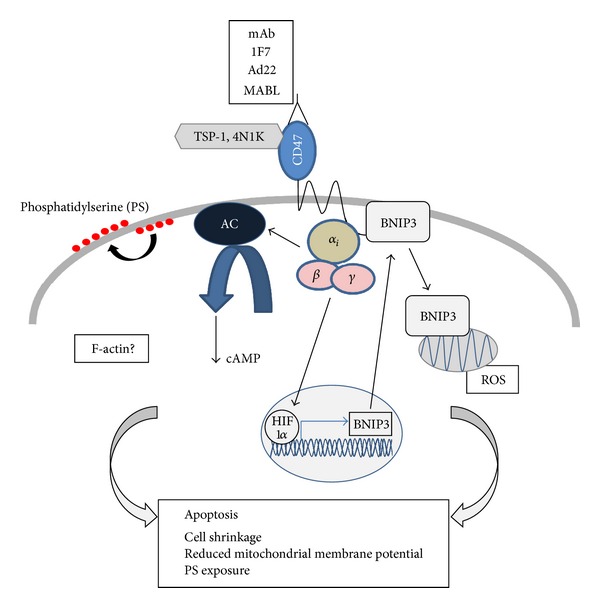
Mechanisms involved in mediating CD47-induced cell death. Ligation of CD47 by mAbs or TSP 1 induces apoptosis, which involves the activation of heterotrimeric Gi proteins, reduction in cAMP, and exposure of phosphatidylserine in the outer leaflet of the plasma membrane. It also induces the expression of the proapoptotic Bcl-2 family member BNIP3, its association with CD47, and subsequent translocation of BNIP3 to mitochondria. CD47-induced apoptosis may also involve ROS and F-actin.
In hematopoietic cells, CD47-induced apoptosis has been described in hematopoietic tumor cells [54, 72–74, 76–78, 81, 82, 84, 85] and in activated primary T or B cells [73, 74, 77, 79]. However, whether apoptosis can be induced through CD47 in nonactivated leukocytes is still somewhat unclear. The only situation where nonactivated T or B cells have been found to undergo CD47-induced apoptosis is when immobilized anti-CD47 mAb has been used [77], but not when using soluble CD47 ligands or mAbs known to induce apoptosis in activated cells or tumor cells [73, 74, 79]. Surprisingly, although CD34+ hematopoietic progenitors express CD47, they are resistant to CD47-induced apoptosis by either immobilized or soluble mAb [76, 77]. In addition, immature human monocyte-derived dendritic cells (iDCs) were described as resistant to CD47-induced apoptosis, following incubation with an immobilized CD47 mAb for 18 hours [77]. However, another study showed that freshly isolated human monocytes or human monocyte-derived iDC undergoes a rapid (within 60 min) cell death in response to the CD47-ligand 4N1K [86]. This cell death, which was described to occur in a subset of cells and where monocytes or iDC not affected by 4N1K remain viable in culture, is associated with cellular features previously described for CD47-induced apoptosis, such as not only PS exposure, increased plasma membrane permeability, reduced mitochondrial membrane potential, caspase independence, but also included DNA fragmentation [86]. Thus, these findings suggest that although a subset of iDC may undergo CD47-induced cell death at an early time point, this may not be detectable at later time points. However, it raises the question if specific subsets of monocytes or iDCs are sensitive to this form of rapid cell death whereas others are resistant and maintain their viability in culture. In addition to hematopoietic cells, overexpression of CD47 can induces cell death of cultured cerebral cortical neurons, which is enhanced by the coexpression of SIRPα and prevented by brain-derived neurotrophic factor (BDNF) when CD47 and SIRPα are coexpressed [87]. Apoptosis in neurons overexpressing CD47, however, is dependent on caspases and apoptotic cells have condensed apoptotic nuclei with fragmented DNA [87]. It has also been shown that endothelial cells incubated under static conditions in the absence of flow increase their expression of TSP-1, which uses the CD47/α v β 3 integrin as a receptor to trigger endothelial cell apoptosis [88]. This mechanism also appears to be involved in endothelial cell apoptosis during proatherogenic turbulent flow conditions [89] and in mechanosensitive induction of apoptosis in fibroblasts [90]. TSP-1-mediated apoptosis, mapped to the type-3 repeat/c-terminal domain of TSP-1, in promyelocytic leukemia cells (NB4-LR1) has also been suggested to depend on the engagement of both CD47 and the α v β 3 integrin [91]. In contrast to the proapoptotic effects of CD47 described above, it was reported that the TSP-1-derived peptide 4N1 could abolish C2-ceramide-induced apoptosis in primary porcine thyroid cells by preventing reduction in intracellular cAMP levels, an effect blocked by the functional blocking antihuman CD47 mAb B6H12 [92]. A similar effect of 4N1 peptide was also found to inhibit the cytotoxic effects of the anticancer drugs camptothecin and doxorubicin in thyroid carcinoma cells [93]. Although it is unclear how these effects of CD47-ligation can be explained in relation to the proapoptotic effects of this molecule, it is interesting to note that tumor cell TSP-1 overexpression has been linked to disease recurrence and decreased survival [94–96], and it was suggested that this pathway could be one explanation behind drug resistance in thyroid cancers [93].
4. Expression of CD47 in Erythrocytes
4.1. Interaction of CD47 with Erythrocyte Membrane Protein Complexes
Mature erythrocytes express high levels of CD47, but do not express integrins, which early indicated that other important functions of CD47 could be expected in these cells. The fact that individuals with the Rhnull phenotype, which do not express any of the proteins of the Rh protein complex, only express about 25% of normal levels of CD47 suggested a close relation between CD47 and erythrocyte Rh proteins [5, 97]. In the erythrocyte cell membrane, Rh polypeptides associate in a complex with many other proteins (e.g., Rh associated glycoprotein (RhAG), glycophorin B, LW, and CD47) [98]. Another erythrocyte membrane protein complex is formed by the band 3 anion exchanger and several other proteins (e.g., glycophorin A, protein 4.2, and ankyrin). The latter multiprotein complex mediates the anchorage of the erythrocyte membrane to the spectrin cytoskeleton [99, 100]. In addition, it has been suggested that the Rh complex and the band 3 complex may in fact be associated in the erythrocyte membrane [101]. Mutations in band 3, or complete band 3 deficiency, in human erythrocytes results in reduced expression of Rh polypeptides and RhAG [102] and results in a virtual lack of CD47 [101]. Moreover, human erythrocytes deficient in protein 4.2 also show a marked deficiency of CD47 as well as an altered glycosylation of RhAG [103]. When combining these finding, a hypothesis was put forward, which suggests that CD47 of the Rh complex may indeed form a link to the band 3 complex by binding to protein 4.2 [101] (Figure 3). Studies of protein mobility within the erythrocyte membrane have shown that CD47 is associated with the erythrocyte cytoskeleton as being a part of the Rh complex, but that CD47 is also present as a noncytoskeleton anchored pool which is more mobile in the erythrocyte membrane [104]. Thus, CD47 in erythrocytes may serve several different functions depending on its grade of mobility in the plasma membrane. It has been suggested that the freely mobile CD47 pool may be of importance to accumulate CD47 in specific membrane areas upon cell-cell contacts, for example, in order to efficiently interact with SIRPα on other cell types such as phagocytic cells [104]. Importantly, data from studies of murine erythrocytes have shown remarkable differences as compared with human erythrocytes, and the link between CD47, the Rh complex, and protein 4.2 in murine erythrocytes is not well understood. First, there are little or no Rh polypeptides in erythrocytes from band 3-deficient mice, while the expression of CD47 is only slightly reduced [101]. Second, erythrocytes from CD47-deficient mice were found to contain normal amounts of murine Rh and RhAG polypeptides [105]. Third, erythrocytes from protein 4.2-deficient mice have normal amounts of CD47 [106]. Taken together, the total picture suggests that CD47 seems to interact with Rh polypeptides and protein 4.2 in human erythrocytes, whereas the interaction between CD47, Rh polypeptides, and proteins of the band 3 complex in murine erythrocytes is still unclear.
Figure 3.
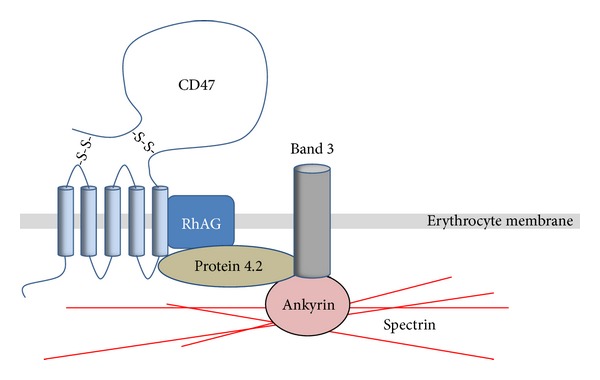
Association of CD47 with the Rh and band 3 complexes in the erythrocyte membrane. CD47 interacts with the Rh/RhAG complex and also associates with protein 4.2, which links CD47 to the band 3/ankyrin complex and the spectrin cytoskeleton. It is important to note that not all CD47 appears to be associated with this multiprotein complex in erythrocyte membranes.
4.2. CD47 in Stored or Aged Erythrocytes
A progressive decrease in the CD47 expression level has been observed on human erythrocytes stored under blood bank conditions at +4°C for more than 14 days [107–109] and in a mouse model of erythrocyte storage which tried to mimic human blood bank conditions [110]. However, the magnitude of reduction in CD47 expression was rather different, from a modest reduction of up to 6% at day 42 of storage [107], around 30% reduction on day 28 of storage [108], to a more than 50% reduction on day 14 of storage [109]. Furthermore, a recent study indicated that the CD47 levels of erythrocytes stored for 35 days were not different from that of fresh cells [111]. The discrepancy between these findings may have several explanations, such as the exact storage conditions or methods for the quantification of erythrocyte CD47 expression levels. In addition, the loss of CD47 may also be sensitive to leukocytes remaining in the erythrocyte concentrates. Indeed, a gradual loss of CD47 from erythrocytes during storage was observed in stored erythrocytes irrespective of whether the buffy coat was removed or not before storage, although buffy coat removal resulted in an increased expression level of CD47 at all time points tested, showing a significant correlation between the number of remaining leukocytes and erythrocyte CD47 levels [108]. A study of cryopreserved leukoreduced erythrocytes was unable to detect any effects on erythrocyte CD47 expression levels [112]. Microparticles released in the blood from blood cells or endothelial cells have recently gained interest due to their possible role in regulating a variety of normal or pathological biological functions [113]. Such microparticles are also released from erythrocytes during cryopreservation or storage at +4°C, and these microparticles do among other erythrocyte membrane proteins also carry CD47 [112, 114]. Although the presence of CD47 on these microparticles suggests a mechanism for the observed loss of CD47 from stored erythrocytes, soluble CD47 has also been detected in the supernatants of erythrocytes stored at +4°C [107]. In addition to a possible loss of CD47 on stored erythrocytes, a recent study also indicated that storage as well as experimental aging in vitro results in a conformational change of the CD47 protein [111]. This modification can be detected as a selectively increased binding of the anti-CD47 mAb 2D3, which recognizes an epitope different from the epitope involved in the binding of TSP-1 or SIRPα and recognized by mAb B6H12 [111]. Normal erythrocyte aging in circulation is rather complicated to study in humans. However, the use of mouse models has been informative to further understand if the level of CD47 is changing during erythrocyte aging. In these models, biotin is injected intravenously to label all blood cells at one or two specific time points [115, 116]. Analyses of circulating erythrocytes at later time points will then allow for discrimination between older biotin-positive and younger biotin-negative circulating erythrocytes [115, 116]. Using this approach, it has been found that CD47 is gradually lost from the surface of circulating murine erythrocytes, where the oldest erythrocytes may have up to 30% lower CD47 expression levels as compared with younger erythrocytes [115, 117].
4.3. CD47 in Sickle Cell or Gaucher Disease
Sickle cell disease, caused by a mutation in the hemoglobin β chain and a formation of insoluble intracellular aggregates of mutated hemoglobin, resulting in characteristic sickle shaped erythrocytes, is associated with severe vasoocclusive crisis [118, 119]. One important mechanism behind this pathology is the enhanced adhesion of sickle erythrocytes to vascular endothelium [118, 119]. CD47 was shown to play a role in the pathologic sequestration of sickle erythrocytes to vascular endothelium under shear stress by binding to TSP [120]. It has been found that the reticulocyte-enriched fractions of sickle erythrocytes are most efficiently binding to endothelial cells, that this function requires heterotrimeric G proteins and tyrosine kinase activity [121], and is mediated by α 4 β 1 integrins on the reticulocytes [19]. Interestingly, sickle erythrocytes also show an increased expression of the CD47 epitope recognized by mAb 2D3 [120]. In Gaucher's disease, a sphingolipidosis caused by glucocerebrosidase deficiency, macrophages accumulate glucosylceramide following the excess phagocytosis of erythrocytes, which converts splenic macrophages to pathogenic Gaucher cells [122]. Erythrocytes from patients with untreated Gaucher's disease have been found to have reduced levels of CD47, which can be reversed upon enzyme-replacement therapy [123]. It has been suggested that the anemia associated with the untreated disease can in part be explained by a combination of reduced CD47 levels together with other morphological erythrocyte abnormalities observed in this disease [123].
5. Self versus Non-Self Discrimination in the Innate Immune System and Regulation of Phagocytosis by the CD47/SIRPα Interaction
5.1. Self versus Non-Self Discrimination
The distinction between self and non-self is central to the maintenance of integrity in a multicellular organism and allows for a powerful and successful elimination of potentially dangerous pathogens, while carefully preserving healthy host cells and tissues. This distinction has been well studied in the adaptive immune response, which is specialized in the recognition of foreign peptides by virtue of small modifications to major histocompatibility complex (MHC) molecules made by these peptides [124]. For the innate immune system, recognition is thought to be based on a large extent on the recognition of specific microbial structures, pathogen-associated molecular patterns (PAMPs) [125]. The innate immune system of the host organism has through the evolution developed a group of receptors, pattern recognition receptors (PRRs), which serves to specifically recognize PAMPs [125]. In this way, the host may utilize a certain number of receptors, encoded in the genome, for the recognition of various evolutionary stable molecular pathogen-associated structures. While some of the PRRs (like the mannose receptor (MR)) recognize PAMPs directly [125], others (like complement receptors (CRs)) are specialized to detect products generated secondary to PAMP recognition [126]. However, a system based on specific recognition of “foreign” is clearly flawed as it prevents the recognition of anything that is also present on the organism's own cells. This can be very elegantly circumvented by a defense system that has very broad recognition, when it is combined with specific molecules that mark host cells and tissues as “self.” Thus, recognition of “self” will inhibit the activation of innate immune cells, whereas the recognition of “missing self” allows for an immune response to proceed. In other words, a system where the own cells express a unique marker of self, not present on foreign cells, would make the distinction between self and foreign very simple. In that way, it would not matter if the organism's own cells express substances or ligands that are similar to those found on foreign cells/particles. Innate immune cells such as macrophages would only look for the presence of the “self marker” on the recognized particle, where the presence of self would release the recognized particle but the absence of self would allow for activation and destruction. This would significantly simplify the recognition process, as there would only be a need for a few broad specific recognition receptors instead of a countless number of foreign-specific receptors. It would also fulfill the criteria for a defense system that effectively recognize and destroy foreign objects but at the same time reduces the risk of damaging the organism's own structures. Such a system for macrophage activation would be analogous to the well-established “missing-self hypothesis” for natural killer (NK) cell activation [127]. NK cells recognize target cells by a range of activating receptors which also recognize ligands on many normal cells. However, in the NK cell system expression of self-MHC class I will protect the recognized cell via the ligation of NK cell inhibitory receptors specific for “self-” MHC class I [128]. It is, however, important to note, that NK cell inhibitory receptors are now described, which seem to have ligands other than MHC class I [129]. Upon the binding of self-MHC class I, ligated NK cell inhibitory receptors will recruit and activate the phosphatases SHP-1/SHP-2 that mediate the inhibition of NK cell activation [129]. NK cells thus spare cells whenever they express “markers of normal self,” and eliminate them when these markers are absent or inadequately expressed. Other leukocytes also express molecules related to NK cell inhibitory receptors, further suggesting that similar mechanisms are operative also, for example, in macrophage activation [130]. Many of these inhibitory receptors recognize MHC class I [130], but the “marker of self” could in principle be any ubiquitously expressed surface molecule.
5.2. CD47 as a Marker of “Self”
A new chapter in the understanding of CD47 and its functions started when CD47 was found to bind SIRPα [52, 53]. At the time, SIRPα was regarded as one of several immunoreceptors with cytoplasmic ITIM motifs, generally suggested to be involved in negative regulation of cellular functions, mediated by the recruitment of the tyrosine phosphatases SHP-1 and/or SHP-2 or the inositol phosphatase SHIP to the tyrosine phosphorylated ITIMs [131]. In addition, SIRPα was found to be highly expressed in primary macrophages and macrophage cell lines [45], as well as in other myeloid cells such as monocytes, granulocytes, and dendritic cells [53]. In macrophages, SIRPα was found to negatively regulate signaling through tyrosine kinase-dependent signaling pathways (e.g., FcεRI) [132]. Although both SHP-1 and SHP-2 can bind to the phosphorylated SIRPα ITIMs, only SHP-1 has been associated with the inhibitory function of SIRPα in macrophages, similar to that of the NK cell inhibitory receptors [128], whereas SHP-2 associated with SIRPα leads to a phosphatase-dependent enhancement of the signal in many situations [133]. When studying CD47-deficient mice, we were struck by the fact that CD47-deficient bone marrow cannot reconstitute and engraft in lethally irradiated syngeneic wild-type recipient mice, while it engrafts normally in CD47-deficient recipient mice [134]. Our work on trying to understand this controversy first involved experiments where different leukocyte populations were transferred from CD47-deficient mice into wild-type recipient mice. However, we later simplified the system by studying transfusion of erythrocytes instead of leukocytes. The reason for this is that erythrocytes in contrast to many leukocyte populations have a long half-life in circulation, they do not divide, do not express MHC class I, and they do not home to extravascular tissues or organs. These studies showed that fresh erythrocytes isolated from the blood of CD47-deficient mice, labeled with a fluorescent cell tracker dye and transfused into wild-type recipient mice, have markedly reduced survival, whereas their half-life is normal in CD47-deficient recipients [68]. The clearance rate of such freshly isolated CD47-deficient erythrocytes is remarkably fast with complete clearance within 24 hours [68]. To put this in perspective, the average lifespan of murine erythrocytes in circulation is somewhere between 45 and 60 days, depending on the mouse strain investigated [135, 136]. The rapid clearance of CD47-deficient erythrocytes from the circulation of wild-type recipient mice does not require complement, since CD47-deficient erythrocytes are also cleared from the circulation of complement factor 3- (C3-) deficient mice. Neither is there a requirement for lymphocytes or antibodies to enable the clearance of CD47-deficient erythrocytes from the circulation, since clearance is normal in Rag1-deficient mice, which lack mature T and B lymphocytes [68]. Rather, the transfused CD47-deficient erythrocytes that are eliminated from the circulation of wild-type mice are recognized and cleared by splenic red pulp macrophages, and the removal of these macrophages by splenectomy or by treatment with macrophage-toxic clodronate liposomes [137] abrogates the elimination of CD47-deficient erythrocytes [68]. Furthermore, macrophage SIRPα is tyrosine phosphorylated upon contact with CD47 on erythrocytes, and when SIRPα on isolated splenic macrophages is blocked, it increases the level of phagocytosis of wild-type erythrocytes to that seen with CD47-deficient erythrocytes, whereas phagocytosis of the CD47-deficient erythrocytes is unaffected by the antibody treatment [68]. In addition, studies in SIRPα-mutant mice, where the cytoplasmic signaling domain of the receptor is deleted, have shown a shorter half-life of normal CD47-expressing erythrocytes in these mice, which also present with mild anemia [138]. Such spontaneous anemia is, however, not seen in CD47-deficient mice, suggesting that CD47 could also be needed on the macrophages to facilitate the clearance of erythrocytes in the spleen. However, lack of CD47 on platelets also results in very rapid clearance when transfused into wild-type recipients, and both CD47-deficient mice and SIRPα-mutant mice have a mild spontaneous antibody-independent thrombocytopenia [139, 140]. Altogether, these findings indicated that all erythrocytes can be phagocytosed by splenic red pulp macrophages when SIRPα is blocked or CD47 is missing, and that these macrophages must have a receptor for erythrocytes. Indeed, we have identified LDL receptor-related protein (LRP-1), which by recognizing calreticulin on the surface of normal erythrocytes can mediate phagocytosis of untreated CD47-deficient erythrocytes [141, 142]. By showing that macrophages in the splenic red pulp can recognize normal circulating erythrocytes, but that these macrophages do not phagocytose erythrocytes as long as they display CD47 on their surface, these findings were the first to prove that macrophages are perfectly capable of recognizing “normal” host cells and rely on “self” recognition for proper function. In addition, it also demonstrated major similarities between mechanisms for “self” recognition in NK cells and macrophages [128]. Based on the findings that unopsonized erythrocytes lacking CD47 can be phagocytosed by splenic macrophages, and that the CD47/SIRPα interaction can potently inhibit the prophagocytic mechanism operating in that situation, we next hypothesized that the CD47/SIRPα interaction might as well be able to negatively regulate phagocytosis of opsonized erythrocytes, and that for the cells of the innate immune systems front line defense (i.e., macrophages and DC) there would be a balance between signals from activating receptors (e.g., FcγR or CR) and the inhibitory signal from SIRPα ligated by target cell CD47. Indeed, the CD47/SIRPα interaction can also regulate phagocytosis of IgG-opsonized or complement opsonized erythrocytes as well as other opsonized host cells, making CD47-deficient cells severely sensitive to being phagocytosed [139, 143–145]. In addition, CD47-deficient mice are severely sensitive to experimental autoimmune hemolytic anemia (AIHA) and experimental thrombocytopenia [139, 146]. On an autoimmune background prone to develop spontaneous AIHA, lack of CD47 results in a more rapid and lethal AIHA [146]. Using motheathen viable (mev/mev) mice, which only have about 20% of normal levels of SHP-1, but normal levels of SHP-2 [147–149], we could pinpoint the role of these phosphatases in mediating the phagocytosis inhibitory effect of the CD47/SIRPα interaction in vivo. When transfused into mev/mev mice, IgG-opsonized wild-type erythrocytes are cleared with the same rapid kinetics as seen with equally opsonized CD47-deficient erythrocytes [145], showing that at that particular signaling strength through prophagocytic Fcγ receptors, the level of CD47 on normal erythrocytes was not enough to prevent phagocytosis if the SHP-1 level was reduced by about 80%. In contrast, the prophagocytic signaling mediating phagocytosis of unopsonized CD47-deficient erythrocytes (presumably mediated by LRP-1) is likely much weaker, since unopsonized wild-type erythrocytes are not cleared with the same rapid kinetics as CD47-deficient cells in mev/mev mice [145]. However, while unopsonized CD47 heterozygous erythrocytes (expressing about 50% of the CD47 found on wild-type erythrocytes) show normal half-life when transfused into wild-type recipients, they were found to be cleared more rapidly when transfused into mev/mev mice [145]. These findings thus proposed that a reduction of the CD47 level to 50% of normal, in combination with a strongly reduced level of SHP-1 in the macrophages, together weakened the inhibitory signaling through SIRPα such that clearance of unopsonized erythrocytes was allowed. Furthermore, the rate of phagocytosis of IgG-opsonized erythrocytes is distinctly regulated by the amount of CD47 present on the surface of the erythrocytes both in vivo and in vitro [150]. Thus, activation of phagocytosis in a macrophage in contact with a target erythrocyte (or any other host cell) can be viewed as a balance between signals from activating receptors (i.e., FcγR, CR, or LRP-1) and the inhibitory signal from SIRPα ligated by target cell CD47. In the macrophage, neither signal appears to be dominant, but rather the decision to phagocytose a target host cell is based on an integration of positive prophagocytic signals and inhibitory CD47/SIRPα signaling (Figure 4(a)). The same functional regulation also seems to be operating in DCs [134] and in microglia [151]. In hemophagocytic lymphohistiocytosis, hematopoietic stem cells are phagocytosed by bone marrow macrophages as a result of systemic inflammation [152]. In this disease, hematopoietic stem cells were found to express reduced levels of CD47 [153], which shows that pathological conditions may occur where a combination of inflammatory macrophage activation and reduced expression of CD47 results in a severe loss of critical cell types [153]. However, not all prophagocytic receptors seem to be regulated by the CD47/SIRPα interaction. Although Fcγ receptor-mediated phagocytosis of IgG-opsonized oxidatively damaged erythrocytes is strongly inhibited by the CD47/SIRPα interaction, scavenger receptor-mediated uptake of unopsonized oxidized erythrocytes turned out to be insensitive to this inhibitory mechanism [154]. The mechanism whereby recruitment of SHP-1 to SIRPα can inhibit phagocytosis has been suggested to involve the tyrosine kinase Syk and phosphoinositide 3 kinase (PI3 kinase) [144]. More recently, it was suggested that SHP-1 mediates dephosphorylation of nonmuscular myosin IIA at the phagocytic synapse between the phagocyte and a host cell, as a result of the interaction between macrophage SIRPα and CD47 on the host cell, which brings further insight into the mechanism behind phagocytosis inhibition by the CD47/SIRPα interaction [155].
Figure 4.
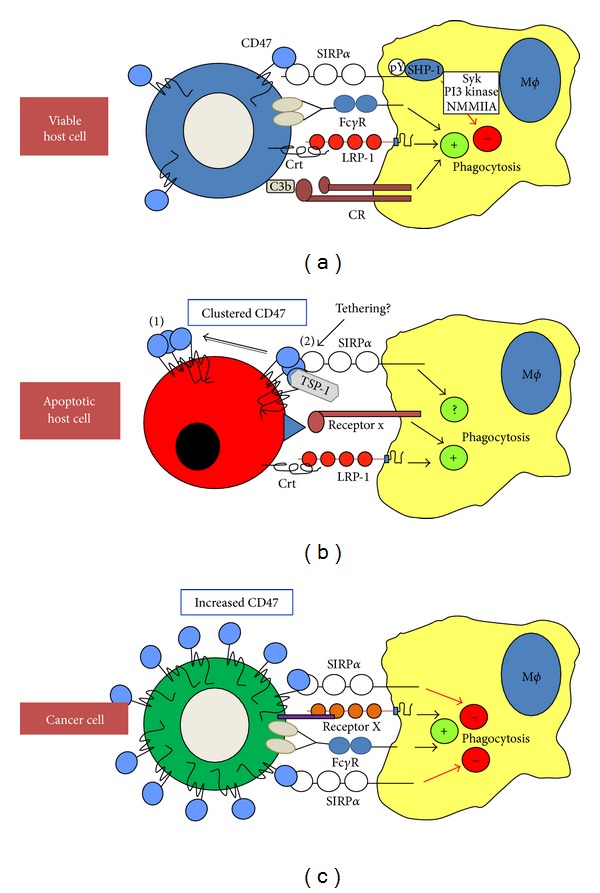
CD47 regulates phagocytosis of host cells by interacting with SIRPα. (a) CD47 on viable normal host cells can bind to SIRPα on a phagocytic cell (e.g., a macrophage (Mϕ)), which induces tyrosine phosphorylation of SIRPα ITIMS and recruitment of SHP-1. This can inhibit prophagocytic signaling through Fcγ receptors, complement receptors (CR), or LRP-1. Phagocytosis inhibition may involve signaling through Syk and PI3 kinase and inhibition of nonmuscle myosin type IIA (NMMIIA). (b) On apoptotic cells, CD47 becomes clustered in the plasma membrane and may be segregated away from domains containing ligands for prophagocytic receptors, here exemplified by calreticulin (crt) binding to LRP-1. However, most likely this principle involves also other prophagocytic receptors (receptor x). Clustered CD47 may also bind to SIRPα without inducing inhibition of phagocytosis, but may rather promote tethering of the apoptotic cell to the phagocyte. This function may also involve TSP-1 and so far uncharacterized mechanisms that can also promote phagocytosis. (c) Cancer cells may increase their expression of CD47 to strengthen the inhibitory signals through SIRPα and to more potently inhibit phagocytosis mediated by Fcγ receptors and other prophagocytic receptors (receptor x).
Based on the seemingly important interaction between CD47 and SIRPα to prevent phagocytosis of host cells, and the fact that CD47-deficient cells are rapidly phagocytosed when transfused into wild-type mice, it is clearly a puzzle why CD47-deficient mice do not present with a more severe phenotype where the macrophages phagocytose a large fraction of the CD47-deficient cells in those mice [68]. However, this phenomenon is rather similar to what is described in β2 microglobulin deficient mice, which lack expression of MHC class I but where the NK cells still do not attack and destroy host cells and also show hyporeactivity to cells carrying ligands for activating NK cell receptors, cells which are efficiently killed by wild-type NK cells [156]. One explanation proposed for this function in NK cells is that inhibitory NK cell receptors need to interact with “self-” MHC class I in order for the NK cell to become “licenced” and able to become activated and kill target cells lacking or expressing reduced levels of “self-” MHC class I [156]. By investigating the ability to phagocytose CD47-deficient cells in mice where CD47 was expressed by hematopoietic cells, nonhematopoietic cells, or both, the phagocytic “tolerance” to cells lacking CD47 in CD47-deficient mice was recently proposed to function in a similar way [157]. It was found that macrophages developing in an environment where nonhematopoietic cells lack CD47 become “tolerant” to cells lacking CD47, which allowed CD47-defcient leukocytes to avoid clearance. Curiously, this did not involve erythrocytes, which were cleared by splenic macrophages also in the bone marrow chimeras where nonhematopoietic cells lacked CD47 and macrophages had become “tolerant” [157]. The latter phenomenon has so far not been explained, but may suggest that phagocytosis of erythrocytes is regulated differently from that of leukocytes. Although CD47-deficient macrophages express normal amounts of SIRPα, which can also become tyrosine phosphorylated upon contact with CD47 expressing cells (Oldenborg et al., unpublished observations and [158]), it is possible that the “tolerance” to CD47-deficient cells that develop in CD47-deficient mice has to do with tuning of intracellular signaling pathways. In favor of such a hypothesis is the observation that transfused IgG-opsonized wild-type erythrocytes are cleared from the circulation at a significantly slower rate in CD47-deficient mice, as compared with that seen in wild-type recipient mice (Oldenborg et al. unpublished observations). This is unlikely due to a different expression of Fcγ receptors in CD47-deficient mice, since CD47-deficient IgG-opsonized erythrocytes are cleared with the same kinetics in both wild-type and CD47-deficient recipient mice (Oldenborg et al. unpublished observations).
5.3. CD47 and Phagocytosis of Apoptotic Cells
Apoptosis is a physiological process of programmed cell death which is important for embryologic development, maintenance of homeostasis, and elimination of damaged cells. An important event related to this process is the rapid uptake of apoptotic cells or apoptotic bodies by phagocytic cells. The efficacy of this process in the body is shown by the fact that apoptotic cells are apparently removed with such extremely high efficiency that apoptotic cells are very hard to detect in tissues under normal physiological conditions. It has even been suggested that if apoptotic cells are in fact detected in vivo, this may indicate a possibility of defects in their clearance or the presence of a large overload of apoptotic cells [159–162]. The largest part of apoptotic cell phagocytosis is mediated not only by professional phagocytes like macrophages and DCs, but also to some extent by nonprofessional phagocytes such as fibroblasts; epithelial cells and stromal cells are equipped with this function [160]. One example of the latter phenomenon is the phagocytosis of apoptotic mammary epithelial cells by bystander epithelial cells during mammary gland involution [163, 164]. When CD47 was identified as a cell surface protein on host cells that can negatively regulate their phagocytosis through SIRPα, one important question was to understand if this signaling pathway is altered during apoptosis to facilitate uptake of apoptotic cells. One hypothesis would be that apoptotic cells downregulate the amount of CD47 on their surface, which together with an increased amount of prophagocytic ligands exposed during apoptosis would result in phagocytosis of the dead cells. Indeed we found a reduced expression of CD47 on apoptotic fibroblasts and neutrophils [141], but curiously not on apoptotic Jurkat T cells [141] or apoptotic murine thymocytes [165]. This issue became even more confusing when apoptotic murine T cells lacking CD47, in contrast to the solid data showing that CD47 on host cells inhibits phagocytosis by macrophages, were found to be phagocytosed less efficiently by macrophages than equally apoptotic CD47 wild-type T cells [143, 165]. In apoptotic murine T cells, CD47 was rather found to be important in mediating tethering of the apoptotic cells to the phagocyte [143, 165]. In addition, CD47 is redistributed into patches on the apoptotic cell surface, areas of the plasma membrane which are different than those harbouring clusters of proteins that function as ligands for prophagocytic receptors [141, 143]. Thus, one possibility would be that the segregation of CD47 away from these phagocytosis promoting ligands could allow for tethering mediated by the CD47/SIRPα interaction, but that SIRPα would be too far away from the prophagocytic receptors to be involved in negative regulation (Figure 4(b)). Another very interesting hypothesis was recently suggested, which shed new light on this process. As described above in Section 4.2 experimentally oxidized erythrocytes, as well as erythrocytes stored in blood bank conditions, showed enhanced expression of the CD47 epitope recognized by mAb 2D3, suggesting a conformational change in the CD47 IgV domain [111]. Importantly, this is associated with an increased binding of TSP-1 to CD47, which together could generate a new binding site for SIRPα that induces a prophagocytic signal instead of the “normal” inhibitory CD47/SIRPα signal [111]. However, this very interesting hypothesis still has to also be investigated in the context of macrophage phagocytosis of apoptotic nucleated cells such as neutrophils or T cells. Thus, CD47 may operate in several different ways during cellular senescence in order to switch its normal phagocytosis-preventing role to rather facilitate and promote the phagocytosis of senescent cells. LRP-1 is a multifunctional scavenger receptor, shown to be involved in mediating uptake of apoptotic cells [141, 166, 167], and as already mentioned it can also bind to calreticulin on viable erythrocytes to induce phagocytosis if the inhibitory CD47/SIRPα interaction is not strong enough [141, 142] (Figure 4(b)). Glucocorticoids are powerful in treatment of inflammatory conditions, which can be attributed to their ability to downregulate production of and cellular responses to proinflammatory cytokines and their ability to inhibit the recruitment of inflammatory cells [168, 169]. However, another mechanism whereby glucocorticoids may also reduce inflammation is to stimulate phagocytosis of apoptotic cells [170, 171]. Interestingly, glucocorticoid treatment of macrophages results in increased macrophage expression of LRP-1 and increased phagocytosis of apoptotic cells or viable CD47-deficient erythrocytes [142].
5.4. CD47 and Regulation of Tumor Cell Phagocytosis
Since CD47 can inhibit phagocytosis of host cells, and the amount of CD47 on the cell surface is clearly important in determining the phagocytosis efficiency of macrophages, this also raised the question on whether cells such as tumor cells can also increase their CD47 expression levels in order to escape macrophage elimination. Indeed, early studies showed that ovarian carcinoma cell lines express increased levels of CD47 [6, 7], the functional significance of which was unclear at that time. More recently, an enhanced expression of CD47 has been reported for murine myeloid leukemias, as well as human normal and leukemic hematopoietic stem cells and many human solid tumors [172–174], and that increased CD47 expression is associated with reduced patient survival in AML and solid tumors [173, 174]. To challenge the hypothesis that CD47 on the leukemic cells protects from phagocytosis by macrophages and supports tumor cell growth, a xenogenic mouse model was used to study a particular human AML clone (MOLM-13) with very low endogenous CD47 expression. Expression of murine CD47 in MOLM-13 cells and transplantation of MOLM-13 clones expressing low or high levels of CD47 showed an enrichment and spreading of the CD47-high expressing clones to many bones of recipient mice, while the CD47-low clones showed minimal engraftment [172]. Depletion of macrophages in recipient mice allows for engraftment of CD47-low clones, and macrophage phagocytosis of AML cells both in vitro and in vivo is selective for tumor cells expressing low amounts of CD47 [172]. Importantly, by blocking SIRPα on macrophages, phagocytosis of CD47-high AML cells is increased to that seen with CD47-low clones [172]. Thus, these findings strongly suggest that an increased CD47 expression level on tumor cells could serve to avoid macrophage clearance and promote dissemination of tumor cells (Figure 4(c)). It was also recently shown that treatment serving to block the CD47/SIRPα interaction can result in elimination of AML stem cells in a xenogenic model [175]. Furthermore, inline with our original finding that the CD47/SIRPα pathway also potently inhibits Fcγ receptor-mediated phagocytosis of IgG-opsonized host cells [139, 145, 146, 150], it has been shown that antibody-mediated blocking of CD47/SIRPα signaling promotes phagocytosis of non-Hodgkin lymphoma cells treated with rituximab [176] and antibody-dependent cellular cytotoxicity (ADCC) against Her2/Neu-positive breast cancer cells treated with traztusumab [177]. Thus, blocking the CD47/SIRPα interaction may prove to be a powerful tool in the treatment of various tumors.
5.5. CD47 and Regulation of Xenografts
While organ transplantation is an important procedure to treat end-stage organ failure, it is hampered by a shortage of organ donors. To solve this problem, it has been suggested that pigs could serve as donors of organs, tissues, or cells to be transplanted into humans (i.e., xenotransplantation) [158, 178]. Both the innate and adaptive immune system participates in the rejection of xenogenic transplants [178], the exact details of which is outside the scope of the present paper. However, of particular interest here are the findings suggesting that even in the absence of a functional adaptive immune system and in the absence of NK cells (both of which react to cell surface determinants of xenogenic cells recognized as non-self), rejection of xenogenic cells and tissues is seen, which suggests that macrophages could play an important role in this process [158]. Indeed, in studies of transplantation of pancreatic islets [179, 180] or hematopoietic cells [181] from pigs to mice, macrophage depletion significantly enhances engraftment. The ability of macrophage SIRPα to inhibit phagocytosis by binding to CD47 on a target cell, and the fact that this interaction is rather species specific suggested that this interaction could play a role in macrophage-mediated clearance of xenogenic cells. Indeed, it was found that porcine CD47 cannot bind to murine SIRPα and induce tyrosine phosphorylation of the SIRPα ITIMs [182], which could explain the rapid clearance of porcine hematopoietic cells by macrophages both in vitro and in vivo [181, 182]. However, expression of murine CD47 in porcine cells markedly inhibits macrophage-mediated phagocytosis of these cells in vitro and prolongs their survival in vivo [182]. Although a human SIRPα fusion protein was found to bind to porcine CD47 [69], porcine CD47 does not activate human SIRPα [183]. As a result, porcine hematopoietic cells are rapidly phagocytosed by human macrophages, but expression of human CD47 in porcine cells results in attenuated phagocytosis by human macrophages [183]. These findings indicate that transgenic pigs expressing human CD47 could be an important possibility to further help reducing the rejection of porcine cells in xenotransplantation.
6. CD47 in Bone Homeostasis
The finding that CD47, by binding to SIRPα, could regulate rat alveolar macrophage fusion and formation of multinucleated giant cells in vitro [184] suggested a function also in regulating formation of bone-resorbing osteoclasts and regulation of bone homeostasis. Using functional blocking monoclonal antibodies against either CD47 or SIRPα, the formation of tartrate resistant acid phosphatase (TRAP)+ multinucleated osteoclasts is strongly inhibited in murine cell culture systems in vitro [66]. This finding can also be confirmed in vivo, since CD47-deficient mice have reduced numbers of osteoclasts in bone [66] and are protected from tumor metastasis and subsequent bone resorption [185]. There are a few hypotheses presented to explain the mechanism whereby the interaction between CD47 and SIRPα promotes osteoclastogenesis. One suggests that binding of CD47 results in SIRPα tyrosine phosphorylation and subsequent recruitment of SHP-1 to the phosphorylated ITIMs, which next would mediate dephosphorylation of nonmuscle cell myosin IIA to promote fusion and formation of osteoclasts [67]. This hypothesis is contrasted by a study of SIRPα mutant mice, expressing a truncated nonsignaling SIRPα cytoplasmic domain, where cultures of bone marrow cells generated the same number of osteoclasts of the same size, as found in wild-type bone marrow cultures [186]. The important observation made in the latter study was that osteoclasts generated in SIRPα mutant mice had an increased bone resorbing activity, suggesting that SIRPα negatively regulates this function in osteoclasts [186]. The contradictory findings presented regarding a possible role of signaling through either CD47 or SIRPα during fusion of preosteoclasts and formation of multinucleated osteoclasts must also be put in context of the originally presented hypothesis that the interaction between CD47 and SIRPα may mainly facilitate cell fusion by promoting cell-cell adhesion and bringing the plasma membranes of two cells close enough to facilitate cell fusion [187]. Here it has also been highlighted that CD47 may interact with the shorter SIRPα isoform having only one extracellular IgV domain, which would further reduce the distance between two opposing cellular membranes where cell fusion could take place [187].
7. Concluding Remarks and Future Perspectives
CD47 can regulate many important physiological cellular mechanisms by interacting with integrins, TSP-1, or SIRPα. As outlined above, much knowledge has been collected over the past two decades, but much is probably still to be discovered regarding the functions of this protein. There are several areas where one can assume that further progress may result in new novel ways to interfere with biological mechanisms in pathological conditions. One such example is the ability to induce apoptosis by ligating CD47. Although CD47 is expressed by virtually all cells in the body, in healthy as well as pathological cells, the data so far indicates that CD47-induced apoptosis may still be a way to kill, for example, tumor cells, since it appears that tumor cells and activated cells are much more sensitive to this cell death mechanism than nonactivated naïve cells or hematopoietic stem cells. The ability of CD47 to inhibit phagocytosis is clearly an important mechanism whereby innate cells as macrophages or dendritic cells can discriminate between self and non-self, and maintain tolerance to host cells. In the field of xenotransplantation, further development and research to create xenotransplants carrying human CD47 may prove important to obtain even better conditions where xenogenic cell, tissues, or organs can be tolerated when transplanted into humans. In the field of cancer research, where tumor cells may express higher levels of CD47 as a way of avoiding phagocytosis and clearance by phagocytic cells, and where CD47-blocking antibodies have been shown to promote clearance of and to reduce the dissemination of tumor cells, brings hope that careful development of reagents that can block the CD47/SIRPα interaction may indeed be useful to treat many forms of cancer without having too much of a negative side effect in terms of inducing clearance of host cells. Also in the field of bone research, one may suggest that manipulation of the interaction between CD47 and SIRPα may be a way to modulate bone resorption and to prevent osteoporosis. However, data showing that these two proteins may also be involved in regulating bone formation [67] indicate that manipulation of this signaling system in bone tissue may be more complicated.
Acknowledgments
This paper is supported by Grants from the Swedish Research Council (2010-4286), the Faculty of Medicine, Umeå University, and a Young Researcher Award from Umeå University.
References
- 1.Lindberg FP, Gresham HD, Schwarz E, Brown EJ. Molecular cloning of integrin-associated protein: an immunoglobulin family member with multiple membrane-spanning domains implicated in α V β 3-dependent ligand binding. Journal of Cell Biology. 1993;123(2):485–496. doi: 10.1083/jcb.123.2.485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Brown E, Hooper L, Ho T, Gresham H. Integrin-associated protein: a 50-kD plasma membrane antigen physically and functionally associated with integrins. Journal of Cell Biology. 1990;111(6):2785–2794. doi: 10.1083/jcb.111.6.2785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lindberg FP, Gresham HD, Reinhold MI, Brown EJ. Integrin-associated protein immunoglobulin domain is necessary for efficient vitronectin bead binding. Journal of Cell Biology. 1996;134(5):1313–1322. doi: 10.1083/jcb.134.5.1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lindberg FP, Bullard DC, Caver TE, Gresham HD, Beaudet AL, Brown EJ. Decreased resistance to bacterial infection and granulocyte defects in IAP-deficient mice. Science. 1996;274(5288):795–798. doi: 10.1126/science.274.5288.795. [DOI] [PubMed] [Google Scholar]
- 5.Lindberg FP, Lublin DM, Telen MJ, et al. Rh-related antigen CD47 is the signal-transducer integrin-associated protein. Journal of Biological Chemistry. 1994;269(3):1567–1570. [PubMed] [Google Scholar]
- 6.Campbell IG, Freemont PS, Foulkes W, Trowsdale J. An ovarian tumor marker with homology to vaccinia virus contains an IgV- like region and multiple transmembrane domains. Cancer Research. 1992;52(19):5416–5420. [PubMed] [Google Scholar]
- 7.Poels LG, Peters D, van Megen Y, et al. Monoclonal antibody against human ovarian tumor-associated antigens. Journal of the National Cancer Institute. 1986;76(5):781–791. [PubMed] [Google Scholar]
- 8.Reinhold MI, Lindberg FP, Plas D, Reynolds S, Peters MG, Brown EJ. In vivo expression of alternatively spliced forms of integrin-associated protein (CD47) Journal of Cell Science. 1995;108(11):3419–3425. doi: 10.1242/jcs.108.11.3419. [DOI] [PubMed] [Google Scholar]
- 9.Lee EHY, Hsieh YP, Yang CL, Tsai KJ, Liu CH. Induction of integrin-associated protein (IAP) mRNA expression during memory consolidation in rat hippocampus. European Journal of Neuroscience. 2000;12(3):1105–1112. doi: 10.1046/j.1460-9568.2000.00985.x. [DOI] [PubMed] [Google Scholar]
- 10.Kaur S, Kuznetsova SA, Pendrak ML, et al. Heparan sulfate modification of the transmembrane receptor CD47 is necessary for inhibition of T Cell receptor signaling by thrombospondin-1. Journal of Biological Chemistry. 2011;286(17):14991–15002. doi: 10.1074/jbc.M110.179663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gresham HD, Goodwin JL, Allen PM, Anderson DC, Brown EJ. A novel member of the integrin receptor family mediates Arg-Gly-Asp-stimulated neutrophil phagocytosis. Journal of Cell Biology. 1989;108(5):1935–1943. doi: 10.1083/jcb.108.5.1935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cooper D, Lindberg FP, Gamble JR, Brown EJ, Vadas MA. The transendothelial migration of neutrophils involves integrin-associated protein (CD47) Proceedings of the National Academy of Sciences of the United States of America. 1995;92(9):3978–3982. doi: 10.1073/pnas.92.9.3978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Parkos CA, Colgan SP, Liang TW, et al. CD47 mediates post-adhesive events required for neutrophil migration across polarized intestinal epithelia. Journal of Cell Biology. 1996;132(3):437–450. doi: 10.1083/jcb.132.3.437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liu Y, Merlin D, Burst SL, Pochet M, Madara JL, Parkos CA. The role of CD47 in neutrophil transmigration: increased rate of migration correlates with increased cell surface expression of CD47. Journal of Biological Chemistry. 2001;276(43):40156–40166. doi: 10.1074/jbc.M104138200. [DOI] [PubMed] [Google Scholar]
- 15.Senior RM, Gresham HD, Griffin GL, Brown EJ, Chung AE. Entactin stimulates neutrophil adhesion and chemotaxis through interactions between its Arg-Gly-Asp (RGD) domain and the leukocyte response integrin. Journal of Clinical Investigation. 1992;90(6):2251–2257. doi: 10.1172/JCI116111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chung J, Gao AG, Frazier WA. Thrombspondin acts via integrin-associated protein to activate the platelet integrin β llb β 3 . Journal of Biological Chemistry. 1997;272(23):14740–14746. doi: 10.1074/jbc.272.23.14740. [DOI] [PubMed] [Google Scholar]
- 17.Chung J, Wang XQ, Lindberg FP, Frazier WA. Thrombospondin-1 acts via IAP/CD47 to synergize with collagen in α2β1- mediated platelet activation. Blood. 1999;94(2):642–648. [PubMed] [Google Scholar]
- 18.Wang XQ, Frazier WA. The thrombospondin receptor CD47 modulates and associates with α2β1 integrin in vascular smooth muscle cells. Molecular Biology of the Cell. 1998;9(4):865–874. doi: 10.1091/mbc.9.4.865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Brittain JE, Han J, Ataga KI, Orringer EP, Parise LV. Mechanism of CD47-induced α4β1 integrin activation and adhesion in sickle reticulocytes. Journal of Biological Chemistry. 2004;279(41):42393–42402. doi: 10.1074/jbc.M407631200. [DOI] [PubMed] [Google Scholar]
- 20.Yoshida H, Tomiyama Y, Ishikawa J, et al. Integrin-associated protein/CD47 regulates motile activity in human B-cell lines through CDC42. Blood. 2000;96(1):234–241. [PubMed] [Google Scholar]
- 21.Koenigsknecht J, Landreth G. Microglial phagocytosis of fibrillar β-amyloid through a β1 integrin-dependent mechanism. Journal of Neuroscience. 2004;24(44):9838–9846. doi: 10.1523/JNEUROSCI.2557-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Orazizadeh M, Lee HS, Groenendijk B, et al. CD47 associates with alpha 5 integrin and regulates responses of human articular chondrocytes to mechanical stimulation in an in vitro model. Arthritis Research & Therapy. 2008;10(1, article R4) doi: 10.1186/ar2350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Adams JC, Lawler J. The thrombospondins. International Journal of Biochemistry and Cell Biology. 2004;36(6):961–968. doi: 10.1016/j.biocel.2004.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gao AG, Lindberg FP, Dimitry JM, Brown EJ, Frazier WA. Thrombospondin modulates α V β 3 function through integrin-associated protein. Journal of Cell Biology. 1996;135(2):533–544. doi: 10.1083/jcb.135.2.533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Frazier WA, Gao AG, Dimitry J, et al. The thrombospondin receptor integrin-associated protein (CD47) functionally couples to heterotrimeric Gi . Journal of Biological Chemistry. 1999;274(13):8554–8560. doi: 10.1074/jbc.274.13.8554. [DOI] [PubMed] [Google Scholar]
- 26.Isenberg JS, Hyodo F, Pappan LK, et al. Blocking thrombospondin-1/CD47 signaling alleviates deleterious effects of aging on tissue responses to ischemia. Arteriosclerosis, Thrombosis, and Vascular Biology. 2007;27(12):2582–2588. doi: 10.1161/ATVBAHA.107.155390. [DOI] [PubMed] [Google Scholar]
- 27.Isenberg JS, Romeo MJ, Yu C, et al. Thrombospondin-1 stimulates platelet aggregation by blocking the antithrombotic activity of nitric oxide/cGMP signaling. Blood. 2008;111(2):613–623. doi: 10.1182/blood-2007-06-098392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Blystone SD, Lindberg FP, LaFlamme SE, Brown EJ. Integrin β 3 cytoplasmic tail is necessary and sufficient for regulation of α 5 β 1 phagocytosis by α V β 3 and integrin-associated protein. Journal of Cell Biology. 1995;130(3):745–754. doi: 10.1083/jcb.130.3.745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Rebres RA, Vaz LE, Green JM, Brown EJ. Normal ligand binding and signaling by CD47 (Integrin-associated protein) requires a long range disulfide bond between the extracellular and membrane-spanning domains. Journal of Biological Chemistry. 2001;276(37):34607–34616. doi: 10.1074/jbc.M106107200. [DOI] [PubMed] [Google Scholar]
- 30.McDonald JF, Zheleznyak A, Frazier WA. Cholesterol-independent interactions with CD47 enhance α V β 3 avidity. Journal of Biological Chemistry. 2004;279(17):17301–17311. doi: 10.1074/jbc.M312782200. [DOI] [PubMed] [Google Scholar]
- 31.Schwartz MA, Brown EJ, Fazeli B. A 50-kDa integrin-associated protein is required for integrin-regulated calcium entry in endothelial cells. Journal of Biological Chemistry. 1993;268(27):19931–19934. [PubMed] [Google Scholar]
- 32.Shahan TA, Fawzi A, Bellon G, Monboisse JC, Kefalides NA. Regulation of tumor cell chemotaxis by type IV collagen is mediated by a Ca2+-dependent mechanism requiring CD47 and the integrin α V β 3 . Journal of Biological Chemistry. 2000;275(7):4796–4802. doi: 10.1074/jbc.275.7.4796. [DOI] [PubMed] [Google Scholar]
- 33.Rebres RA, Green JM, Reinhold MI, Ticchioni M, Brown EJ. Membrane raft association of CD47 is necessary for actin polymerization and protein kinase C θ translocation in its synergistic activation of T cells. Journal of Biological Chemistry. 2001;276(10):7672–7680. doi: 10.1074/jbc.M008858200. [DOI] [PubMed] [Google Scholar]
- 34.Waclavicek M, Majdic O, Stulnig T, et al. T cell stimulation via CD47: agonistic and antagonistic effects of CD47 monoclonal antibody 1/1A4. Journal of Immunology. 1997;159(11):5345–5354. [PubMed] [Google Scholar]
- 35.Wu AL, Wang J, Zheleznyak A, Brownt EJ. Ubiquitin-related proteins regulate interaction of vimentin intermediate filaments with the plasma membrane. Molecular Cell. 1999;4(4):619–625. doi: 10.1016/s1097-2765(00)80212-9. [DOI] [PubMed] [Google Scholar]
- 36.Miyashita M, Ohnishi H, Okazawa H, et al. Promotion of neurite and filopodium formation by CD47: roles of integrins, rac, and Cdc42. Molecular Biology of the Cell. 2004;15(8):3950–3963. doi: 10.1091/mbc.E04-01-0019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Murata T, Ohnishi H, Okazawa H, et al. CD47 promotes neuronal development through Src- and FRG/Vav2-mediated activation of Rac and Cdc42. Journal of Neuroscience. 2006;26(48):12397–12407. doi: 10.1523/JNEUROSCI.3981-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Broom OJ, Zhang Y, Oldenborg PA, Massoumi R, Sjölander A. CD47 regulates collagen I-induced cyclooxygenase-2 expression and intestinal epithelial cell migration. PLoS ONE. 2009;4(7) doi: 10.1371/journal.pone.0006371.e6371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chuang W, Lagenaur CF. Central nervous system antigen P84 can serve as a substrate for neurite outgrowth. Developmental Biology. 1990;137(2):219–232. doi: 10.1016/0012-1606(90)90249-i. [DOI] [PubMed] [Google Scholar]
- 40.Comu S, Weng W, Olinsky S, et al. The murine P84 neural adhesion molecule is SHPS-1, a member of the phosphatase-binding protein family. Journal of Neuroscience. 1997;17(22):8702–8710. doi: 10.1523/JNEUROSCI.17-22-08702.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Fujioka Y, Matozaki T, Noguchi T, et al. A novel membrane glycoprotein, SHPS-1, that binds the SH2-domain- containing protein tyrosine phosphatase SHP-2 in response to mitogens and cell adhesion. Molecular and Cellular Biology. 1996;16(12):6887–6899. doi: 10.1128/mcb.16.12.6887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kharitonenkov A, Chen Z, Sures I, Wang H, Schilling J, Ullrich A. A family of proteins that inhibit signalling through tyrosine kinase receptors. Nature. 1997;386(6621):181–186. doi: 10.1038/386181a0. [DOI] [PubMed] [Google Scholar]
- 43.Saginario C, Sterling H, Beckers C, et al. MFR, a putative receptor mediating the fusion of macrophages. Molecular and Cellular Biology. 1998;18(11):6213–6223. doi: 10.1128/mcb.18.11.6213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sano SI, Ohnishi H, Omori A, Hasegawa J, Kubota M. BIT, an immune antigen receptor-like molecule in the brain. FEBS Letters. 1997;411(2-3):327–334. doi: 10.1016/s0014-5793(97)00724-2. [DOI] [PubMed] [Google Scholar]
- 45.Veillette A, Thibaudeaut E, Latour S. High expression of inhibitory receptor SHPS-1 and its association with protein-tyrosine phosphatase SHP-1 in macrophages. Journal of Biological Chemistry. 1998;273(35):22719–22728. doi: 10.1074/jbc.273.35.22719. [DOI] [PubMed] [Google Scholar]
- 46.Timms JF, Swanson KD, Marie-Gardine A, et al. SHPS-1 is a scaffold for assembling distinct adhesion-regulated multi-protein complexes in macrophages. Current Biology. 1999;9(16):927–930. doi: 10.1016/s0960-9822(99)80401-1. [DOI] [PubMed] [Google Scholar]
- 47.Seiffert M, Brossart P, Cant C, et al. Signal-regulatory protein α (SIRPα) but not SIRPβ is involved in T-cell activation, binds to CD47 with high affinity, and is expressed on immature CD34+CD38− hematopoietic cells. Blood. 2001;97(9):2741–2749. doi: 10.1182/blood.v97.9.2741. [DOI] [PubMed] [Google Scholar]
- 48.Ichigotani Y, Matsuda S, Machida K, et al. Molecular cloning of a novel human gene (SIRP-B2) which encodes a new member of the SIRP/SHPS-1 protein family. Journal of Human Genetics. 2000;45(6):378–382. doi: 10.1007/s100380070013. [DOI] [PubMed] [Google Scholar]
- 49.Dietrich J, Cella M, Seiffert M, Bühring HJ, Colonna M. Cutting edge: signal-regulatory protein β1 is a DAP12-associated activating receptor expressed in myeloid cells. Journal of Immunology. 2000;164(1):9–12. doi: 10.4049/jimmunol.164.1.9. [DOI] [PubMed] [Google Scholar]
- 50.Tomasello E, Cant C, Buhring HJ, et al. Association of signal-regulatory proteins β with KARAP/DAP-12. European Journal of Immunology. 2000;30:2147–2156. doi: 10.1002/1521-4141(2000)30:8<2147::AID-IMMU2147>3.0.CO;2-1. [DOI] [PubMed] [Google Scholar]
- 51.Barclay AN, Brown MH. The SIRP family of receptors and immune regulation. Nature Reviews Immunology. 2006;6(6):457–464. doi: 10.1038/nri1859. [DOI] [PubMed] [Google Scholar]
- 52.Jiang P, Lagenaur CF, Narayanan V. Integrin-associated protein is a ligand for the P84 neural adhesion molecule. Journal of Biological Chemistry. 1999;274(2):559–562. doi: 10.1074/jbc.274.2.559. [DOI] [PubMed] [Google Scholar]
- 53.Seiffert M, Cant C, Chen Z, et al. Human signal-regulatory protein is expressed on normal, but not on subsets of leukemic myeloid cells and mediates cellular adhesion involving its counterreceptor CD47. Blood. 1999;94(11):3633–3643. [PubMed] [Google Scholar]
- 54.Brooke G, Holbrook JD, Brown MH, Barclay AN. Human lymphocytes interact directly with CD47 through a novel member of the signal regulatory protein (SIRP) family. Journal of Immunology. 2004;173(4):2562–2570. doi: 10.4049/jimmunol.173.4.2562. [DOI] [PubMed] [Google Scholar]
- 55.Piccio L, Vermi W, Boles KS, et al. Adhesion of human T cells to antigen-presenting cells through SIRPβ2-CD47 interaction costimulates T-cell proliferation. Blood. 2005;105(6):2421–2427. doi: 10.1182/blood-2004-07-2823. [DOI] [PubMed] [Google Scholar]
- 56.Legrand N, Huntington ND, Nagasawa M, et al. Functional CD47/signal regulatory protein alpha (SIRPα) interaction is required for optimal human T- and natural killer- (NK) cell homeostasis in vivo . Proceedings of the National Academy of Sciences of the United States of America. 2011;108:13224–13229. doi: 10.1073/pnas.1101398108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sato-Hashimoto M, Saito Y, Ohnishi H, et al. Signal regulatory protein α regulates the homeostasis of T lymphocytes in the spleen. The Journal of Immunology. 2011;187:291–297. doi: 10.4049/jimmunol.1100528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Latour S, Tanaka H, Demeure C, et al. Bidirectional negative regulation of human T and dendritic cells by CD47 and its cognate receptor signal-regulator protein-α: down-regulation of IL-12 responsiveness and inhibition of dendritic cell activation. Journal of Immunology. 2001;167(5):2547–2554. doi: 10.4049/jimmunol.167.5.2547. [DOI] [PubMed] [Google Scholar]
- 59.Hagnerud S, Manna PP, Cella M, et al. Deficit of CD47 results in a defect of marginal zone dendritic cells, blunted immune response to particulate antigen and impairment of skin dendritic cell migration. Journal of Immunology. 2006;176(10):5772–5778. doi: 10.4049/jimmunol.176.10.5772. [DOI] [PubMed] [Google Scholar]
- 60.Saito Y, Iwamura H, Kaneko T, et al. Regulation by SIRPα of dendritic cell homeostasis in lymphoid tissues. Blood. 2010;116(18):3517–3525. doi: 10.1182/blood-2010-03-277244. [DOI] [PubMed] [Google Scholar]
- 61.Van VQ, Lesage S, Bouguermouh S, et al. Expression of the self-marker CD47 on dendritic cells governs their trafficking to secondary lymphoid organs. EMBO Journal. 2006;25(23):5560–5568. doi: 10.1038/sj.emboj.7601415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.De Vries HE, Hendriks JJA, Honing H, et al. Signal-regulatory protein α-CD47 interactions are required for the transmigration of monocytes across cerebral endothelium. Journal of Immunology. 2002;168(11):5832–5839. doi: 10.4049/jimmunol.168.11.5832. [DOI] [PubMed] [Google Scholar]
- 63.Liu Y, Bühring HJ, Zen K, et al. Signal regulatory protein (SIRPα), a cellular ligand for CD47, regulates neutrophil transmigration. Journal of Biological Chemistry. 2002;277(12):10028–10036. doi: 10.1074/jbc.M109720200. [DOI] [PubMed] [Google Scholar]
- 64.Barclay AN. Signal regulatory protein alpha (SIRPα)/CD47 interaction and function. Current Opinion in Immunology. 2009;21(1):47–52. doi: 10.1016/j.coi.2009.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Matozaki T, Murata Y, Okazawa H, Ohnishi H. Functions and molecular mechanisms of the CD47-SIRPα signalling pathway. Trends in Cell Biology. 2009;19(2):72–80. doi: 10.1016/j.tcb.2008.12.001. [DOI] [PubMed] [Google Scholar]
- 66.Lundberg P, Koskinen C, Baldock PA, et al. Osteoclast formation is strongly reduced both in vivo and in vitro in the absence of CD47/SIRPα-interaction. Biochemical and Biophysical Research Communications. 2007;352(2):444–448. doi: 10.1016/j.bbrc.2006.11.057. [DOI] [PubMed] [Google Scholar]
- 67.Maile LA, DeMambro VE, Wai C, et al. An essential role for the association of CD47 to SHPS-1 in skeletal remodeling. Journal of Bone and Mineral Research. 2011;26(9):2068–2081. doi: 10.1002/jbmr.441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Oldenborg PA, Zheleznyak A, Fang YF, Lagenaur CF, Gresham HD, Lindberg FP. Role of CD47 as a marker of self on red blood cells. Science. 2000;288(5473):2051–2054. doi: 10.1126/science.288.5473.2051. [DOI] [PubMed] [Google Scholar]
- 69.Subramanian S, Parthasarathy R, Sen S, Boder ET, Discher DE. Species- and cell type-specific interactions between CD47 and human SIRPα . Blood. 2006;107(6):2548–2556. doi: 10.1182/blood-2005-04-1463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Subramanian S, Boder ET, Discher DE. Phylogenetic divergence in human SIRPalpha-CD47 interactions reveals locus of species-specificity: implications for the binding site. Journal of Biological Chemistry. 2007;282(3):1805–1818. doi: 10.1074/jbc.M603923200. [DOI] [PubMed] [Google Scholar]
- 71.Ogura T, Noguchi T, Murai-Takebe R, Hosooka T, Honma N, Kasuga M. Resistance of B16 melanoma cells to CD47-induced negative regulation of motility as a result of aberrant N-glycosylation of SHPS-1. Journal of Biological Chemistry. 2004;279(14):13711–13720. doi: 10.1074/jbc.M310276200. [DOI] [PubMed] [Google Scholar]
- 72.Mateo V, Lagneaux L, Bron D, et al. CD47 ligation induces caspase-independent cell death in chronic lymphocytic leukemia. Nature Medicine. 1999;5(11):1277–1284. doi: 10.1038/15233. [DOI] [PubMed] [Google Scholar]
- 73.Pettersen RD, Hestdal K, Olafsen MK, Lie SO, Lindberg FP. CD47 signals T cell death. Journal of Immunology. 1999;162(12):7031–7040. [PubMed] [Google Scholar]
- 74.Manna PP, Frazier WA. The mechanism of CD47-dependent killing of T cells: heterotrimeric Gi-dependent inhibition of protein kinase A. Journal of Immunology. 2003;170(7):3544–3553. doi: 10.4049/jimmunol.170.7.3544. [DOI] [PubMed] [Google Scholar]
- 75.Manna PP, Frazier WA. CD47 mediates killing of breast tumor cells via Gi-dependent inhibition of protein kinase A. Cancer Research. 2004;64(3):1026–1036. doi: 10.1158/0008-5472.can-03-1708. [DOI] [PubMed] [Google Scholar]
- 76.Uno S, Kinoshita Y, Azuma Y, et al. Antitumor activity of a monoclonal antibody against CD47 in xenograft models of human leukemia. Oncology Reports. 2007;17(5):1189–1194. [PubMed] [Google Scholar]
- 77.Mateo V, Brown EJ, Biron G, et al. Mechanisms of CD47-induced caspase-independent cell death in normal and leukemic cells: link between phosphatidylserine exposure and cytoskeleton organization. Blood. 2002;100(8):2882–2890. doi: 10.1182/blood-2001-12-0217. [DOI] [PubMed] [Google Scholar]
- 78.Lamy L, Ticchioni M, Rouquette-Jazdanian AK, et al. CD47 and the 19 kDA interacting protein-3 (BNIP3) in T cell apoptosis. Journal of Biological Chemistry. 2003;278(26):23915–23921. doi: 10.1074/jbc.M301869200. [DOI] [PubMed] [Google Scholar]
- 79.Lamy L, Foussat A, Brown EJ, Bornstein P, Ticchioni M, Bernard A. Interactions between CD47 and thrombospondin reduce inflammation. Journal of Immunology. 2007;178(9):5930–5939. doi: 10.4049/jimmunol.178.9.5930. [DOI] [PubMed] [Google Scholar]
- 80.Blundell MP, Worth A, Bouma G, Thrasher AJ. The Wiskott-Aldrich syndrome: the actin cytoskeleton and immune cell function. Disease Markers. 2010;29(3-4):157–175. doi: 10.3233/DMA-2010-0735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Roué G, Bitton N, Yuste VJ, et al. Mitochondrial dysfunction in CD47-mediated caspase-independent cell death: ros production in the absence of cytochrome c and AIF release. Biochimie. 2003;85(8):741–746. doi: 10.1016/s0300-9084(03)00129-9. [DOI] [PubMed] [Google Scholar]
- 82.Sagawa M, Shimizu T, Fukushima N, et al. A new disulfide-linked dimer of a single-chain antibody fragment against human CD47 induces apoptosis in lymphoid malignant cells via the hypoxia inducible factor-1α pathway. Cancer Science. 2011;102(6):1208–1215. doi: 10.1111/j.1349-7006.2011.01925.x. [DOI] [PubMed] [Google Scholar]
- 83.Manna PP, Dimitry J, Oldenborg PA, Frazier WA. CD47 augments fas/CD95-mediated apoptosis. Journal of Biological Chemistry. 2005;280(33):29637–29644. doi: 10.1074/jbc.M500922200. [DOI] [PubMed] [Google Scholar]
- 84.Kikuchi Y, Uno S, Yoshimura Y, et al. A bivalent single-chain Fv fragment against CD47 induces apoptosis for leukemic cells. Biochemical and Biophysical Research Communications. 2004;315(4):912–918. doi: 10.1016/j.bbrc.2004.01.128. [DOI] [PubMed] [Google Scholar]
- 85.Kikuchi Y, Uno S, Kinoshita Y, et al. Apoptosis inducing bivalent single-chain antibody fragments against CD47 showed antitumor potency for multiple myeloma. Leukemia Research. 2005;29(4):445–450. doi: 10.1016/j.leukres.2004.09.005. [DOI] [PubMed] [Google Scholar]
- 86.Johansson U, Higginbottom K, Londei M. CD47 ligation induces a rapid caspase-independent apoptosis-like cell death in human monocytes and dendritic cells. Scandinavian Journal of Immunology. 2004;59(1):40–49. doi: 10.1111/j.0300-9475.2004.01355.x. [DOI] [PubMed] [Google Scholar]
- 87.Koshimizu H, Araki T, Takai S, et al. Expression of CD47/integrin-associated protein induces death of cultured cerebral cortical neurons. Journal of Neurochemistry. 2002;82(2):249–257. doi: 10.1046/j.1471-4159.2002.00965.x. [DOI] [PubMed] [Google Scholar]
- 88.Freyberg MA, Kaiser D, Graf R, Vischer P, Friedl P. Integrin-associated protein and thrombospondin-1 as endothelial mechanosensitive death mediators. Biochemical and Biophysical Research Communications. 2000;271(3):584–588. doi: 10.1006/bbrc.2000.2678. [DOI] [PubMed] [Google Scholar]
- 89.Freyberg MA, Kaiser D, Graf R, Buttenbender J, Friedl P. Proatherogenic flow conditions initiate endothelial apoptosis via thrombospondin-1 and the integrin-associated protein. Biochemical and Biophysical Research Communications. 2001;286(1):141–149. doi: 10.1006/bbrc.2001.5314. [DOI] [PubMed] [Google Scholar]
- 90.Graf R, Freyberg M, Kaiser D, Friedl P. Mechanosensitive induction of apoptosis in fibroblasts is regulated by thrombospondin-1 and integrin associated protein (CD47) Apoptosis. 2002;7(6):493–498. doi: 10.1023/a:1020634924760. [DOI] [PubMed] [Google Scholar]
- 91.Saumet A, Slimane MB, Lanotte M, Lawler J, Dubernard V. Type 3 repeat/C-terminal domain of thrombospondin-1 triggers caspase-independent cell death through CD47/α v β 3 in promyelocytic leukemia NB4 cells. Blood. 2005;106(2):658–667. doi: 10.1182/blood-2004-09-3585. [DOI] [PubMed] [Google Scholar]
- 92.Rath GM, Schneider C, Dedieu S, et al. Thrombospondin-1 C-terminal-derived peptide protects thyroid cells from ceramide-induced apoptosis through the adenylyl cyclase pathway. International Journal of Biochemistry and Cell Biology. 2006;38(12):2219–2228. doi: 10.1016/j.biocel.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 93.Rath GM, Schneider C, Dedieu S, et al. The C-terminal CD47/IAP-binding domain of thrombospondin-1 prevents camptothecin- and doxorubicin-induced apoptosis in human thyroid carcinoma cells. Biochimica et Biophysica Acta. 2006;1763(10):1125–1134. doi: 10.1016/j.bbamcr.2006.08.001. [DOI] [PubMed] [Google Scholar]
- 94.Grossfeld GD, Ginsberg DA, Stein JP, et al. Thrombospondin-1 expression in bladder cancer: association with p53 alterations, tumor angiogenesis, and tumor progression. Journal of the National Cancer Institute. 1997;89(3):219–227. doi: 10.1093/jnci/89.3.219. [DOI] [PubMed] [Google Scholar]
- 95.Sargiannidou I, Zhou J, Tuszynski GP. The role of thrombospondin-1 in tumor progression. Experimental Biology and Medicine. 2001;226(8):726–733. doi: 10.1177/153537020222600803. [DOI] [PubMed] [Google Scholar]
- 96.Sutton CD, O’Byrne K, Goddard JC, et al. Expression of thrombospondin-1 in resected colorectal liver metastases predicts poor prognosis. Clinical Cancer Research. 2005;11(18):6567–6573. doi: 10.1158/1078-0432.CCR-05-0439. [DOI] [PubMed] [Google Scholar]
- 97.Avent N, Judson PA, Parsons SF, et al. Monoclonal antibodies that recognize different membrane proteins that are deficient in Rhnull human erythrocytes. One group of antibodies reacts with a variety of cells and tissues whereas the other group is erythroid-specific. Biochemical Journal. 1988;251(2):499–505. doi: 10.1042/bj2510499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Cartron JP. RH blood group system and molecular basis of Rh-deficiency. Best Practice & Research Clinical Haematology . 1999;12(4):655–689. doi: 10.1053/beha.1999.0047. [DOI] [PubMed] [Google Scholar]
- 99.Rybicki AC, Schwartz RS, Hustedt EJ, Cobb CE. Increased rotational mobility and extractability of band 3 from protein 4.2-deficient erythrocyte membranes: evidence of a role for protein 4.2 in strengthening the band 3-cytoskeleton linkage. Blood. 1996;88(7):2745–2753. [PubMed] [Google Scholar]
- 100.Tanner MJA. Band 3 anion exchanger and its involvement in erythrocyte and kidney disorders. Current Opinion in Hematology. 2002;9(2):133–139. doi: 10.1097/00062752-200203000-00009. [DOI] [PubMed] [Google Scholar]
- 101.Bruce LJ, Beckmann R, Ribeiro ML, et al. A band 3-based macrocomplex of integral and peripheral proteins in the RBC membrane. Blood. 2003;101(10):4180–4188. doi: 10.1182/blood-2002-09-2824. [DOI] [PubMed] [Google Scholar]
- 102.Booth PB, Serjeantson S, Woodfield DG, Amato D. Selective depression of blood group antigens associated with hereditary ovalocytosis among Melanesians. Vox Sanguinis. 1977;32(2):99–110. doi: 10.1111/j.1423-0410.1977.tb00612.x. [DOI] [PubMed] [Google Scholar]
- 103.Bruce LJ, Ghosh S, King MJ, et al. Absence of CD47 in protein 4.2-deficient hereditary spherocytosis in man: an interaction between the Rh complex and the band 3 complex. Blood. 2002;100(5):1878–1885. doi: 10.1182/blood-2002-03-0706. [DOI] [PubMed] [Google Scholar]
- 104.Dahl KN, Westhoff CM, Discher DE. Fractional attachment of CD47 (IAP) to the erythrocyte cytoskeleton and visual colocalization with Rh protein complexes. Blood. 2003;101(3):1194–1199. doi: 10.1182/blood-2002-04-1187. [DOI] [PubMed] [Google Scholar]
- 105.Mouro-Chanteloup I, Johansen M, Brown EJ, Cartron JPCY. Normal red cell membrane expression of Rh and RhAG polypeptides in CD47-deficient mice. Vox Sanguinis. 2000;78, article P030 [Google Scholar]
- 106.Mouro-Chanteloup I, Delaunay J, Gane P, et al. Evidence that the red cell skeleton protein 4.2 interacts with the Rh membrane complex member CD47. Blood. 2003;101(1):338–344. doi: 10.1182/blood-2002-04-1285. [DOI] [PubMed] [Google Scholar]
- 107.Anniss AM, Sparrow RL. Expression of CD47 (integrin-associated protein) decreases on red blood cells during storage. Transfusion and Apheresis Science. 2002;27(3):233–238. doi: 10.1016/s1473-0502(02)00070-8. [DOI] [PubMed] [Google Scholar]
- 108.Kamel N, Goubran F, Ramsis N, Ahmed AS. Effects of storage time and leucocyte burden of packed and buffy-coat depleted red blood cell units on red cell storage lesion. Blood Transfusion. 2010;8(4):260–266. doi: 10.2450/2009.0131-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Stewart A, Urbaniak S, Turner M, Bessos H. The application of a new quantitative assay for the monitoring of integrin-associated protein CD47 on red blood cells during storage and comparison with the expression of CD47 and phosphatidylserine with flow cytometry. Transfusion. 2005;45(9):1496–1503. doi: 10.1111/j.1537-2995.2005.00564.x. [DOI] [PubMed] [Google Scholar]
- 110.Gilson CR, Kraus TS, Hod EA, et al. A novel mouse model of red blood cell storage and posttransfusion in vivo survival. Transfusion. 2009;49(8):1546–1553. doi: 10.1111/j.1537-2995.2009.02173.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Burger P, Hilarius-Stokman P, de Korte D, van den Berg TK, van Bruggen R. CD47 functions as a molecular switch for erythrocyte phagocytosis. Blood. 2012;119:5512–5521. doi: 10.1182/blood-2011-10-386805. [DOI] [PubMed] [Google Scholar]
- 112.Holovati JL, Wong KA, Webster JM, Acker JP. The effects of cryopreservation on red blood cell microvesiculation, phosphatidylserine externalization, and CD47 expression. Transfusion. 2008;48(8):1658–1668. doi: 10.1111/j.1537-2995.2008.01735.x. [DOI] [PubMed] [Google Scholar]
- 113.Rubin O, Canelli G, Delobel J, Lion N, Tissot JD. Red blood cell microparticles: clinical relevance. Transfusion Medicine and Hemotherapy. 2012;39:342–347. doi: 10.1159/000342228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Kriebardis AG, Antonelou MH, Stamoulis KE, Economou-Petersen E, Margaritis LH, Papassideri IS. RBC-derived vesicles during storage: ultrastructure, protein composition, oxidation, and signaling components. Transfusion. 2008;48(9):1943–1953. doi: 10.1111/j.1537-2995.2008.01794.x. [DOI] [PubMed] [Google Scholar]
- 115.Fossati-Jimack L, Azeredo Da Silveira S, Moll T, et al. Selective increase of autoimmune epitope expression on aged erythrocytes in mice: implications in anti-erythrocyte autoimmune responses. Journal of Autoimmunity. 2002;18(1):17–25. doi: 10.1006/jaut.2001.0563. [DOI] [PubMed] [Google Scholar]
- 116.Saxena RK, Bhardwaj N, Sachar S, Puri N, Khandelwal S. A double in vivo biotinylation technique for objective assessment of aging and clearance of mouse erythrocytes in blood circulation. Transfusion Medicine and Hemotherapy. 2012;39:335–341. doi: 10.1159/000342524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Khandelwal S, Van Rooijen N, Saxena RK. Reduced expression of CD47 during murine red blood cell (RBC) senescence and its role in RBC clearance from the circulation. Transfusion. 2007;47(9):1725–1732. doi: 10.1111/j.1537-2995.2007.01348.x. [DOI] [PubMed] [Google Scholar]
- 118.Ballas SK, Mohandas N. Pathophysiology of vaso-occlusion. Hematology/Oncology Clinics of North America. 1996;10(6):1221–1239. doi: 10.1016/s0889-8588(05)70396-8. [DOI] [PubMed] [Google Scholar]
- 119.Ballas SK, Mohandas N. Sickle red cell microrheology and sickle blood rheology. Microcirculation. 2004;11(2):209–225. doi: 10.1080/10739680490279410. [DOI] [PubMed] [Google Scholar]
- 120.Brittain JE, Mlinar KJ, Anderson CS, Orringer EP, Parise LV. Integrin-associated protein is an adhesion receptor on sickle red blood cells for immobilized thrombospondin. Blood. 2001;97(7):2159–2164. doi: 10.1182/blood.v97.7.2159. [DOI] [PubMed] [Google Scholar]
- 121.Brittain JE, Mlinar KJ, Anderson CS, Orringer EP, Parise LV. Activation of sickle red blood cell adhesion via integrin-associated protein/CD47-induced signal transduction. Journal of Clinical Investigation. 2001;107(12):1555–1562. doi: 10.1172/JCI10817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Grabowski GA. Phenotype, diagnosis, and treatment of Gaucher’s disease. The Lancet. 2008;372(9645):1263–1271. doi: 10.1016/S0140-6736(08)61522-6. [DOI] [PubMed] [Google Scholar]
- 123.Bratosin D, Tissier JP, Lapillonne H, et al. A cytometric study of the red blood cells in gaucher disease reveals their abnormal shape that may be involved in increased erythrophagocytosis. Cytometry Part B. 2011;80(1):28–37. doi: 10.1002/cyto.b.20539. [DOI] [PubMed] [Google Scholar]
- 124.Jiang H, Chess L. How the immune system achieves self-nonself discrimination during adaptive immunity. Advances in Immunology. 2009;102:95–133. doi: 10.1016/S0065-2776(09)01202-4. [DOI] [PubMed] [Google Scholar]
- 125.Janeway CA, Medzhitov R. Innate immune recognition. Annual Review of Immunology. 2002;20:197–216. doi: 10.1146/annurev.immunol.20.083001.084359. [DOI] [PubMed] [Google Scholar]
- 126.Zipfel PF, Skerka C. Complement regulators and inhibitory proteins. Nature Reviews Immunology. 2009;9(10):729–740. doi: 10.1038/nri2620. [DOI] [PubMed] [Google Scholar]
- 127.Kärre K, Ljunggren HG, Piontek G, Kiessling R. Selective rejection of H-2-deficient lymphoma variants suggests alternative immune defence strategy. Nature. 1986;319(6055):675–678. doi: 10.1038/319675a0. [DOI] [PubMed] [Google Scholar]
- 128.Lanier LL. NK cell recognition. Annual Review of Immunology. 2004;23:225–274. doi: 10.1146/annurev.immunol.23.021704.115526. [DOI] [PubMed] [Google Scholar]
- 129.Kumar V, McNerney ME. A new self: MHC-class-I-independent natural-killer-cell self-tolerance. Nature Reviews Immunology. 2005;5(5):363–374. doi: 10.1038/nri1603. [DOI] [PubMed] [Google Scholar]
- 130.Steevels TAM, Meyaard L. Immune inhibitory receptors: essential regulators of phagocyte function. European Journal of Immunology. 2011;41(3):575–587. doi: 10.1002/eji.201041179. [DOI] [PubMed] [Google Scholar]
- 131.Ravetch JV, Lanier LL. Immune inhibitory receptors. Science. 2000;290(5489):84–89. doi: 10.1126/science.290.5489.84. [DOI] [PubMed] [Google Scholar]
- 132.Liénard H, Bruhns P, Malbec O, Fridman WH, Daëron M. Signal regulatory proteins negatively regulate immunoreceptor-dependent cell activation. Journal of Biological Chemistry. 1999;274(45):32493–32499. doi: 10.1074/jbc.274.45.32493. [DOI] [PubMed] [Google Scholar]
- 133.Tonks NK, Neel BG. From form to function: signaling by protein tyrosine phosphatases. Cell. 1996;87(3):365–368. doi: 10.1016/s0092-8674(00)81357-4. [DOI] [PubMed] [Google Scholar]
- 134.Blazar BR, Lindberg FP, Ingulli E, et al. CD47 (Integrin-associated Protein) engagement of dendritic cell and macrophage counterreceptors is required to prevent the clearance of donor lymphohematopoietic cells. Journal of Experimental Medicine. 2001;194(4):541–549. doi: 10.1084/jem.194.4.541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Goodman JW, Smith LH. Erythrocyte life span in normal mice and in radiation bone marrow chimeras. American Journal of Physiology. 1961;200:764–770. doi: 10.1152/ajplegacy.1961.200.4.764. [DOI] [PubMed] [Google Scholar]
- 136.Horky J, Vacha J, Znojil V. Comparison of life span of erythrocytes in some inbred strains of mouse using 14C-labelled glycine. Physiologia Bohemoslovaca. 1978;27(3):209–217. [PubMed] [Google Scholar]
- 137.Van Rooijen N, Sanders A. Liposome mediated depletion of macropbages: mechanism of action, preparation of liposomes and applications. Journal of Immunological Methods. 1994;174(1-2):83–93. doi: 10.1016/0022-1759(94)90012-4. [DOI] [PubMed] [Google Scholar]
- 138.Ishikawa-Sekigami T, Kaneko Y, Okazawa H, et al. SHPS-1 promotes the survival of circulating erythrocytes through inhibition of phagocytosis by splenic macrophages. Blood. 2006;107(1):341–348. doi: 10.1182/blood-2005-05-1896. [DOI] [PubMed] [Google Scholar]
- 139.Olsson M, Bruhns P, Frazier WA, Ravetch JV, Oldenborg PA. Platelet homeostasis is regulated by platelet expression of CD47 under normal conditions and in passive immune thrombocytopenia. Blood. 2005;105(9):3577–3582. doi: 10.1182/blood-2004-08-2980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Yamao T, Noguchi T, Takeuchi U, et al. Negative regulation of platelet clearance and of the macrophage phagocytic response by the transmembrane glycoprotein SHPS-1. Journal of Biological Chemistry. 2002;277(42):39833–39839. doi: 10.1074/jbc.M203287200. [DOI] [PubMed] [Google Scholar]
- 141.Gardai SJ, McPhillips KA, Frasch SC, et al. Cell-surface calreticulin initiates clearance of viable or apoptotic cells through trans-activation of LRP on the phagocyte. Cell. 2005;123(2):321–334. doi: 10.1016/j.cell.2005.08.032. [DOI] [PubMed] [Google Scholar]
- 142.Nilsson A, Vesterlund L, Oldenborg PA. Macrophage expression of LRP1, a receptor for apoptotic cells and unopsonized erythrocytes, can be regulated by glucocorticoids. Biochemical and Biophysical Research Communications. 2012;417(4):1304–1309. doi: 10.1016/j.bbrc.2011.12.137. [DOI] [PubMed] [Google Scholar]
- 143.Nilsson A, Oldenborg PA. CD47 promotes both phosphatidylserine-independent and phosphatidylserine-dependent phagocytosis of apoptotic murine thymocytes by non-activated macrophages. Biochemical and Biophysical Research Communications. 2009;387(1):58–63. doi: 10.1016/j.bbrc.2009.06.121. [DOI] [PubMed] [Google Scholar]
- 144.Okazawa H, Motegi SI, Ohyama N, et al. Negative regulation of phagocytosis in macrophages by the CD47-SHPS-1 system. Journal of Immunology. 2005;174(4):2004–2011. doi: 10.4049/jimmunol.174.4.2004. [DOI] [PubMed] [Google Scholar]
- 145.Oldenborg PA, Gresham HD, Lindberg FP. CD47-signal regulatory protein α (SIRPα) regulates Fcγ and complement receptor-mediated phagocytosis. Journal of Experimental Medicine. 2001;193(7):855–861. doi: 10.1084/jem.193.7.855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Oldenborg PA, Gresham HD, Chen Y, Izui S, Lindberg FP. Lethal autoimmune hemolytic anemia in CD47-deficient nonobese diabetic (NOD) mice. Blood. 2002;99(10):3500–3504. doi: 10.1182/blood.v99.10.3500. [DOI] [PubMed] [Google Scholar]
- 147.Kozlowski M, Mlinaric-Rascan I, Feng GS, Shen R, Pawson T, Siminovitch KA. Expression and catalytic activity of the tyrosine phosphatase PTP1C is severely impaired in motheaten and viable motheaten mice. Journal of Experimental Medicine. 1993;178(6):2157–2163. doi: 10.1084/jem.178.6.2157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Schultz LD, Coman DR, Bailey CL, et al. “Viable motheaten”, a new allele at the motheaten locus. I. Pathology. American Journal of Pathology. 1984;116(2):179–192. [PMC free article] [PubMed] [Google Scholar]
- 149.Shultz LD, Schweitzer PA, Rajan TV, et al. Mutations at the murine motheaten locus are within the hematopoietic cell protein-tyrosine phosphatase (Hcph) gene. Cell. 1993;73(7):1445–1454. doi: 10.1016/0092-8674(93)90369-2. [DOI] [PubMed] [Google Scholar]
- 150.Olsson M, Nilsson A, Oldenborg PA. Dose-dependent inhibitory effect of CD47 in macrophage uptake of IgG-opsonized murine erythrocytes. Biochemical and Biophysical Research Communications. 2007;352(1):193–197. doi: 10.1016/j.bbrc.2006.11.002. [DOI] [PubMed] [Google Scholar]
- 151.Gitik M, Liraz-Zaltsman S, Oldenborg PA, Reichert F, Rotshenker S. Myelin down-regulates myelin phagocytosis by microglia and macrophages through interactions between CD47 on myelin and SIRPα (signal regulatory protein-α) on phagocytes. Journal of Neuroinflammation. 2011;8, article 24 doi: 10.1186/1742-2094-8-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Janka GE. Hemophagocytic syndromes. Blood Reviews. 2007;21(5):245–253. doi: 10.1016/j.blre.2007.05.001. [DOI] [PubMed] [Google Scholar]
- 153.Kuriyama T, Takenaka K, Kohno K, et al. Engulfment of hematopoietic stem cells caused by down-regulation of CD47 is critical in the pathogenesis of hemophagocytic lymphohistiocytosis. Blood. 2012;120(19):4058–4067. doi: 10.1182/blood-2012-02-408864. [DOI] [PubMed] [Google Scholar]
- 154.Olsson M, Oldenborg PA. CD47 on experimentally senescent murine RBCs inhibits phagocytosis following Fcγ receptor-mediated but not scavenger receptor-mediated recognition by macrophages. Blood. 2008;112(10):4259–4267. doi: 10.1182/blood-2008-03-143008. [DOI] [PubMed] [Google Scholar]
- 155.Tsai RK, Discher DE. Inhibition of “self” engulfment through deactivation of myosin-II at the phagocytic synapse between human cells. Journal of Cell Biology. 2008;180(5):989–1003. doi: 10.1083/jcb.200708043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Jonsson AH, Yokoyama WM. Natural killer cell tolerance: licensing and other mechanisms. Advances in Immunology. 2009;101:27–79. doi: 10.1016/S0065-2776(08)01002-X. [DOI] [PubMed] [Google Scholar]
- 157.Wang H, Madariaga ML, Wang S, Van Rooijen N, Oldenborg PA, Yang YG. Lack of CD47 on nonhematopoietic cells induces split macrophage tolerance to CD47null cells. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(34):13744–13749. doi: 10.1073/pnas.0702881104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Yang YG. CD47 in xenograft rejection and tolerance induction: review article. Xenotransplantation. 2010;17(4):267–273. doi: 10.1111/j.1399-3089.2010.00601.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Elliott MR, Ravichandran KS. Clearance of apoptotic cells: implications in health and disease. Journal of Cell Biology. 2010;189(7):1059–1070. doi: 10.1083/jcb.201004096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Gardai SJ, Bratton DL, Ogden CA, Henson PM. Recognition ligands on apoptotic cells: a perspective. Journal of Leukocyte Biology. 2006;79(5):896–903. doi: 10.1189/jlb.1005550. [DOI] [PubMed] [Google Scholar]
- 161.Ravichandran KS, Lorenz U. Engulfment of apoptotic cells: signals for a good meal. Nature Reviews Immunology. 2007;7(12):964–974. doi: 10.1038/nri2214. [DOI] [PubMed] [Google Scholar]
- 162.Ravichandran KS. Beginnings of a good apoptotic meal: the find-me and eat-me signaling pathways. Immunity. 2011;35(4):445–455. doi: 10.1016/j.immuni.2011.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Hanayama R, Nagata S. Impaired involution of mammary glands in the absence of milk fat globule EGF factor 8. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(46):16886–16891. doi: 10.1073/pnas.0508599102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Sandahl M, Hunter DM, Strunk KE, Earp HS, Cook RS. Epithelial cell-directed efferocytosis in the post-partum mammary gland is necessary for tissue homeostasis and future lactation. BMC Developmental Biology. 2010;10, article 122 doi: 10.1186/1471-213X-10-122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Tada K, Tanaka M, Hanayama R, et al. Tethering of apoptotic cells to phagocytes through binding of CD47 to src homology 2 domain-bearing protein tyrosine phosphatase substrate-1. Journal of Immunology. 2003;171(11):5718–5726. doi: 10.4049/jimmunol.171.11.5718. [DOI] [PubMed] [Google Scholar]
- 166.Ogden CA, DeCathelineau A, Hoffmann PR, et al. C1q and mannose binding lectin engagement of cell surface calreticulin and CD91 initiates macropinocytosis and uptake of apoptotic cells. Journal of Experimental Medicine. 2001;194(6):781–795. doi: 10.1084/jem.194.6.781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Vandivier RW, Ogden CA, Fadok VA, et al. Role of surfactant proteins A, D, and C1q in the clearance of apoptotic cells in vivo and in vitro: calreticulin and CD91 as a common collectin receptor complex. Journal of Immunology. 2002;169(7):3978–3986. doi: 10.4049/jimmunol.169.7.3978. [DOI] [PubMed] [Google Scholar]
- 168.Löwenberg M, Stahn C, Hommes DW, Buttgereit F. Novel insights into mechanisms of glucocorticoid action and the development of new glucocorticoid receptor ligands. Steroids. 2008;73(9-10):1025–1029. doi: 10.1016/j.steroids.2007.12.002. [DOI] [PubMed] [Google Scholar]
- 169.Schleimer RP. An overview of glucocorticoid anti-inflammatory actions. European Journal of Clinical Pharmacology. 1993;45(1):S3–S7. doi: 10.1007/BF01844196. [DOI] [PubMed] [Google Scholar]
- 170.Giles KM, Ross K, Rossi AG, Hotchin NA, Haslett C, Dransfield I. Glucocorticoid augmentation of macrophage capacity for phagocytosis of apoptotic cells is associated with reduced p130Cas expression, loss of paxillin/pyk2 phosphorylation, and high levels of active rac. Journal of Immunology. 2001;167(2, supplement 1):976–986. doi: 10.4049/jimmunol.167.2.976. [DOI] [PubMed] [Google Scholar]
- 171.Liu Y, Cousin JM, Hughes J, et al. Glucocorticoids promote nonphlogistic phagocytosis of apoptotic leukocytes. Journal of Immunology. 1999;162(6):3639–3646. [PubMed] [Google Scholar]
- 172.Jaiswal S, Jamieson CHM, Pang WW, et al. CD47 is upregulated on circulating hematopoietic stem cells and leukemia cells to avoid phagocytosis. Cell. 2009;138(2):271–285. doi: 10.1016/j.cell.2009.05.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Majeti R, Chao MP, Alizadeh AA, et al. CD47 is an adverse prognostic factor and therapeutic antibody target on human acute myeloid leukemia stem cells. Cell. 2009;138(2):286–299. doi: 10.1016/j.cell.2009.05.045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Willingham SB, Volkmer JP, Gentles AJ, et al. The CD47-signal regulatory protein alpha (SIRPa) interaction is a therapeutic target for human solid tumors. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:6662–6667. doi: 10.1073/pnas.1121623109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Theocharides APA, Jin L, Cheng PY, et al. Disruption of SIRPα signaling in macrophages eliminates human acute myeloid leukemia stem cells in xenografts. The Journal of Experimental Medicine. 2012;209(10):1883–1899. doi: 10.1084/jem.20120502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Chao MP, Alizadeh AA, Tang C, et al. Anti-CD47 antibody synergizes with rituximab to promote phagocytosis and eradicate non-hodgkin lymphoma. Cell. 2010;142(5):699–713. doi: 10.1016/j.cell.2010.07.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Zhao XW, van Beek EM, Schornagel K, et al. CD47-signal regulatory protein-α (SIRPα) interactions form a barrier for antibody-mediated tumor cell destruction. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:18342–118347. doi: 10.1073/pnas.1106550108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Yang YG, Sykes M. Xenotransplantation: current status and a perspective on the future. Nature Reviews Immunology. 2007;7(7):519–531. doi: 10.1038/nri2099. [DOI] [PubMed] [Google Scholar]
- 179.Karlsson-Parra A, Ridderstad A, Wallgren AC, Möller E, Ljunggren HG, Korsgren O. Xenograft rejection of porcine islet-like cell clusters in normal and natural killer cell-depleted mice. Transplantation. 1996;61(9):1313–1320. doi: 10.1097/00007890-199605150-00005. [DOI] [PubMed] [Google Scholar]
- 180.Wu G, Korsgren O, Zhang J, Song Z, Van Rooijen N, Tibell A. Pig islet xenograft rejection is markedly delayed in macrophage-depleted mice: a study in streptozotocin diabetic animals. Xenotransplantation. 2000;7(3):214–220. doi: 10.1034/j.1399-3089.2000.00071.x. [DOI] [PubMed] [Google Scholar]
- 181.Abe M, Cheng J, Qi J, et al. Elimination of porcine hemopoietic cells by macrophages in mice. Journal of Immunology. 2002;168(2):621–628. doi: 10.4049/jimmunol.168.2.621. [DOI] [PubMed] [Google Scholar]
- 182.Wang H, Verhalen J, Madariaga ML, et al. Attenuation of phagocytosis of xenogeneic cells by manipulating CD47. Blood. 2007;109(2):836–842. doi: 10.1182/blood-2006-04-019794. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Ide K, Wang H, Tahara H, et al. Role for CD47-SIRPα signaling in xenograft rejection by macrophages. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(12):5062–5066. doi: 10.1073/pnas.0609661104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Han X, Sterling H, Chen Y, et al. CD47, a ligand for the macrophage fusion receptor, participates in macrophage multinucleation. Journal of Biological Chemistry. 2000;275(48):37984–37992. doi: 10.1074/jbc.M002334200. [DOI] [PubMed] [Google Scholar]
- 185.Uluçkan Ö, Becker SN, Deng H, et al. CD47 regulates bone mass and tumor metastasis to bone. Cancer Research. 2009;69(7):3196–3204. doi: 10.1158/0008-5472.CAN-08-3358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Van Beek EM, De Vries TJ, Mulder L, et al. Inhibitory regulation of osteoclast bone resorption by signal regulatory protein α . FASEB Journal. 2009;23(12):4081–4090. doi: 10.1096/fj.09-131557. [DOI] [PubMed] [Google Scholar]
- 187.Vignery A. Osteoclasts and giant cells: macrophage-macrophage fusion mechanism. International Journal of Experimental Pathology. 2000;81(5):291–304. doi: 10.1111/j.1365-2613.2000.00164.x. [DOI] [PMC free article] [PubMed] [Google Scholar]