

Microtubules are the basic elements of the cytoskeleton. This study demonstrates that a specific mutation in mec-7/β-tubulin is necessary for the correct number of neurites a neuron extends in vivo and the neuron’s capacity for axonal regeneration following injury.
Abstract
Microtubules have been known for decades to be basic elements of the cytoskeleton. They form long, dynamic, rope-like structures within the cell that are essential for mitosis, maintenance of cell shape, and intracellular transport. More recently, in vitro studies have implicated microtubules as signaling molecules that, through changes in their stability, have the potential to trigger growth of axons and dendrites in developing neurons. In this study, we show that specific mutations in the Caenorhabditis elegans mec-7/β-tubulin gene cause ectopic axon formation in mechanosensory neurons in vivo. In mec-7 mutants, the ALM mechanosensory neuron forms a long ectopic neurite that extends posteriorly, a phenotype that can be mimicked in wild-type worms with a microtubule-stabilizing drug (paclitaxel), and suppressed by mutations in unc-33/CRMP2 and the kinesin-related gene, vab-8. Our results also reveal that these ectopic neurites contain RAB-3, a marker for presynaptic loci, suggesting that they have axon-like properties. Interestingly, in contrast with the excessive axonal growth observed during development, mec-7 mutants are inhibited in axonal regrowth and remodeling following axonal injury. Together our results suggest that MEC-7/β-tubulin integrity is necessary for the correct number of neurites a neuron generates in vivo and for the capacity of an axon to regenerate.
INTRODUCTION
A neuron becomes polarized early in its development, when it breaks its symmetry by extending an axon and dendrites. The polarity of a neuron is defined by its orientation within the animal, the number of processes it extends, and the specification of neurite identity. In the nematode Caenorhabditis elegans, alterations to neuronal polarity have been detected by changes in cell orientation in the body of the worm, leading to the discovery of several molecules in the Wnt/Frizzled and Netrin/DCC pathway that are critical for this process in vivo (Adler et al., 2006; Chang et al., 2006; Hilliard and Bargmann, 2006; Prasad and Clark, 2006; Quinn et al., 2008). In parallel, studies using cultured hippocampal neurons have provided insights into the molecules that specify neurite identity, thereby establishing whether a neuronal process develops into an axon or a dendrite (Polleux and Snider, 2010). Local destabilization of the actin network at the tip of a neurite has been shown to act as an early signal within the cell for axon specification (Bradke and Dotti, 1999; Witte and Bradke, 2008; Neukirchen and Bradke, 2011). However, several studies have identified molecules, such as collapsin response–mediated protein 2 (CRMP2), synapses of amphids defective (SAD) kinases, and glycogen synthase kinase-3β (GSK3β), that trigger axon specification independently of actin through microtubule dynamics (Jiang et al., 2005; Kishi et al., 2005; Yoshimura et al., 2005; Maniar et al., 2012). Furthermore, growing evidence from in vitro experiments suggests that microtubules do not merely act as simple building blocks or cargo tracks in this process, but may provide instructive signals for axon specification (Baas, 2002; Witte et al., 2008; Hoogenraad and Bradke, 2009; Stiess and Bradke, 2011). Interestingly, microtubule dynamics have also been implicated in axonal regeneration, indicating microtubules are important not only during axon development but also for regrowth following injury (Erturk et al., 2007; Stone et al., 2010; Chen et al., 2011; Hellal et al., 2011; Sengottuvel and Fischer, 2011).
Although significant advances have been made in understanding the molecular mechanisms of neuronal polarization governed by neuron orientation and neurite identity, we know much less about how a neuron is able to control the number of neurites it forms in vivo. Recent results have shown the planar cell polarity molecules VANG-1 and PRKL-1 are involved in this process (Sanchez-Alvarez et al., 2011).
In this study, we performed an unbiased genetic screen in C. elegans mechanosensory neurons to identify mutations affecting the number of neurites that a neuron generates. We show that a mutation in mec-7, which encodes β-tubulin, causes ALM mechanosensory neurons to develop long, ectopic neurites containing presynaptic loci, in addition to their normal axons. Time-lapse morphological characterization suggests that this defect results from a disruption in the maintenance of neuronal polarity, with pharmacological experiments revealing that similar defects arise from hyperstable microtubules. Converse to the excessive axon growth observed during development, we also demonstrate that axons of mec-7 animals display a reduced capacity for regeneration following laser-induced injury. Our results reveal that β-tubulin mutations can alter the number of axons generated in vivo and inhibit multiple stages of axonal regeneration.
RESULTS
Semidominant mutations in MEC-7/β-tubulin induce neurite outgrowth in vivo
The C. elegans ALM mechanosensory neuron is bilaterally symmetrical and has a single process that extends anteriorly, with a ventral branch of this process forming synaptic connections in the nerve ring (Figure 1A). Occasionally, a short posterior neurite that is less than twice the length of the cell body is observed, although this is regarded as a protrusion, rather than a full process (White, 1986; Watari-Goshima et al., 2007). The ALM neuron was visualized using the zdIs5(Pmec-4::GFP) transgene, which expresses green fluorescent protein (GFP) in the ALM, AVM, PVM, and PLM mechanosensory neurons. In an unbiased, forward genetic screen aimed at finding mutants with ectopic neurite outgrowth in the ALM neuron, we isolated a mutation, ky852, in which both the left and right ALM neurons extended a long posterior neurite, essentially converting them into bipolar neurons (Figure 1, C and D). This phenotype was fully penetrant and semidominant, with heterozygotes (ky852/+) displaying a less severe phenotype with shorter posterior processes than those of homozygotes (Figure 1, B and D). Interestingly, we never observed a reversed ALM phenotype in which the anterior process was misrouted posteriorly, a common occurrence in other known ALM polarity mutants (Wolf et al., 1998; Hilliard and Bargmann, 2006; Prasad and Clark, 2006; Levy-Strumpf and Culotti, 2007; Watari-Goshima et al., 2007; Pan et al., 2008). This suggests that the ky852 phenotype might result from disruption of a different molecular process that, rather than affecting orientation of a neuron within the animal, specifically regulates the number of neurites it develops. Moreover, unlike other polarity mutants, the ky852 mutant also exhibited ectopic neurite formation in the mechanosensory neurons AVM and PVM, and overextension of the posterior neurite in the PLM neuron (Figure S1). Behavioral analysis of ky852 mutants with the light-touch assay (Chalfie and Sulston, 1981) revealed a mild touch insensitivity compared with wild-type animals, suggesting that the functional integrity of the mechanosensory neurons had been affected (Figure S2). To determine whether the mutation affected other classes of neurons, we visualized the D-type GABAergic motor neurons (Punc-25::GFP transgene) and the oxygen sensory neurons, AQR, PQR, and URX (Pgcy-36::GFP transgene) in the ky852 mutants. The morphology of these motor and oxygen sensory neurons appeared unaltered, suggesting that the ky852 mutation specifically affected the mechanosensory neurons.
FIGURE 1:

Posterior neurite outgrowth defects in ALM neurons of ky852 mutants. Morphology of ALM in wild type (A), ky852 heterozygotes (B), and ky852 homozygotes (C). White arrowheads indicate the ectopic posterior process. (D) Quantification of ALM phenotypes, classified as normal (no posterior process), moderate (medium length posterior process), and severe (long posterior process). Each data set is based on an n of at least 100 animals. Scale bar: 25 μm.
The ALM mechanosensory neurons develop during embryogenesis, a time at which mec-7 also begins to be expressed (Hamelin et al., 1992), and are fully extended to the head of the animal at hatching. We reasoned that the ectopic ALM process in ky852 mutants could either emerge during early development, which would suggest a disruption in the establishment of neuronal polarity, or arise after the development of the anterior neurite, which would indicate a defect in the maintenance of the neuronal polarity. To investigate this, we performed time-lapse analysis of ALM development and found that the ectopic neurite emerged during the L1 stage, after the normal anterior neurite had already extended to the head of the animal (Figure 2). This supports the notion that the ky852 phenotype differs from other neuronal polarity mutants, in which polarity defects are already seen in the embryo when the cell first begins to develop asymmetry (Hilliard and Bargmann, 2006), and therefore indicates the ky852 mutation affects signaling processes occurring later in development.
FIGURE 2:

Ectopic neurites in ky852 mutants develop following embryogenesis. Morphology of ALM neurons in wild type and ky852 mutants in the embryo (A and B), at hatching (C and D), and 5 h after hatching (E and F). (G) Quantification of ALM neurons with posterior neurite defects in ky852 mutants during developmental stages corresponding to the images in (A–F). Each data set is based on an n of at least 100 animals. Error bars represent the SE of proportion. Scale bars: 20 μm.
Using single-nucleotide polymorphism (SNP) analysis (Wicks et al., 2001; Davis et al., 2005; Davis and Hammarlund, 2006), we mapped the ky852 mutation to an interval of chromosome X including the mec-7 gene, which encodes a β-tubulin that is specifically expressed in the six mechanosensory neurons of C. elegans and forms a component of the 15 protofilament microtubules unique to these neurons (Chalfie and Thomson, 1982). Several lines of evidence indicated the ky852 phenotype was due to a mutation in the mec-7 gene. First, sequence analysis of the ky852 mutant revealed a single base-pair change (ccg to tcg) in the mec-7 gene that generated a missense mutation (proline to serine at amino acid position 220) in the encoded protein (Figure 3A). Second, when we introduced a wild-type copy of mec-7 under the control of the mec-4 promoter (Pmec-4::mec-7) into ky852 mutant animals, we found that this transgene could partially rescue the ALM defect, with the length of the ectopic process being reduced or returned to wild-type length in the majority of animals (Figure 3B). This partial rescue was consistent with the mutation being semidominant, as heterozygous animals showed ectopic processes of intermediate lengths (Figure 1, B and D), due to the presence of both wild type and mutant copies of the gene. Third, expression of the mec-7(ky852) mutant allele in wild-type animals induced ectopic posterior outgrowth in some ALM neurons (Figure 3C), further supporting the semidominant role of this gene. Finally, the mec-7 gene has been extensively characterized for mutations that affect the touch sensitivity of C. elegans, with more than 50 alleles identified (Savage et al., 1989, 1994). Although visualization in these studies of MEC-7 with antibodies indicated that the vast majority of these alleles cause degenerative phenotypes in ALM, two alleles, u278 and u170, presented posterior outgrowth in ALM similar to that observed in the ky852 mutant animals (Savage et al., 1994). We visualized and quantified the phenotypes of the mec-7(u278) allele in the zdIs5(Pmec-4::GFP) background and found it had a qualitatively similar but less severe phenotype compared with the ky852 mutant, with a lower penetrance of the ALM defect (Figure 3D). Complementation analysis between the ky852 and mec-7(u278) alleles revealed a significant worsening of the defect in the double heterozygotes (noncomplementation) compared with either individual heterozygote (Figure 3D), confirming ky852 to be a novel allele of mec-7.
FIGURE 3:

The ky852 phenotype is caused by a mutation in the mec-7 gene. (A) Diagram of the mec-7 gene. Solid bars represent exons and connecting lines represent introns. The ky852 mutation and two other alleles that cause similar ALM neurite outgrowth phenotypes are indicated. (B) Rescue of ALM defects by insertion of wild-type mec-7 into mec-7(ky852) mutants. Two independent lines that were generated with different concentrations of injected transgene are shown. Phenotypic distributions between transgenics and nontransgenic controls were significantly different in both cases (chi-square test: p < 0.0001). (C) Induction of mec-7(ky852) phenotype by insertion of mutant mec-7 into wild-type animals. Two independent lines that were generated with different concentrations of injected transgene are shown. In both cases, transgenics had a reduced percentage of normal ALMs and an increased percentage of ALMs with intermediate length posterior process defects (Fisher’s exact test: p < 0.0001). (D) ALM phenotype of mec-7(u278) homozygotes, mec-7(u278) heterozygotes, and mec-7(u278)/mec-7(ky852) double heterozygotes. Data sets for (B–D) are based on an n of at least 100 animals.
Ectopic neurite outgrowth is triggered by hyperstable microtubules
Local stabilization of microtubules in immature neurites of dissociated neurons is sufficient to trigger axon formation (Witte et al., 2008). To investigate whether the ectopic processes in mec-7(ky852) animals were a result of hyperstable microtubules, we treated animals with a microtubule-destabilizing agent, colchicine, or a microtubule-stabilizing agent, paclitaxel. Colchicine is known to inhibit the assembly of microtubules by binding to tubulin, thereby disrupting microtubule dynamics (Fojo, 2008). Treatment of mec-7(ky852) animals with colchicine prevented outgrowth of the ectopic posterior process in the ALM neuron, leaving the anterior process intact (Figure 4A). To determine the developmental period during which colchicine produces its effect, we restricted treatment to the first 24 h after hatching (when the animals are largely in the L1/L2 stages) or to the following 24 h (during L3/L4). Approximately 70% of ALM neurons formed normally in mec-7(ky852) animals treated with colchicine during the first 24 h following hatching; in contrast, no rescue of the ALM posterior neurite was observed when the colchicine treatment was started 24 h after hatching (Figure 4B). The ability to specifically block development of the ALM ectopic process during the early L1 stage supports our observation that the defect emerges later than the normal ALM process, which is formed during embryogenesis. Notably, concentrations of colchicine that blocked development of the ALM ectopic process also affected neurite outgrowth of the AVM and PVM mechanosensory neurons in wild-type animals, both of which develop within the first 24 h after hatching, suggesting that the microtubules in mec-7(ky852) animals have similar sensitivity to colchicine to those of wild-type animals (i.e., the mutation does not appear to affect the ability of colchicine to bind to β-tubulin).
FIGURE 4:
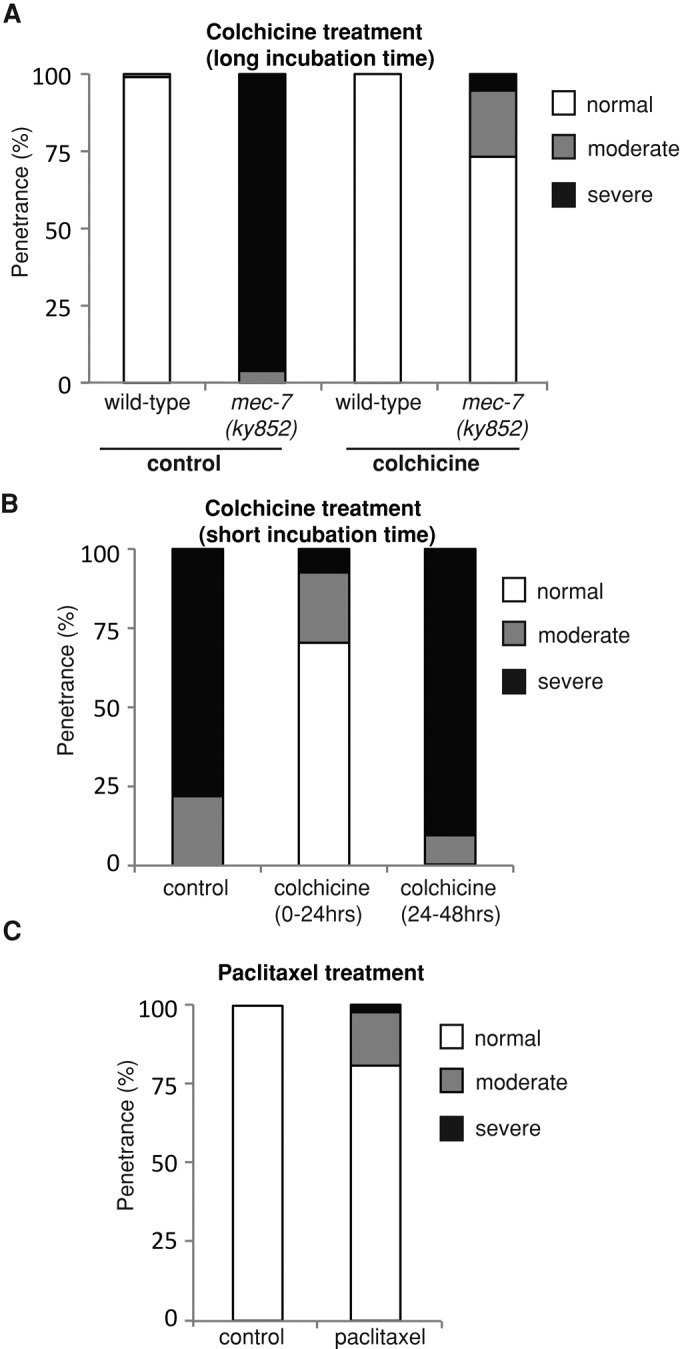
Manipulation of microtubule stability using colchicine and paclitaxel affects development of ectopic ALM neurites. (A) Rescue of ALM posterior neurite defects in mec-7(ky852) animals grown on colchicine throughout development. The distribution of ALM phenotypes in mec-7(ky852) animals grown on colchicine was significantly different from that of control mec-7(ky852) animals (chi-square test: p < 0.0001). (B) Rescue of ALM defects in mec-7(ky852) animals when treated with colchicine during the first 24 h but not the second 24 h after hatching. Animals treated with colchicine from 0 to 24 h were significantly different than controls (chi-square test: p < 0.0001). (C) Ectopic neurites generated in ALM neurons of wild-type (bus-17; zdIs5) animals grown on paclitaxel. ALM phenotypes of animals treated with paclitaxel were significantly different than controls (Fisher’s exact test: p < 0.03). Each data set is based on an n of at least 50 animals in (A) and (B) and 59 and 24 animals for paclitaxel-treated and control animals, respectively, in (C).
We next tested the effect of a microtubule-stabilizing agent, paclitaxel, on development of the mechanosensory neurons. Paclitaxel binds to microtubule polymers, stabilizing them by suppressing microtubule dynamics (Fojo, 2008). We hypothesized that pharmacological stabilization of microtubules would produce a similar phenotype to that of mec-7(ky852) mutants. bus-17(e2800) mutant animals were used for this experiment due to their thinner cuticles, which enhance the permeability of drugs (Bounoutas et al., 2009). In the absence of paclitaxel treatment, the mechanosensory neurons were normal in these animals, as visualized using the Pmec-4::GFP transgene (zdIs5; bus-17). However, treatment with paclitaxel during development induced the formation of ectopic processes, producing a phenotype that was similar to that of the mec-7(ky852) mutants, albeit to a lower penetrance (Figure 4C). This demonstrates that microtubule-stabilizing agents can induce ectopic neurite formation in vivo. Moreover, it suggests that the mec-7(ky852) phenotype could arise from excessive stabilization of microtubules, providing a dosage effect of hyperstable β-tubulin consistent with the semidominant nature of the mutation.
Ectopic outgrowth in mec-7(ky852) is suppressed by unc-33 and vab-8 mutations
Microtubule growth and stability is regulated by a variety of tubulin-binding molecules. To investigate whether β-tubulin could interact with such molecules to produce ectopic neurite outgrowth, we searched for suppressors of the mec-7(ky852) ALM phenotype. We first investigated the ptl-1 gene, which is the sole C. elegans homologue of the Tau/MAP2/MAP4 family of microtubule-associated proteins (MAPs) and promotes microtubule assembly (Goedert et al., 1996). A deletion mutation of the ptl-1 gene had no effect on the ALM outgrowth defects of mec-7(ky852) animals (Figure 5A and Table S1), suggesting that PTL-1 does not contribute to the mutant phenotype.
FIGURE 5:

Genetic interactions of mec-7(ky852) with MAPs and Wnts. (A) ALM phenotypes in double mutants of the microtubule-binding proteins, PTL-1/Tau and UNC-33/CRMP2, and the kinesin-like molecule VAB-8. mec-7(ky852); unc-33(e204) and mec-7(ky852); vab-8(e1017) double mutants have significantly more wild-type ALMs and less severe ALM defects compared with the mec-7(ky852) mutant (chi-square test: p < 0.001). (B) ALM phenotypes in mec-7(ky852); cwn-1; cwn-2 triple mutants. The mec-7(ky852) mutation converts the reversed unipolar neurons generated in cwn-1; cwn-2 double mutant animals into bipolar neurons. Each data set is based on an n of at least 100 animals.
Next we analyzed the C. elegans member of the collapsin response–mediated protein (CRMP) family, UNC-33. UNC-33 binds tubulin, promoting its assembly into microtubules, and has been shown to be necessary for axon outgrowth, microtubule structure, and polarized axonal transport (Hedgecock et al., 1985; Inagaki et al., 2001; Tsuboi et al., 2005; Maniar et al., 2012). We found that the unc-33(e204) mutation could partially suppress the mec-7(ky852) ALM outgrowth phenotype, as double mutants displayed a significant number of normal ALMs (never seen in mec-7(ky852) single mutants) and a dramatic reduction in the length of the ectopic process outgrowth (Figure 5A and Table S1). In contrast, the normal anterior process remained unaffected by the unc-33 mutation. This suggests that the ectopic outgrowth phenotype caused by the mutant MEC-7/β-tubulin could be caused by an enhanced or altered interaction with UNC-33, which may contribute to excessive growth of microtubules in these mutants.
An intriguing question is how a single mutation in MEC-7/β-tubulin can produce such precise, posterior outgrowth of a neuronal process. We hypothesized that this phenotype may be caused by interaction with VAB-8, a novel kinesin-like protein of C. elegans that is specifically required for posterior migration of both cells and growth cones (Wightman et al., 1996; Wolf et al., 1998; Lai and Garriga, 2004). Indeed, mutations in vab-8 could partially suppress the mec-7(ky852) ALM phenotype (Figure 5A and Table S1). Furthermore, several molecules that have been proposed to act downstream of VAB-8 in axon outgrowth, such as UNC-40 and UNC-73 (Levy-Strumpf and Culotti, 2007; Watari-Goshima et al., 2007) with its interacting partner CED-10/Rac (Steven et al., 1998; Spencer et al., 2001), could also partially suppress the ALM outgrowth phenotype (Table S1). Together these results suggest that VAB-8 and other downstream molecules interact with mutant MEC-7/β-tubulin to coordinate posterior neurite outgrowth.
Finally, we examined interactions with Wnt molecules, which act redundantly in ALM polarity (Hilliard and Bargmann, 2006; Prasad and Clark, 2006). Loss of specific combinations of Wnt molecules, such as in cwn-1 cwn-2 double mutants, results in the development of bipolar and reversed ALMs. Reversed ALMs remain monopolar but have an opposite orientation within the animal’s body, with their single process extending posteriorly toward the tail. We asked whether the mec-7(ky852) mutation was able to transform a reversed unipolar ALM into a bipolar ALM, thus indicating an independent mechanism of action. We found that in triple mutants mec-7(ky852) cwn-1 cwn-2, 100% of the reversed ALM neurons extended an ectopic anterior process (Figure 5B and Table S1), distinguishable from a normal ALM process, because it had not extended in the L1 and did not reach the nerve ring (unpublished data). This suggests that the function of mec-7(ky852) is preserved regardless of the orientation of the neuron, and is independent from this polarity effect of the Wnt mutations.
Mutant β-tubulin alters synaptic localization and axonal transport
Presynaptic loci are important elements that define the axonal identity of a neurite (Nonet, 1999; Hilliard and Bargmann, 2006; Mahoney et al., 2006; Ou et al., 2011). We investigated whether the ectopic posterior neurite in the ALM neuron of mec-7(ky852) mutants had axon-like properties, and whether the mutation could alter the normal distribution of presynaptic loci in the anterior process. In wild-type animals, ALM neurons have presynaptic sites located on a short branch of the anterior process that enters the nerve ring (Figure 6, A and E). Presynaptic densities can be labeled by the vesicle-associated Ras GTPase, RAB-3 (Nonet et al., 1997; Nonet, 1999; Mahoney et al., 2006). We used a Pmec-4::mCherry::rab-3 transgene to visualize the presynaptic sites specifically in the mechanosensory neurons of wild type and mec-7(ky852) mutants. A high percentage of mec-7(ky852) animals displayed mCherry::RAB-3 expression on the ectopic posterior process of the ALM neurons (Figure 6, B and E). The presence of the presynaptic marker on the ectopic process indicates that it has acquired an axon-like identity, indicating that the ALM neuron in the mec-7(ky852) animals has been changed from a cell with a single axon to a cell with two axon-like neurites. However, among those animals that ectopically expressed mCherry::RAB-3 on the posterior ALM process, approximately half also showed a loss of synapses from the normal anterior process of ALM, suggesting that the ky852 mutation might compromise the normal transport of vesicles to the anterior neurite. To determine whether this could occur in other neurons, we investigated the presynaptic distribution in the bipolar PLM neuron of mec-7(ky852) mutants. Interestingly, we found that, similar to ALM, the transport of presynaptic vesicles in PLM was altered in the mutants. In the PLM neurons of mutant animals, the RAB-3 presynaptic marker was mislocalized to the posterior neurite, with only a small percentage of selective localization on the anterior neurite, as in wild-type animals (Figure S3).
FIGURE 6:

Distribution of RAB-3 and UNC-104/kinesin in ALM neurons of mec-7(ky852) mutants. (A) Normal localization of the RAB-3::mCherry presynaptic marker in ALM neurons of wild-type animals. White arrowheads indicate RAB-3 in the nerve ring branch of ALM. Gray dashed circles represent coelomocytes expressing GFP that were used as a transgenic marker. (B) RAB-3::mCherry localizes to the posterior processes of ALM neurons in mec-7(ky852) mutants. White arrowheads indicate RAB-3 in the nerve ring branch and ectopic posterior process of ALM. (C) UNC-104::GFP localization in ALM neurons of wild-type animals. Localization of UNC-104 is indicated (white arrowheads). (D) UNC-104::GFP localizes to the posterior processes of ALM neurons in mec-7(ky852) mutants. (E) Quantification of RAB-3::mCherry localization on the anterior processes and on the posterior processes of ALM neurons. (F) Quantification of UNC-104::GFP localization on the anterior and posterior processes of ALM neurons. Each data set is based on an n of at least 50 animals. Scale bar: 25 μm.
Anterograde transport of synaptic vesicles along axonal microtubules is mediated by the UNC-104/KIF1A motor protein (Hall and Hedgecock, 1991). To investigate whether this machinery was compromised in the mec-7(ky852) animals, we examined its localization in the ALM mechanosensory neurons of wild-type animals and mec-7(ky852) mutants using a Pmec-4::unc-104::GFP transgene. In wild-type mechanosensory neurons, UNC-104::GFP is expressed throughout the cell body and axon and is strongly localized to presynaptic sites (Kumar et al., 2011; Figure 6C). In mec-7(ky852) mutants, UNC-104::GFP appeared to localize normally in the anterior ALM process; however, it was also enriched in the terminals of the ectopic posterior processes (Figure 6, D and F), indicating a failure to restrict transport to the anterior process. As expected, the ectopic UNC-104::GFP appeared to colocalize with RAB-3 (unpublished data). Together these results suggest that mec-7(ky852) animals have mislocalized transport of synaptic vesicles into the ectopic posterior processes, which gives them an axon-like identity.
Axonal regeneration is inhibited in mec-7(ky852) mutants
The likelihood that an axon will regrow following injury has been closely linked to the presence of presynaptic loci, as axons regenerate poorly when they are severed distally from presynaptic sites (Wu et al., 2007). Our observation that mec-7(ky852) animals displayed mislocalization of the RAB-3 presynaptic marker suggested that this might influence their capacity for axonal regeneration. To test whether this was the case, we used laser axotomy to sever the axons of ALM neurons in wild-type and mutant animals and then quantified the level of regeneration. As previously shown by us and by others, ALM presents a high rate of regeneration following injury, with the regrowing proximal axon extending toward the distal fragment and often reconnecting with it by means of axonal fusion, which restores the axonal tract (Yanik et al., 2006; Wu et al., 2007; Ghosh-Roy et al., 2011; Neumann et al., 2011). Interestingly, regeneration of both the anterior and posterior neurites was reduced in mec-7(ky852) mutants (Figure 7). mec-7(ky852) animals displayed a reduced capacity to initiate axon regrowth (Figure 7A); however, axon extension was normal, as the average length of regrowth in animals that could initiate regeneration was similar to that of wild-type animals (Figure 7B). Furthermore, regrowing ALM neurites in mec-7(ky852) mutants displayed a reduced rate of axonal reconnection and fusion compared with wild-type animals (Figure 7, C and D). To determine whether these defects also extended to other neurons, we tested the regenerative capacity of PLM. Neurite regeneration was also altered in this cell, similar to ALM, with strongly reduced initiation of regrowth and reduced fusion (Figure S4). These results suggest that the mec-7(ky852) mutation causes defects in multiple aspects of axonal regeneration, including regrowth initiation and reconnection with separated distal fragments.
FIGURE 7:

Axonal regeneration defects in mec-7(ky852) mutants. Regeneration of the anterior ALM process in wild-type animals and both the anterior and posterior ALM processes in mutants is shown in each graph. (A) Percentage of ALM neurons showing regrowth following axotomy in wild-type and mec-7(ky852) animals. (B) Quantification of the average length of regrowth of regenerating axons shown in (A). (C and D) Percentage of ALM neurons showing reconnection of the regenerating process to the distal fragment, and of these reconnected axons, the percentage showing maintenance of the connection (fusion). (E and F) Images showing axonal regeneration in a wild-type animal (E) and defective axonal regeneration a mec-7(ky852) mutant (F). In wild-type animals, the regenerating axon regrows from the proximal fragment (closest to the cell body) to the distal fragment of the ALM neuron. n for each data set is indicated on the bars of the graph. Error bars represent the SE of proportion. *, significant difference between wild-type and mutant animals (Student’s t test: p < 0.05).
DISCUSSION
Studies in cultured neurons have demonstrated that regulation of microtubule dynamics plays a crucial role in determining whether an immature neurite develops into an axon. Nevertheless, evidence that this can occur in vivo has been lacking. Witte et al. (2008) recently demonstrated that local stabilization of microtubules in cultured hippocampal neurons is sufficient to induce axon formation, and suggested that microtubules play an active, instructive role in specifying axonal identity. Our study provides evidence that a single point mutation in the mec-7/β-tubulin gene can lead to outgrowth of secondary axon-like neurites. Interestingly, we find that the same mutation inhibits axonal regeneration in these neurons. Together our results indicate that the mutant form of β-tubulin affects multiple aspects of neuronal development and repair, and suggest that there are critical differences between axonal growth and regeneration.
We have identified a novel allele of mec-7/β-tubulin that causes an unusual polarity phenotype in the ALM mechanosensory neurons, inducing the outgrowth of an additional neurite, which transforms them into bipolar neurons. The precision of this phenotype is remarkable: the neurons in the mutant animals are consistently bipolar (rather than multipolar), and the long ectopic processes always grow in the posterior direction. Such precise ectopic neurite outgrowth along the anterior–posterior axis has also been observed in the six C. elegans VC motor neurons (VC1–6), suggesting the involvement of common factors (Sanchez-Alvarez et al., 2011). These phenotypes may be explained by the presence of extracellular factors, or intrinsic cell signals, that bias the direction of outgrowth. A number of studies have identified mutations in guidance cues, receptors, and intracellular proteins that cause ALM polarity defects (Wolf et al., 1998; Hilliard and Bargmann, 2006; Levy-Strumpf and Culotti, 2007; Watari-Goshima et al., 2007; Pan et al., 2008). However, the phenotypes observed in these studies differed from those in the mec-7(ky852) animals, as they predominantly showed complete inversion of ALM polarity and were generated during the embryonic development of the ALM neuroblast (Hilliard and Bargmann, 2006), rather than postembryonically. Finally, our finding that inverted ALM neurons caused by Wnt mutations were transformed into bipolar neurons in a mec-7(ky852) mutant background suggests that the outgrowth of the mec-7(ky852) ectopic process is mediated by an intrinsic cell bias, which may operate independently of neuronal polarity established by Wnts. Our study suggests that the kinesin-like protein,VAB-8, recruits molecules required for the posterior neurite outgrowth seen in the mec-7(ky852) mutants. Whether VAB-8 binds directly to tubulin remains to be investigated.
In cultured cells, neuronal polarization is considered to occur when a preexisting immature neurite transforms into an axon (Polleux and Snider, 2010; Namba et al., 2011). In such cases, the number of potential axons and their directions of outgrowth are dependent on the location and number of immature neurites. In contrast, C. elegans ALM neurons usually lack secondary neurites, with only a small proportion of animals presenting a short posterior protrusion, which may be indicative of a preexisting intrinsic cell bias. Thus the ALM ectopic posterior process observed in the mec-7(ky852) mutants usually emerges directly from the cell body. This suggests that mutations in mec-7/β-tubulin can alter the entire process of axon formation in vivo, from the initial emergence of the neurite to its full extension into an axon-like process containing presynaptic sites.
Our results suggest that microtubule stabilization contributes to the regulation of axon outgrowth in vivo. First, application of paclitaxel to wild-type animals was sufficient to trigger the outgrowth of ectopic processes. Second, colchicine treatment could rescue the ectopic ALM outgrowth in mec-7(ky852) mutants, suggesting that microtubules could mitigate the effect of the mutation. Whether the ky852 mutation can specifically confer microtubule hyperstability remains unclear. One possibility is that it might alter the structure of tubulin polymers, such that they interfere with normal microtubule dynamics, slowing or preventing microtubule breakdown. Alternatively, the mutation may alter the binding or transport of molecules that regulate microtubule stability. Our finding that UNC-33/CRMP could suppress the mec-7(ky852) ALM phenotype suggests that the binding of this molecule, which has been shown to regulate microtubule structure, dynamics, and transport (Fukata et al., 2002; Maniar et al., 2012), may contribute to excessive axon outgrowth, consistent with the results of overexpression studies in hippocampal neurons (Inagaki et al., 2001).
The ectopic localization of the RAB-3 presynaptic marker and the UNC-104/KIF1A motor protein in the posterior ALM processes of mec-7(ky852) animals suggests that the mutant microtubules affect normal intracellular transport. Notably, defects in synapse and axon outgrowth have recently been observed in C. elegans mutants of other tubulin subunits (Baran et al., 2010), further indicating that the nature of microtubules and their potential interactions with motor proteins is crucial for normal axonal transport. Intracellular transport of proteins is dependent on the intrinsic polarity of microtubules, which have dynamic plus ends that undergo rapid growth or collapse, and more stable minus ends (Howard and Hyman, 2003). These polarized polymers serve as one-way conduits for intracellular trafficking by plus end–directed or minus end–directed motor proteins (Baas and Lin, 2011) and are thus crucial for axon development and maintenance. Interestingly, UNC-33 is required for polarization of microtubule subunits and directed transport of synaptic proteins into the axon of PVD neurons (Maniar et al., 2012). Polarized transport has also been shown to be essential for correct synapse localization to the axon of the C. elegans DA9 motor neuron and for transport of polarity proteins into the axon in cultured neurons (Nishimura et al., 2004; Shi et al., 2004; Ou et al., 2011). The plus end–directed kinesin-1 motor domain has been reported to selectively accumulate in a neurite prior to its specification as an axon (Jacobson et al., 2006). Therefore it is possible that intrinsic microtubule polarity could provide an initial cue for axon formation by promoting movement of cargo by axon-specific kinesins into the developing axon.
The altered transport of synaptic proteins in mec-7(ky852) mutants may also be a cause of their axonal regeneration defects. Following injury, rapid signaling must occur between the site of injury and the cell body in order to initiate regeneration (Abe and Cavalli, 2008). This is reliant on the transport of proteins, as impairment of axonal transport mechanisms has been found to affect regrowth of PLM in C. elegans (Chen et al., 2011). Therefore it is possible that mec-7(ky852) regeneration defects are partly due to a failure of axonal transport following injury. Another possible explanation is that regeneration may be triggered only when the cell body detects a loss of synapses. This hypothesis is consistent with the findings of Wu et al. (2007), who showed that axons of C. elegans mechanosensory neurons severed distally to the presynaptic sites (i.e., with presynaptic sites still connected to the cell body) fail to regenerate and, furthermore, that regeneration can be rescued by severing the branch containing presynaptic sites. Thus, it is possible that in mec-7(ky852) mutants, which often lack presynaptic sites in the axons, the cell fails to detect a loss of synapses following severing of its axon and consequently fails to signal for regeneration to occur. Future studies are required to determine whether the defects in axonal regeneration observed in these mutants are caused by impairment of axonal transport or by alterations to the synaptic sites, or whether an alternative mechanism is in place.
Although emerging evidence suggests that the regulation of microtubule dynamics is a crucial determinant of axonal regenerative capacity (Erturk et al., 2007; Stone et al., 2010; Chen et al., 2011; Hellal et al., 2011; Sengottuvel and Fischer, 2011), a direct role for β-tubulin in regeneration has not been shown. Regeneration of an axon requires drastic remodeling of cell shape, which relies on dynamic instability of the cytoskeleton. An axon must first sprout a new process with a growth cone at its tip; this must then extend in the correct direction to reconnect with its target tissue. Interestingly, although regrowth initiation and reconnection to the distal process are inhibited in the mec-7(ky852) mutants, there are no apparent deficits in extension of regrowing axons. It is therefore likely that regrowth initiation and reconnection by axonal fusion require comparatively greater structural changes to the cytoskeleton than axon extension. Thus mec-7(ky852) mutations may differentially affect distinct stages of regeneration, depending on their specific requirements for dynamic microtubules.
This study of a C. elegans β-tubulin mutant has revealed a central role for microtubules in axon outgrowth and regeneration in vivo. Our findings contribute to growing evidence that precise microtubule dynamics are essential for neuronal development, maintenance, and repair. Mutations in human tubulin have been found to cause neurodevelopmental and neurodegenerative diseases (Kuijpers and Hoogenraad, 2011; Tischfield et al., 2011). As β-tubulin is highly conserved across species, understanding its function in model organisms has the potential to provide important insights into microtubule-related neuropathies in humans.
MATERIALS AND METHODS
Strains
Nematodes were cultured under standard conditions (Brenner, 1974). All experiments were performed at 22°C on hermaphrodite animals. The Hawaiian CB4856 strain was used for SNP mapping of the mec-7(ky852) mutation (Wicks et al., 2001; Davis et al., 2005; Davis and Hammarlund, 2006). The wild-type Bristol N2 strain and the following mutations were used: LGX: mec-7(ky852), mec-7(u278); LGI: unc-73(e936), mig-2(mu28), unc-40(e1430); LGII: cwn-1(ok546); LGIII: mig-10(ct41), ptl-1(ok621); LGIV: ced-10(n3246), unc-33(e204), cwn-2(ok895); and LGV: vab-8(e1017). Transgenes used were: zdIs5[Pmec-4::GFP]; juIs76[Punc-25::GFP]; vdEx262[Pmec-4::mCherry::rab-3 (0.5 ng/μl), Punc-122::GFP (25 ng/μl)]; vdEx309[Pmec-4::mec-7 (5 ng/μl), Podr-1::dsRed (30 ng/μl)]; vdEx312[Pmec-4::mec-7 (10 ng/μl), Podr-1::dsRed (30 ng/μl)]; jsIs1111[Pmec-4::unc-104::GFP]; vdEx263[(Pmec-4::mCherry (5 ng/μl), Podr-1::dsRed (30 ng/μl)]; vdEx326 [Pmec-4::mec-7(ky852) (5 ng/μl), Podr-1::dsRed (30 ng/μl)]; vdEx329[Pmec-4::mec-7(ky852)(10 ng/μl), Podr-1::dsRed (30 ng/μl)]. The jsIs1111 strain was a kind gift from Sandhya Koushika (Tata Institute of Fundamental Research, Bangalore, India).
Molecular biology
Standard molecular biology techniques were used (Sambrook and Russell, 2001). The Pmec-4::mec-7::unc-54 3′UTR plasmid was generated through excision of the GFP coding sequences from the Pmec-4::GFP plasmid and subsequent insertion of the entire mec-7 coding sequence (amplified from N2 DNA, with the following primers: forward, 5′-tcagtgggatccaagcaacatgcgcgagatcg-3′; reverse, 5′-tcagtgggatccatttactctccgtcgaacgc-3′). The Pmec-4::mec-7(ky852)::unc-54 3′UTR plasmid was generated by QuikChange Site-Directed Mutagenesis (Agilent, Santa Clara, CA) using the Pmec-4::mec-7::unc-54 3′UTR as template DNA and the following primers to introduce the ky852 mutation: sense, 5′-cactctgaaactgacgacatcgacatatggagacc-3′; antisense, 5′-ggtctccatatgtcgatgtcgtcagtttcagagtg-3′. The Pmec-4::mCherry::RAB-3::unc-54 3′UTR plasmid was created through modification of the Pmec-4::GFP plasmid. First, the coding sequence for GFP was excised and replaced with that of mCherry. Next the rab-3 coding sequence, which was amplified from the Prab-3::eGFP::RAB-3::rim-3 3′UTR plasmid (a kind gift from Michael Nonet, St. Louis, MO), was inserted downstream from mCherry.
Phenotypic analyses
ALM morphology was analyzed at the L4 or adult stage in a zdIs5(Pmec-4::GFP) background. Although ALM neurons in wild-type zdIs5 animals are usually monopolar, some have a small posterior protrusion that is less than two cell bodies in length. The ALM posterior process defect in mec-7(ky852) animals was considered to be moderate when the process was between 2 and 15 cell bodies in length, and severe when greater than 15 cell bodies in length. AVM, PVM, and PLM mechanosensory neurons were also analyzed for axon outgrowth and overextension phenotypes in L4 and adult animals.
Developmental analysis of ALM morphology in the mec-7(ky852) mutant was performed in embryos and in synchronized populations of hatched animals. Animals were synchronized by collecting newly hatched larvae from a plate containing only eggs. Newly hatched L1 animals were either scored immediately or transferred to new plates and grown on food for 5 h or until adulthood before scoring. In embryos and L1 animals, ALM neurons were scored as defective if a posterior process was observed (in wild-type animals, posterior processes were never observed at these early stages).
Microscopy and laser axotomy
Animals were immobilized in 0.01–0.05% tetramisole hydrochloride on 4% agar pads and visualized using Zeiss Axioimager Z1 and Zeiss Axioimager A1 microscopes (Jena, Germany). Images were taken using a Photometrics camera (Cool Snap HQ2; Tucson, AZ) and analyzed with MetaMorph software (Molecular Devices, Sunnyvale, CA). Individual planes from Z-stacks were combined in Adobe Photoshop (San Jose, CA). Axotomies were performed as previously described (Neumann et al., 2011). A MicroPoint laser system basic unit (Andor, Belfast, UK) was used to perform axonal transection on L4 animals at a point ∼50 μm anterior to the ALM and PLM cell bodies. For analysis of regeneration in the posterior ALM axon in mec-7(ky852) animals, axotomy was performed ∼50 μm posterior to the cell body. Axonal regrowth was quantified by measuring the length of the longest neurite beyond the cut site 24 h postaxotomy; neurons that underwent axonal fusion were excluded from these quantifications.
Colchicine and paclitaxel treatment
Animals were grown on nematode growth medium agar plates containing colchicine (Sigma-Aldrich, St. Louis, MO) or paclitaxel (Sigma-Aldrich) with dimethyl sulfoxide, as previously described (Chalfie and Thomson, 1982).
Statistical analyses
Primer of Biostatistics 3.01 and GraphPad Prism (La Jolla, CA) were used for statistical analyses. Error of proportions was used to estimate variation within a single population. The Student’s t test or Mann-Whitney rank sum test were used as indicated. The chi-square test was performed to estimate variance in the distribution of phenotypes between wild-type and mutant populations.
Supplementary Material
Acknowledgments
We thank Cori Bargmann, in whose lab this work started; Martin Chalfie, Sandhya Koushika, Michael Nonet, Nick Valmas, and Casey Linton for sharing reagents; Paolo Bazzicalupo, Rowan Tweedale, Helen Cooper, and members of the Hilliard Lab for helpful discussion and comments on the manuscript; and Luke Hammond and Paula Mugno for technical assistance. Nematode strains used in this work were provided by the Caenorhabditis Genetics Center, which is funded by the National Institutes of Health (NIH) National Center for Research Resources, and the International C. elegans Gene Knockout Consortium. This work was funded by the National Health and Medical Research Council of Australia (NHMRC) and the NIH and was supported by the Queensland Brain Institute.
Abbreviations used:
- CRMP
collapsin response–mediated protein
- GFP
green fluorescent protein
- MAP
microtubule-associated protein
- SNP
single-nucleotide polymorphism
Footnotes
This article was published online ahead of print in MBoC in Press (http://www.molbiolcell.org/cgi/doi/10.1091/mbc.E12-06-0441) on December 5, 2012.
The authors declare no conflict of interest.
REFERENCES
- Abe N, Cavalli V. Nerve injury signaling. Curr Opin Neurobiol. 2008;18:276–283. doi: 10.1016/j.conb.2008.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adler CE, Fetter RD, Bargmann CI. UNC-6/Netrin induces neuronal asymmetry and defines the site of axon formation. Nat Neurosci. 2006;9:511–518. doi: 10.1038/nn1666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baas PW. Neuronal polarity: microtubules strike back. Nat Cell Biol. 2002;4:E194–E195. doi: 10.1038/ncb0802-e194. [DOI] [PubMed] [Google Scholar]
- Baas PW, Lin S. Hooks and comets: the story of microtubule polarity orientation in the neuron. Dev Neurobiol. 2011;71:403–418. doi: 10.1002/dneu.20818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baran R, Castelblanco L, Tang G, Shapiro I, Goncharov A, Jin Y. Motor neuron synapse and axon defects in a C. elegans alpha-tubulin mutant. PLoS One. 2010;5:e9655. doi: 10.1371/journal.pone.0009655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bounoutas A, O’Hagan R, Chalfie M. The multipurpose 15-protofilament microtubules in C. elegans have specific roles in mechanosensation. Curr Biol. 2009;19:1362–1367. doi: 10.1016/j.cub.2009.06.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradke F, Dotti CG. The role of local actin instability in axon formation. Science. 1999;283:1931–1934. doi: 10.1126/science.283.5409.1931. [DOI] [PubMed] [Google Scholar]
- Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974;77:71–94. doi: 10.1093/genetics/77.1.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chalfie M, Sulston J. Developmental genetics of the mechanosensory neurons of Caenorhabditis elegans. Dev Biol. 1981;82:358–370. doi: 10.1016/0012-1606(81)90459-0. [DOI] [PubMed] [Google Scholar]
- Chalfie M, Thomson JN. Structural and functional diversity in the neuronal microtubules of Caenorhabditis elegans. J Cell Biol. 1982;93:15–23. doi: 10.1083/jcb.93.1.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang C, Adler CE, Krause M, Clark SG, Gertler FB, Tessier-Lavigne M, Bargmann CI. MIG-10/lamellipodin and AGE-1/PI3K promote axon guidance and outgrowth in response to slit and netrin. Curr Biol. 2006;16:854–862. doi: 10.1016/j.cub.2006.03.083. [DOI] [PubMed] [Google Scholar]
- Chen L, Wang Z, Ghosh-Roy A, Hubert T, Yan D, O’Rourke S, Bowerman B, Wu Z, Jin Y, Chisholm AD. Axon regeneration pathways identified by systematic genetic screening in C. elegans. Neuron. 2011;71:1043–1057. doi: 10.1016/j.neuron.2011.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis MW, Hammarlund M. Single-nucleotide polymorphism mapping. Methods Mol Biol. 2006;351:75–92. doi: 10.1385/1-59745-151-7:75. [DOI] [PubMed] [Google Scholar]
- Davis MW, Hammarlund M, Harrach T, Hullett P, Olsen S, Jorgensen EM. Rapid single nucleotide polymorphism mapping in C. elegans. BMC Genomics. 2005;6:118. doi: 10.1186/1471-2164-6-118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erturk A, Hellal F, Enes J, Bradke F. Disorganized microtubules underlie the formation of retraction bulbs and the failure of axonal regeneration. J Neurosci. 2007;27:9169–9180. doi: 10.1523/JNEUROSCI.0612-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fojo T. The Role of Microtubules in Cell Biology, Neurobiology, and Oncology. Totowa, NJ: Humana; 2008. [Google Scholar]
- Fukata Y, et al. CRMP-2 binds to tubulin heterodimers to promote microtubule assembly. Nat Cell Biol. 2002;4:583–591. doi: 10.1038/ncb825. [DOI] [PubMed] [Google Scholar]
- Ghosh-Roy A, Wu Z, Goncharov A, Jin Y, Chisholm AD. Calcium and cyclic AMP promote axonal regeneration in Caenorhabditis elegans and require DLK-1 kinase. J Neurosci. 2011;30:3175–3183. doi: 10.1523/JNEUROSCI.5464-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goedert M, Baur CP, Ahringer J, Jakes R, Hasegawa M, Spillantini MG, Smith MJ, Hill F. PTL-1, a microtubule-associated protein with tau-like repeats from the nematode Caenorhabditis elegans. J Cell Sci. 1996;109:2661–2672. doi: 10.1242/jcs.109.11.2661. [DOI] [PubMed] [Google Scholar]
- Hall DH, Hedgecock EM. Kinesin-related gene unc-104 is required for axonal transport of synaptic vesicles in C. elegans. Cell. 1991;65:837–847. doi: 10.1016/0092-8674(91)90391-b. [DOI] [PubMed] [Google Scholar]
- Hamelin M, Scott IM, Way JC, Culotti JG. The mec-7 β-tubulin gene of Caenorhabditis elegans is expressed primarily in the touch receptor neurons. EMBO J. 1992;11:2885–2893. doi: 10.1002/j.1460-2075.1992.tb05357.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedgecock EM, Culotti JG, Thomson JN, Perkins LA. Axonal guidance mutants of Caenorhabditis elegans identified by filling sensory neurons with fluorescein dyes. Dev Biol. 1985;111:158–170. doi: 10.1016/0012-1606(85)90443-9. [DOI] [PubMed] [Google Scholar]
- Hellal F, et al. Microtubule stabilization reduces scarring and causes axon regeneration after spinal cord injury. Science. 2011;331:928–931. doi: 10.1126/science.1201148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hilliard MA, Bargmann CI. Wnt signals and frizzled activity orient anterior-posterior axon outgrowth in C. elegans. Dev Cell. 2006;10:379–390. doi: 10.1016/j.devcel.2006.01.013. [DOI] [PubMed] [Google Scholar]
- Hoogenraad CC, Bradke F. Control of neuronal polarity and plasticity—a renaissance for microtubules. Trends Cell Biol. 2009;19:669–676. doi: 10.1016/j.tcb.2009.08.006. [DOI] [PubMed] [Google Scholar]
- Howard J, Hyman AA. Dynamics and mechanics of the microtubule plus end. Nature. 2003;422:753–758. doi: 10.1038/nature01600. [DOI] [PubMed] [Google Scholar]
- Inagaki N, Chihara K, Arimura N, Menager C, Kawano Y, Matsuo N, Nishimura T, Amano M, Kaibuchi K. CRMP-2 induces axons in cultured hippocampal neurons. Nat Neurosci. 2001;4:781–782. doi: 10.1038/90476. [DOI] [PubMed] [Google Scholar]
- Jacobson C, Schnapp B, Banker GA. A change in the selective translocation of the kinesin-1 motor domain marks the initial specification of the axon. Neuron. 2006;49:797–804. doi: 10.1016/j.neuron.2006.02.005. [DOI] [PubMed] [Google Scholar]
- Jiang H, Guo W, Liang X, Rao Y. Both the establishment and the maintenance of neuronal polarity require active mechanisms: critical roles of GSK-3β and its upstream regulators. Cell. 2005;120:123–135. doi: 10.1016/j.cell.2004.12.033. [DOI] [PubMed] [Google Scholar]
- Kishi M, Pan YA, Crump JG, Sanes JR. Mammalian SAD kinases are required for neuronal polarization. Science. 2005;307:929–932. doi: 10.1126/science.1107403. [DOI] [PubMed] [Google Scholar]
- Kuijpers M, Hoogenraad CC. Centrosomes, microtubules and neuronal development. Mol Cell Neurosci. 2011;48:349–358. doi: 10.1016/j.mcn.2011.05.004. [DOI] [PubMed] [Google Scholar]
- Kumar J, Choudhary BC, Metpally R, Zheng Q, Nonet ML, Ramanathan S, Klopfenstein DR, Koushika SP. The Caenorhabditis elegans kinesin-3 motor UNC-104/KIF1A is degraded upon loss of specific binding to cargo. PLoS Genet. 2011;6:e1001200. doi: 10.1371/journal.pgen.1001200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai T, Garriga G. The conserved kinase UNC-51 acts with VAB-8 and UNC-14 to regulate axon outgrowth in C. elegans. Development. 2004;131:5991–6000. doi: 10.1242/dev.01457. [DOI] [PubMed] [Google Scholar]
- Levy-Strumpf N, Culotti JG. VAB-8, UNC-73 and MIG-2 regulate axon polarity and cell migration functions of UNC-40 in C. elegans. Nat Neurosci. 2007;10:161–168. doi: 10.1038/nn1835. [DOI] [PubMed] [Google Scholar]
- Mahoney TR, Liu Q, Itoh T, Luo S, Hadwiger G, Vincent R, Wang ZW, Fukuda M, Nonet ML. Regulation of synaptic transmission by RAB-3 and RAB-27 in Caenorhabditis elegans. Mol Biol Cell. 2006;17:2617–2625. doi: 10.1091/mbc.E05-12-1170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maniar TA, Kaplan M, Wang GJ, Shen K, Wei L, Shaw JE, Koushika SP, Bargmann CI. UNC-33 (CRMP) and ankyrin organize microtubules and localize kinesin to polarize axon-dendrite sorting. Nat Neurosci. 2012;15:48–56. doi: 10.1038/nn.2970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Namba T, Nakamuta S, Funahashi Y, Kaibuchi K. The role of selective transport in neuronal polarization. Dev Neurobiol. 2011;71:445–457. doi: 10.1002/dneu.20876. [DOI] [PubMed] [Google Scholar]
- Neukirchen D, Bradke F. Neuronal polarization and the cytoskeleton. Semin Cell Dev Biol. 2011;22:825–833. doi: 10.1016/j.semcdb.2011.08.007. [DOI] [PubMed] [Google Scholar]
- Neumann B, Nguyen KC, Hall DH, Ben-Yakar A, Hilliard MA. Axonal regeneration proceeds through specific axonal fusion in transected C. elegans neurons. Dev Dyn. 2011;240:1365–1372. doi: 10.1002/dvdy.22606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura T, Kato K, Yamaguchi T, Fukata Y, Ohno S, Kaibuchi K. Role of the PAR-3-KIF3 complex in the establishment of neuronal polarity. Nat Cell Biol. 2004;6:328–334. doi: 10.1038/ncb1118. [DOI] [PubMed] [Google Scholar]
- Nonet ML. Visualization of synaptic specializations in live C. elegans with synaptic vesicle protein-GFP fusions. J Neurosci Methods. 1999;89:33–40. doi: 10.1016/s0165-0270(99)00031-x. [DOI] [PubMed] [Google Scholar]
- Nonet ML, Staunton JE, Kilgard MP, Fergestad T, Hartwieg E, Horvitz HR, Jorgensen EM, Meyer BJ. Caenorhabditis elegans rab-3 mutant synapses exhibit impaired function and are partially depleted of vesicles. J Neurosci. 1997;17:8061–8073. doi: 10.1523/JNEUROSCI.17-21-08061.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ou CY, et al. Two cyclin-dependent kinase pathways are essential for polarized trafficking of presynaptic components. Cell. 2011;141:846–858. doi: 10.1016/j.cell.2010.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan CL, Baum PD, Gu M, Jorgensen EM, Clark SG, Garriga G. AP-2 and retromer control Wnt signaling by regulating mig-14/Wntless. Dev Cell. 2008;14:132–139. doi: 10.1016/j.devcel.2007.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polleux F, Snider W. Initiating and growing an axon. Cold Spring Harb Perspect Biol. 2010;2:a001925. doi: 10.1101/cshperspect.a001925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasad BC, Clark SG. Wnt signaling establishes anteroposterior neuronal polarity and requires retromer in C. elegans. Development. 2006;133:1757–1766. doi: 10.1242/dev.02357. [DOI] [PubMed] [Google Scholar]
- Quinn CC, Pfeil DS, Wadsworth WG. CED-10/Rac1 mediates axon guidance by regulating the asymmetric distribution of MIG-10/lamellipodin. Curr Biol. 2008;18:808–813. doi: 10.1016/j.cub.2008.04.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J, Russell DW. Molecular Cloning. 3rd. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory; 2001. [Google Scholar]
- Sanchez-Alvarez L, Visanuvimol J, McEwan A, Su A, Imai JH, Colavita A. VANG-1 and PRKL-1 cooperate to negatively regulate neurite formation in Caenorhabditis elegans. PLoS Genet. 2011;7:e1002257. doi: 10.1371/journal.pgen.1002257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savage C, Hamelin M, Culotti JG, Coulson A, Albertson DG, Chalfie M. mec-7 is a β-tubulin gene required for the production of 15-protofilament microtubules in Caenorhabditis elegans. Genes Dev. 1989;3:870–881. doi: 10.1101/gad.3.6.870. [DOI] [PubMed] [Google Scholar]
- Savage C, Xue Y, Mitani S, Hall D, Zakhary R, Chalfie M. Mutations in the Caenorhabditis elegans β-tubulin gene mec-7: effects on microtubule assembly and stability and on tubulin autoregulation. J Cell Sci. 1994;107:2165–2175. doi: 10.1242/jcs.107.8.2165. [DOI] [PubMed] [Google Scholar]
- Sengottuvel V, Fischer D. Facilitating axon regeneration in the injured CNS by microtubule stabilization. Commun Integr Biol. 2011;4:391–393. doi: 10.4161/cib.4.4.15552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi SH, Cheng T, Jan LY, Jan YN. APC and GSK-3β are involved in mPar3 targeting to the nascent axon and establishment of neuronal polarity. Curr Biol. 2004;14:2025–2032. doi: 10.1016/j.cub.2004.11.009. [DOI] [PubMed] [Google Scholar]
- Spencer AG, Orita S, Malone CJ, Han M. A RHO GTPase-mediated pathway is required during P cell migration in Caenorhabditis elegans. Proc Natl Acad Sci USA. 2001;98:13132–13137. doi: 10.1073/pnas.241504098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steven R, Kubiseski TJ, Zheng H, Kulkarni S, Mancillas J, Ruiz Morales A, Hogue CW, Pawson T, Culotti J. UNC-73 activates the Rac GTPase and is required for cell and growth cone migrations in C. elegans. Cell. 1998;92:785–795. doi: 10.1016/s0092-8674(00)81406-3. [DOI] [PubMed] [Google Scholar]
- Stiess M, Bradke F. Neuronal polarization: the cytoskeleton leads the way. Dev Neurobiol. 2011;71:430–444. doi: 10.1002/dneu.20849. [DOI] [PubMed] [Google Scholar]
- Stone MC, Nguyen MM, Tao J, Allender DL, Rolls MM. Global up-regulation of microtubule dynamics and polarity reversal during regeneration of an axon from a dendrite. Mol Biol Cell. 2010;21:767–777. doi: 10.1091/mbc.E09-11-0967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tischfield MA, Cederquist GY, Gupta ML, Jr., Engle EC. Phenotypic spectrum of the tubulin-related disorders and functional implications of disease-causing mutations. Curr Opin Genet Dev. 2011;21:286–294. doi: 10.1016/j.gde.2011.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsuboi D, Hikita T, Qadota H, Amano M, Kaibuchi K. Regulatory machinery of UNC-33 Ce-CRMP localization in neurites during neuronal development in Caenorhabditis elegans. J Neurochem. 2005;95:1629–1641. doi: 10.1111/j.1471-4159.2005.03490.x. [DOI] [PubMed] [Google Scholar]
- Watari-Goshima N, Ogura K, Wolf FW, Goshima Y, Garriga G. C. elegans VAB-8 and UNC-73 regulate the SAX-3 receptor to direct cell and growth-cone migrations. Nat Neurosci. 2007;10:169–176. doi: 10.1038/nn1834. [DOI] [PubMed] [Google Scholar]
- White JG. The structure of the nervous system of the nematode C. elegans. Philos Trans R Soc Lond B Biol Sci. 1986;314:1–340. doi: 10.1098/rstb.1986.0056. [DOI] [PubMed] [Google Scholar]
- Wicks SR, Yeh RT, Gish WR, Waterston RH, Plasterk RH. Rapid gene mapping in Caenorhabditis elegans using a high density polymorphism map. Nat Genet. 2001;28:160–164. doi: 10.1038/88878. [DOI] [PubMed] [Google Scholar]
- Wightman B, Clark SG, Taskar AM, Forrester WC, Maricq AV, Bargmann CI, Garriga G. The C. elegans gene vab-8 guides posteriorly directed axon outgrowth and cell migration. Development. 1996;122:671–682. doi: 10.1242/dev.122.2.671. [DOI] [PubMed] [Google Scholar]
- Witte H, Bradke F. The role of the cytoskeleton during neuronal polarization. Curr Opin Neurobiol. 2008;18:479–487. doi: 10.1016/j.conb.2008.09.019. [DOI] [PubMed] [Google Scholar]
- Witte H, Neukirchen D, Bradke F. Microtubule stabilization specifies initial neuronal polarization. J Cell Biol. 2008;180:619–632. doi: 10.1083/jcb.200707042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolf FW, Hung MS, Wightman B, Way J, Garriga G. vab-8 is a key regulator of posteriorly directed migrations in C. elegans and encodes a novel protein with kinesin motor similarity. Neuron. 1998;20:655–666. doi: 10.1016/s0896-6273(00)81006-5. [DOI] [PubMed] [Google Scholar]
- Wu Z, Ghosh-Roy A, Yanik MF, Zhang JZ, Jin Y, Chisholm AD. Caenorhabditis elegans neuronal regeneration is influenced by life stage, ephrin signaling, and synaptic branching. Proc Natl Acad Sci USA. 2007;104:15132–15137. doi: 10.1073/pnas.0707001104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yanik MF, Cinar H, Cinar HN, Chisholm A, Jin Y, Yakar AB. Nerve regeneration in C. elegans after femtosecond laser axotomy. IEEE J Sel Top Quant. 2006:1283–1291. [Google Scholar]
- Yoshimura T, Kawano Y, Arimura N, Kawabata S, Kikuchi A, Kaibuchi K. GSK-3β regulates phosphorylation of CRMP-2 and neuronal polarity. Cell. 2005;120:137–149. doi: 10.1016/j.cell.2004.11.012. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.