

Abstract
The ability to predict how another individual will behave is useful in social competition. Chimpanzees can predict the behaviour of another based on what they observe her to see, hear, know and infer. Here we show that chimpanzees act on the assumption that others have preferences that match their own. All subjects began with a preference for a box with a picture of food over one with a picture of nothing, even though the pictures had no causal relation to the contents. In a back-and-forth food competition, chimpanzees then avoided the box with the picture of food when their competitor had chosen one of the boxes before them—presumably on the assumption that the competitor shared their own preference for it and had already chosen it. Chimpanzees predicted that their competitor's preference would match their own and adjusted their behavioural strategies accordingly.
Keywords: theory of mind, competition, preference, social cognition, chimpanzee
1. Introduction
Individuals who can predict the behavioural decisions of others are at a huge competitive advantage. A number of recent studies have shown that primates and corvids, in particular, are highly skilled at predicting the behaviour of others by assessing their goals and perceptions in the situation [1–4]. Schmelz et al. [5] used a competitive paradigm to assess whether chimpanzees also predict behaviour by assessing others' inferences. Two chimpanzees facing each other took turns making choices in private. The subject witnessed (in private) (i) one piece of food being placed inside a hole drilled on a platform and covered by a small board and (ii) another piece being placed on the opposite side of the platform and also covered by an identical board. This resulted in one board resting flat on the platform while the other acquired a slanted orientation. After the competitor was confronted with this arrangement and allowed to choose (in private) between the flat and the slanted board, the subject could select between the two alternatives but without seeing which one had been depleted. Although chimpanzees showed a marked preference for the slanted board both in the absence of a competitor or when they chose before the competitor, they showed no such preference when they chose after the competitor.
Schmelz et al. [5] interpreted this result as evidence that chimpanzees attributed to others the same causal inference that they themselves made [6]. Here we investigated whether chimpanzees would also think that another individual's preference matched their own when the information provided was not grounded on causal relations. We used the same competitive paradigm as before but replaced the causal relationship between the food locations and the hidden food by the presence of two different photographs on the locations. In a pre-test, chimpanzees first experienced the same options that their competitor would face later. We wanted to know whether they would assume that their competitor would make the same choice they themselves had made in that situation. Would they predict that their competitor's preference would match their own even though they had by then acquired more information and knew that there was no reason to prefer one location over the other?
2. Material and methods
(a). Subjects
We tested 15 socially housed chimpanzees (Pan troglodytes) at the Wolfgang Köhler Primate Research Center in Leipzig, Germany. Eight met our test prerequisites and advanced to the testing phase (six females and two males, mean age = 27.4 years, range = 6–35 years).
(b). Apparatus
A platform (80 × 20 cm) that the experimenter could slide back and forth was placed between two opposite cages (figure 1a). Chimpanzees could see and access this platform from their location by poking their fingers through any of three holes on the bottom of a Plexiglas panel. Two identical white cardboard boxes (10 cm high; 14.5 × 10 cm bottom surface and 10 × 10 cm top surface) rested on the platform so that food could be hidden under them. On the front side, the boxes were slanted so that a photograph (9 × 13 cm) with a grey background of either ‘TEN’ food pellets or ‘ZERO’ could be propped against them (figure 1b).
Figure 1.

(a) Schematic of the experimental setup. Subject (S) and Competitor (C) face each other across the table. On the platform in the middle, there is identical food under the ‘ZERO’ and ‘TEN’ boxes. The Competitor's view is blocked by an occluder. (b) Picture of the ‘TEN’ and ‘ZERO’ boxes.
(c). Procedure
In all pre-test and test conditions, the side of the two pictures and the reward were counterbalanced and randomized with the stipulation that they could not be on the same side in more than three consecutive trials.
(i). Pre-test
Subjects received 12 trials in which E hid a single food pellet inside each of the two boxes with the ‘TEN’ and ‘ZERO’ pictures and chimpanzees were allowed to select one by poking their finger through one of the outer holes of the panel. Nine (out of 15) subjects chose ‘TEN’ above chance level (more than or equal to 10 trials out of 12, Binomial test, p < 0.05). A second round of 12 trials on a different day confirmed that those nine subjects displayed a robust preference for the picture of food by choosing the ‘TEN’ box in every trial.
(ii). Prerequisite 1
We tested those nine subjects in two prerequisites. Subjects faced a conspecific competitor (one of the chimpanzees who had not shown a preference in the pre-test) opposite to their location. While both chimpanzees watched, E placed a pellet on the platform under each of the two identical opaque cups without attached pictures and slid the platform towards the competitor so that she could choose and receive the pellet under that chosen cup. Then E slid the platform with one baited and one empty cup to the subject's side and she could choose a cup. This prerequisite established the back-and-forth paradigm between subject and competitor that was later used in the experimental conditions, and confirmed that subjects knew that the piece of food chosen by the competitor was no longer available. Subjects received 12 trials in one session and passed this prerequisite when they had chosen the baited cup in at least 10 out of 12 trials. Eight subjects passed in their first session, one after three sessions.
(iii). Prerequisite 2
E placed a food pellet on one side of the platform under one of the boxes (‘TEN’ or ‘ZERO’) while the subject watched. The other box was left empty. After baiting, E turned the boxes around so that the pictures faced the other cage and were no longer visible to the subject. He then slid the platform to the other side behind an occluder blocking the subject's view and brought it back for the subject to choose. The opposite cage was always empty and subjects knew it because they saw the empty cage before the occluder's placement. This prerequisite established that subjects (i) could remember a specific side while the platform was out of sight, (ii) knew that the pictures did not show what was underneath the box and (iii) were able to choose against their initial preference for ‘TEN’ when they knew that it was not baited. To pass this prerequisite, subjects had to select the baited box in at least 10 out of 12 trials on two consecutive days. Eight subjects passed (requiring between two and 11 sessions) while one subject always chose her right-hand side and was excluded from further testing.
(iv). Experimental conditions
Subjects received non-social (opposite cage empty) and social (conspecific competitor present in the opposite cage) 12-trial sessions. Half of the eight subjects started with two non-social, the other half with two social sessions and all received four non-social and four social sessions in an AABBAABB or BBAABBAA design, respectively. Competitors were chimpanzees who had shown no preference in the pre-test. Before test trials, subjects received six ‘warm-up’ trials identical to those in prerequisite 1 except that the subject chose first in half of the trials (randomized order). In non-social sessions, the platform was slid to the (empty) competitor's cage and subjects could choose either baited cup. These warm-up trials served to remind chimpanzees of the back-and-forth character of the study and that food that was chosen by the competitor in the social conditions was no longer available, whereas nothing happened to the food in the non-social conditions.
Following the ‘warm-up’, each experimental test session consisted of six ‘subject chooses first’ and six ‘subject chooses last’ trials presented in random order (with no more than three consecutive trials of the same kind). Identical food pellets were hidden under both boxes by E while the competitor's view was blocked behind an occluder. After the subject watched the baiting and the position of the ‘TEN’ and ‘ZERO’ pictures, the boxes were turned around so that they faced the competitor's side. In ‘subject chooses first’ trials, E moved the platform towards the subject and she could make a choice. In ‘subject chooses last’ trials, E first slid the platform towards the competitor's side behind the occluder. Whatever the competitor did, she was given a pellet from the experimenter's pocket and both hidden pellets were left under the boxes without the subject's knowledge. Then E slid the platform back to the subject's side and she could make her choice. The subject was always rewarded regardless of her choice to avoid potential learning effects over time.
3. Results
Figure 2 presents the percentage of trials in which subjects selected the ‘TEN’ box as a function of choosing order and condition (see the electronic supplementary material for individual data). There was a significant difference between conditions (Friedman test: 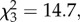 n = 8, p < 0.001). In the social conditions, subjects chose the ‘TEN’ box significantly less often when they chose last than when they chose first (Wilcoxon test: T = 0, n = 7 (one tie), p = 0.016). In contrast, no such difference was found in the non-social conditions (Wilcoxon test: T = 10, n = 7 (one tie), p = 0.594). When subjects chose last, they chose the ‘TEN’ box significantly less often in the social condition than in the non-social condition (Wilcoxon test: T = 0, n = 8, p = 0.008). When subjects chose first, there was no difference between social and non-social conditions (Wilcoxon test: T = 5, n = 6 (two ties), p = 0.281).
n = 8, p < 0.001). In the social conditions, subjects chose the ‘TEN’ box significantly less often when they chose last than when they chose first (Wilcoxon test: T = 0, n = 7 (one tie), p = 0.016). In contrast, no such difference was found in the non-social conditions (Wilcoxon test: T = 10, n = 7 (one tie), p = 0.594). When subjects chose last, they chose the ‘TEN’ box significantly less often in the social condition than in the non-social condition (Wilcoxon test: T = 0, n = 8, p = 0.008). When subjects chose first, there was no difference between social and non-social conditions (Wilcoxon test: T = 5, n = 6 (two ties), p = 0.281).
Figure 2.
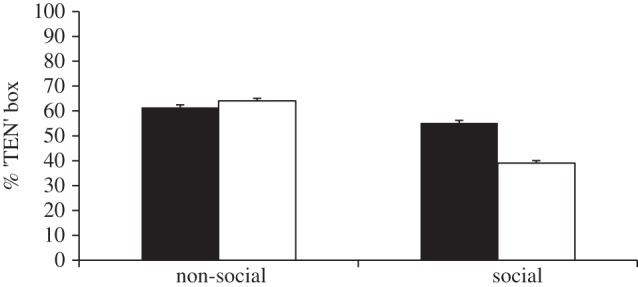
Mean percentage of trials in which subjects selected the ‘TEN’ box (±s.e.m.; n = 8) in different conditions. Filled bars denote subject chooses first, whereas unfilled bars denote subject chooses last.
Subjects selected the ‘TEN’ box significantly below chance in the ‘subject chooses last’ social condition (paired samples t-test: t7 = −2.9, n = 8, p = 0.023) whereas they selected it either above chance or at chance levels in all other conditions (paired samples t-tests: ‘subject chooses first’ non-social: t7 = 2.0, n = 8, p = 0.085; ‘subject chooses last’ non-social: t7 = 3.2, n = 8, p = 0.015; ‘subject chooses first’ social: t7 = 0.9, n = 8, p = 0.388).
4. Discussion
Chimpanzees showing a robust initial preference for the ‘TEN’ box later avoided it after their competitors had a chance to select one of the boxes before them. This result cannot be explained by avoidance learning because subjects always received the same reward regardless of their choice. In contrast, subjects showed no such avoidance in test trials without competitors or when they chose boxes before their competitors. Although subjects' preference for the ‘TEN’ box during the test was much less marked than it had been during the pre-test, note that subjects during the test saw that both boxes contained the same amount of food but they did not see the boxes' contents in the pre-test. Also in contrast to the pre-test, at the moment of choosing the pictures were not visible to the subjects but were facing the other way.
These results highlight three key issues. First, in the absence of any other information chimpanzees predicted that their competitor's choice would match their own former preference and reversed it when choosing after them. In another study based on the same competitive back-and-forth paradigm [7], chimpanzees did not do this as a categorical behavioural rule. When the competitor had not seen the baiting (equivalent to the current study), chimpanzees would not avoid a preferred food and choose a lower-quality food when they chose after their competitor.
Second, Schmelz et al. [5] suggested that chimpanzees attributed causal inferences to others. The present study shows that the causal inference component is not strictly necessary. In fact, both results could be explained in terms of chimpanzees attributing preferences to others, but without specifying how those preferences arise. This explanation, however, does not completely fit the results of the slanted board task because apes showed no preference for the slanted board when the food did not cause its inclination, even though the reinforcement contingencies were the same as when the food caused the slant [6]. In contrast, the majority of chimpanzees in the current study showed a strong preference for the box associated with the picture of multiple pellets. It is conceivable that subjects used the picture of food as an iconic cue [8] of the content of the box. Furthermore, subjects may have assumed that the iconic cue would have also caused their competitors to select it, and consequently avoided the box with the picture of food after the competitor made her choice. Although intriguing, this possibility remains untested since it is unclear that the picture possessed any iconic value.
Third, one of the shortcomings of our previous study [6] was that chimpanzees reduced their preference for the slanted board after the competitor chose first, but they did not come to prefer the other alternative. By contrast, our current results demonstrated that chimpanzees reversed their preference for the box displaying the picture of pellets. In sum, chimpanzees predicted that their competitor's choice would match their own non-causal preference and adjusted their choices accordingly.
Acknowledgements
M.S. was funded by the Volkswagen Foundation.
References
- 1.Call J, Tomasello M. 2008. Does the chimpanzee have a theory of mind? 30 years later. Trends Cogn. Sci. 12, 187–192 10.1016/j.tics.2008.02.010 (doi:10.1016/j.tics.2008.02.010) [DOI] [PubMed] [Google Scholar]
- 2.MacLean EL, Hare B. 2012. Bonobos and chimpanzees infer the target of another's attention. Anim. Behav. 83, 345–353 10.1016/j.anbehav.2011.10.026 (doi:10.1016/j.anbehav.2011.10.026) [DOI] [Google Scholar]
- 3.Clayton NS, Dally JM, Emery NJ. 2007. Social cognition by food-caching corvids. The western scrub-jay as a natural psychologist. Phil. Trans. R. Soc. B 362, 507–522 10.1098/rstb.2006.1992 (doi:10.1098/rstb.2006.1992) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bugnyar T, Heinrich B. 2005. Ravens, Corvus corax, differentiate between knowledgeable and ignorant competitors. Proc. R. Soc. B 272, 1641–1646 10.1098/rspb.2005.3144 (doi:10.1098/rspb.2005.3144) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Schmelz M, Call J, Tomasello M. 2011. Chimpanzees know that others make inferences. Proc. Natl Acad. Sci. USA 108, 3077–3079 10.1073/pnas.1000469108 (doi:10.1073/pnas.1000469108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Call J. 2007. Apes know that hidden objects can affect the orientation of other objects. Cognition 105, 1–25 10.1016/j.cognition.2006.08.004 (doi:10.1016/j.cognition.2006.08.004) [DOI] [PubMed] [Google Scholar]
- 7.Kaminski J, Call J, Tomasello M. 2008. Chimpanzees know what others know but not what they believe. Cognition 109, 224–234 10.1016/j.cognition.2008.08.010 (doi:10.1016/j.cognition.2008.08.010) [DOI] [PubMed] [Google Scholar]
- 8.Herrmann E, Melis A, Tomasello M. 2006. Apes’ use of iconic cues in the object choice task. Anim. Cogn. 9, 118–130 10.1007/s10071-005-0013-4 (doi:10.1007/s10071-005-0013-4) [DOI] [PubMed] [Google Scholar]