

Links between leaf morphology and temperature have been established at a range of ecological scales [1,2]. Narrower leaves can lose heat without evapotranspiration during hot, dry summers [3]. We proposed that an observed decrease in leaf width in Dodonaea viscosa subsp. angustissima was consistent with expectations from a changing climate based on a latitudinal cline [4].
Duncan [5] proposes a different interpretation by analysing leaf width data dating from before and after the onset of a significant warming trend separately. Duncan proposes the shift occurred before this warming trend and is not linked to temperature. Although this data split could be considered arbitrary, there is merit to the approach, in that a strong warming trend began from 1950. However, other analytical permutations may also be informative.
If temperature increases post-1950 have driven decreases in leaf width, we would expect to observe a shift between pre-1950 and more recent collections. We would also expect to observe narrowing through the latter period. In addition, it makes sense to consider a pre-1950 baseline with enough years to account for patchier collection. Finally, we would expect stronger narrowing of leaves at more northerly latitudes where temperatures are more extreme.
Here, we use alternative data splits and statistical methods to test our original findings, and to test the conclusion that temporal shifts in leaf width do not match warming trends. We include new data from specimens collected recently within the study region as vouchers for genetic sampling and systematic surveys (table 1). We know of no relevant bias in their locality or morphology and selection of leaves for measurement was conservative.
Table 1.
New leaf width data.
| voucher (AD) | leaf width (mm) | latitude | year |
|---|---|---|---|
| G.R.Guerin 1128 & E.Biffin | 1.0 | −30.778611 | 2011 |
| G.R.Guerin 1142 & E.Biffin | 0.9 | −30.454167 | 2011 |
| G.R.Guerin 1149 & E.Biffin | 1.5 | −31.338333 | 2011 |
| G.R.Guerin BRA1-A-15 & D.I.Jardine | 1.7 | −31.32717 | 2011 |
| G.R.Guerin BRA2-A-6 & D.I.Jardine | 1.2 | −31.31363 | 2011 |
| G.R.Guerin DUT1-A-16 & D.I.Jardine | 1.9 | −32.322188 | 2011 |
| G.R.Guerin DUT2-A-28 & D.I.Jardine | 3.6 | −32.311448 | 2011 |
| G.R.Guerin REM1-B-2 & D.I.Jardine | 3.1 | −32.828129 | 2011 |
| G.R.Guerin REM2-B-2 & D.I.Jardine | 4.5 | −32.750078 | 2011 |
| G.R.Guerin WAR-A-20 & E.Biffin | 1.0 | −30.77635 | 2011 |
| G.R.Guerin WIL1-A-22 & D.I.Jardine | 2.0 | −31.543914 | 2011 |
We calculated coefficients for linear leaf width decrease with time and their 95% CI with 10 000 bootstraps, including latitude as a covariate, using the following data splits: pre-1950, post-1950; 1880–2011, post-1950 between –30 and –31° N; 1920–1980, 1920–2011. We fit a GLM with year pre-1950 (n = 30) or post-1990 (n = 36) as categorical predictor and tested coefficients with 10 000 bootstraps.
Year was supported as a predictor of leaf width in all cases except the linear model using only pre-1950 data (figure 1 and table 2). Leaf width decreased with more recent data whether or not pre-1950 data were included. With pre-1950 data, bootstrapped 95% CI for coefficients overlapped zero. Data and R code to reproduce figure 1 are available in the electronic supplementary material.
Figure 1.
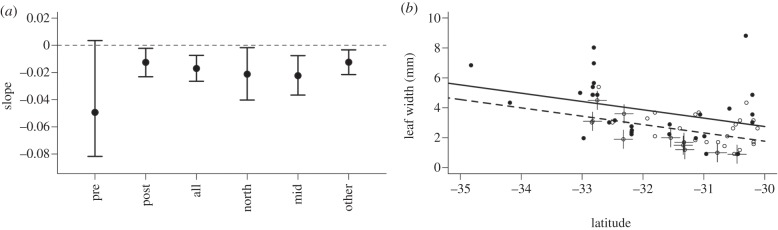
Regressions of leaf width as a function of year with latitude as covariate: (a) bootstrapped coefficients (points) and 95% CI (bars): ‘pre’ (1880–1950), ‘post’ (1950–2011), ‘all’ (1880–2011), ‘north’ (1950–2011, northernmost degree of latitude only), ‘mid’ (1920–1980), ‘other’ (1920–2011); (b) pre-1950 (filled circles and solid line) versus post-1990 (open circles and dashed line) as categorical predictor. Crosses indicate new data.
Table 2.
Statistics for temporal leaf width decrease based on 10 000 bootstraps (latitude covariate).
| years included | n | coefficient | upper 95% CI | statistically robust? |
|---|---|---|---|---|
| 1880–1950 (‘pre’) | 30 | −0.05 | 0.004a | no |
| 1950–2011 (all latitudes; ‘post’) | 232 | −0.01 | −0.002 | yes |
| 1880–2011 (‘all’) | 262 | −0.02 | −0.007 | yes |
| 1950–2011 (northern latitudes; ‘north’) | 68 | −0.02 | −0.002 | yes |
| 1920–1980 (‘mid’) | 201 | −0.02 | −0.008 | yes |
| 1920–2011 (‘other’) | 254 | −0.01 | −0.003 | yes |
| categorical: pre-1950/post-1990 | 66 | −0.98 | −0.071 | yes |
aPositive value indicates insufficient confidence in width decrease, as 95% CI overlap zero.
Pre-1950 data suggest narrowing before commencement of warming. However, this is not supported by bootstrap resampling of the data, implying the trend is not statistically robust. This analysis relies on few data (30) and is less robust than alternative splits including post-1950 data using between 66 and 262 data, especially given noise owing to sporadic, non-systematic collection and population variation.
With alternative subsampling and new data, our original interpretation is clearly supported. Narrowing occurred over the latter period and relative to a pre-1950 baseline. The apparent decrease of leaf width prior to 1950 relies on few data points and is not well supported. These conclusions are significant, given this is the first proposed evidence for a leaf morphology response to recent climate change [5].
These results do not rule out additional or alternative drivers of leaf width within the study taxon and region, although we know of no other established links between leaf width decrease and environmental drivers that operated over this period. These results support our original conclusion that observed shifts are consistent with a response to climate change. We question narrowing pre-1950 and rule out static leaf width post-1950.
Footnotes
The accompanying comment can be viewed at http://dx.doi.org/10.1098/rsbl.2012.0659.
References
- 1.Millien V, Lyons SK, Olson L, Smith FA, Wilson AB, Yom-Tov Y. 2006. Ecotypic variation in the context of global climate change: revisiting the rules. Ecol. Lett. 9, 853–869 10.1111/j.1461-0248.2006.00928.x (doi:10.1111/j.1461-0248.2006.00928.x) [DOI] [PubMed] [Google Scholar]
- 2.Peppe DJ, et al. 2011. Sensitivity of leaf size and shape to climate: global patterns and paleoclimatic applications. New Phytol. 190, 724–739 10.1111/j.1469-8137.2010.03615.x (doi:10.1111/j.1469-8137.2010.03615.x) [DOI] [PubMed] [Google Scholar]
- 3.Yates MJ, Verboom GA, Rebelo AG, Cramer MD. 2010. Ecophysiological significance of leaf size variation in Proteaceae from the Cape Floristic Region. Funct. Ecol. 24, 485–492 10.1111/j.1365-2435.2009.01678.x (doi:10.1111/j.1365-2435.2009.01678.x) [DOI] [Google Scholar]
- 4.Guerin GR, Wen H, Lowe AJ. 2012. Leaf morphology shift linked to climate change. Biol. Lett. 8, 882–886 10.1098/rsbl.2012.0458 (doi:10.1098/rsbl.2012.0458) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Duncan RP. 2012. Leaf morphology shift is not linked to climate change. Biol. Lett. 10.1098/rsbl.2012.0458 (doi:10.1098/rsbl.2012.0458) [DOI] [PMC free article] [PubMed] [Google Scholar]