

Abstract
Vocal duetting occurs in many taxa, but its function remains much-debated. Like species in which only one sex sings, duetting birds can use their song repertoires to signal aggression by singing song types that match those of territorial intruders. However, when pairs do not share specific combinations of songs (duet codes), individuals must choose to signal aggression by matching the same-sex rival, or commitment by replying appropriately to their mate. Here, we examined the song types used by female happy wrens (Pheugopedius felix) forced to make this decision in a playback experiment. We temporarily removed the male from the territory and then played songs from two loudspeakers to simulate an intruding female and the removed mate's response, using song types that the pair possessed but did not naturally combine into duets. Females were aggressive towards the female playback speaker, approaching it and overlapping the female playback songs, but nevertheless replied appropriately to their mate's songs instead of type matching the intruding female. This study indicates that females use song overlapping to signal aggression but use their vocal repertoires to create pair-specific duet codes with their mates, suggesting that duetting functions primarily to demonstrate pair commitment.
Keywords: duetting, song matching, happy wren, territory defence, cooperation
1. Introduction
Song repertoires are common in animals [1,2]. Where some or all of the repertoire is shared between individuals, rivals can match song types in territorial disputes [3,4]. Song matching can signal aggression [5] and is key in the communication system of many temperate birds, where normally only males sing [1]. Tropical birds often differ from their temperate counterparts in that both males and females sing [1], sometimes forming synchronized duets. In many duetting birds, each pair has a unique duet ‘code’ [6], linking particular song types in combinations that differ from other pairs even if the individual song types are shared [6,7].
Like temperate zone males, duetting birds can use their song repertoires for aggressive type matching [8,9] and playback experiments have demonstrated that both males and females match the song types of same-sex intruders [7,10]. In some species, this is relatively easy because pairs share male–female duet combinations across the population [10]. However, when duet codes are not standardized across pairs, signalling poses a greater challenge: aggressively type matching the same-sex intruder necessitates not singing the appropriate duet reply to a mate who is also matching its same-sex intruder. By examining the song types selected when a bird is forced to choose between effectively signalling aggression to an intruder and signalling cooperation to its mate, we can begin to disentangle the roles of cooperation and conflict in vocal duetting.
We conducted a mate removal and playback experiment to test the roles of conflict and cooperation in duetting in the happy wren (Pheugopedius felix, formerly Thryothorus [11]). Pairs of happy wrens jointly defend territories [12]. Each individual has a repertoire consisting of around 30–40 different, sex-specific song types. These are sung as solos and combined into duets, which can be initiated by males or females. Song sharing occurs in the population, but each pair maintains a unique duet code [11–13]. We asked whether female happy wrens would preferentially match the songs of an intruding female or sing the appropriate reply to playback of their mate's songs, in order to better understand the nature and function of vocal duetting.
2. Material and methods
(a). Experimental preparation
Research was conducted at the UNAM Chamela Research Station in Jalisco, Mexico. We have uniquely marked most happy wrens in the population with coloured leg rings. Prior to the experiment, we mapped territories and recorded duet and solo songs of each focal pair (n = 10). The evening before the experiment (approx. 20.00), we captured and removed the male from the focal territory using mist nets. Males were held overnight in the laboratory, given food and water, and released the next morning following the playback experiment (approx. 08.00). Males responded well to captivity and all individuals sang duets with their partner immediately upon release, retaining their territories and mates.
(b). Playback stimuli
We used two different song duet types for each pair to make the playback stimuli (figure 1). Songs were recorded (wave files, 41 kHz sampling rate) using Sennheiser ME67/K6 shotgun microphones and Marantz PMD 660 solid-state recorders. For one duet, we used the frequency cursor filter function of Syrinx (www.syrinxpc.com) to remove all vocalizations of the male. From the other duet, we removed all vocalizations of the female. We then created a playback loop of six phrases, with each bout lasting 12 s and spaced 30 s apart, maintaining the natural cadence. Using Audacity (audacity.sourceforge.net), we normalized the amplitude of each song (−3 dB) and created two-channel playback stimuli, with female songs in one channel and male songs in the other channel. The playback sequence consisted of a female song (six phrases) followed immediately by a male song (six phrases), with the male playback delayed by 1 s from the end of the last female phrase. This was repeated six times, for a 500 s playback sequence.
Figure 1.

(a,b) Natural duets produced by focal pairs were used to create (c) 2-channel playback stimuli by inserting female songs from one duet followed by male songs from the other duet. Actual stimuli included six song phrases of each sex, repeated six times during the trial. We measured whether females (d) type matched the intruder or (e) sang the appropriate reply song to their mate as predicted from each hypothesis.
Playback sequences, with female songs taken from one duet and male songs from another, were designed to force the subject to choose between matching the song type of the intruding female or singing the appropriate reply song to her male, as it was not possible to do both with the same song type. We used self song for the female playback to standardize the stimuli across females and ensure that each female had the opportunity to match the playback stimuli.
(c). Playback procedure
We used a two-speaker design [14], consisting of a wireless playback speaker (FoxPro GX-7) connected by a 10 m cable to a second speaker (FoxPro SP-55; frequency responses: 40 Hz–22 kHz). Each speaker played one channel of the two-channel recording, and therefore broadcasted songs of a single sex (male or female). The relative speaker locations and sex played through each speaker were randomly determined. Speakers were hung from branches (approx. 2 m high) along a trail within the territory boundary where we had witnessed the pair singing. The playback was remotely controlled by one observer, located between the two speakers and approximately 10 m from the trail. Additional observers were located 10 m beyond each speaker.
Playback experiments began shortly after dawn (approx. 07.00). Two minutes prior to the experimental playback, we played three to four songs from the resident male to reassure the subject that her mate was near (though he was actually still in captivity). Male and female Pheugopedius wrens roost separately and reunite to sing together at first light (N. I. Mann 2001–2003, personal observation). We broadcast male mate songs from a small speaker (Radio Shack no. 277-1008; frequency response 100 Hz–10 kHz), located halfway between the two experimental speakers, to avoid biasing the female's movements. The centre observer recorded all vocalizations and dictated behavioural observations from all three observers (via walkie-talkies) to a digital recorder. In addition to recording female vocalizations, we quantified her distance from both speakers during the 8 min trial and for 20 min subsequently, as a measure of her territorial response [12].
(d). Analyses
We visualized all songs with Syrinx (FFT = 1024 bits, Blackman window). We compared whether these song types either (i) matched the intruder female playback or (ii) were the appropriate reply song to the male playback (i.e. matched the song type she sung when we originally recorded the duet) at rates higher than expected by chance (random song selection). Because we did not have exact repertoire sizes for all subjects, we used 1/20 as an estimate of chance matching rates. All happy wrens have more than 20 song types (typically 30–40) in their repertoires [11,13], so this estimate was conservative. We compared repertoire use with two-tailed binomial tests (SPSS v. 19). Song timing was examined by measuring the proportion of female songs that (i) temporally overlapped or (ii) were sung immediately following (to create antiphonal duets) the songs broadcast during the female and male portions of the playback. We limited these analyses to subjects that sang during both male and female playback periods (n = 5), and analysed these data using a two-tailed paired t-test. We examined female movements by analysing (i) whether the initial approach was towards the male or female speaker, (ii) the proportion of the trial spent nearer each speaker, and (iii) the closest approach to each speaker. We tested these with two-tailed binomial or paired t-tests. Raw data are included as electronic supplementary material.
3. Results
Female happy wrens sang the appropriate reply song to their mate playback rather than matching the intruder female playback (figure 2a). Most focal females did not match the song type of the female playback (0/10 matched with their first song, 1/10 switched to a matching song type; 1/20 expected by chance, Binomial tests p > 0.8). In contrast, almost all females sang the appropriate reply song to their mate when they first began singing (8/10 first sang the appropriate song; 1/20 expected, Binomial test, p < 0.0001).
Figure 2.
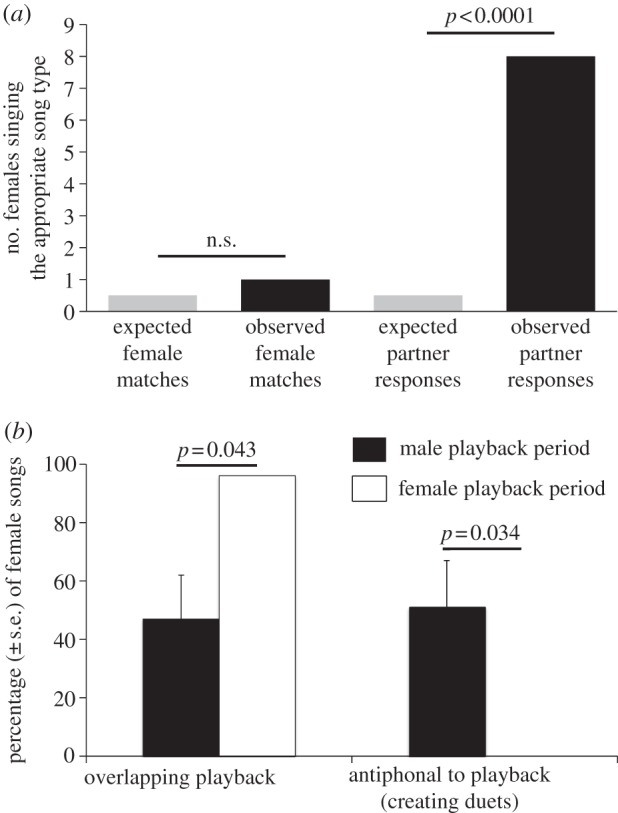
(a) Females sang the appropriate reply song to playback of their mate's song but did not type match the female intruder playback more than expected by chance. (b) Subjects also altered the timing of their songs to overlap the intruder and create well-timed duets with their mate.
Subjects timed their vocal responses (figure 2b) so that most songs sung during the male playbacks began immediately after the end of the male song phrase, creating antiphonal duets (51 ± 16% songs sung during male playback versus 0% of songs during female playback; paired t-test: t = 3.16, p = 0.034). In contrast, they were more likely to overlap female playback songs (96 ± 4% songs sung during female playbacks versus 47 ± 15% during male playbacks; t = 2.92, p = 0.043).
Most females' initial approach was towards the female playback speaker (8/10; 5/10 expected, Binomial test, p = 0.11). After the initial approach, females tended to move between the male and female playback speakers and did not spend more time near one (five were nearer the female, three nearer the male and two were equally near both; Binomial test, p = 0.72), nor did they approach one speaker more closely (closest approach to female speaker = 8.7 ± 0.88 m, closest to male speaker = 11.1 ± 1.2 m, paired t-test, t9 = 1.82, p = 0.10).
4. Discussion
Female happy wrens sang the appropriate reply song to form a duet with their mate rather than type matching the songs of the intruding female, despite showing an aggressive vocal response to the female intruder and a tendency to first approach this speaker. We suggest that this result indicates that duets function primarily as a cooperative signal in this species. If duetting functioned simply to advertise that a bird was mated, then any song type should suffice (as many intruders would not be familiar with the specific code of a given pair) and females should select the song type that would most effectively deter the intruder. Instead, it appears that by monitoring the song selection of her mate and singing the appropriate reply, a female may be signalling her commitment to the partnership [15]. In addition to being a cooperative display, signalling commitment could also function selfishly since signalling a strong bond with her mate, by singing well-practiced and carefully timed duets, may also provide a strongly aggressive signal to intruding females [16].
Subjects varied the timing of their songs depending on the context, tending to form duets with male playback songs but temporally overlap female playback songs. Overlapping has been well-studied in male singing but has not previously been examined in singing females. Overlapping has been proposed as an aggressive signal [17]; (but see critique in [5]). Here, female songs are shorter than male songs, so overlapping female playback by chance [5] should be less likely than overlapping male playback. Female happy wrens could potentially alter the meaning and intended receiver of the same song type simply by changing its timing.
In a natural context, when the vocal behaviour of both sexes is more flexible, rapid switching between song types may allow birds to both signal aggression to intruders through matching song types and overlapping and signal commitment to their partner by singing the appropriate duet code and timing [7]. We expect happy wrens, and many other duetting animals, to display such behaviour.
Our study provides a novel approach—short-term removals and playback simulating the missing bird—to examine the function of duetting, and we expect that much could be learned by applying this method to other species of duetting animals.
Acknowledgements
We thank Constantino Macias, Dave Gammon and three anonymous referees for helpful suggestions and NSF and the Leverhulme Trust for funding.
References
- 1.Catchpole CK, Slater PJB. 2008. Bird song: biological themes and variations, 2nd edn Cambridge, UK: Cambridge University Press [Google Scholar]
- 2.Janik VM, Slater PJB. 1997. Vocal learning in mammals. Adv. Study Behav. 26, 59–99 10.1016/S0065-3454(08)60377-0 (doi:10.1016/S0065-3454(08)60377-0) [DOI] [Google Scholar]
- 3.Krebs JR, Ashcroft R, Vanorsdol K. 1981. Song matching in the great tit Parus major L. Anim. Behav. 29, 918–923 10.1016/S0003-3472(81)80029-2 (doi:10.1016/S0003-3472(81)80029-2) [DOI] [Google Scholar]
- 4.Stoddard PK, Beecher MD, Campbell SE, Horning CL. 1992. Song-type matching in the song sparrow. Can. J. Zool. 70, 1440–1444 10.1139/z92-200 (doi:10.1139/z92-200) [DOI] [Google Scholar]
- 5.Searcy WA, Beecher MD. 2009. Song as an aggressive signal in songbirds. Anim. Behav. 78, 1281–1292 10.1016/j.anbehav.2009.08.011 (doi:10.1016/j.anbehav.2009.08.011) [DOI] [Google Scholar]
- 6.Logue DM. 2006. The duet code of the female black-bellied wren. Condor 108, 326–335 10.1650/0010-5422(2006)108[326:TDCOTF]2.0.CO;2 (doi:10.1650/0010-5422(2006)108[326:TDCOTF]2.0.CO;2) [DOI] [Google Scholar]
- 7.Marshall-Ball L, Slater PJB. 2004. Duet singing and repertoire use in threat signalling of individuals and pairs. Proc. R. Soc. Lond. B 271, S440–S443 10.1098/rsbl.2004.0186 (doi:10.1098/rsbl.2004.0186) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sonnenschein E, Reyer HU. 1983. Mate-guarding and other functions of antiphonal duets in the slate-colored boubou (Laniarius funebris). Z. Tierpsychol. 63, 112–140 10.1111/j.1439-0310.1983.tb00083.x (doi:10.1111/j.1439-0310.1983.tb00083.x) [DOI] [Google Scholar]
- 9.Grafe TU, Bitz JH, Wink M. 2004. Song repertoire and duetting behaviour of the tropical boubou, Laniarius aethiopicus. Anim. Behav. 68, 181–191 10.1016/j.anbehav.2003.11.004 (doi:10.1016/j.anbehav.2003.11.004) [DOI] [Google Scholar]
- 10.Rogers AC, Mulder RA, Langmore NE. 2006. Duet duels: sex differences in song matching in duetting eastern whipbirds. Anim. Behav. 72, 53–61 10.1016/j.anbehav.2005.08.019 (doi:10.1016/j.anbehav.2005.08.019) [DOI] [Google Scholar]
- 11.Mann NI, Dingess KA, Barker FK, Graves JA, Slater PJB. 2009. A comparative study of song form and duetting in neotropical Thryothorus wrens. Behaviour 146, 1–43 10.1163/156853908X390913 (doi:10.1163/156853908X390913) [DOI] [Google Scholar]
- 12.Templeton CN, Rivera-Caceres KD, Mann NI, Slater PJB. 2011. Song duets function primarily as cooperative displays in pairs of happy wrens. Anim. Behav. 82, 1399–1407 10.1016/j.anbehav.2011.09.024 (doi:10.1016/j.anbehav.2011.09.024) [DOI] [Google Scholar]
- 13.Brown RN, Lemon RE. 1979. Structure and evolution of song form in the wrens Thryothorus sinaloa and T. felix. Behav. Ecol. Sociobiol. 5, 111–131 10.1007/BF00293301 (doi:10.1007/BF00293301) [DOI] [Google Scholar]
- 14.Douglas SB, Mennill DJ. 2010. A review of acoustic playback techniques for studying avian vocal duets. J. Field Ornithol. 81, 115–129 10.1111/j.1557-9263.2010.00268.x (doi:10.1111/j.1557-9263.2010.00268.x) [DOI] [Google Scholar]
- 15.Wickler W. 1980. Vocal duetting and the pair bond. I. Coyness and partner commitment, a hypothesis. Z. Tierpsychol. 52, 201–209 10.1111/j.1439-0310.1980.tb00711.x (doi:10.1111/j.1439-0310.1980.tb00711.x) [DOI] [Google Scholar]
- 16.Hall ML. 2004. A review of hypotheses for the functions of avian duetting. Behav. Ecol. Sociobiol. 55, 415–430 10.1007/s00265-003-0741-x (doi:10.1007/s00265-003-0741-x) [DOI] [Google Scholar]
- 17.Naguib M, Mennill DJ. 2010. The signal value of birdsong: empirical evidence suggests song overlapping is a signal. Anim. Behav. 80, E11–E15 10.1016/j.anbehav.2010.06.001 (doi:10.1016/j.anbehav.2010.06.001) [DOI] [Google Scholar]