

Abstract
In cooperative breeders, sexually mature subordinates can either queue for chances to inherit the breeding position in their natal group, or disperse to reproduce independently. The choice of one or the other option may be flexible, as when individuals respond to attractive dispersal options, or they may reflect fixed life-history trajectories. Here, we show in a permanently marked, natural population of the cooperatively breeding cichlid fish Neolamprologus pulcher that subordinate helpers reduce investment in territory defence shortly before dispersing. Such reduction of effort is not shown by subordinates who stay and inherit the breeding position. This difference suggests that subordinates ready to leave reduce their investment in the natal territory strategically in favour of future life-history perspectives. It seems to be part of a conditional choice of the dispersal tactic, as this reduction in effort appears only shortly before dispersal, whereas philopatric and dispersing helpers do not differ in defence effort earlier in life. Hence, cooperative territory defence is state-dependent and plastic rather than a consistent part of a fixed life-history trajectory.
Keywords: cooperative breeding, dispersal, consistent behavioural variation, life history, helping, Neolamprologus pulcher
1. Introduction
In cooperative breeders, offspring usually delay dispersal from their natal territory [1]. When reproductively mature, they have two options to obtain a breeding position: they may either queue in their natal group to inherit the dominant breeding position or disperse elsewhere when breeding vacancies arise [1,2]. These strategies can be flexible, e.g. individuals may respond to current changes in expected fitness benefits from queuing or dispersing, irrespective of their previous behaviour and developmental history [3]. Alternatively, individuals may be predetermined for philopatry or dispersal by genetic, parental or early environmental effects, i.e. these strategies would reflect fixed life-history trajectories [4].
Field studies investigating dispersal often suffer from limited possibilities to recapture dispersers reliably. A recent study of paper wasps Polistes dominulus demonstrates convincingly that helpers with long nest tenure spend higher cooperative effort than those with short tenure but it was unable, for instance, to exclude that individuals classified as dispersed just might have died early [5]. Hence, studies that can reliably differentiate dispersal from mortality can provide valuable insight into the ecology of cooperative breeding [6].
This is particularly important when studying the strategic adjustment of helpers' behaviour to the decision to stay in or leave the natal territory. In the cichlid Neolamprologus pulcher, laboratory experiments showed that large subordinates reduce their investment in energetically costly territory maintenance when dispersal options are available [7]. Strategic helping adjusted to expected future fitness payoffs has also been demonstrated in the hover wasp Liostenogaster flavolineata, where subordinates adjust helping effort to the chance to inherit the territory and the expected number of helpers they will then have [3]. Furthermore, observational evidence suggests that extra territorial prospecting of male subordinate meerkats Suricata suricatta is traded off against cooperative effort in the home territory [8].
It is yet unclear, however, whether and how subordinate group members of cooperative breeders in nature adjust their helping effort before leaving the natal territory, even when a dispersal option has not yet turned up. Helpers that have decided to leave should anticipate dispersal by a strategic reduction of effort at home. Here, we report on the effort in cooperative territory defence and maintenance of subordinates in groups of N. pulcher before and after (i) dispersal or (ii) territory inheritance. We monitored dispersed individuals and compare their behaviour pre - and post-dispersal with that of philopatric group members pre- and post-inheritance of their territory. We predicted that if subordinates adjust helping effort strategically to their future life-history trajectory, individuals that have decided to disperse or to stay should differ in cooperative effort before dispersal or territory inheritance. Prospective heirs of their natal territory should show high investment, particularly in territory defence, whereas prospective dispersers should reduce their investment in costly cooperation before leaving home.
2. Methods
(a). Study species
Neolamprologus pulcher is a cooperatively breeding cichlid endemic to Lake Tanganyika, East Africa. The fish were studied in 10–12 m depth at Kasakalawe point, Zambia (8°46.849′ S, 31°04.882′ E) from September to December 2009 and September to December 2010. Groups contain up to 25 subordinates participating in cooperative territory defence, territory maintenance and brood care [9]. Subordinates remain in their natal group queuing for dominance, or they disperse into other groups, usually long after sexual maturity [2,9]. Dispersal distances of up to 50 m have been recorded, but most dispersal distances do not exceed 10 m [10].
(b). The study protocol
Individuals were captured, measured, fin clipped and individually marked with elastomer before release back in their original group. We marked 341 individuals in 96 cooperative groups. We surveyed marked individuals weekly together with group size and composition, and recorded dispersal and territory inheritance. Their behaviour was recorded 3–7 times during the study for 7 min each using a standardized focal observation protocol and Observer software (Noldus Inc.) on waterproof packed Psion computers. Focal recordings included the frequency of territory defence (attacks against predators and conspecifics from other territories) and territory maintenance (digging events to remove sand from a shelter). Data are deposited in the Dryad repository [11].
(c). Behavioural analyses
(i). Long-term behavioural differences
We assessed whether the individuals that dispersed or inherited a breeding position between December 2009 and September 2010 already differed in their behaviour one to ten months prior to the takeover events. This analysis includes all observations (N = 118) conducted in the field season 2009 on individuals that were recaptured in 2010, taking into account the takeover type of the breeding position. This dataset includes n = 31 subordinate individuals of which 20 inherited the natal territory and 11 dispersed. Behavioural data were averaged before they were analysed with a linear model using the program R [12]. The data were log transformed and after fitting the model, residual plots were used to confirm that no assumption of the statistical model was violated. If linear models could not be used, non-parametric tests were applied.
(ii). Behavioural differences shortly before takeover
We analysed a dataset of 16 individuals observed shortly before and shortly after dispersal or inheritance (maximum time interval: two months). These individuals were observed in 2010 when they had either dispersed (n = 7) or inherited a breeding position at home (n = 9) during this field season. In total 80 observations were conducted on these focal individuals (32 on dispersers and 48 on territory heirs). We fitted generalized linear mixed models with the R packages lme4 [13] and nlme [14], and included the takeover type (dispersal or inheritance) and the observation period (before or after takeover) as two fixed factors with two levels each. In addition, we included the individual identity as a random factor. First, we fitted full models including the interaction of the fixed factors and reduced non-significant factors stepwise, starting with the highest order interaction. Terms that dropped out of the model are presented with the coefficient and p-value at which they did so.
3. Results
(a). Behaviour of individuals long before their dispersal or territory inheritance
The behaviour of N. pulcher 1–9 months before obtaining a breeding position did not differ between individuals that dispersed and individuals inheriting the territory, neither in their investment in territory defence against predators (mean ± s.e. heirs: 1.50 ± 0.36; mean dispersers: 2.34 ± 0.46; linear model: heirs: Est = 0.75 ± 0.12; dispersers: Est = 0.34 ± 0.21, p = 0.11; N = 31) and conspecific members of other groups (mean heirs: 0.18 ± 0.06; mean dispersers: 0.11 ± 0.06; Mann–Whitney test, p = 0.48; N = 31) nor in territory maintenance (mean heirs: 0.66 ± 0.30; mean dispersers: 1.10 ± 0.33; Mann–Whitney test, p = 0.10; N = 31).
(b). Behaviour of individuals shortly before their dispersal or territory inheritance
Prior to dispersal and inheritance, dispersers showed significantly less defence effort against predators than philopatric individuals (figure 1 and table 1). However, after dispersal or inheritance, territory defence rates did not differ, suggesting that defence effort increased more strongly after dispersal than after inheritance (significant interaction of takeover type × period; figure 1 and table 1). Territory defence against conspecific intruders was higher prior to dispersal or inheritance than thereafter, but did not differ between individuals pursuing different takeover types (table 1). Dispersers and heirs did not differ in their territory maintenance, but all individuals enhanced territory maintenance after acquiring dominance status (table 1).
Figure 1.
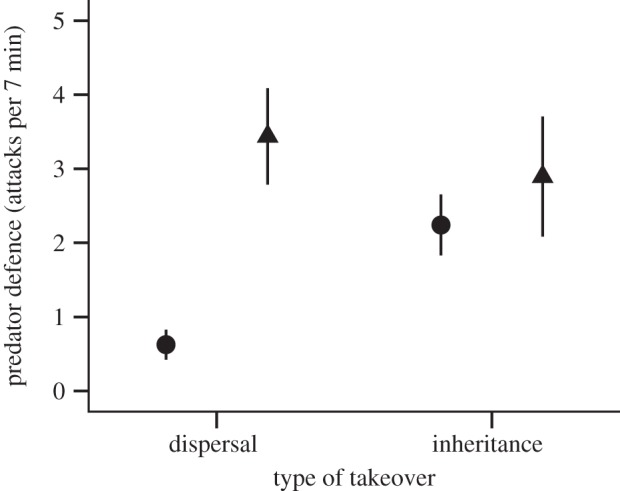
Territory defense of N. pulcher in dependence of their type of breeding position takeover before (circle) and after (triangle) takeover. Depicted are means ± s.e. of the raw data. N = 80 observations on n = 16 individuals.
Table 1.
Factors explaining the cooperative investment of helpers. Full models included the observation period (before or after takeover) and the type of takeover (inheritance or dispersal) as fixed factors, the interaction and the individual identity as random factor. All displayed models are based on N = 80 observations on n = 16 individuals.
| estimate | t/z-value | p-value | |
|---|---|---|---|
| territory defence against predators | |||
| intercept | 0.38 ± 0.18 | 2.13 | 0.04 |
| period (before/after takeover) | 0.92 ± 0.24 | 3.80 | <0.001 |
| takeover type (inheritance/dispersal) | 0.56 ± 0.22 | 2.50 | 0.03 |
| period × takeover type | −0.83 ± 0.32 | −2.60 | 0.01 |
| territory defence against conspecifics | |||
| intercept | −0.78 ± 0.31 | −2.54 | 0.01 |
| period (before/after takeover) | −0.80 ± 0.40 | −2.00 | 0.046 |
| takeover type (inheritance/dispersal)a | −0.62 ± 0.53 | −1.18 | 0.24 |
| period × takeover typea | −1.07 ± 0.92 | −1.155 | 0.25 |
| territory maintenance | |||
| intercept | 0.19 ± 0.0.20 | 0.91 | 0.36 |
| period (before/after takeover) | 0.69 ± 0.17 | 4.00 | <0.001 |
| takeover type (inheritance/dispersal)a | −0.03 ± 0.36 | −0.08 | 0.94 |
| period × takeover typea | 0.02 ± 0.35 | 0.05 | 0.96 |
aThese terms are displayed with the parameter estimates at the point they were dropped from the model.
4. Discussion
Helpers of N. pulcher invested differently in cooperative territory defence, depending on their future life history perspective. Subordinates that later dispersed exhibited less territory defence shortly before dispersal, suggesting an anticipatory adaptive response to their specific future life history. Participation in cooperative tasks in this species can serve as payment from subordinates to the dominant individuals to be accepted in the territory [15,16]. In addition, subordinates can benefit from territory defence if they inherit breeding status in their territory and benefit from group augmentation caused by their investment. Both mechanisms, independently or in conjunction, can select for costly investment of helpers that stay in the territory, but not of individuals that will soon disperse to breed elsewhere. It is possible that dispersers left voluntarily, or alternatively they got evicted. Either way, dispersers changed their behaviour shortly before dispersal, suggesting contingency between the change of behaviour and subsequent dispersal.
A similar strategic reduction of investment prior to dispersal is shown by paper wasps, where subordinates reduce cooperative foraging and tend to rest more in the nest prior to dispersal [5]. To our knowledge, however, this field study is the first to combine information about the survival of subordinate group members to successful inheritance or dispersal, and the cooperative behaviour preceding distinctively different forms of group takeover. Furthermore, post dispersal behavioural data reveal that individuals that dispersed increased their investment in predator defence in the new group in comparison to the behaviour they showed in their group of origin. Hence, the detected differences in cooperative behaviour are not due to differential body condition or survival potential (cf. [5]), which provides new insight into the ecology and evolution of dispersal and cooperative breeding.
Our results are consistent with a previous laboratory experiment showing that potential dispersal options cause large N. pulcher helpers to reduce investment in their home territory [7]. However, the implications of our results differ in important aspects from this previous evidence. We compared individuals successfully dispersing or inheriting a breeding position in a natural population, whereas in the previous study dispersal options were experimentally provided, but actual dispersal did not happen in most cases [7]. Hence, it was not clear if the behavioural change was a response to a specific life-history trajectory, or if helpers merely respond to changes in dispersal and breeding opportunities as indicated by habitat parameters. Furthermore, we found in this study that adaptive behavioural decisions were made already before breeding vacancies arose. This implies that subordinate N. pulcher anticipate dispersal and adjust their investment at home before their decision to leave can be implemented. This has important implications for the social system, because vacancies for dispersing individuals are usually created by predation events, which are difficult to predict. Nevertheless, N. pulcher helpers appeared to adjust their helping effort strategically to upcoming dispersal already when the territory to which they would disperse was not yet determined and available. As contributions to territory defence at a longer interval (1–9 months) before successful dispersal or inheritance did not differ from each other, this behaviour is apparently strategically adjusted rather shortly before dispersal, suggesting strategic investment reduction rather than a consequence of fixed life-history trajectories [4]. If fixed life-history trajectories cause consistent behavioural variation among individuals, theoretical models predict that this variation should be observed already early in life, and should persist beyond dispersal or territory inheritance [4]. The second prediction was also not met by our data: after acquiring a breeding position, individuals did not differ in their territory defence, regardless whether they had dispersed or inherited.
Acknowledgements
We thank Stefanie Meindl for help with data collection, Ruben Shapola and Danny Sinyinza for their logistic support. We are grateful to Peter Biedermann and Ben Dantzer for comments on previous versions of the manuscript. Funding: SNF-grant no. 3100A0–122511 to M.T.
References
- 1.Koenig WD, Pitelka FA, Carmen WJ, Mumme RL, Stanback MT. 1992. The evolution of delayed dispersal in cooperative breeders. Q. Rev. Biol. 5, 111–150 10.1086/417552 (doi:10.1086/417552) [DOI] [PubMed] [Google Scholar]
- 2.Stiver KA, Fitzpatrick J, Desjardins JK, Balshine S. 2006. Sex differences in rates of territory joining and inheritance in a cooperatively breeding cichlid fish. Anim. Behav. 71, 449–456 10.1016/j.anbehav.2005.06.011 (doi:10.1016/j.anbehav.2005.06.011) [DOI] [Google Scholar]
- 3.Field J, Cronin A, Bridge C. 2006. Future fitness and helping in social queues. Nature 441, 214–217 10.1038/nature04560 (doi:10.1038/nature04560) [DOI] [PubMed] [Google Scholar]
- 4.Wolf M, van Doorn G, Leimar O, Weissing FJ. 2007. Life-history trade-offs favour the evolution of animal personalities. Nature 447, 581–584 10.1038/nature05835 (doi:10.1038/nature05835) [DOI] [PubMed] [Google Scholar]
- 5.Tibbetts EA. 2007. Dispersal decisions and predispersal behavior in Polistes paper wasp ‘workers’. Behav. Ecol. Sociobiol. 61, 1877–1883 10.1007/s00265-007-0427-x (doi:10.1007/s00265-007-0427-x) [DOI] [Google Scholar]
- 6.Walters JR, Doerr PD, Carter JH. 1992. Delayed dispersal and reproduction as a life-history tactic in cooperative breeders: fitness calculations from red-cockaded woodpeckers. Am. Nat. 139, 623–643 10.1086/285347 (doi:10.1086/285347) [DOI] [Google Scholar]
- 7.Bergmuller R, Heg D, Taborsky M. 2005. Helpers in a cooperatively breeding cichlid stay and pay or disperse and breed, depending on ecological constraints. Proc. R. Soc. Lond. B 272, 325–331 10.1098/rspb.2004.2960 (doi:10.1098/rspb.2004.2960) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Young AJ, Carlson AA, Clutton-Brock T. 2005. Trade-offs between extraterritorial prospecting and helping in a cooperative mammal. Anim. Behav. 70, 829–837 10.1016/j.anbehav.2005.01.019 (doi:10.1016/j.anbehav.2005.01.019) [DOI] [Google Scholar]
- 9.Taborsky M, Limberger D. 1981. Helpers in fish. Behav. Ecol. Sociobiol. 8, 143–145 10.1007/BF00300826 (doi:10.1007/BF00300826) [DOI] [Google Scholar]
- 10.Zöttl M. 2012. The social and ecological influences on commodity trading in the cooperatively breeding cichlid Neolamprologus pulcher. PhD thesis, University of Bern [Google Scholar]
- 11.Zöttl M, Chapuis L, Freiburghaus M, Taborsky M. 2012. Strategic reduction of help before dispersal in a cooperative breeder. Dryad Digital Respository. (doi:10.5061/dryad.8fg95) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.R Development Core Team 2011. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing [Google Scholar]
- 13.Bates D, Maechler M, Bolker B. 2011. lme4: Linear mixed-effects models using S4 classes. Version 0.999375–39
- 14.Pinheiro J, Bates D, DebRoy S, Sarkar D, the R Development Core Team 2010. nlme: linear and nonlinear mixed effects models. Version 3.1–97
- 15.Bergmuller R, Taborsky M. 2005. Experimental manipulation of helping in a cooperative breeder: helpers ‘pay to stay’ by pre-emptive appeasement. Anim. Behav. 69, 19–28 10.1016/j.anbehav.2004.05.009 (doi:10.1016/j.anbehav.2004.05.009) [DOI] [Google Scholar]
- 16.Zöttl M, Heg D, Chervet N, Taborsky M. In press Kinship lowers alloparental brood care when helpers pay-to-stay. Nat. Commun . [DOI] [PubMed] [Google Scholar]