

Abstract
The rise of dinosaurs was a major event in vertebrate history, but the timing of the origin and early diversification of the group remain poorly constrained. Here, we describe Nyasasaurus parringtoni gen. et sp. nov., which is identified as either the earliest known member of, or the sister–taxon to, Dinosauria. Nyasasaurus possesses a unique combination of dinosaur character states and an elevated growth rate similar to that of definitive early dinosaurs. It demonstrates that the initial dinosaur radiation occurred over a longer timescale than previously thought (possibly 15 Myr earlier), and that dinosaurs and their immediate relatives are better understood as part of a larger Middle Triassic archosauriform radiation. The African provenance of Nyasasaurus supports a southern Pangaean origin for Dinosauria.
Keywords: Dinosauria, Archosaur radiation, Triassic, Manda beds, phylogeny
1. Introduction
The appearance of dinosaurs in the Triassic was an important event in the history of life because it marked the onset of a faunal diversification that has dominated terrestrial ecosystems for almost 230 Myr. The exact timing of dinosaur origins and the pace of early dinosaur diversification remain unclear [1,2], but a growing consensus suggests that dinosaurs were initially rare components of Early–Late Triassic terrestrial ecosystems and that their ascent was gradual and regionally diachronous [3]. The oldest well-dated and unambiguously identified dinosaurs are from the late Carnian (approx. 230 Ma) of Argentina [4], and by the end of the Carnian, all three major dinosaur lineages were present (Ornithischia, Sauropodomorpha, Theropoda) [5,6]. The surprising early diversity and morphological disparity of early dinosaurs occurring together in the oldest dinosaur-bearing strata indicate that either (i) dinosaurs experienced an ‘early burst’ of diversification in the Carnian, or (ii) dinosaurs had a deeper, cryptic origination in the Triassic and that their diversification occurred more slowly than shown by current knowledge of their fossil record. The closest relatives of dinosaurs, silesaurids, are known from the late Anisian (Middle Triassic); ghost lineages inferred from phylogenetic analyses of ornithodirans predict that Dinosauria and its immediate stem lineage should be at least as old if these topologies are correct [7].
To date, every reported pre-late Carnian dinosaur body-fossil or footprint(s) has proved to be non-dinosaurian, referable to taxa with convergent morphology, or ambiguously identified [1,8]. Here, we present the best available evidence for a Middle Triassic dinosaur (or at least the closest dinosaur relative yet found) from the late Anisian of Tanzania.
2. Systematic palaeontology
Archosauria Cope 1869 (sensu Gauthier & Padian 1985)
Dinosauriformes Novas 1992
Dinosauria Owen 1842 (sensu Padian & May 1993)
Nyasasaurus parringtoni gen. et sp. nov.
(a). Etymology
Nyasa, from Lake Nyasa near the type locality, and sauros, Greek for lizard; parringtoni, in honour of Francis Rex Parrington, collector of the holotype.
(b). Holotype and referred material
Holotype: NHMUK R6856 (The Natural History Museum, London, UK); right humerus, three partial presacral vertebrae and three sacral vertebrae. Referred specimen: SAM-PK-K10654 (Iziko South African Museum, Cape Town, South Africa) (=Thecodontosaurus alophos); three cervical vertebrae and two posterior presacral vertebrae.
(c). Locality and age
The holotype was collected from a single locality (Parrington's locality B36) from the Lifua Member of the Manda beds, Ruhuhu Basin, southern Tanzania. The Lifua Member is late Anisian (Middle Triassic) in age, on the basis of biostratigraphic comparisons with the tetrapod fauna of the Cynognathus Assemblage Zone (subzone C) of South Africa [9]. Cynodonts (Aleodon brachyrhamphus and Scalenodon angustifrons), dicynodonts (Sangusaurus edentatus and Angonisaurus cruickshanki) and the rhynchosaur Stenaulorhynchus stockleyi co-occur at the same horizon in the immediate vicinity.
(d). Diagnosis
Nyasasaurus parringtoni was a 2–3 metre long (estimated from vertebral dimensions) dinosauriform with the following unique combination of humeral character states: ventrally elongated deltopectoral crest; laterally deflected apex of the deltopectoral crest; distinct notch central to the apex of the deltopectoral crest; pointed expansion on the proximal surface near the dorsal extent of the deltopectoral crest; proximal surface of the humerus continuous with the lateral surface of the deltopectoral crest; and distinct fossa present on the posterodorsal surface, just ventral to the proximal surface. Humeral histology indicates rapid growth characterized by complex vascularization, highly woven bone tissue and the absence of any lines of arrested growth. The vertebrae also have a distinct combination of character states, including: at least three sacral vertebrae; dorsoventrally tall sacral ribs; and hyposphene–hypantrum intervertebral articulations in the presacral vertebrae (last character also present in the referred specimen; see the electronic supplementary material for additional information).
3. Description
The humerus of Nyasasaurus parringtoni bears a subrectangular, distally elongated deltopectoral crest that extends down the shaft for more than 30 per cent of its estimated total length, an unambiguously optimized dinosaur character state in all recent phylogenies [1,2]. The apex of the deltopectoral crest is deflected laterally at its anterior tip, a feature present exclusively in dinosaurs (e.g. Tawa, Saturnalia, Heterodontosaurus) as found in our phylogenetic analysis. The humerus does not share any synapomorphies exclusively with any other Triassic archosaur clade.
The preserved axial column consists of partial posterior presacral vertebrae and elements of the sacrum. The presacral vertebrae bear hypantrum intervertebral articulations as indicated by a clear gap between the prezygapophyses, as in some pseudosuchians (e.g. Batrachotomus), silesaurids (e.g. Silesaurus) and dinosaurs (e.g. Dilophosaurus). The sacrum consists of three partial vertebrae: two primordial/ancestral vertebrae frame an additional vertebra between the two (see the electronic supplementary material). Parts of each sacral rib are preserved, and the articular facet on the first sacral rib indicates that the ilium was nearly vertical and the sacral rib was dorsoventrally elongated like that of all early dinosaurs [10].
The referred specimen (SAM-PK-K10654) consists of presacral vertebrae, three of which are anteroposteriorly elongated cervical vertebrae with significant vertical offset between the anterior and posterior articular faces. One specimen (figure 1j) preserves the bases of epipophyses immediately dorsal to the postzygapophyses, a character state present in, but not exclusive to, Dinosauriformes. The cervical vertebrae bear deep fossae (i) lateral to the neural canal in anterior view, and (ii) on the posterolateral surface of the neural arch just anteroventral to the prezygapophyses. Both of these character states are present in Silesaurus [11], but not in the earlier diverging silesaurid Asilisaurus, and in early theropod dinosaurs [10] and have been interpreted as possibly homologous with features that represent unambiguous postcranial pneumaticity in theropod dinosaurs [12].
Figure 1.

Holotype (a–h; NHMUK R6856) and referred (i–j; SAM-PK-K10654) specimens of Nyasasaurus parringtoni gen. et sp. nov. Right humerus in (a) anterior and (b) posterior views. Histological section of humerus in (c) complete cross-section in transmitted light, (d) cross-section through the entire cortex in crossed Nicols, and (e) cross-section through the outer portion of cortex in crossed Nicols. (f) Rearticulated sacrum in right lateral view with interpretive drawing (below). (g) Posterior presacral vertebra in right lateral view. (h) Partial posterior presacral vertebra in dorsal view. (i) Anterior cervical vertebra in left lateral view with interpretive drawing (below). (j) Anterior cervical vertebra in left lateral view with interpretive drawing (below). Arrows point anteriorly. Scale bars, (a,b,f–j) 1 cm, (c) 4 mm, (d) 1 mm, (e) 500 nm. Dep, depression; df, deep fossa; dia, diapophysis; dp, deltopectoral crest; epi, epipophysis; hs, histology section; hyp, hypantrum; no, notch; ns, neural spine; par, parapophysis; pre, prezygapophysis; pz, postzygapophysis; r, ridge; s1–3, sacral vertebra number; sr1–3, sacral rib number; st, striations.
The histology of the holotypic humerus (figure 1c–e) suggests sustained, elevated growth rates similar to those of dinosaurs (e.g. Megapnosaurus (Syntarsus) rhodesiensis, Herrerasaurus) and higher than those of most other early archosauriforms with the exception of pterosauromorphs [13,14]. Unaltered, primary, woven-fibered bone comprises much of the humeral cortex, and all vascular canals are primary osteons. Many of these canals are longitudinal canals, but at least half of these anastomose with other canals (in all directions), and locally, short radial canals may dominate. The outermost cortex shows a decrease in osteocytes and a temporary transition from woven-fibred to paralleled-fibred bone (figure 1e), which we tentatively identify as an annulus (see the electronic supplementary material for further details).
This condition differs from that of Silesaurus, which shows low levels of vascular connectivity and lacks the complex vascular patterns seen in Nyasasaurus [15], and is histologically more similar to Megapnosaurus rhodesiensis (see the electronic supplementary material).
4. Relationships
Nyasasaurus bears two character states that are considered autapomorphic for dinosaurs in the holotype (elongated and laterally deflected deltopectoral crest) and one character state that could represent a dinosaur plesiomorphy, but which has a complex distribution among dinosauriforms (three sacral vertebrae). Additionally, the referred specimen bears two character states in the cervical vertebrae consistent with, but not exclusive to, theropod dinosaurs (deep fossae lateral to the neural canal in anterior view, deep lateral fossae). Together with all other aspects of the morphology, this combination of character states is unique for any dinosaur or close relative; so we scored the holotype and the referred specimen independently in a comprehensive early archosauriform phylogeny [16], but then combined their scorings in a second analysis. The methods of the phylogenetic analysis were identical to those of Butler et al. [17] and the Nexus file, and high-resolution histological and osteological images are deposited online at http://morphobank.org/permalink/?P485 (see the electronic supplementary material for accession numbers). The holotype was found in a polytomy with all members of Dinosauria included in the analysis in the strict consensus tree, but was found as the sister–taxon of Dinosauria or within Dinosauria in the various most parsimonious trees. The referred specimen was recovered within Dinosauria as a theropod, and when the scores of the holotype and referred specimen were combined, Nyasasaurus was found as either the sister taxon of Dinosauria or to lie within the clade.
The shared synapomorphies of Nyasasaurus and Dinosauria are present in axial and appendicular elements, thus demonstrating that the characters are spread across the skeleton rather than restricted to a single element. We interpret these results as supporting our identification of Nyasasaurus as either the sister–taxon to, or the earliest-known member of, Dinosauria, but do not consider the phylogenetic positions within Dinosauria as a robust portrayal of the available evidence. The unique character combination of Nyasasaurus may represent the plesiomorphic states present just outside or within Dinosauria given its age, but there are considerable uncertainties about (i) the plesiomorphic character states of early ornithischians [17], (ii) character optimizations of vertebral fossae interpreted as pneumatic-like [18], and (iii) the phylogenetic position of early saurischians such as Herrerasaurus [1].
5. Discussion
All of the available morphological, phylogenetic and histological evidence indicates that Nyasasaurus represents either the sister–taxon to, or the earliest member of, Dinosauria (figure 2). The occurrence of a late Anisian ‘stem’ dinosaur or true dinosaur has previously been predicted by the sister–taxon relationship with the Middle–Late Triassic silesaurids [7]. It is noteworthy that the oldest known silesaurid, Asilisaurus kongwe, also occurs in the Lifua Member. This co-occurrence indicates (i) that dinosaurs and their nearest relatives coexisted in the same terrestrial environments, and (ii) if Nyasasaurus belongs to Dinosauria or the dinosaurian stem lineage, the current 10–15 Myr ghost lineage separating these taxa is immediately eliminated (figure 2).
Figure 2.
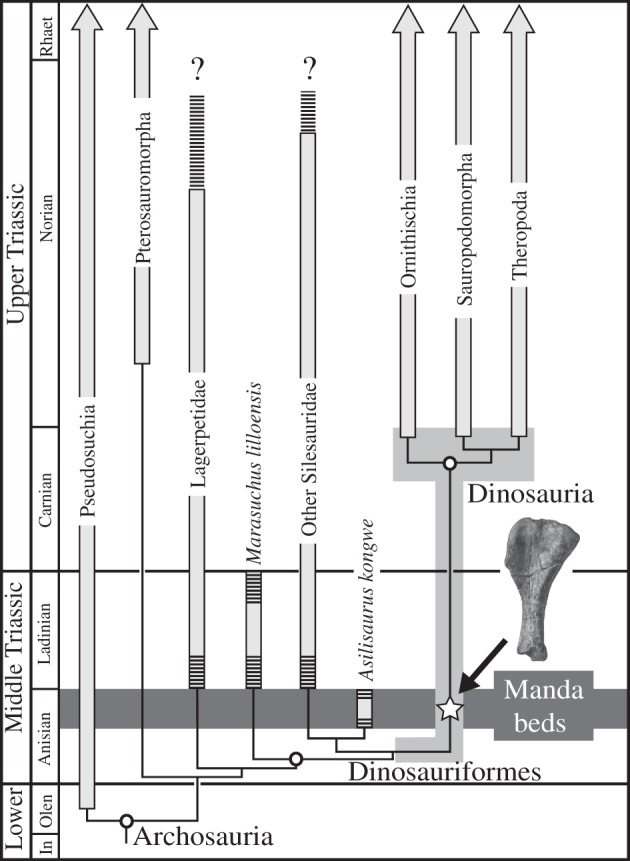
Possible phylogenetic position of Nyasasaurus parringtoni gen. et sp. nov. as either within, or the sister-taxon of, Dinosauria (indicated by the medium grey box), illustrating the decrease in the dinosaurian ghost lineage. Dashed bars represent uncertainty. In, Induan; Olen, Olenekian.
The relationships of Nyasasaurus and its age have important implications regardless of whether this taxon is a dinosaur or ‘near-dinosaur.’ The late Anisian age demonstrates that essentially dinosaur-like dinosauriforms (or the earliest dinosaurs) were just another component of an extensive archosauriform radiation that existed in the aftermath of the end-Permian mass extinction. Moreover, if Nyasasaurus is a dinosaur, it establishes that dinosaurs evolved earlier than previously expected and that the dinosaurs from the Ischigualasto Formation and elsewhere do not necessarily record an ‘early burst’ of diversification in the Late Triassic. The limited material available for Nyasasaurus in comparison with that for other members of the same assemblages (e.g. synapsids, rhynchosaurs; [19]) also suggests that dinosaurs or ‘near-dinosaurs’ were rare in Middle Triassic vertebrate assemblages. Finally, the occurrence of Nyasasaurus parringtoni in Tanzania supports a southern Pangaean origin for dinosaurs, as predicted by the distributions of the earliest unambiguous dinosaur taxa from southern South America [2].
Acknowledgements
We thank S. Chapman (NHMUK) for permission to section and cast the humerus and A. C. Milner (NHMUK) for access to A. Charig's personal papers. Photographs of the specimens were provided by P. Crabb (NHMUK). We thank R. Irmis, M. Stocker, K. Padian, P. Makovicky, R. Butler and N. Fraser for discussion and helpful comments. This work was supported by NSF EAR-1024036 and a NHMUK collections improvement grant.
References
- 1.Brusatte S, Nesbitt SJ, Irmis RB, Butler R, Benton MJ, Norell MA. 2010. The origin and early radiation of dinosaurs. Earth-Sci. Rev. 101, 68–100 10.1016/j.earscirev.2010.04.001 (doi:10.1016/j.earscirev.2010.04.001) [DOI] [Google Scholar]
- 2.Langer MC, Ezcurra MD, Bittencourt JS, Novas FE. 2010. The origin and early evolution of dinosaurs. Biol. Rev. 85, 55–110 10.1111/j.1469-185X.2009.00094.x (doi:10.1111/j.1469-185X.2009.00094.x) [DOI] [PubMed] [Google Scholar]
- 3.Irmis RB. 2011. Evaluating hypotheses for the early diversification of dinosaurs. Earth Environ. Sci. Trans. R. Soc. Edinburgh 101, 397–426 10.1017/S1755691011020068 (doi:10.1017/S1755691011020068) [DOI] [Google Scholar]
- 4.Martinez RN, Sereno PC, Alcober OA, Colombi CE, Renne PR, Montañez IP, Currie BS. 2011. A basal dinosaur from the dawn of the dinosaur era in southwestern Pangaea. Science 331, 206–210 10.1126/science.1198467 (doi:10.1126/science.1198467) [DOI] [PubMed] [Google Scholar]
- 5.Martinez RN, Alcober OA. 2009. A basal sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the early evolution of Sauropodomorpha. PLoS ONE 4, e4397. 10.1371/journal.pone.0004397 (doi:10.1371/journal.pone.0004397) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sereno PC. 1999. The evolution of dinosaurs. Science 284, 2137–2147 10.1126/science.284.5423.2137 (doi:10.1126/science.284.5423.2137) [DOI] [PubMed] [Google Scholar]
- 7.Nesbitt SJ, Sidor CA, Irmis RB, Angielczyk KD, Smith RMH, Tsuji LA. 2010. Ecologically distinct dinosaurian sister-group shows early diversification of Ornithodira. Nature 464, 95–98 10.1038/nature08718 (doi:10.1038/nature08718) [DOI] [PubMed] [Google Scholar]
- 8.Benton MJ. 1986. The Late Triassic reptile Teratosaurus a rauisuchian not a dinosaur. Palaeontology 29, 293–301 [Google Scholar]
- 9.Rubidge BS. 2005. Re-uniting lost continents: fossil reptiles from the ancient Karoo and their wanderlust. S. Afr. J. Geol. 108, 135–172 10.2113/108.1.135 (doi:10.2113/108.1.135) [DOI] [Google Scholar]
- 10.Langer MC, Benton MJ. 2006. Early dinosaurs: a phylogenetic study. J. Syst. Palaeontol. 4, 309–358 10.1017/S1477201906001970 (doi:10.1017/S1477201906001970) [DOI] [Google Scholar]
- 11.Piechowski R, Dzik J. 2010. The axial skeleton of Silesaurus opolensis. J. Vertebr. Paleontol. 30, 1127–1141 10.1080/02724634.2010.483547 (doi:10.1080/02724634.2010.483547) [DOI] [Google Scholar]
- 12.Benson RBJ, Butler RJ, Carrano MT, O'Connor MT. 2012. Air-filled postcranial bones in theropod dinosaurs: physiological implications and the ‘reptile’: bird transition. Biol. Rev. 87, 168–193 10.1111/j.1469-185X.2011.00190.x (doi:10.1111/j.1469-185X.2011.00190.x) [DOI] [PubMed] [Google Scholar]
- 13.de Ricqlès AJ, Padian K, Horner JR. 2003. On the bone histology of some Triassic pseudosuchian archosaurs and related taxa. Ann. Paleontol. 89, 67–101 10.1016/S0753-3969(03)00005-3 (doi:10.1016/S0753-3969(03)00005-3) [DOI] [Google Scholar]
- 14.Padian K, Horner JR, de Ricqlès A. 2003. Growth in small dinosaurs and pterosaurs: the evolution of archosauria growth strategies. J. Vertebr. Paleontol. 24, 555–571 10.1671/0272-4634(2004)024[0555:GISDAP]2.0.CO;2 (doi:10.1671/0272-4634(2004)024[0555:GISDAP]2.0.CO;2) [DOI] [Google Scholar]
- 15.Fostowicz-Frelik Ł., Sulej T. 2010. Bone histology of Silesaurus opolensis Dzik, 2003 from the Late Triassic of Poland. Lethaia 43, 137–148 10.1111/j.1502-3931.2009.00179.x (doi:10.1111/j.1502-3931.2009.00179.x) [DOI] [Google Scholar]
- 16.Nesbitt SJ. 2011. The early evolution of Archosauria: relationships and the origin of major clades. Bull. Am. Mus. Nat. Hist. 352, 1–292 10.1206/352.1 (doi:10.1206/352.1) [DOI] [Google Scholar]
- 17.Butler RJ, Upchurch P, Norman DB. 2008. The phylogeny of the ornithischian dinosaurs. J. Syst. Palaeontol. 6, 1–40 10.1017/S1477201907002271 (doi:10.1017/S1477201907002271) [DOI] [Google Scholar]
- 18.Butler RJ, Barrett PM, Gower DJ. 2009. Postcranial skeletal pneumaticity and air-sacs in the earliest pterosaurs. Biol. Lett. 5, 557–560 10.1098/rsbl.2009.0139 (doi:10.1098/rsbl.2009.0139) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bishop WW. 1968. The evolution of fossil environments in East Africa. Trans. Leicestershire Literary Philos. Soc. 62, 22–44 [Google Scholar]