

Abstract
Mean trophic level (MTL) of landings and primary production required (PPR) by fisheries are increasingly used in the assessment of sustainability in fisheries. However, in their present form, MTL and PPR are prone to misinterpretation. We show that it is important to account for actual catch data, define an appropriate historical and spatial domain, and carefully consider the effects of fisheries management, based on results from a case study of Swedish fisheries during the past century.
Keywords: discard, fisheries, management, MTL, PPR, seafood
1. Introduction
Understanding trophic interactions and how fisheries affect them is essential for management of fisheries [1–3]. Indicators related to a species' position in the food web are accordingly used in several ways in attempts to quantify sustainability of fisheries.
One of these indicators is the mean trophic level (MTL) of landings, intended to represent, and account for, the phenomenon of ‘fishing down the food web’ [4]. If fishing pressure depletes top predators more severely than low-trophic-level species, a decline in the MTL of fisheries landings can be expected. This concept has been adopted by the Convention of Biological Diversity (CBD) (i.e. to be ‘ready for global use’ [5]).
Factors determining the trophic composition of landings, however, are complex and difficult to disentangle. For example, a decrease in MTL can be caused by an increase in the contribution of low-trophic-level species to landings rather by depletion of top predators [6]. Moreover, top predators are not the main driver behind fishing revenues as often assumed previously [7], and there is evidence that low-trophic-level species collapse more frequently than do top predators [8]. In fact, the negative trend in global MTL observed at the end of the 1990s is no longer supported by the most recent two decades of data [9]. Given the unresolved complexities in the factors and mechanisms that determine MTL of fishery landings at different scales [10] further evaluation of MTL as an indicator of sustainable fishery exploitation is needed.
The primary production required (PPR) by fisheries is an indicator that is closely related to MTL. PPR represents an estimate of the carbon used by photosynthesis to produce one kilogramme of biomass in the population of a species at a certain trophic level (TL) [11]. The present global rate of biomass removal by fisheries in terms of PPR is thought to exceed the limits required for long-term sustainable marine ecosystem production [12]. Lower PPR values would accordingly be associated with lower ecosystem costs. PPR has been suggested as a common currency or ecological footprint that enables comparison of the ecological cost of fishing over time or between ecosystems [13,14]. PPR is also increasingly applied in environmental systems analysis of seafood production; in this case it serves as a measure of biological resource use from aquaculture or fisheries [15,16].
We have studied the patterns of MTL and PPR using data that represent over a century of Swedish fishing in the Kattegat and Skagerrak and related these results to other sources of information on fisheries development and ecosystem dynamics for this area. Our intent is to evaluate the strengths and weaknesses of MTL and PPR in detecting trends related to fishing pressure on ecosystem functioning and on their potential use as indicators of sustainability for fisheries and their management.
2. Material and methods
In this study, we used Swedish landings data from the International Council for Exploration of the Sea (ICES; area IIIa, years 1903–2010). Because data from IIIa were pooled with IIIc or IVb + c in 1932–1933 and 1962–1974, these years were excluded from consideration. Estimates of TLs were obtained from FishBase and SeaAroundUs and assumed to be invariant over time (see [17] for constraints on these assumptions). Landings with insufficient species-specific information were excluded (on average 2% of the total biomass). For details see the electronic supplementary material, S1.
PPR was estimated as in [11], by assuming a conservative 9 : 1 conversion ratio of wet weight to carbon:
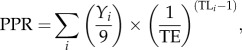 |
where Yi is the yield for species i (measured as landings) with trophic level TLi, and transfer efficiency TE (assumed to be 14% in this study, as it is higher than the standard 10% TE in northerly regions [12]). TE was assumed to be constant during the time period of this study.
The MTL was estimated as
for each year, and Y represents the yield from species i.
Fishery-independent MTL was calculated using catch-per-unit-effort data from the International Bottom Trawl Survey (IBTS, quarter 1, 1979–2010; electronic supplementary material, S2). Our study also included analysis of the PPR and MTL of actual catch data (landings and discard) from pre-separated fishing segments for one year (2009), including all fish and commercial invertebrates (for details see electronic supplementary material, S3). Estimates of primary production for 1985–2010 (PP, mean mC*m2*year) were provided by the Swedish Meteorological and Hydrological Institute (SMHI).
3. Results
A progressive increase in the total quantity of landings is seen until the end of the 1990s, followed by a sharp decline (figure 1a). Initially, gadoids contributed more to the landings volume, but at the end of the time covered by these data, shrimp and small pelagic fishes were more dominant (see the electronic supplementary material, S4). A breakpoint regression analysis revealed an increasing trend in MTL prior to the 1930s, followed by a decreasing trend (figure 1b). Trends in MTL and volume of landings exhibit independent patterns during the entire period, with both variables declining in recent years (see the electronic supplementary material, S5).
Figure 1.

(a) Landings by Swedish fisheries in mass per TL (ICES area IIIa), data for 1962–1974 are lacking. The species dominating TL, less than 2.99 were invertebrates; TL, 3–3.49 was dominated by sprat and herring; for TL, 3.5–4 it was mackerel; and for TL more than 4 gadoids dominated. The large drop in landings in 1996 is related to changes in quota access at the time Sweden joined the European Union. (b) Trends observed in MTL, and (c) trends in PPR are for the same set of data as shown in (a).
PPR followed a pattern similar to that exhibited by MTL, but the breakpoint was more sensitive to the number of iterations in the analysis, finally stabilizing in the 1990s (figure 1c). Combining the two, it can be seen that, over the past two decades, landings exhibit both low MTL and PPR relative to the total available ecosystem production (% PPR) (figure 2a), with occasional peaks due to increased landings of blue whiting (Micromesistius poutassou).
Figure 2.
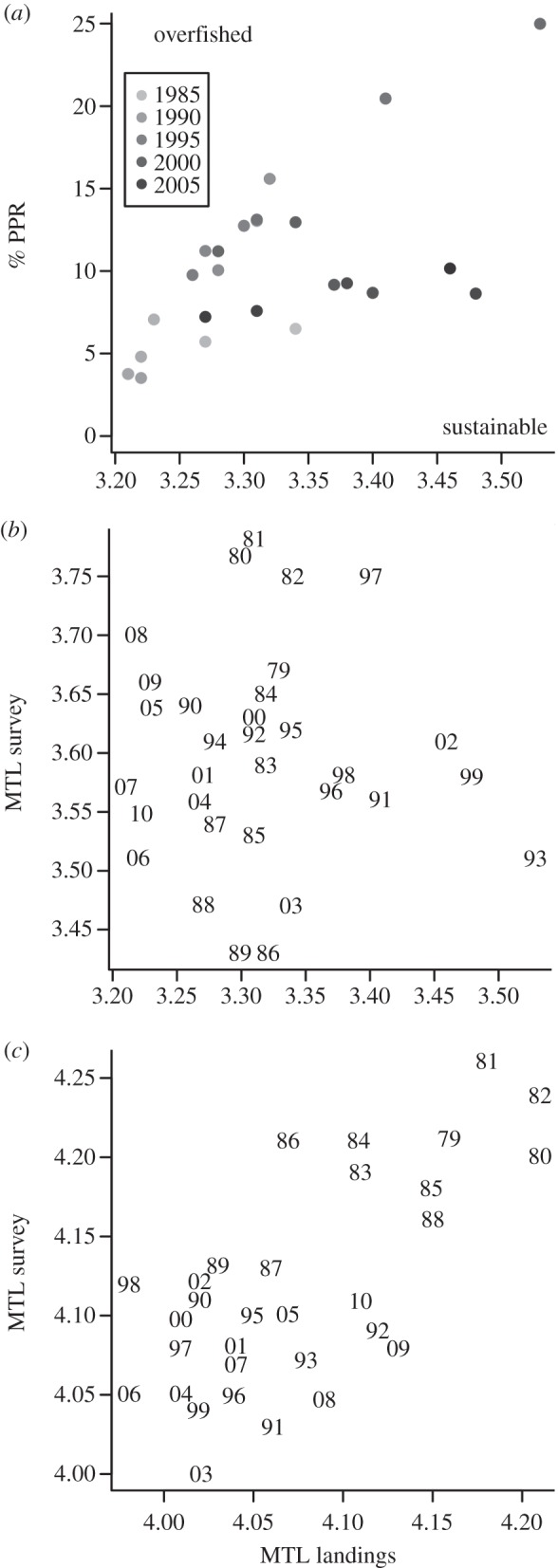
In (a) landings are expressed as % PPR out of total ecosystem production plotted against MTL, format adopted from [2]. In (b), MTL based on survey data (1979–2010) are compared with landings (fish and commercial crustaceans) and in (c) MTL for species with a TL more than 3.25 are compared with landings using the same data as shown in (b).
We found no correlation between the MTL of landings and that of survey data for fish and commercial crustacean species (r2 = 0.001, p < 0.843; figure 2b). However, with only species above TL 3.25 in the sample, there was a weak correlation (r2 = 0.448, p < 0.001; figure 2c). This is explained on the grounds that herring, the major contributor to catches, is removed when calculating MTL for species above 3.25.
On a more detailed scale than represented by figures 1 and 2, landings represented a highly variable part of the actual catch PPR (i.e. including discards) depending on the fishing segment, where the PPR from landings ranged between 22 and 83 per cent of the total catch PPR of the fishery (see the electronic supplementary material, S3). Likewise, MTL also differed between the total catch and the landed portion, ranging between 2.50 and 4.18 for the total catch) (see the electronic supplementary material, S3).
Overall, measures of the CPUE showed a positive trend for all species in the survey data (including non-commercial species; r2 = 0.400, p < 0.001; electronic supplementary material, S2).
4. Discussion
The major fishing pattern behind the trend in MTL within the areas of the Kattegat and Skagerrak is a reduction in the contribution of stocks of large predator fish to landings, consistent with earlier reports [18,19]. Taken together, the observed peak of small pelagics in the 1990s and the decreased contribution from top predators in recent years favour a ‘fishing through’ scenario [6] in which lower TLs are increasingly exploited. However, this trend itself does not necessarily indicate that fishing practices at present are unsustainable. In part, this is because recent decreases in landing MTL are highly influenced by management efforts aiming at protecting and rebuilding gadoid stocks [20,21]. Furthermore, there was no correlation between landings and survey data MTL (unless the low-trophic-level species are ignored). The trend in landing MTL appears to be, at most, a weak measure of the ecosystem state and pressures on biodiversity in the area. From this, we conclude that inferences concerning global fishing mortality and abundance trends in top predators using relative patterns in aggregate MTL are difficult to interpret without the consideration of actual total catch data (including discards or survey data).
A clear conclusion from our work is that PPR estimates based on data restricted to landings are inadequate and possibly misleading. Including PPR of discards is in fact essential to enable evaluations of the ecological costs from different fishing practices [20]. Nevertheless, before making detailed comparisons between regions, further refinement of appropriate values for TEs might be needed [22].
It is additionally clear that care must be taken in interpreting data involving estimated PPR. For example, declining PPR from fisheries can be erroneously interpreted as a fishery with decreasing costs to the ecosystem. However, low PPR values in the Kattegat and Skagerrak may very well involve commercial landings at an all-time low because of commercial stocks that are severely depleted, which cannot be interpreted as advantageous. Regarding other metrics, survey data in our study do not indicate a lower overall production in the area. Such indications of good health can also be misleading when considered alone, as other measures could easily indicate serious ecosystem-level problems caused by the synergistic effects of overfishing and eutrophication [23–26]. Therefore, before any fisheries indices related to trophic interactions can be interpreted properly, it is essential to have a much better understanding of which factors contribute to local fish production [27] and how fishing affects the dynamics of energy within ecosystems [3].
Complex systems involve complex sets of interactions, and it is most probable that a large suite of indicators is needed to assess the sustainability of fisheries; single trends in PPR or MTL could send conflicting messages. Decreases in MTL (interpreted as a negative signal by the CBD) can contribute to decreasing PPR (considered as a necessary transition towards more sustainable fisheries [12,14] and as a positive factor in the environmental systems analysis of seafood [15]).
To conclude, there are serious constraints on the conclusions that can be drawn from information on trends and levels of estimated MTL and PPR. As one significant step towards improvement, we strongly recommend that future efforts use actual catch data (including discards/surveys) and ecologically sound spatial resolution to account for obvious differences among different regions (e.g. fisheries management systems and ecosystem functioning), and that they take into account temporal factors, especially the influence attributable to the duration of fisheries exploitation.
Acknowledgements
We thank Katja Ringdahl and Barbara Bland for assisting with data and Per Nilsson, Leif Pihl, Andreas Emanuelsson, Owen Petchey, Chuck Fowler and two anonymous reviewers for helpful suggestions for improvements of this paper. This work was funded by FORMAS—The Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning and the European Commission (Grant Agreement 243827).
References
- 1.Casini M, Hjelm J, Molinero JC, Lövgren J, Cardinale M, Bartolino V, Belgrano A, Kornilovs G. 2009. Trophic cascades promote threshold-like shifts in pelagic marine ecosystems. Proc. Natl. Acad. Sci. USA 106, 197–202 10.1073/pnas.0806649105 (doi:10.1073/pnas.0806649105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Conti L, Scardi M. 2010. Fisheries yield and primary productivity in large marine ecosystems. Mar. Ecol. Progr. Ser. 410, 233–244 10.3354/meps08630 (doi:10.3354/meps08630) [DOI] [Google Scholar]
- 3.Frank KT, Petrie B, Fisher JAD, Leggett WC. 2011. Transient dynamics of an altered large marine ecosystem. Nature 477, 86–89 10.1038/nature10285 (doi:10.1038/nature10285) [DOI] [PubMed] [Google Scholar]
- 4.Pauly D, Christensen V, Dalsgaard J, Froese R, Torres F., Jr 1998. Fishing down marine food webs. Science 279, 860–863 10.1126/science.279.5352.860 (doi:10.1126/science.279.5352.860) [DOI] [PubMed] [Google Scholar]
- 5.2010 Biodiversity Indicators Partnership 2010. Biodiversity indicators and the 2010 Target: Experiences and lessons learnt from the 2010 Biodiversity Indicators Partnership. Secretariat of the Convention on Biological Diversity, Montreal, Canada. Technical Series no. 53, pp. 196
- 6.Essington TE, Beaudreau AH, Wiedenmann J. 2006. Fishing through marine food webs. Proc. Natl Acad. Sci. USA 103, 3171–3175 10.1073/pnas.0510964103 (doi:10.1073/pnas.0510964103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sethi SA, Branch TA, Watson R. 2010. Fishery development patterns are driven by profit but not trophic level. Proc. Natl Acad. Sci. USA 107, 12 163–12 167 10.1073/pnas.1003236107 (doi:10.1073/pnas.1003236107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pinsky ML, Jensen OP, Ricard D, Palumbi SR. 2011. Unexpected patterns of fisheries collapse in the world's oceans. Proc. Natl Acad. Sci. USA 108, 8317–8322 10.1073/pnas.1015313108 (doi:10.1073/pnas.1015313108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Branch TA, Watson R, Fulton EA, Jennings S, McGilliard CR, Pablico GT, Ricard D, Tracey SR. 2010. The trophic fingerprint of marine fisheries. Nature 468, 431–435 10.1038/nature09528 (doi:10.1038/nature09528) [DOI] [PubMed] [Google Scholar]
- 10.Stergiou KI, Tsikliras AC. 2011. Fishing down, fishing through and fishing up: fundamental process versus technical details. Mar. Ecol. Progr. Ser. 441, 295–301 10.3354/meps09377 (doi:10.3354/meps09377) [DOI] [Google Scholar]
- 11.Pauly D, Christensen V. 1995. Primary production required to sustain global fisheries. Nature 374, 255–257 10.1038/374255a0 (doi:10.1038/374255a0) [DOI] [Google Scholar]
- 12.Coll M, Libralato S, Tudela S, Palomera I, Pranovi F. 2008. Ecosystem overfishing in the ocean. PLoS ONE 3, e3881. 10.1371/journal.pone.0003881 (doi:10.1371/journal.pone.0003881) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.ICES 2005. Ecosystems effects of fishing: impacts, metrics, and management strategies. ICES Cooperative Research Report, no. 272, 177 pp
- 14.Swartz W, Sala E, Tracey S, Watson R, Pauly D. 2010. The spatial expansion and ecological footprint of fisheries (1950 to Present). PLoS ONE 5, e15143. 10.1371/journal.pone.0015143 (doi:10.1371/journal.pone.0015143) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pelletier N, Tyedmers P, Sonesson U, Scholz A, Ziegler F, Flysjo A, Kruse S, Cancino B, Silverman H. 2009. Not all salmon are created equal: life cycle assessment (LCA) of global salmon farming systems. Environ. Sci. Technol. 43, 8730–8736 10.1021/es9010114 (doi:10.1021/es9010114) [DOI] [PubMed] [Google Scholar]
- 16.Vázquez-Rowe I, Moreira M, Feijoo G. 2012. Inclusion of discard assessment indicators in fisheries life cycle assessment studies. Expanding the use of fishery-specific impact categories. Int. J. Life Cycle Assess. 17, 535–549 10.1007/s11367-012-0395-x (doi:10.1007/s11367-012-0395-x) [DOI] [Google Scholar]
- 17.Jennings S, Greenstreet SPR, Hill L, Piet GJ, Pinnegar JK, Warr KJ. 2002. Long-term trends in the trophic structure of the North Sea fish community: evidence from stable-isotope analysis, size-spectra and community metrics. Mar. Biol. 141, 1085–1097 10.1007/s00227-002-0905-7 (doi:10.1007/s00227-002-0905-7) [DOI] [Google Scholar]
- 18.Cardinale M, Hagberg J, Svedang H, Bartolino V, Gedamke T, Hjelm J, Börjesson P, Norén F. 2010. Fishing through time: population dynamics of plaice (Pleuronectes platessa) in the Kattegat–Skagerrak over a century. Popul. Ecol. 52, 251–262 10.1007/s10144-009-0177-x (doi:10.1007/s10144-009-0177-x) [DOI] [Google Scholar]
- 19.Bartolino V, Cardinale M, Svedäng H, Linderholm HW, Casini M, Grimwall A. 2012. Historical spatiotemporal dynamics of eastern North Sea cod. Can. J. Fish. Aquat. Sci. 69, 833–841 10.1139/f2012-028 (doi:10.1139/f2012-028) [DOI] [Google Scholar]
- 20.Hornborg S, Nilsson P, Valentinsson D, Ziegler F. 2012. Integrated environmental assessment of fisheries management: Swedish Nephrops trawl fisheries evaluated with a life cycle approach. Mar. Pol. 36, 1193–1201 10.1016/j.marpol.2012.02.017 (doi:10.1016/j.marpol.2012.02.017) [DOI] [Google Scholar]
- 21.Catchpole TL, Tidda AN, Kell LT, Revilla AS, Dunlinb G. 2007. The potential for new Nephrops trawl designs to positively effect North Sea stocks of cod, haddock and whiting. Fish. Res. 86, 262–267 10.1016/j.fishres.2007.06.023 (doi:10.1016/j.fishres.2007.06.023) [DOI] [Google Scholar]
- 22.Baumann M. 1995. A comment on transfer efficiencies. Fish. Oceanogr. 4, 264–266 10.1111/j.1365-2419.1995.tb00150.x (doi:10.1111/j.1365-2419.1995.tb00150.x) [DOI] [Google Scholar]
- 23.Pihl L. 1994. Changes in the diet of demersal fish due to eutrophication-induced hypoxia in the Kattegat, Sweden. Can. J. Fish. Aquat. Sci. 51, 321–336 10.1139/f94-033 (doi:10.1139/f94-033) [DOI] [Google Scholar]
- 24.Diaz RJ, Rosenberg R. 2008. Spreading dead zones and consequences for marine ecosystems. Science 321, 926–929 10.1126/science.1156401 (doi:10.1126/science.1156401) [DOI] [PubMed] [Google Scholar]
- 25.Baden S, Emanuelsson A, Pihl L, Svensson CJ, Åberg P. 2012. Shift in seagrass food web structure over decades is linked to overfishing. Mar. Ecol. Progr. Ser. 451, 61–73 10.3354/meps09585 (doi:10.3354/meps09585) [DOI] [Google Scholar]
- 26.Nielsen E, Richardson K. 1996. Can changes in the fisheries yield in the Kattegat (1953–1992) be linked to changes in primary production? ICES J. Mar. Sci. 53, 988–994 10.1006/jmsc.1996.0123 (doi:10.1006/jmsc.1996.0123) [DOI] [Google Scholar]
- 27.Hunt GL, Jr, McKinnel S. 2006. Interplay between top-down, bottom-up, and wasp-waist control in marine ecosystems. Progr. Oceanogr. 68, 115–124 10.1016/j.pocean.2006.02.008 (doi:10.1016/j.pocean.2006.02.008) [DOI] [Google Scholar]