

Abstract
Gillisia limnaea Van Trappen et al. 2004 is the type species of the genus Gillisia, which is a member of the well characterized family Flavobacteriaceae. The genome of G. limnea R-8282T is the first sequenced genome (permanent draft) from a type strain of the genus Gillisia. Here we describe the features of this organism, together with the permanent-draft genome sequence and annotation. The 3,966,857 bp long chromosome (two scaffolds) with its 3,569 protein-coding and 51 RNA genes is a part of the Genomic Encyclopedia of Bacteria and Archaea project.
Keywords: aerobic, motile, rod-shaped, moderately halotolerant, psychrophilic, chemoheterotrophic, proteorhodopsin, microbial mat, yellow-pigmented, Flavobacteriaceae, GEBA
Introduction
Strain R-8282T (= DSM 15749 = LMG 21470 = CIP 108418) is the type strain of the species Gillisia limnaea [1], which in turn is the type species of the Gillisia, a genus currently encompassing six known species [1]. The strain was isolated from a microbial mat in Lake Fryxell, Antarctica [1] during the MICROMAT project, which systematically collected novel strains from Antarctic lakes [2]. The genus was named after the Belgian bacteriologist Monique Gillis for her work on bacterial taxonomy [1]. The species epithet was derived from the Neo-Latin adjective ‘limnaeae’, living in the water, referring to the microbial mats in Lake Fryxell where the organism was first isolated [1]. PubMed records do not indicate any follow-up research with strain R-8282T after the initial description and valid publication of the new species name G. limnaea, and genus Gillisia [1]. Here we present a summary classification and a set of features for G. limnaea R-8282T, together with the description of the genomic sequencing and annotation.
Classification and features
A representative genomic 16S rRNA sequence of G. limnaea R-8282T was compared using NCBI BLAST [3,4] under default settings (e.g., considering only the high-scoring segment pairs (HSPs) from the best 250 hits) with the most recent release of the Greengenes database [5] and the relative frequencies of taxa and keywords (reduced to their stem [6]) were determined, weighted by BLAST scores. The most frequently occurring genera were Flavobacterium (80.2%), Gillisia (17.8%), Chryseobacterium (1.0%) and Cytophaga (1.0%) (94 hits in total). Regarding the single hit to sequences from members of the species, the average identity within HSPs was 99.1%, whereas the average coverage by HSPs was 98.2%. Regarding the five hits to sequences from other members of the genus, the average identity within HSPs was 95.6%, whereas the average coverage by HSPs was 94.3%. Among all other species, the one yielding the highest score was Gillisia hiemivivida (AY694006), which corresponded to an identity of 97.1% and an HSP coverage of 90.8%. (Note that the Greengenes database uses the INSDC (= EMBL/NCBI/DDBJ) annotation, which is not an authoritative source for nomenclature or classification.) The highest-scoring environmental sequence was EU735617 (Greengenes short name: 'archaeal structures and pristine soils China oil contaminated soil Jidong Oilfield clone SC78'), which showed an identity of 99.0% and an HSP coverage of 98.4%. The most frequently occurring keywords within the labels of all environmental samples which yielded hits were 'librari' (3.2%), 'dure' (3.0%), 'bioremedi, broader, chromat, groundwat, microarrai, polylact, sampl, stimul, subsurfac, typic, univers' (2.9%), 'spring' (2.5%) and 'soil' (2.4%) (156 hits in total). The most frequently occurring keywords within the labels of those environmental samples which yielded hits of a higher score than the highest scoring species were 'soil' (15.4%), 'archaeal, china, contamin, jidong, oil, oilfield, pristin, structur' (7.7%) and 'antarct, cover, lake' (7.7%) (2 hits in total). Whereas some of these keywords confirm the environment of G. limnaea, others are indicative of other habitats in which related taxa are found.
Figure 1 shows the phylogenetic neighborhood of G. limnaea in a 16S rRNA based tree. The sequences of the two 16S rRNA gene copies in the genome differ from each other by up to eleven nucleotides, and differ by up to eight nucleotides from the previously published 16S rRNA sequence (AJ440991), which contains seven ambiguous base calls.
Figure 1.

Phylogenetic tree highlighting the position of G. limnaea relative to the type strains of the type species of the genera within the family Flavobacteriaceae. The tree was inferred from 1,366 aligned characters [7,8] of the 16S rRNA gene sequence under the maximum likelihood (ML) criterion [9]. Rooting was done initially using the midpoint method [10] and then checked for its agreement with the current classification (Table 1). The branches are scaled in terms of the expected number of substitutions per site. Numbers adjacent to the branches are support values from 1,000 ML bootstrap replicates [11] (left) and from 1,000 maximum-parsimony bootstrap replicates [12] (right) if larger than 60%. Lineages with type strain genome sequencing projects registered in GOLD [13] are labeled with one asterisk, those also listed as 'Complete and Published' with two asterisks [14-16]; for Ornithobacterium rhinotracheale see CP003283)
Table 1. Classification and general features of G. limnaea G-8282T according to the MIGS recommendations [17] and NamesforLife [18].
| MIGS ID | Property | Term | Evidence code |
|---|---|---|---|
| Domain Bacteria | TAS [19] | ||
| Phylum Bacteroidetes | TAs [20,21] | ||
| Class Flavobacteria | TAS [22-24] | ||
| Current classification | Order Flavobacteriales | TAS [21,25] | |
| Family Flavobacteriaceae | TAS [26-29] | ||
| Genus Gillisia | TAS [1] | ||
| Species Gillisia limnaea | TAS [1] | ||
| Type strain R-8282 | TAS [1] | ||
| Gram stain | negative | TAS [1] | |
| Cell shape | rod-shaped | TAS [1] | |
| Motility | gliding motility likely, but not proven | NAS | |
| Sporulation | none | TAS [1] | |
| Temperature range | psychrophile, 5-30°C | TAS [1] | |
| Optimum temperature | 20°C | TAS [1] | |
| Salinity | 0-5% NaCl (w/v) | TAS [1] | |
| MIGS-22 | Oxygen requirement | aerobe | TAS [1] |
| Carbon source | yeast extract, peptone | TAS [1] | |
| Energy metabolism | chemoheterotrophic, phototrophic | TAS [1] | |
| MIGS-6 | Habitat | fresh water | TAS [1] |
| MIGS-15 | Biotic relationship | free living | TAS [1] |
| MIGS-14 | Pathogenicity | none | NAS |
| Biosafety level | 1 | TAS [30] | |
| MIGS-23.1 | Isolation | microbial mats | TAS [1] |
| MIGS-4 | Geographic location | Lake Fryxell, McMurdo Dry Valleys, Antarctica | TAS [1] |
| MIGS-5 | Sample collection time | between November 1998 and February 2001 | TAS [1,2] |
| MIGS-4.1 | Latitude | -77.614 | NAS |
| MIGS-4.2 | Longitude | 163.184 | NAS |
| MIGS-4.3 | Depth | not reported | |
| MIGS-4.4 | Altitude | not reported |
Evidence codes - TAS: Traceable Author Statement (i.e., a direct report exists in the literature); NAS: Non-traceable Author Statement (i.e., not directly observed for the living, isolated sample, but based on a generally accepted property for the species, or anecdotal evidence). Evidence codes are from the Gene Ontology project [31].
Cells of strain G. limnaea R-8282T are Gram-negative and rod-shaped [Figure 2] [1]. They are 0.7 µm in width and 3.0 µm in length [1], whereas scanning electron micrographs of strain R-8282T revealed a cell diameter that varies from 0.4 µm to 0.5 µm, and a length that varies from 1.6 µm to longer than 4.9 µm [Figure 2], which is more consistent with data previously reported for several Gillisia strains [32-34]. Motility, especially gliding motility, was not observed [1], despite the presence of numerous genes associated with gliding motility (see below), and the presence of pili-containing cells in scanning electron micrographs of strain R-8282T. It is unclear if these pili are involved in gliding motility or bacterial adhesion to surfaces. Cells are strictly aerobic, psychrophilic and chemoheterotrophic [1]. Growth occurs between 5°C and 30°C with an optimum at 20°C [1]; the strain is unable to grow at temperatures above 37°C [1]. Growth occurs within a salinity range of 0% to 5% NaCl, but not in 10% NaCl, indicating moderate halotolerance [1]. Peptone and yeast extract were required for growth [1]. When cultivated on marine agar, colonies are yellow in color, convex and translucent with diameters of 1-3 mm forming entire margins after 6 days of incubation [1]. When cultivated on Anacker & Ordal’s agar, colonies become flat and round with entire margins and 0.7 to 0.9 mm in diameter after 14 days incubation [1]. Additionally growth is both detectable on nutrient agar and R2A, but the strain does not grow on trypticase soy agar [1]. Further detailed physiological data such as carbon source utilization, carbon degradation, and enzyme activities have been reported previously [1].
Figure 2.

Scanning electron micrograph of G. limnaea R-8282T
Chemotaxonomy
The principal cellular fatty acids of strain R-8282T are iso-C15:0, anteiso-C15:0, iso-C15:1, iso-C16:0, C17:0 2-OH, iso-C17:0 3-OH, iso-C17:1 ω9c, anteiso-C17:1 ω9c and summed feature 3 (containing iso-C15:0 2-OH and/or C16:1 ω7c) [1]. The major polar lipids were not reported for strain R-8282T.
Genome sequencing and annotation
Genome project history
This organism was selected for sequencing on the basis of its phylogenetic position [35], and is part of the Genomic Encyclopedia of Bacteria and Archaea project [36]. The genome project is deposited in the Genomes On Line Database [13] and the complete genome sequence is deposited in GenBank. Sequencing, finishing and annotation were performed by the DOE Joint Genome Institute (JGI). A summary of the project information is shown in Table 2.
Table 2. Genome sequencing project information.
| MIGS ID | Property | Term |
|---|---|---|
| MIGS-31 | Finishing quality | Non-contiguous |
| MIGS-28 | Libraries used | Four genomic libraries: one 454 pyrosequence standard library, two 454 PE libraries (4 kb and 10 kb insert size), one Illumina library |
| MIGS-29 | Sequencing platforms | Illumina GAii, 454 GS FLX Titanium |
| MIGS-31.2 | Sequencing coverage | 309.5 × Illumina; 36.0 × pyrosequence |
| MIGS-30 | Assemblers | Newbler version 2.3, Velvet 1.0.13, phrap version SPS - 4.24 |
| MIGS-32 | Gene calling method | Prodigal |
| INSDC ID | PAHKR00000000 | |
| GenBank Date of Release | January 24, 2012 | |
| GOLD ID | Gc04190 | |
| NCBI project ID | 50579 | |
| Database: IMG-GEBA | 2506783053 | |
| MIGS-13 | Source material identifier | DSM 15749 |
| Project relevance | Tree of Life, GEBA |
Growth conditions and DNA isolation
G. limnaea strain R-8282T, DSM 15749, was grown in DSMZ medium 514 (BACTO Marine Broth) [37] at 20°C. DNA was isolated from 0.5-1 g of cell paste using MasterPure Gram Positive DNA Purification kit (Epicentre MGP04100) following the standard protocol as recommended by the manufacturer with modification st/DL as described by Wu et al. 2009 [36] for optimized cell lysis. DNA is available through the DNA Bank Network [38].
Genome sequencing and assembly
The genome was sequenced using a combination of Illumina and 454 sequencing platforms. All general aspects of library construction and sequencing can be found at the JGI website [39]. Pyrosequencing reads were assembled using the Newbler assembler (Roche). The initial Newbler assembly consisting of 93 contigs in one scaffold was converted into a phrap [40] assembly by making fake reads from the consensus, to collect the read pairs in the 454 paired end library. Illumina GAii sequencing data (1,096.5Mb) was assembled with Velvet [41] and the consensus sequences were shredded into 1.5 kb overlapped fake reads and assembled together with the 454 data. The 454 draft assembly was based on 178.7 Mb 454 draft data and all of the 454 paired end data. Newbler parameters are -consed -a 50 -l 350 -g -m -ml 20. The Phred/Phrap/Consed software package [40] was used for sequence assembly and quality assessment in the subsequent finishing process. After the shotgun stage, reads were assembled with parallel phrap (High Performance Software, LLC). Possible mis-assemblies were corrected with gapResolution [39], Dupfinisher [42], or sequencing cloned bridging PCR fragments with subcloning. Gaps between contigs were closed by editing in Consed, by PCR and by Bubble PCR primer walks (J.-F. Chang, unpublished). A total of 893 additional reactions and one shatter library were necessary to close gaps and to raise the quality of the final sequence. Illumina reads were also used to correct potential base errors and increase consensus quality using the software Polisher developed at JGI [43]. The error rate of the final genome sequence is less than 1 in 100,000. Together, the combination of the Illumina and 454 sequencing platforms provided 127.9 x coverage of the genome. The final assembly contained 597,282 pyrosequence and 33,599,185 Illumina reads.
Genome annotation
Genes were identified using Prodigal [44] as part of the Oak Ridge National Laboratory genome annotation pipeline, followed by a round of manual curation using the JGI GenePRIMP pipeline [45]. The predicted CDSs were translated and used to search the National Center for Biotechnology Information (NCBI) non-redundant database, UniProt, TIGRFam, Pfam, PRIAM, KEGG, COG, and InterPro databases. These data sources were combined to assert a product description for each predicted protein. Additional gene prediction analysis and functional annotation was performed within the Integrated Microbial Genomes - Expert Review (IMG-ER) platform [46].
Genome properties
The genome consists of two scaffolds with 3,558,876 bp and 407,981 bp length, respectively, with a G+C content of 37.6% (Table 3 and Figure 3). Of the 3,620 genes predicted, 3,569 were protein-coding genes, and 51 RNAs; 135 pseudogenes were also identified. The majority of the protein-coding genes (66.0%) were assigned a putative function while the remaining ones were annotated as hypothetical proteins. The distribution of genes into COGs functional categories is presented in Table 4.
Table 3. Genome Statistics.
| Attribute | Value | % of Total |
|---|---|---|
| Genome size (bp) | 3,966,857 | 100.00% |
| DNA coding region (bp) | 3,414,922 | 85.09% |
| DNA G+C content (bp) | 1,490,901 | 37.61% |
| Number of scaffolds | 2 | |
| Total genes | 3,620 | 100.00% |
| RNA genes | 51 | 1.41% |
| rRNA operons | 1* | |
| tRNA genes | 44 | 1.22% |
| Protein-coding genes | 3,569 | 98.59% |
| Pseudo genes | 135 | 3.73% |
| Genes with function prediction (proteins) | 2,388 | 65.97% |
| Genes in paralog clusters | 1,727 | 47.71% |
| Genes assigned to COGs | 2,489 | 68.76% |
| Genes assigned Pfam domains | 2,606 | 71.99% |
| Genes with signal peptides | 867 | 23.95% |
| Genes with transmembrane helices | 826 | 22.82% |
| CRISPR repeats | 0 |
* one 23S rRNA gene, two 16S rRNA genes
Figure 3.
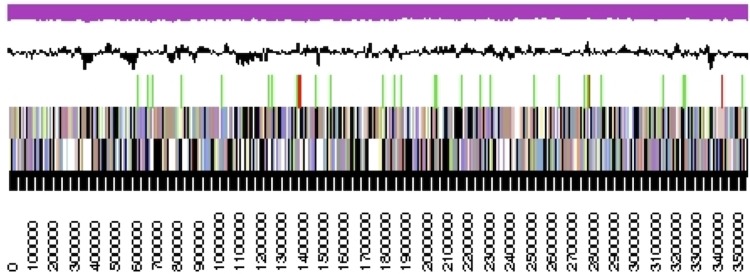
Graphical map of the largest scaffold. From bottom to top: Genes on forward strand (colored by COG categories), Genes on reverse strand (colored by COG categories), RNA genes (tRNAs green, rRNAs red, other RNAs black), GC content(black), GC skew (purple/olive).
Table 4. Number of genes associated with the general COG functional categories.
| Code | value | %age | Description |
|---|---|---|---|
| J | 165 | 6.1 | Translation, ribosomal structure and biogenesis |
| A | 0 | 0.0 | RNA processing and modification |
| K | 148 | 5.5 | Transcription |
| L | 159 | 5.9 | Replication, recombination and repair |
| B | 1 | 0.0 | Chromatin structure and dynamics |
| D | 28 | 1.0 | Cell cycle control, cell division, chromosome partitioning |
| Y | 0 | 0.0 | Nuclear structure |
| V | 70 | 2.6 | Defense mechanisms |
| T | 125 | 4.7 | Signal transduction mechanisms |
| M | 277 | 10.3 | Cell wall/membrane biogenesis |
| N | 12 | 0.5 | Cell motility |
| Z | 0 | 0.0 | Cytoskeleton |
| W | 0 | 0.0 | Extracellular structures |
| U | 49 | 1.8 | Intracellular trafficking and secretion, and vesicular transport |
| O | 105 | 3.9 | Posttranslational modification, protein turnover, chaperones |
| C | 125 | 4.7 | Energy production and conversion |
| G | 146 | 5.4 | Carbohydrate transport and metabolism |
| E | 217 | 8.1 | Amino acid transport and metabolism |
| F | 61 | 2.3 | Nucleotide transport and metabolism |
| H | 131 | 4.9 | Coenzyme transport and metabolism |
| I | 94 | 3.5 | Lipid transport and metabolism |
| P | 131 | 4.9 | Inorganic ion transport and metabolism |
| Q | 59 | 2.2 | Secondary metabolites biosynthesis, transport and catabolism |
| R | 336 | 12.5 | General function prediction only |
| S | 250 | 9.3 | Function unknown |
| - | 1,131 | 31.2 | Not in COGs |
Insights into the genome sequence
Genome analysis of G. limnaea R-8282T revealed the presence of three rhodopsin genes related to proteorhodopsin (PR, GenBank Accession No. EHQ04368, Gilli_0216) and xanthorhodopsin (XR, EHQ02967, Gilli_2340) protein-encoding sequences, whereas a third rhodopsin protein sequence (EHQ02971, Gilli_2344) seems to be truncated. Another finding was a set of genes involved in β-carotene biosynthesis, together with a gene encoding a β-carotene 15,15'-monooxygenase (EHQ04367, Gilli_0215), an enzyme that oxidatively cleaves β-carotene into two molecules of retinal, which is necessary for rhodopsin function. PRs and XRs are photoactive transmembrane opsins that bind retinal and which belong to the microbial rhodopsin superfamily [47]. When exposed to light, a change in protein conformation causes a proton translocation with respect to its cofactor retinal from the inside to the outside of the cell [48]. This proton-pump activity generates a proton motive force across the cell membrane, which can be used in heterologously PR-expressing E. coli cells for ATP synthesis [49] as well as to power general cellular functions like transmembrane nutrient transport or flagella rotation [50]. In contrast to PRs, XRs are light-driven proton pumps containing a dual chromophore: one retinal molecule and one carotenoid antenna [51,52], that was first discovered in Salinibacter ruber M31T [53,54]. Its carotenoid antenna salinixanthin transfers as much as 40-45% of the absorbed photons to retinal [55], resulting in a potentially much more efficient light capturing system compared to PRs from Bacteria [56,57] or bacteriorhodopsins from Archaea [58].
NCBI BLAST analysis [3] revealed that the protein encoded by Gilli_0216 shares distinct identities with many PR protein sequences, found in other species within the Flavobacteriaceae (Figure 4). It shows typical features necessary for proton pump activity: K224 (K231) for retinal-binding, and D88 (D97) as well as E99 (E108) (EBAC31A08 numbering shown in brackets), which act as a proton acceptor and proton donor in the retinylidene Schiff’s base transfer during the PR photocycle [60,61]. Furthermore, the putative PR (Gilli_0216 protein) has a M96 (L105) (EBAC31A08 numbering in parentheses), which mainly indicates that it is a green light-absorbing proteorhodopsin [48,62].
Figure 4.

Rhodopsin tree for Gillisia and relatives. Amino acid sequences were processed in the same way as the 16S rRNA sequences used in Figure 1 except for the explicit determination of an optimal maximum-likelihood model, which turned out to be Lateral Gene Transfer [59]. GenBank Accession Numbers are shown in parentheses.
The gene encoding the putative XR (Gilli_2340) of strain R-8282T shows identities to XR-related proteins, but provides evidence of a new cluster of rhodopsins found in very few flavobacterial isolates like Dokdonia donghaensis PRO95 (EHQ04368) [63] and Krokinobacter sp. 4H-3-7-5 (AEE18495) [64], which was reclassified into the genus Dokdonia [65,66] (Figure 4). This rhodopsin-encoding sequence also reveals typical features necessary for rhodopsin function: K316 (K231) for retinal binding and L181 (L105), which mainly indicates a green-light absorbing rhodopsin [48,62] (EBAC31A08 numbering shown in brackets). But amino acid residues functioning as proton acceptor and proton donor in proteorhodopsin differ from those commonly known. Instead of D97 and E108 (EBAC31A08 numbering), the related amino acids N173 and Q184 are found in the protein sequence encoded by Gilli_2340, which indicates a possible new kind of rhodopsins.
Interestingly, no rhodopsin-encoding sequence could be detected in the genome sequence of Gillisia sp. strain CBA3202 [67], which was isolated from the littoral zone on Jeju Island, Republic of Korea [67]. Digital DNA-DNA hybridization (DDH) [68] between strain R-8282T and CBA3202 revealed an estimate between 9.7% and 13.9% (depending on the formula used), indicating that Gillisia sp. strain CBA3202 does not belong to the species G. limnaea.
Compared to free-living bacteria, representatives of the Bacteroidetes phylum were frequently found attached to aggregates [69] and during an algae-bloom collapse [70,71]. They were also known to move over surfaces by gliding motility [72,73]. In strain R-8282T several genes were detected that are thought to be involved in gliding motility (gldA (Gilli_1140), gldB (Gilli_2923), gldC (Gilli_2942), gldD (Gilli_1840), gldE (Gilli_1841), gldF (Gilli_3447), gldG (Gilli_3446), gldH (Gilli_2158), gldI (Gilli_0258), gldJ (Gilli_1638), gldK (Gilli_2747), gldL (Gilli_2748), gldM (Gilli_2749), gldN (Gilli_2750), espA (Gilli_3049), espB (Gilli_3050), remB (Gilli_2697), sprA (Gilli_2693) and sprE (Gilli_2130)). This observation indicates the possible gliding motility of strain R-8282T, but has never been reported in literature.
Acknowledgements
We would like to gratefully acknowledge the help of Helga Pomrenke for growing G. limnaea cultures and Evelyne-Marie Brambilla for DNA extraction and quality control (both at the DSMZ). This work was performed under the auspices of the US Department of Energy Office of Science, Biological and Environmental Research Program, and by the University of California, Lawrence Berkeley National Laboratory under contract No. DE-AC02-05CH11231, Lawrence Livermore National Laboratory under Contract No. DE-AC52-07NA27344, and Los Alamos National Laboratory under contract No. DE-AC02-06NA25396, UT-Battelle and Oak Ridge National Laboratory under contract DE-AC05-00OR22725, as well as German Research Foundation (DFG) INST 599/1-2 as well as TRR 51.
References
- 1.Van Trappen S, Vandecandelaere I, Mergaert J, Swings J. Gillisia limnaea gen. nov., sp. nov., a new member of the family Flavobacteriaceae isolated from a microbial mat in Lake Fryxell, Antarctica. Int J Syst Evol Microbiol 2004; 54:445-448 10.1099/ijs.0.02922-0 [DOI] [PubMed] [Google Scholar]
- 2.Van Trappen S, Mergaert J, Van Eygen S, Dawyndt P, Cnockaert MC, Swings J. Diversity of 746 heterotrophic bacteria isolated from microbial mats from ten Antarctic lakes. Syst Appl Microbiol 2002; 25:603-610 10.1078/07232020260517742 [DOI] [PubMed] [Google Scholar]
- 3.Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol 1990; 215:403-410 [DOI] [PubMed] [Google Scholar]
- 4.Korf I, Yandell M, Bedell J. BLAST, O'Reilly, Sebastopol, 2003. [Google Scholar]
- 5.DeSantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K, Huber T, Dalevi D, Hu P, Andersen GL. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl Environ Microbiol 2006; 72:5069-5072 10.1128/AEM.03006-05 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Porter MF. An algorithm for suffix stripping. Program: electronic library and information systems 1980; 14:130-137.
- 7.Lee C, Grasso C, Sharlow MF. Multiple sequence alignment using partial order graphs. Bioinformatics 2002; 18:452-464 10.1093/bioinformatics/18.3.452 [DOI] [PubMed] [Google Scholar]
- 8.Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol 2000; 17:540-552 10.1093/oxfordjournals.molbev.a026334 [DOI] [PubMed] [Google Scholar]
- 9.Stamatakis A, Hoover P, Rougemont J. A rapid bootstrap algorithm for the RAxML web servers. Syst Biol 2008; 57:758-771 10.1080/10635150802429642 [DOI] [PubMed] [Google Scholar]
- 10.Hess PN, De Moraes Russo CA. An empirical test of the midpoint rooting method. Biol J Linn Soc Lond 2007; 92:669-674 10.1111/j.1095-8312.2007.00864.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pattengale ND, Alipour M, Bininda-Emonds ORP, Moret BME, Stamatakis A. How many bootstrap replicates are necessary? Lect Notes Comput Sci 2009; 5541:184-200 10.1007/978-3-642-02008-7_13 [DOI] [PubMed] [Google Scholar]
- 12.Swofford DL. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0 b10. Sinauer Associates, Sunderland, 2002. [Google Scholar]
- 13.Pagani I, Liolios K, Jansson J, Chen IM, Smirnova T, Nosrat B, Markowitz VM, Kyrpides NC. The Genomes OnLine Database (GOLD) v.4: status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res 2012; 40:D571-D579 10.1093/nar/gkr1100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Qin QL, Zhang XY, Wang XM, Liu GM, Chen XL, Xie BB, Dang HY, Zhou BC, Yu J, Zhang YZ. The complete genome of Zunongwangia profunda SM-A87 reveals its adaptation to the deep-sea environment and ecological role in sedimentary organic degradation. BMC Genomics 2010; 11:247 10.1186/1471-2164-11-247 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pati A, Abt B, Teshima H, Nolan M, Lapidus A, Lucas S, Hammon N, Deshpande S, Cheng JF, Tapia R, et al. Complete genome sequence of Cellulophage lytica type strain (LIM21T). Stand Genomic Sci 2011; 4:221-232 10.4056/sigs.1774329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mavrommatis K, Gronow S, Saunders E, Land M, Lapidus A, Copeland A, Glavina Del Rio T, Nolan M, Lucas S, Chen F, et al. Complete genome sequence of Capnocytophaga ochracea type strain (VPI 2845T). Stand Genomic Sci 2009; 1:101-109 10.4056/sigs.15195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, Tatusova T, Thomson N, Allen MJ, Angiuoli SV, et al. The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol 2008; 26:541-547 10.1038/nbt1360 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Garrity G. NamesforLife. BrowserTool takes expertise out of the database and puts it right in the browser. Microbiol Today 2010; 37:9 [Google Scholar]
- 19.Woese CR, Kandler O, Wheelis ML. Towards a natural system of organisms. Proposal for the domains Archaea and Bacteria. Proc Natl Acad Sci USA 1990; 87:4576-4579 10.1073/pnas.87.12.4576 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Krieg NR, Ludwig W, Euzéby J, Whitman WB. Phylum XIV. Bacteroidetes phyl. nov. In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, Ludwig W, Whitman WB (eds), Bergey's Manual of Systematic Bacteriology, Second Edition, Volume 4, Springer, New York, 2011, p. 25. [Google Scholar]
- 21.Editor L. Validation List No. 143. [PubMed]. Int J Syst Evol Microbiol 2012; 62:1-4 [Google Scholar]
- 22.Bernardet JF. Class II. Flavobacteriia class. nov. In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, Ludwig W, Whitman WB (eds), Bergey's Manual of Systematic Bacteriology, Second Edition, Volume 4, Springer, New York, 2011, p. 105. [Google Scholar]
- 23.Ludwig W, Euzeby J, Whitman WG. Draft taxonomic outline of the Bacteroidetes, Planctomycetes, Chlamydiae, Spirochaetes, Fibrobacteres, Fusobacteria, Acidobacteria, Verrucomicrobia, Dictyoglomi, and Gemmatimonadetes
- 24.Judicial Commission of the International Committee on Systematics of Prokaryotes The nomenclatural types of the orders Acholeplasmatales, Halanaerobiales, Halobacteriales, Methanobacteriales, Methanococcales, Methanomicrobiales, Planctomycetales, Prochlorales, Sulfolobales, Thermococcales, Thermoproteales and Verrucomicrobiales are the genera Acholeplasma, Halanaerobium, Halobacterium, Methanobacterium, Methanococcus, Methanomicrobium, Planctomyces, Prochloron, Sulfolobus, Thermococcus, Thermoproteus and Verrucomicrobium, respectively. Opinion 79. Int J Syst Evol Microbiol 2005; 55:517-518 10.1099/ijs.0.63548-0 [DOI] [PubMed] [Google Scholar]
- 25.Bernardet JF. Order I. Flavobacteriales ord. nov. In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, Ludwig W, Whitman WB (eds), Bergey's Manual of Systematic Bacteriology, Second Edition, Volume 4, Springer, New York, 2011, p. 105. [Google Scholar]
- 26.Reichenbach H. Order 1. Cytophagales Leadbetter 1974, 99AL. In: Holt JG (ed), Bergey's Manual of Systematic Bacteriology, First Edition, Volume 3, The Williams and Wilkins Co., Baltimore, 1989, p. 2011-2013. [Google Scholar]
- 27.Bernardet JF, Segers P, Vancanneyt M, Berthe F, Kersters K, Vandamme P. Cutting a Gordian knot: emended classification and description of the genus Flavobacterium, emended description of the family Flavobacteriaceae, and proposal of Flavobacterium hydatis nom. nov. (Basonym, Cytophaga aquatilis Strohl and Tait 1978). Int J Syst Bacteriol 1996; 46:128-148 10.1099/00207713-46-1-128 [DOI] [Google Scholar]
- 28.Bernardet JF, Nakagawa Y, Holmes B. Proposed minimal standards for describing new taxa of the family Flavobacteriaceae, and emended description of the family. [PubMed]. Int J Syst Evol Microbiol 2002; 52:1049-1070 10.1099/ijs.0.02136-0 [DOI] [PubMed] [Google Scholar]
- 29.Validation of the publication of new names and new combinations previously effectively published outside the IJSB. List No. 41. Int J Syst Bacteriol 1992; 42:327-328 10.1099/00207713-42-2-327 [DOI] [PubMed] [Google Scholar]
- 30.BAuA. 2010, Classification of Bacteria and Archaea in risk groups. http://www.baua.de TRBA 466, p. 93.
- 31.Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 2000; 25:25-29 10.1038/75556 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lee OO, Lau SC, Tsoi MM, Li X, Plakhotnikova I, Dobretsov S, Wu MC, Wong PK, Qian PY. Gillisia myxillae sp. nov., a novel member of the family Flavobacteriaceae, isolated from the marine sponge Myxilla incrustans. Int J Syst Evol Microbiol 2006; 56:1795-1799 10.1099/ijs.0.64345-0 [DOI] [PubMed] [Google Scholar]
- 33.Bowman JP, Nichols DS. Novel members of the family Flavobacteriaceae from Antarctic maritime habitats including Subsaximicrobium wynnwilliamsii gen. nov., sp. nov., Subsaximicrobium saxinquilinus sp. nov., Subsaxibacter broadyi gen. nov., sp. nov., Lacinutrix copepodicola gen. nov., sp. nov., and novel species of the genera Bizionia, Gelidibacter and Gillisia. Int J Syst Evol Microbiol 2005; 55:1471-1486 10.1099/ijs.0.63527-0 [DOI] [PubMed] [Google Scholar]
- 34.Nedashkovskaya OI, Kim SB, Lee KH, Mikhailov VV, Bae KS. Gillisia mitskevichiae sp. nov., a novel bacterium of the family Flavobacteriaceae, isolated from sea water. Int J Syst Evol Microbiol 2005; 55:321-323 10.1099/ijs.0.63256-0 [DOI] [PubMed] [Google Scholar]
- 35.Klenk HP, Göker M. En route to a genome-based classification of Archaea and Bacteria? Syst Appl Microbiol 2010; 33:175-182 10.1016/j.syapm.2010.03.003 [DOI] [PubMed] [Google Scholar]
- 36.Wu D, Hugenholtz P, Mavromatis K, Pukall R, Dalin E, Ivanova NN, Kunin V, Goodwin L, Wu M, Tindall BJ, et al. A phylogeny-driven Genomic Encyclopaedia of Bacteria and Archaea. Nature 2009; 462:1056-1060 10.1038/nature08656 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.List of growth media used at DSMZ: http://www.dsmz.de/catalogues/catalogue-microorganisms/culture-technology/list-of-media-for-microorganisms.html
- 38.Gemeinholzer B, Dröge G, Zetzsche H, Haszprunar G, Klenk HP, Güntsch A, Berendsohn WG, Wägele JW. The DNA Bank Network: the start from a German initiative. Biopreserv Biobank 2011; 9:51-55 10.1089/bio.2010.0029 [DOI] [PubMed] [Google Scholar]
- 39.The DOE Joint Genome Institute www.jgi.doe.gov
- 40.Phrap and Phred for Windows. MacOS, Linux, and Unix. www.phrap.com
- 41.Zerbino DR, Birney E. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res 2008; 18:821-829 10.1101/gr.074492.107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Han C, Chain P. Finishing repeat regions automatically with Dupfinisher. In: Proceedings of the 2006 international conference on bioinformatics & computational biology. Arabnia HR, Valafar H (eds), CSREA Press. June 26-29, 2006: 141-146. [Google Scholar]
- 43.Lapidus A, LaButti K, Foster B, Lowry S, Trong S, Goltsman E. POLISHER: An effective tool for using ultra short reads in microbial genome assembly and finishing. AGBT, Marco Island, FL, 2008. [Google Scholar]
- 44.Hyatt D, Chen GL, Locascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal Prokaryotic Dynamic Programming Genefinding Algorithm. BMC Bioinformatics 2010; 11:119 10.1186/1471-2105-11-119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pati A, Ivanova N, Mikhailova N, Ovchinikova G, Hooper SD, Lykidis A, Kyrpides NC. GenePRIMP: A Gene Prediction Improvement Pipeline for microbial genomes. Nat Methods 2010; 7:455-457 10.1038/nmeth.1457 [DOI] [PubMed] [Google Scholar]
- 46.Markowitz VM, Ivanova NN, Chen IMA, Chu K, Kyrpides NC. IMG ER: a system for microbial genome annotation expert review and curation. Bioinformatics 2009; 25:2271-2278 10.1093/bioinformatics/btp393 [DOI] [PubMed] [Google Scholar]
- 47.Spudich JL, Yang CS, Jung KH, Spudich EN. Retinylidene proteins: structures and functions from archaea to humans. Annu Rev Cell Dev Biol 2000; 16:365-392 10.1146/annurev.cellbio.16.1.365 [DOI] [PubMed] [Google Scholar]
- 48.Fuhrman JA, Schwalbach MS, Stingl U. Proteorhodopsins: an array of physiological roles? Nat Rev Microbiol 2008; 6:488-494 [DOI] [PubMed] [Google Scholar]
- 49.Martinez A, Bradley AS, Waldbauer JR, Summons RE, DeLong EF. Proteorhodopsin photosystem gene expression enables photophosphorylation in a heterologous host. Proc Natl Acad Sci USA 2007; 104:5590-5595 10.1073/pnas.0611470104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Walter JM, Greenfield D, Bustamante C. Liphardt. Light-powering Escherichia coli with proteorhodopsin. Proc Natl Acad Sci USA 2007; 104:2408-2412 10.1073/pnas.0611035104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Balashov SP, Imasheva ES, Boichenko VA, Anton J, Wang JM, Lanyi JK. Xanthorhodopsin. A proton pump with a light-harvesting carotenoid antenna. Science 2005; 309:2061-2064 10.1126/science.1118046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Balashov SP, Lanyi JK. Xanthorhodopsin: Proton pump with a carotenoid antenna. Cell Mol Life Sci 2007; 64:2323-2328 10.1007/s00018-007-7167-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Antón J, Oren A, Benlloch S, Rodriguez-Valera F, Amann R, Rossello-Mora R. Salinibacter ruber gen. nov., sp. nov., a novel, extremely halophilic member of the Bacteria from saltern crystallizer ponds. Int J Syst Evol Microbiol 2002; 52:485-491 [DOI] [PubMed] [Google Scholar]
- 54.Mongodin EF, Nelson KE, Daugherty S, Deboy RT, Wister J, Khouri H, Weidman J, Walsh DA, Papke RT, Sanchez PG, et al. The genome of Salinibacter ruber: convergence and gene exchange among hyperhalophilic bacteria and archaea. Proc Natl Acad Sci USA 2005; 102:18147-18152 10.1073/pnas.0509073102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Balashov SP, Imasheva ES, Wang JM, Lanyi JK. Excitation energy-transfer and the relative orientation of retinal and carotenoid in xanthorhodopsin. Biophys J 2008; 95:2402-2414 10.1529/biophysj.108.132175 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Béjà O, Aravind L, Koonin EV, Suzuki MT, Hadd A, Nguyen LP, Jovanovich SB, Gates CM, Feldman RA, Spudich JL, et al. Bacterial rhodopsin: evidence for a new type of phototrophy in the sea. Science 2000; 289:1902-1906 10.1126/science.289.5486.1902 [DOI] [PubMed] [Google Scholar]
- 57.Béjà O, Spudich EN, Spudich JL, Leclerc M, DeLong EF. Proteorhodopsin phototrophy in the ocean. Nature 2001; 411:786-789 10.1038/35081051 [DOI] [PubMed] [Google Scholar]
- 58.Oesterhelt D, Stoeckenius W. Rhodopsin-like protein from the purple membrane of Halobacterium halobium. Nat New Biol 1971; 233:149-152 [DOI] [PubMed] [Google Scholar]
- 59.Le SQ, Gascuel O. An improved general amino acid replacement matrix. Mol Biol Evol 2008; 25:1307-1320 10.1093/molbev/msn067 [DOI] [PubMed] [Google Scholar]
- 60.Dioumaev AK, Brown LS, Shih J, Spudich EN, Spudich JL, Lanyi JK. Proton transfers in the photochemical reaction cycle of proteorhodopsin. Biochemistry 2002; 41:5348-5358 10.1021/bi025563x [DOI] [PubMed] [Google Scholar]
- 61.Wang WW, Sineshchekov A, Spudich EN, Spudich JL. Spectroscopic and photochemical characterization of a deep ocean proteorhodopsin. J Biol Chem 2003; 278:33985-33991 10.1074/jbc.M305716200 [DOI] [PubMed] [Google Scholar]
- 62.Man D, Wang W, Sabehi G, Aravind L, Post AF, Massana R, Spudich EN, Spudich JL, Beja O. Diversification and spectral tuning in marine proteorhodopsins. EMBO J 2003; 22:1725-1731 10.1093/emboj/cdg183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Riedel T, Tomasch J, Buchholz I, Jacobs J, Kollenberg M, Gerdts G, Wichels A, Brinkhoff T, Cypionka H, Wagner-Döbler I. Constitutive expression of the proteorhodopsin gene by a flavobacterium strain representative of the proteorhodopsin-producing microbial community in the North Sea. Appl Environ Microbiol 2010; 76:3187-3197 10.1128/AEM.02971-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Klippel B, Lochner A, Bruce DC, Walston DK, Detter C, Goodwin LA, Han J, Han S, Hauser L, Land ML, et al. Complete Genome Sequences of Krokinobacter sp. Strain 4H-3-7-5 and Lacinutrix sp. Strain 5H-3-7-4, Polysaccharide-Degrading Members of the Family Flavobacteriaceae. J Bacteriol 2011; 193:4545-4546 10.1128/JB.05518-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Yoon JH, Kang SJ, Lee CH, Oh TK. Dokdonia donghaensis gen. nov., sp. nov., isolated from sea water. Int J Syst Evol Microbiol 2005; 55:2323-2328 10.1099/ijs.0.63817-0 [DOI] [PubMed] [Google Scholar]
- 66.Yoon JH, Kang SJ, Park S, Oh TK. Reclassification of the three Krokinobacter species into the genus Dokdonia as Dokdonia genika comb. nov., Dokdonia diaphora comb. nov. and Dokdonia eikasta comb. nov. and emended description of the genus Dokdonia Yoon et al. 2005. Int J Syst Evol Microbiol 2011; [Epup ahead of print].
- 67.Nam YD, Lee HW, Lee M, Yim KJ, Kim KN, Roh SW, Kim D. Draft Genome Sequence of Gillisia sp strain CBA3202, a novel member of the genus Gillisia, which belongs to the family Flavobacteriaceae. J Bacteriol 2012; 194:3739 10.1128/JB.00631-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Auch AF, Klenk HP. Göker, M. Standard operating procedure for calculating genome-to-genome distances based on high-scoring segment pairs. Stand Genomic Sci 2010; 2:142-148 10.4056/sigs.541628 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.DeLong EF, Franks DG, Alldredge AL. Phylogenetic diversity of aggregate-attached versus free-living marine bacterial assemblages. Limnol Oceanogr 1993; 38:924-934 10.4319/lo.1993.38.5.0924 [DOI] [Google Scholar]
- 70.Pinhassi J, Sala MM, Havskum H, Peters F, Guadayol O, Malits A, Marrasé C. Changes in bacterioplancton composition under different phytoplankton regimes. Appl Environ Microbiol 2004; 70:6753-6766 10.1128/AEM.70.11.6753-6766.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Riemann L, Steward GF, Azam F. Dynamics of bacterial community composition and activity during a mesocosm diatom bloom. Appl Environ Microbiol 2000; 66:578-587 10.1128/AEM.66.2.578-587.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.McBride MJ. Bacterial gliding motility: multiple mechanisms for cell movement over surfaces. Annu Rev Microbiol 2001; 55:49-75 10.1146/annurev.micro.55.1.49 [DOI] [PubMed] [Google Scholar]
- 73.McBride MJ. Cytophaga- Flavobacterium gliding motility. J Mol Microbiol Biotechnol 2004; 7:63-71 10.1159/000077870 [DOI] [PubMed] [Google Scholar]