

Abstract
Trachoma, caused by Chlamydia trachomatis (Ct), is the leading infectious blinding disease worldwide. Chronic conjunctival inflammation develops in childhood and leads to eyelid scarring and blindness in adulthood. The immune response to Ct provides only partial protection against re-infection, which can be frequent. Moreover, the immune response is central to the development of scarring pathology, leading to loss of vision. Here we review the current literature on both protective and pathological immune responses in trachoma. The resolution of Ct infection in animal models is IFNγ-dependent, involving Th1 cells, but whether this is the case in human ocular infection still needs to be confirmed. An increasing number of studies indicate that innate immune responses arising from the epithelium and other innate immune cells, along with changes in matrix metalloproteinase activity, are important in the development of tissue damage and scarring. Current trachoma control measures, which are centred on repeated mass antibiotic treatment of populations, are logistically challenging and have the potential to drive antimicrobial resistance. A trachoma vaccine would offer significant advantages. However, limited understanding of the mechanisms of both protective immunity and immunopathology to Ct remain barriers to vaccine development.
Introduction
Trachoma results from infection of the conjunctiva with Chlamydia trachomatis (Ct) and is the commonest infectious cause of blindness worldwide. The World Health Organization (WHO) estimates that at least 1.3 million people are blind from trachoma and 40 million have active disease; it is also part of the Neglected Tropical Diseases Programme [1], [2]. It is caused by Ct serovars A to C and is generally found in poor, rural areas in less developed countries.
Genital tract Ct infection, the commonest bacterial sexually transmitted disease worldwide, infecting 90 million people each year, is caused by serovars D to K [3]. The ocular and genital strains are strictly differentiated with ocular strains lacking tryptophan synthase function [4]. Both ocular and genital infections produce inflammatory reactions, which are often asymptomatic, leading to scarring complications with significant morbidity in a subset of those infected. The WHO recommends the SAFE Strategy for trachoma control: Surgery for trichiasis, mass Antibiotic distribution to treat infection, improved Facial hygiene, and Environmental improvements to interrupt Ct transmission [5]–[7]. While implementation of this strategy appears to be effective in reducing active ocular infection with Ct, successfully carrying it out on a programmatic level can be challenging in the areas where trachoma is found, and the long-term effect on the scarring stages of trachoma is unknown.
The immune response to Ct provides incomplete protection and is known to be important in the development of tissue damage and sequelae including blinding complications. However, protective immune responses and disease pathogenesis in trachoma remain poorly understood. The “immunological paradigm” suggested that disease pathology is a result of cell-mediated immune (CMI) responses against specific chlamydial antigens. The “cellular paradigm” states that infected epithelial cell responses drive pathology through the release of various mediators, and this is supported by recent studies in humans and animal models, which have highlighted the importance of an innate immune response in active and scarring trachoma.
This article reviews the literature on immunity and immunopathogenesis in trachoma. Human studies of the pathology, immune response, and immunopathogenesis in trachoma are comprehensively summarised. Whilst focusing on these human studies of ocular Ct infection, we also draw on relevant studies of genital tract infection and animal models. Ocular infection of nonhuman primates leads to a self-limiting follicular conjunctivitis after a single innoculum, and chronic inflammation, conjunctival scarring, and trichiasis with repeated inoculations [8]–[12]. Such a response is similar to that seen in humans, and this model has useful parallels for human disease [13]. Other animal models, often involving mice [14], [15] and guinea pigs [16], [17], are limited by major differences in immune responses with humans [18]. Additionally, they have usually relied on the use of other chlamydial species such as Chlamydia caviae, psittaci, or muridarum, which may be evolutionarily separated from Ct and are adapted to their respective hosts.
Methods
Ethical approval was not required for this review article. References were identified through searches of PubMed for articles published at any date, by use of the terms “trachoma” and “immunology,” “pathogenesis,” “pathology,” “scarring,” or “histology.” Articles resulting from these searches and relevant articles cited in those articles were reviewed. Articles published in English were included. Relevant human studies, which are the focus of this review, are summarised in Tables S1, S2, S3, S4, S5, S6 in Supporting Information S1.
Clinical Features, Natural History, and Infection
Infection with Ct causes chronic inflammation of the conjunctiva (conjunctivitis) or “active trachoma,” which is characterised by follicles (subepithelial collections of lymphoid cells appearing as small, yellow-white elevations) and papillae (engorgement of small vessels with inflammatory conjunctival thickening). Infection may be detected in the absence of clinical disease, and conversely, active disease may be present without detectable current Ct infection [19]. While this may appear paradoxical, there are a number of reasons to explain these findings, including how sensitive the diagnostic test is compared to the clinical examination, and also the kinetics of the disease [19], [20]. These include a latent phase with infection before the onset of clinical signs and a recovery phase where infection has been cleared but clinical signs persist. Children experiencing repeated episodes of infection are at increased risk of subsequent conjunctival scarring, which causes in-turning of the eyelids (entropion) so that the eyelashes scratch the cornea (trichiasis). Eventually sight is lost as irreversible corneal opacification develops from a combination of insults, which compromise the ocular surface (trichiasis, dryness, secondary bacterial, or fungal infections). Clinical features are shown in Figure 1. The scarring complications of trachoma usually develop slowly over many years. Several longitudinal studies support this pathway to blindness, although there is considerable variation in reported disease progression rates, possibly reflecting differences in both the natural history in different populations and in study methodology (Table 1).
Figure 1. Clinical features and grades of trachoma.
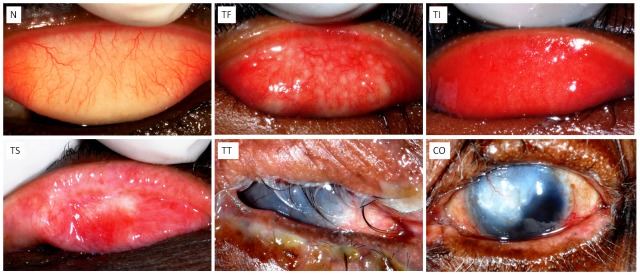
N, normal; TF, Trachomatous inflammation–follicular; TI, Trachomatous inflammation–intense; TS, trachomatous scarring; TT, trachomatous trichiasis; CO, corneal opacity.
Table 1. Summary of the incidence and progression rates of trachomatous scarring.
| Progression Factor | Sample Size | Follow-Up Interval | Rate | Setting | Prospective Design? | Associated Factors |
| Incident conjunctival scarring [28], [151] | 367 | 5 y | 20.4% | Tanzania | Yes | In children <10 y: active disease/persistent infection; female gender; age |
| Incident conjunctival scarring [24] | 236 (age <7 y) | 7 y | 29.2% versus 9.6% | Tanzania | No | Higher rate was in children with severe-constant active disease; female gender; age |
| Worsening of conjunctival scarring [151] | 85 | 5 y | 47.1% | Tanzania | Yes | Not specified |
| Worsening of conjunctival scarring [25] | 213a | 14 y | 68.5% | Tunisia | No | Active disease; household density |
| From conjunctival scarring to trichiasis [26] | 523 (all women) | 7 y | 9.2% | Tanzania | No | Active disease; chlamydial infection; increasing age |
| From conjunctival scarring to trichiasis [152] | 297 | 12 y | 6.4% | Gambia | No | Mandinka ethnicity |
| From conjunctival scarring to trichiasis [27] | 4,898 | 5 y | 3.2%–15.1% | Tanzania | Nob | Increasing age |
| From minor to major trichiasis [153] | 55 | 1 y | 33% | Gambia | No | None mentioned |
| From minor to major trichiasis [27] | 75 | 4 y | 37% | Gambia | No | Conjunctival inflammation |
| From unilateral to bilateral trichiasis [153] | 46 | 1 y | 46% | Gambia | No | Baseline pannus; hot ash as an aid to epilation |
| From conjunctival scarring +/− trichiasis to corneal scarring [152] | 302 | 12 y | 6.0% | Gambia | No | Baseline trichiasis |
| From trichiasis to corneal scarring [27] | 211 | 4 y | 7.6% | Gambia | No | Increasing trichiasis severity; conjunctival inflammation |
| From trichiasis to corneal opacity [154] | 4,898 | 10 y | 27.2%–53.5% | Tanzania | Nob | Increasing age |
| Worsening of corneal scarring [153] | 96 | 1 y | 34% | Gambia | No | Conjunctival inflammation; bacterial growth |
Including 82 people with no scarring at baseline.
Estimated incidence rates based on age-specific prevalence of scarring, trichiasis, and corneal opacity among women.
A key question is why only a minority of people living in trachoma endemic regions develop severe scarring complications. It is likely that the answer to this is a complex interaction between the individual's lifetime burden of infection and their local immune response. Risk factors for trachoma have been extensively reviewed elsewhere [19], [21]–[23] and include poor facial hygiene with unwashed ocular/nasal secretions, crowded living conditions, flies (which act as a vector for the disease), and migration between communities. Severe, persistent, or recurrent conjunctival inflammation is strongly associated with the risk of scarring complications [24]–[28]. Females are at increased risk of conjunctival scarring and trichiasis, possibly from greater lifetime Ct exposure through close contact with children [21], [29]. In regions where trachoma is highly endemic, scarring sequelae are more common and occur at a younger age [30], [31]. There is good evidence that the immune response to Ct is partly determined by host genetic variations, with several studies from different populations having identified genetic polymorphisms related to immune system function that are associated with trachomatous scarring. These include polymorphisms in interferon-γ (IFNγ), tumour necrosis factor-α, and matrix metalloproteinase (MMP) 9 (Table S1) and indicate the importance of host immunogenetic variations in modifying the risk of developing blinding disease.
Trachoma was initially thought to result from a single episode of infection [32]. However, the importance of repeated re-infection in the pathogenesis of scarring disease is now supported by various lines of investigation: animal models require repeated inoculation to induce disease [9], [10], absence of scarring following a single inoculation in human volunteer studies [32], and cohort studies from trachoma-endemic areas demonstrating repeated infection [20], [28], [33]. However, in vitro studies have shown that persistent, nonreplicating forms of Ct develop in response to various stress stimuli (penicillin, IFNγ, and iron or nutrient depletion) [34]. The relevance of these findings to human infection is not known. There is very limited in vivo evidence for persistent ocular infection [35]. An important observation about Ct infection and ocular disease is that infection is infrequently detected in older adults, who are the group in which progressive scarring and blinding complications usually develop [27], [36]–[38]. This is partly explained by the shorter duration of infection with increasing age [39]. However, other factors may also be important in scarred conjunctiva, such as nonchlamydial bacterial infection (discussed below).
Histopathology
Histological examination of conjunctival biopsy samples from subjects with trachoma has given some insights into the disease process (Table S2). However, interpretation of studies conducted to date is limited by the relatively small sample sizes involved and few or no control participants. Comparison with nontrachomatous subjects is essential to establish background conjunctival morphology and cellular compositions in these trachoma-endemic areas, as this may differ markedly from those populations where trachoma is not endemic.
Active Disease
Histological studies of conjunctival biopsies from children with active trachoma found mild to moderate epithelial hyperplasia with a mixed inflammatory infiltrate consisting of many macrophages with some T cells and polymorphonuclear leucocytes (Table S2) [40]–[42]. Dendritic cells are seen in the deeper epithelium and underlying stroma. Plasma cells form a band directly beneath the epithelium and around accessory lacrimal glands. Lymphoid follicles, the clinical and pathological hallmark of active trachoma, are found in the stroma (Figure 2). In children these consist mainly of B cells with some macrophages and T cells and are surrounded with a lymphocytic mantle. Around the follicle, a diffuse infiltrate consisting of a mixed population of leucocytes (T cells, neutrophils, macrophages, mast cells, and eosinophils) is seen. Schematic diagrams of normal conjunctiva and active disease are shown in Figure 3.
Figure 2. Histological section of the conjunctiva from a child with active trachoma.

A subepithelial follicle is seen. Kindly provided by Professor A. El-Asrar, King Saud University.
Figure 3. Schemas of normal conjunctiva, active trachomatous disease, and trachomatous scarring.

Scarring Disease
Adults with conjunctival scarring have an epithelium that may show either squamous metaplasia or atrophy with focally denuded areas replaced by fibrinous exudate and cellular debris (Figure 4) [42], [43]. A chronic inflammatory cell infiltrate is found, most marked in the substantia propria with lymphocytes predominating [40], [44]. CD4+ and CD8+ T cells are present, and these generally outnumber B cells. Follicles may be seen histologically in very inflamed subjects, even when absent clinically; these contain monocytes, macrophages, and plasma cells but lack the germinal centres or centrally located B cells seen in follicles of children. Underneath the epithelium, the stroma is replaced by abnormal, compact, thick, and mostly avascular scar tissue [43]. This subepithelial fibrous membrane has fibres parallel to the surface epithelium and is firmly adherent to the tarsal plate (Figure 3).
Figure 4. Histological sections of healthy conjunctiva and trachomatous scarring.
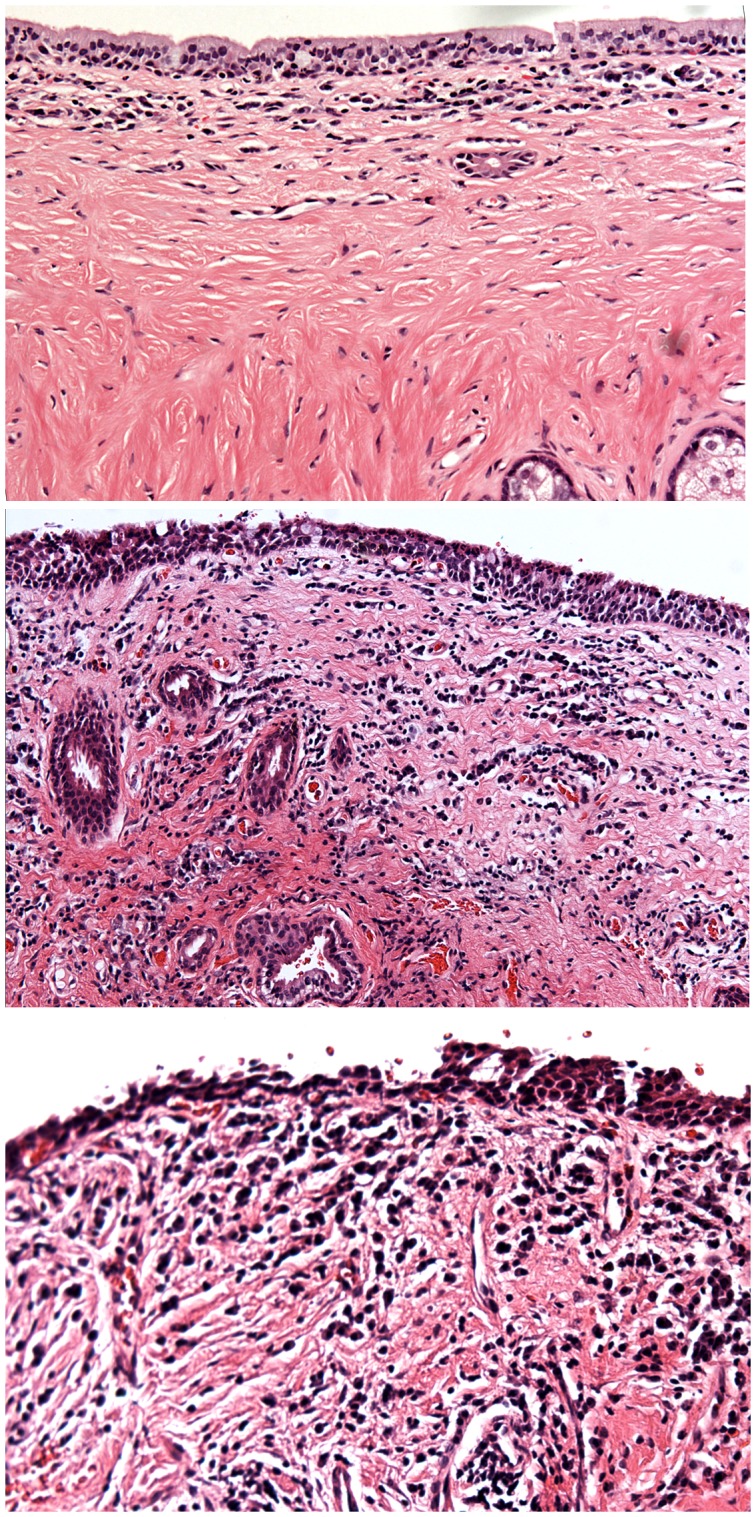
(Top) Healthy. (Middle and bottom) Scarred, note disruption of the epithelial and connective tissue morphology, and an increased inflammatory cells.
Resolution of Ct Infection
A number of immune-mediated responses have been demonstrated to Ct infection that are thought to be important in clearing infection. The area requires further clarification, however, with many of these responses yet to be unequivocally associated with resolution of infection in human Ct infection.
Epithelial Cells Mount a Pro-Inflammatory and Chemotactic Response
Several in vitro studies of Ct-infected epithelial cell lines have found that this infection provokes pro-inflammatory responses with epithelial cells producing inflammatory and chemotactic cytokines such as interleukin-6 (IL-6), IL-8, growth-regulated oncogene α (GROα), and granulocyte macrophage colony-stimulating factor (GM-CSF) [45]–[47]. The secretion of these factors was prolonged and required synthesis of chlamydial proteins. In contrast, other bacterial species provoked a rapid but transient cytokine induction [45].
Human studies are consistent with the epithelium-producing pro-inflammatory factors constituting an initial innate response to Ct. Immunohistochemistry of conjunctival epithelium from children with active trachoma has shown IL-1 expression [48]. Gene expression studies on conjunctival surface swab samples find enriched expression of multiple pro-inflammatory/chemotactic factors (Table S3). In active trachoma, the epithelium shows increased Major Histocompatibility Complex (MHC) class I expression (found at a lower level in normal conjunctiva) and induction of MHC class II (not normally found at this site) [41], [49]. There is also evidence for natural killer (NK) cell and neutrophil recruitment and activation in active disease/infection as part of an acute inflammatory response [16], [50], [51].
Importance of a CMI Response and IFNγ Production
The results of animal studies of genital chlamydial infection suggest that resolution of infection is dependent on a Type-1 CD4+ T-helper lymphocyte (Th1) response-mediated primarily through IFNγ. Congenitally athymic (nude) mice are unable to clear C. muridarum genital tract infection [52]. However, adoptive transfer to nude mice of CD4+ or CD8+ T cells derived from normal mice previously infected with C. muridarum resolved the infection [53]. The CD4+-enriched cell line was much more efficient at clearing infection. However, recent murine studies using Ct instead of C. muridarum as the infectious agent did not find that the absence of CD4+ cells altered the course of infection and limitations of animal models of Ct infection need to be acknowledged [54]. Although CD8+ T cells can help resolve infection, they seem less important in animal models, as they were not an absolute requirement for clearing infection [55].
IFNγ is produced by several cell types including Th1 lymphocytes, CD8+ cytotoxic T lymphocytes (CTLs), and NK cells. The production of IFNγ characterises the Th1 response and has several antichlamydial actions, which probably play a central role in clearing infection. IFNγ induces indoleamine-2,3-dioxygenase (IDO) expression, which lowers the intracellular pool of the essential amino acid tryptophan, which is necessary for chlamydial metabolism [56], [57]. IFNγ can also up-regulate inducible nitric oxide synthase (iNOS), which may help control Ct and protect against chronic sequelae in murine models [58]. In vitro, chlamydial growth can also be limited by IFNγ-mediated intracellular iron depletion [59].
Several human studies point to the importance of the CMI response to ocular Ct infection, although direct longitudinal evidence that resolution is IFNγ-dependent is limited. Biopsies from children with active trachoma show heavy cellular infiltrates with both CD4+ and CD8+ lymphocytes (Table S2) [41]. Children who resolved active disease (associated with infection) had stronger peripheral blood lymphoproliferative responses to Ct antigens compared to children with persistent disease (Table S4) [60]. Gene expression studies have shown increased expression of IFNγ, IL12p40, Perforin, IDO, IL-4, IL-10, and forkhead box p3 (FOXP3) in infection and/or disease, with levels generally being highest if both were present (Table S3) [33], [50], [61], [62]. Increased expression of IFNγ, IL-10, and FOXP3 has been associated with longer episodes of infection [63]. A recent study using Ct elementary bodies (EBs) to stimulate PBMC from individuals exposed to ocular infection showed that cells with a classical NK cell phenotype (CD3-CD56+) were an additional major source of IFNγ [51].
Protective Immunity to Ct
Human volunteer studies of ocular Ct infection have provided some evidence that a partial, protective immune response develops [64]–[66]. Conjunctival inoculation with Ct led to a characteristic follicular/papillary conjunctivitis in nearly all volunteers after 2–14 d. However, when previously infected individuals were rechallenged with the same serovar, there was an attenuated clinical response with reduced re-isolation rates. This apparent immunity to re-infection was serovar-specific: if a different serovar was used, then disease and infection levels were similar to the primary infection. Presumed complete immunity after primary infection, in which no clinical signs developed and the organism was never detected, despite multiple rechallenge inoculations, was rare. Despite the concerns over the induction of presumed delayed-type hypersensitivity (DTH), the trachoma vaccine trials in the 1960s did demonstrate short-term, serovar-specific immunity could be induced [67], [68]. Trachoma models in nonhuman primates also showed that, after recovery from a primary infection, on secondary challenge there was less severe disease that resolved more rapidly [9]. With regular weekly re-inoculations, it also became progressively more difficult to detect the organism, suggesting the development of protective immunity. The human studies discussed above were conducted over 40 years ago and need to be interpreted in that context. Ethical considerations, however, limit contemporary studies to being of an observational design.
A longitudinal study in which a cohort from a trachoma endemic population was regularly assessed for disease and infection found the duration of both shortened considerably with increasing age [39], [69]. This is thought to be the result of an increasingly effective protective immune response developing with repeated exposure to Ct, although other factors could also have been important, such as behavioural changes. Another study has shown that in 4- to 15-y-olds, there was evidence of age-related modulation of cytokine expression, with older individuals tending to have greater expression of IFNγ, IDO, and IL-10 [62]. The clinical manifestations also appeared to be modulated: adults experienced short bursts of intense disease and children more prolonged episodes of follicular disease [39].
Acquired T-cell–dependent CMI responses, as outlined above, probably contribute to protection against or more rapid resolution of re-infection, although direct evidence for this from human trachoma studies is limited. Fading immunity in animal models is correlated with a reduction in CD4+ T cells, which leave the genital tract following resolution of infection [70]. A study of the incidence of genital tract infection amongst sex workers in Nairobi, Kenya, found that IFNγ production by PBMC stimulated with cHsp60 was associated with reduced risk of re-infection compared to women without such responses [71].
The importance of humoral immunity to Ct is unclear. Several human studies have detected antichlamydial antibodies in tear fluid and serum (Table S5). However, in a longitudinal study, antichlamydial IgG in tears from clinically normal individuals was associated with an increased incidence of subsequently developing clinically active trachoma [72]. In contrast, there was an opposite (nonsignificant) trend with antichlamydial IgA. Individuals with conjunctival scarring have been found to have significantly higher plasma titres of antichlamydial IgG and lower titres of IgA compared with normal controls [73].
However, a consistent picture on the role of antibody in protection from trachoma has not emerged from these studies. What is clear is that the overall level of the antibody response can be a useful proxy of the lifetime exposure to ocular infection. Within the overall antibody response, a profile of specific antibodies directed against a limited number of individual antigens are associated with the presence or absence of clinical signs of disease [74]. However, how such antibodies mediate protection and pathology requires elucidation.
Various animal models of genital tract infection have yielded a mixed pattern of results on the relative importance of the humoral response. Some have shown that B-cell–deficient mice can resolve a primary chlamydial infection and are only slightly delayed in clearing secondary infections, compared to control mice [75], [76]. However, other studies found that B-cell–deficient mice that also had their CD4+ cells depleted were completely unable to control secondary infection, whereas mice depleted of CD4+ cells alone only showed slight delay in clearing secondary infection [77], [78]. In addition, transfer of monoclonal antibodies to Ct major outer membrane protein (MOMP) and LPS into B-cell–deficient, CD4+ T-cell–depleted mice restored the ability of these mice to control secondary infection [79]. Antibodies to polymorphic membrane protein D (PmpD) have also been shown to be neutralising in vitro [80]. However, whether similar protective effects occur in human ocular infection is unknown.
Immunopathogenesis of Trachoma
In 1974 Silverstein proposed that tissue damage and scarring sequelae in trachoma are the result of a chronic, immunopathogenic response to an otherwise seemingly innocuous, superficial infection [81]. He observed that Ct infection itself is limited to a small minority of epithelial cells and is not particularly cytopathic, with cells accommodating large chlamydial inclusion bodies. The infection can be quite prolonged, suggesting that the immune response is relatively inefficient or that Ct is effective at avoiding immune responses. At the tissue level, the inflammatory infiltrate and subsequent scarring is in the conjunctival stroma, below the level of the epithelium, where the infection is located.
A key clinical observation pointing to the immunopathogenic nature of trachoma is that visible conjunctival inflammation is frequently found in the absence of detectable Ct [19], [20], [33], [61], [82]. In longitudinal studies, discrete episodes of active disease persist long after the associated initiating infectious episode has resolved [39]. In one trachoma vaccine trial, the incidence of trachoma was higher in the vaccinated group and another reported a possible increase in disease severity [68], [83].
Monkey models of Ct infection support the importance of immunopathology in trachoma [9]. Treatment of infected animals with corticosteroids markedly reduced the inflammation, and in monkey vaccine studies, rechallenge was associated with more severe inflammatory disease and scarring complications [84], [85].
Several theories for the pathogenesis and scarring caused by chlamydial infection have been proposed, none of which entirely explain the observed data. Stephens helpfully subdivided these into two broad paradigms: the “cellular paradigm” in which the response of the epithelial cell layer is central and sufficient to explain the pathology, and the “immunological paradigm” in which cell-mediated immunity causing either DTH or autoimmune mimicry explains disease pathology [86]. These processes are not mutually exclusive and may both contribute.
The “Immunological Paradigm” and Trachoma
The “immunological paradigm” suggests that tissue damage and fibrosis in chlamydia-related disease results from CMI responses against specific chlamydial antigens [46], [55], [81], [86]. It argues that specific T-cell responses, which are important in clearing Ct infection, also cause collateral tissue damage. There are two major lines of investigation that have explored this hypothesis, which are discussed below.
DTH and cHsp60
During early trachoma vaccine studies on nonhuman primates, it was observed that the disease phenotype could be more severe on rechallenge. This led the investigators to propose that a DTH reaction to chlamydial antigens was responsible for the disease [10]. Subsequently, in a guinea pig model of ocular infection, a Ct antigen, extracted using the detergent triton X-100, was able to provoke an inflammatory reaction in animals previously infected with Ct [87]. The active component was identified as the 57 kDa protein, cHsp60 [88], [89]. Similar experiments were conducted in cynomolgus monkeys, with comparable results [11]. There was some concern that the conjunctival inflammatory response observed in these monkeys might have been partly provoked by the triton X-100 detergent itself, as repeated installation of the buffer alone also led to significant inflammation. However, when experiments were repeated with recombinant cHsp60, inflammation was provoked in guinea pig eyes and in the subcutaneous abdominal salpingitis pocket model in monkeys [88]–[91]. Interpretation of the role of cHsp60 from these animal models is difficult, since a further study showed that initial immunisation with recombinant antigen in guinea pigs did not produce exacerbated disease after ocular infection with viable organisms but rather may have led to some protective effect [92].
Human studies on cHsp60 have delivered mixed results on its role in pathogenesis. Antibodies to cHsp60 are associated with trachomatous scarring, pelvic inflammatory disease, and tubal infertility [93]–[95]. It is unclear, however, whether these antibodies have a pathogenic role or are simply markers of previous infection. Other studies have suggested that immune responses to cHsp60 may even be protective: examination of PBMC proliferation responses to chlamydial antigens in trachoma endemic populations found that conjunctival scarring was associated with weaker responses to cHsp60, while the resolution of infection was associated with increased responses [60], [73]. Also, as noted above, IFNγ production by PBMC stimulated with cHsp60 is associated with protection against subsequent genital tract re-infection [71]. Overall, there is little evidence from human studies to suggest that scarring trachoma is the result of a DTH reaction to cHsp60.
Differential Activity of CD4+ T-Cell Subsets
A Th2-dominated response has been linked to the development of scarring complications in some infectious diseases such as schistosomiasis [96]. However, there is little evidence for a similar role in trachoma. Conjunctival gene expression profiling in children with active disease and Ct infection found an increase in both Th1 (IFNγ) and Th2 (IL-4) cytokine expression (Table S3) [33], [61]. Adults with scarring, compared to controls, have reduced lymphoproliferative responses and IFNγ production in response to stimulation with Ct EB and some antigens, but an increased number of IL-4–producing cells in response to cHsp60 [73], [97]. This seems to support the hypothesis that individuals with scarring may have weaker Th1 cell-mediated responses to Ct, leading to prolonged infection and inflammation possibly as a result of Th2 responses with pro-fibrotic effects. However, recent studies comparing the conjunctival transcriptome by microarray, and RT-PCR expression of selected targets, in subjects with scarring trachoma and matched controls found no evidence for Th2 responses and only indirect evidence of increased expression of genes associated with Th1 cells [98]–[100]. Furthermore, the expression of IL-13 was lower in individuals with established conjunctival scarring and inflammation, compared to healthy controls. Additionally, Th2 cytokine levels in tear fluid were not increased in scarred individuals (Table S6) [101].
The role other types of CD4+ T cells play in trachoma pathogenesis has received little attention. One study showed that the expression of FOXP3, used as an indicator of regulatory T cells, was increased in children who had clinical signs of trachoma but in whom infection had resolved [62]. However, whether this role is beneficial, by dampening the immune response and subsequent tissue damage once infection has been cleared, or detrimental, by impeding removal of infectious agent, is unknown. The expression of IL-17A, suggestive of Th17 cell activity, has been found to be significantly increased in active trachoma (discussed below) [50], [102], [103].
The “Cellular Paradigm” and Trachoma
In the “cellular paradigm,” tissue damage and scarring are thought to be driven by infected epithelial cells, which serve as early innate system responders [46], [86]. Pro-inflammatory chemokines, cytokines, and growth factors produced by Ct-infected epithelial cells initiate the recruitment of inflammatory immune cells (neutrophils, macrophages, and NK cells). Infected epithelial cells express multiple factors, which also promote the CMI response and are chemotactic for lymphocytes. The “cellular paradigm” does not differentiate between damage resulting from professional innate immune cells, such as neutrophils and monocytes, and adaptive immune cells [46]. However, it does propose that chronic inflammatory responses, tissue damage, and fibrosis in Ct-related disease is largely driven from the epithelium rather than by CMI. There is now increasing evidence that innate immune responses are a prominent feature of responses in trachoma and likely interact with the adaptive response.
Animal studies have shown an influx of neutrophils into genital tract tissue following Ct inoculation [104]. Neutrophils were not required to clear infection, however the intensity of the neutrophil infiltrate was related to subsequent fibrotic sequelae [105]. A guinea pig model of trachoma examining neutrophil depletion showed no effect on the burden of infection, however there was less clinical inflammation and fewer mucosal erosions histologically [16]. In this study, adaptive immune responses were also affected with reduced CD4+ and CD8+ cell recruitment and changes in the expression of various cytokines and chemokines such as decreased CCL5. This study only evaluated animals for up to 7 days, and once again caution needs to be exercised in extrapolating results of animal models to humans. However, it does provide an insight into the role of innate immunity in trachoma pathogenesis and how this might partly be mediated through its interaction with adaptive responses.
The production of pro-inflammatory cytokines in response to Ct infection is mediated in part through recognition of pathogen-associated molecular patterns (PAMPs) by Toll-like receptors (TLRs) and other pattern recognition receptors (PRRs) [106]. While TLR2 knockout (KO) mice were able to eradicate infection in a similar manner to control mice, they had reduced TNFα and CXCL2 [107]. Of particular note is the observation that TLR2-KO mice also had a marked reduction in late oviduct pathology. Human genetic studies on TLR polymorphisms have yet to identify significant associations with chlamydial diseases [108]–[110].
In children with active trachoma, IL-1 has been found in the surface epithelial cells of cases, but not controls [48]. This might promote recruitment of innate immune cells, including neutrophils and macrophages, which are indeed seen in large numbers in the subepithelial substantia propria of children with active trachoma [40], [41]. In vivo confocal microscopy of the conjunctival surface has shown that most of the cellular infiltrate in active and scarring trachoma is concentrated just below the epithelium, supporting the importance of the epithelial cell layer as a source of chemotactic factors [111].
For both active and scarring trachoma conjunctival transcriptome, studies found prominent innate immune responses [50], [99]. In children with active disease and/or Ct infection, there was marked enrichment of neutrophil and NK-cell-related transcripts [50]. In addition, several PRR and chemokines including the neutrophil chemotactic factor CXCL5 were increased [103]. In adults with scarring and little Ct infection, there was also strong evidence for an innate immune response, with some of the most abundant increases in gene expression found for pro-inflammatory mediators such as Psoriasin-1 (S100A7), IL-1B, and CXCL5 [99], [100]. These factors induce neutrophil chemotaxis and were particularly increased in inflamed cases. The importance of the chemokine response in trachoma is further supported by the finding that genetic variation across the IL-8 locus, defined by haplotypes of multiple single nucleotide polymorphisms (SNPs), was associated with scarring [112].
TNFα, while not specific to innate immune processes, is a key cytokine in acute inflammation and has been associated with scarring trachoma in several studies: a SNP in the TNFA promoter region, elevated levels in tear fluid, and increased secretion from peripheral blood mononuclear cells (PBMCs) from scarred subjects stimulated with EBs [101], [113]–[115]. Increased transcript levels of TNFA, as well as IL-1B, have also been associated with active disease/infection [33], [61], [116].
There is increasing evidence that nonchlamydial bacterial infection could play a role in the pathogenesis of trachoma and that this is likely to be through innate mechanisms. While infection with Ct is often found in children with active trachoma, it is only rarely identified in adults, as discussed above. Infection with bacteria other than Ct, however, is more common in individuals with conjunctival scarring, trichiasis, or its recurrence after surgery, compared to controls, and it is also more common in active disease [36], [37], [117]–[119]. This nonchlamydial bacterial infection is associated with elevated expression of a number of antimicrobial peptides, pro-inflammatory mediators, and modifiers of the extracellular matrix (ECM) including CXCL5, S100A7, DEFB4A, IL-1B, and MMPs 1, 9, 10, and 12 [100], [103], [120]. It is tempting to postulate that the conjunctiva of individuals who have been repeatedly infected with Ct undergoes morphological/immunological changes, rendering it susceptible to nonchlamydial bacterial infection, and that this subsequent infection causes an exaggerated inflammatory response with propagation of the scarring process. However, it is also possible that nonchlamydial bacterial infection may itself promote Ct infection or the different types of infection may act synergistically.
A Role for IL-17A in Trachoma?
Neither the “immunological” nor “cellular paradigms” completely accommodate the published data regarding trachoma pathogenesis. Initial studies suggest that IL-17A may be important in trachoma pathogenesis [50], [103]. IL-17A is the signature cytokine of Th17 cells, a CD4+ T-cell population that act in an antigen-specific manner [121]. However, it is also produced by several other cell types, notably innate immune cells (γδ T cells, NK cells, macrophages, neutrophils), and it can contribute to innate inflammatory responses [122], [123]. The IL-17A receptor is found on various cell types including dendritic cells, lymphocytes, epithelial cells, and fibroblasts [124]. IL-17A is pro-inflammatory and plays an important role in host immunity to extracellular and some intracellular pathogens [125]. Recently, it has become apparent that IL-17A may contribute to fibrosis through several mechanisms including epithelial-mesenchymal transition (EMT) and increased collagen production in a TGFβ1-dependant manner [126]. Infection of mice with Mycobacterium tuberculosis followed by repeated BCG injections led to an IL-17A–dominated response, which was refractory to regulation by IFNγ [127]. This caused extensive lung tissue damage by neutrophils that could be reduced by anti-IL17A antibody. These observations from TB may have parallels with trachoma, with IL-17A playing a major part in immunopathology, and could have implications for vaccine strategies.
Tissue Damage and Fibrogenesis in Trachoma
Scarring develops when normal tissue architecture is disrupted and replaced by excessive connective tissue through the abnormal accumulation of ECM components. Tissue damage can be mediated through a variety of cell types and mechanisms. Neutrophil infiltration appears important in trachoma: they have been identified in conjunctival biopsies of trachomatous tissue, they produce toxic reactive oxygen and nitrogen species that damage host tissue in animal models of genital tract infection, and they can also produce MMPs [48], [128]. Macrophages are also found in trachomatous tissue and may be important effector cells for tissue damage in chlamydial infection [48].
The matrix metalloproteinases are a group of more than 25 endopeptidases with multiple, complex functions. While MMPs are required for normal tissue homeostasis, there is also evidence that they play a role in the pathogenesis of a range of inflammatory-fibrotic diseases [129]–[131]. An important step is the disruption of the basement membrane, aiding the recruitment of inflammatory cells [132]. MMPs also have wide-ranging effects on inflammatory and immune processes such as modulating chemokine activity and activation of TGFβ, IL-1β, and TNFα. [133]. They are known to be important in a number of ocular surface diseases, and inhibition of MMP activity has been shown to reduce conjunctival scarring after glaucoma surgery [134], [135].
MMP9 in particular has been studied extensively in trachoma. It is part of the neutrophil lysosome and mediates epithelial dissolution associated with infection through degradation of type IV collagen [128]. Mouse models of genital tract infection have shown a reduced rate of scarring sequelae in MMP9 KO animals and that infection is associated with increased MMP9 in genital tract tissue (determined by zymography and gene expression) [15], [136]. Children with active trachoma have increased amounts of MMP9 (determined by immunohistochemistry, zymography, and gene expression analysis) [61], [137]. Scarring trachoma in adults is associated with increased expression of MMP9 and a coding SNP that is adjacent to the active binding site of the MMP9 enzyme [61], [100], [138]. Scarring trachoma is also associated with differential expression of MMPs 7, 9, 10, and 12 and tissue inhibitor of MMP (TIMP)-1, and recurrence of trichiasis after surgery is associated with an altered MMP1/TIMP1 transcript ratio [98]–[100], [120].
The production of scar tissue has not been extensively studied in trachoma. However, it probably originates from activated fibroblasts that are stimulated to produce collagen by various pro-fibrogenic mediators (TGF-β, PDGF, CTGF, and bFGF) [48], [103], [139]. Chemokines have also been shown to act as fibrogenic mediators, in particular the CC- and CXC-chemokine families, and various members of these families have been associated with scarring including the pro-fibrogenic CCL18 [96], [99], [101]–[103], [132].
Programmatic Implications
Trachoma is still endemic in over 50 countries and current trachoma control strategies face major obstacles. With regards to mass antibiotic distribution, the coverage levels in practice are often disappointing; there are concerns that mass distribution of azithromycin may lead to an increase in antibiotic resistance, and the “arrested immunity hypothesis,” put forward in the context of genital tract infection, suggests that shortening the duration of chlamydial infection with treatment results in population-wide reductions in protective immunity [140]–[142]. As well as being potentially the most effective way of reducing blindness from trachoma, a chlamydial vaccine could be very cost effective and have major benefits for genital tract disease caused by Ct [143], [144].
The human trachoma vaccine studies in the 1960s using inactivated whole EB tended to show only partial, short-term, serovar-specific protection, which was little better than natural immunity and may have resulted in earlier disease onset in some vaccinated individuals [67], [68], [83], [145]–[148]. Understanding the immunobiology of trachoma, including both how infection is successfully cleared and its pathogenesis, is important in the rational design of an effective vaccine that avoids immunopathology. A recent study employed a plasmid-deficient strain of Ct to immunise cynomolgus macaque monkeys [149]. Attenuated organisms were inoculated directly onto the conjunctival surface and caused minimal/absent ocular inflammation, despite repeated innoculation. On rechallenge with plasmid-bearing organisms, previously infected animals shed markedly less infectious organisms than controls, and three out of six animals showed minimal inflammatory changes compared to marked inflammation in all six control animals. These encouraging results, as well as implicating the Ct plasmid as an important virulence factor, support further work into vaccine development. While plasmid-deficient strains of Ct are naturally isolated from populations at risk of trachoma, the manipulation of virulence factors and the development of vaccine candidates may be helped by recent advances in Ct genetic transformation [150].
Conclusion
The immunology and pathogenesis of trachoma is a challenging but fascinating area of study. Despite considerable efforts, the mechanisms of protective immunity in trachoma remain elusive, although data from animal models suggest these involve IFNγ-dependent CMI responses. Recent data point to the importance of the human innate immune response, epithelial cell responses, EMT, and possible IL-17A involvement in both active disease and the pathogenesis of scarring. The MMPs are important effector enzymes in this tissue damage. There is still a pressing need for further research to better understand this ancient disease. The development of a vaccine would help to overcome the many obstacles that lie in the way of eliminating blinding trachoma.
Key Learning Points
The immune response to Ct provides only partial protection against re-infection and is important in causing tissue scarring and blindness.
Recent evidence has supported the “cellular paradigm” of disease pathogenesis, which states that infected epithelial cell responses drive pathology through the release of various mediators.
A trachoma vaccine would offer significant advantages to trachoma control, but vaccine development is impeded by a limited understanding of disease mechanisms.
Key Papers in the Field
Stephens RS (2003) The cellular paradigm of chlamydial pathogenesis. Trends Microbiol 11: 44–51.
Darville T, Hiltke TJ Pathogenesis of genital tract disease due to Chlamydia trachomatis. J Infect Dis 201 Suppl 2: S114–S125.
Burton MJ, Rajak SN, Bauer J, Weiss HA, Tolbert SB, et al. (2011) Conjunctival transcriptome in scarring trachoma. Infect Immun 79: 499–511.
Kari L, Whitmire WM, Olivares-Zavaleta N, Goheen MM, Taylor LD, et al. (2011) A live-attenuated chlamydial vaccine protects against trachoma in nonhuman primates. J Exp Med 208: 2217–2223.
Supporting Information
Table S1: Human trachoma genetic studies; Table S2: Histology and immunohistochemistry studies using human tarsal conjunctival biopsies; Table S3: Quantitative gene expression and microarray studies from human tarsal conjunctival swab samples; Table S4: Lymphoproliferative and cytokine studies using human PBMCs; Table S5: Antibody/B-cell responses from human serum, conjunctival, and tear samples; Table S6: Miscellaneous human trachoma studies.
(DOCX)
Funding Statement
This study was funded by grants from the Wellcome Trust (080741/Z/06/Z and 079246/Z/06/Z) and by a grant for the British Council for the Prevention of Blindness. VHH is supported by a fellowship grant from the British Council for the Prevention of Blindness (Barrie Jones Fellowship). The funders had no part in the study design; in the collection, analysis, and interpretation of data; in the writing of the report; and in the decision to submit the paper for publication.
References
- 1. Resnikoff S, Pascolini D, Etya'ale D, Kocur I, Pararajasegaram R, et al. (2004) Global data on visual impairment in the year 2002. Bull World Health Organ 82: 844–851. [PMC free article] [PubMed] [Google Scholar]
- 2. Mariotti SP, Pascolini D, Rose-Nussbaumer J (2009) Trachoma: global magnitude of a preventable cause of blindness. Br J Ophthalmol 93: 563–568. [DOI] [PubMed] [Google Scholar]
- 3.World Health Organization (2002) Global prevalence and incidence of selectted curable sexually transmitted infections overview and estimates. http://www.who.int/hiv/pub/sti/en/who_hiv_aids_2001.02.pdf.
- 4. Caldwell HD, Wood H, Crane D, Bailey R, Jones RB, et al. (2003) Polymorphisms in Chlamydia trachomatis tryptophan synthase genes differentiate between genital and ocular isolates. J Clin Invest 111: 1757–1769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.WHO (1998) Global elimination of blinding trachoma. World Health Assembly Resolutin WHA 51.11. Geneva, Switzerland: WHO.
- 6. Kuper H, Solomon AW, Buchan J, Zondervan M, Foster A, et al. (2003) A critical review of the SAFE strategy for the prevention of blinding trachoma. Lancet Infect Dis 3: 372–381. [DOI] [PubMed] [Google Scholar]
- 7. Emerson PM, Burton M, Solomon AW, Bailey R, Mabey D (2006) The SAFE strategy for trachoma control: using operational research for policy, planning and implementation. Bull World Health Organ 84: 613–619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Taylor HR, Prendergast RA, Dawson CR, Schachter J, Silverstein AM (1981) An animal model for cicatrizing trachoma. Invest Ophthalmol Vis Sci 21: 422–433. [PubMed] [Google Scholar]
- 9. Taylor HR, Johnson SL, Prendergast RA, Schachter J, Dawson CR, et al. (1982) An animal model of trachoma II. The importance of repeated reinfection. Invest Ophthalmol Vis Sci 23: 507–515. [PubMed] [Google Scholar]
- 10. Grayston JT, Wang SP, Yeh LJ, Kuo CC (1985) Importance of reinfection in the pathogenesis of trachoma. Rev Infect Dis 7: 717–725. [DOI] [PubMed] [Google Scholar]
- 11. Taylor HR, Johnson SL, Schachter J, Caldwell HD, Prendergast RA (1987) Pathogenesis of trachoma: the stimulus for inflammation. J Immunol 138: 3023–3027. [PubMed] [Google Scholar]
- 12. Taylor HR (1985) Ocular models of chlamydial infection. Rev Infect Dis 7: 737–740. [DOI] [PubMed] [Google Scholar]
- 13.Taylor-Robinson D (1986) The role of animal models in chlamydial research. In: Oriel D, Ridgway G, Schachter J, Taylor-Robinson D, Ward M, editors. Surrey. Cambridge University Press. pp. 355–366.
- 14. Shah AA, Schripsema JH, Imtiaz MT, Sigar IM, Kasimos J, et al. (2005) Histopathologic changes related to fibrotic oviduct occlusion after genital tract infection of mice with Chlamydia muridarum. Sex Transm Dis 32: 49–56. [DOI] [PubMed] [Google Scholar]
- 15. Ramsey KH, Sigar IM, Schripsema JH, Shaba N, Cohoon KP (2005) Expression of matrix metalloproteinases subsequent to urogenital Chlamydia muridarum infection of mice. Infect Immun 73: 6962–6973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Lacy HM, Bowlin AK, Hennings L, Scurlock AM, Nagarajan UM, et al. (2011) Essential role for neutrophils in pathogenesis and adaptive immunity in Chlamydia caviae ocular infections. Infect Immun 79: 1889–1897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Treharne JD, Shallal A (1991) The antigenic specificity of the humoral immune response to primary and repeated ocular infections of the guinea pig with the GPIC agent (Chlamydia psittaci). Eye 5 (Pt 3) 299–304. [DOI] [PubMed] [Google Scholar]
- 18. Mestas J, Hughes CC (2004) Of mice and not men: differences between mouse and human immunology. J Immunol 172: 2731–2738. [DOI] [PubMed] [Google Scholar]
- 19. Wright HR, Taylor HR (2005) Clinical examination and laboratory tests for estimation of trachoma prevalence in a remote setting: what are they really telling us? Lancet Infect Dis 5: 313–320. [DOI] [PubMed] [Google Scholar]
- 20. Bailey RL, Hampton TJ, Hayes LJ, Ward ME, Whittle HC, et al. (1994) Polymerase chain reaction for the detection of ocular chlamydial infection in trachoma-endemic communities. J Infect Dis 170: 709–712. [DOI] [PubMed] [Google Scholar]
- 21. West SK (2004) Trachoma: new assault on an ancient disease. Prog Retin Eye Res 23: 381–401. [DOI] [PubMed] [Google Scholar]
- 22. Emerson PM, Cairncross S, Bailey RL, Mabey DC (2000) Review of the evidence base for the ‘F’ and ‘E’ components of the SAFE strategy for trachoma control. Trop Med Int Health 5: 515–527. [DOI] [PubMed] [Google Scholar]
- 23. Hu VH, Harding-Esch EM, Burton MJ, Bailey RL, Kadimpeul J, et al. (2010) Epidemiology and control of trachoma: systematic review. Trop Med Int Health 15: 673–691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. West SK, Munoz B, Mkocha H, Hsieh YH, Lynch MC (2001) Progression of active trachoma to scarring in a cohort of Tanzanian children. Ophthalmic Epidemiol 8: 137–144. [DOI] [PubMed] [Google Scholar]
- 25.Dawson CR MR, Daghfous R, Juster R, Schachter J (1990) What clinical signs are critical in evaluating the impact of intervention in trachoma? Chlamydia Infections. Proceedings of the Seventh International Symposium on Human Chlamydial Infections. London: Cambridge University Press. pp. 271–275.
- 26. Munoz B, Bobo L, Mkocha H, Lynch M, Hsieh YH, et al. (1999) Incidence of trichiasis in a cohort of women with and without scarring. Int J Epidemiol 28: 1167–1171. [DOI] [PubMed] [Google Scholar]
- 27. Burton MJ, Bowman RJ, Faal H, Aryee EA, Ikumapayi UN, et al. (2006) The long-term natural history of trachomatous trichiasis in the Gambia. Invest Ophthalmol Vis Sci 47: 847–852. [DOI] [PubMed] [Google Scholar]
- 28. Wolle MA, Munoz BE, Mkocha H, West SK (2009) Constant ocular infection with Chlamydia trachomatis predicts risk of scarring in children in Tanzania. Ophthalmology 116: 243–247. [DOI] [PubMed] [Google Scholar]
- 29. Congdon N, West S, Vitale S, Katala S, Mmbaga BB (1993) Exposure to children and risk of active trachoma in Tanzanian women. Am J Epidemiol 137: 366–372. [DOI] [PubMed] [Google Scholar]
- 30. King JD, Ngondi J, Gatpan G, Lopidia B, Becknell S, et al. (2008) The burden of trachoma in ayod county of southern Sudan. PLoS Negl Trop Dis 2: e299 doi:10.1371/journal.pntd.0000299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Ngondi J, Ole-Sempele F, Onsarigo A, Matende I, Baba S, et al. (2006) Blinding trachoma in postconflict southern Sudan. PLoS Med 3: e478 doi:10.1371/journal.pmed.0030478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.TaylorHR (2008) Trachoma: a blinding scourge from the Bronze Age to the twenty-first century. Melbourne: Haddington Press.
- 33. Faal N, Bailey RL, Sarr I, Joof H, Mabey DC, et al. (2005) Temporal cytokine gene expression patterns in subjects with trachoma identify distinct conjunctival responses associated with infection. Clin Exp Immunol 142: 347–353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Wyrick PB (2010) Chlamydia trachomatis persistence in vitro: an overview. J Infect Dis 201 Suppl 2: S88–S95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Smith A, Munoz B, Hsieh YH, Bobo L, Mkocha H, et al. (2001) OmpA genotypic evidence for persistent ocular Chlamydia trachomatis infection in Tanzanian village women. Ophthalmic Epidemiol 8: 127–135. [DOI] [PubMed] [Google Scholar]
- 36. Burton MJ, Kinteh F, Jallow O, Sillah A, Bah M, et al. (2005) A randomised controlled trial of azithromycin following surgery for trachomatous trichiasis in the Gambia. Br J Ophthalmol 89: 1282–1288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Burton MJ, Adegbola RA, Kinteh F, Ikumapayi UN, Foster A, et al. (2007) Bacterial infection and trachoma in the gambia: a case control study. Invest Ophthalmol Vis Sci 48: 4440–4444. [DOI] [PubMed] [Google Scholar]
- 38. West ES, Mkocha H, Munoz B, Mabey D, Foster A, et al. (2005) Risk factors for postsurgical trichiasis recurrence in a trachoma-endemic area. Invest Ophthalmol Vis Sci 46: 447–453. [DOI] [PubMed] [Google Scholar]
- 39. Bailey R, Duong T, Carpenter R, Whittle H, Mabey D (1999) The duration of human ocular Chlamydia trachomatis infection is age dependent. Epidemiol Infect 123: 479–486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Abrahams C, Ballard RC, Sutter EE (1979) The pathology of trachoma in a black South African population. Light microscopical, histochemical and electron microscopical findings. S Afr Med J 55: 1115–1118. [PubMed] [Google Scholar]
- 41. el-Asrar AM, Van den Oord JJ, Geboes K, Missotten L, Emarah MH, et al. (1989) Immunopathology of trachomatous conjunctivitis. Br J Ophthalmol 73: 276–282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Guzey M, Ozardali I, Basar E, Aslan G, Satici A, et al. (2000) A survey of trachoma: the histopathology and the mechanism of progressive cicatrization of eyelid tissues. Ophthalmologica 214: 277–284. [DOI] [PubMed] [Google Scholar]
- 43. al-Rajhi AA, Hidayat A, Nasr A, al-Faran M (1993) The histopathology and the mechanism of entropion in patients with trachoma. Ophthalmology 100: 1293–1296. [DOI] [PubMed] [Google Scholar]
- 44. Reacher MH, Pe'er J, Rapoza PA, Whittum-Hudson JA, Taylor HR (1991) T cells and trachoma. Their role in cicatricial disease. Ophthalmology 98: 334–341. [DOI] [PubMed] [Google Scholar]
- 45. Rasmussen SJ, Eckmann L, Quayle AJ, Shen L, Zhang YX, et al. (1997) Secretion of proinflammatory cytokines by epithelial cells in response to Chlamydia infection suggests a central role for epithelial cells in chlamydial pathogenesis. J Clin Invest 99: 77–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Darville T, Hiltke TJ (201) Pathogenesis of genital tract disease due to Chlamydia trachomatis. J Infect Dis 201 Suppl 2: S114–S125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Johnson RM (2004) Murine oviduct epithelial cell cytokine responses to Chlamydia muridarum infection include interleukin-12-p70 secretion. Infect Immun 72: 3951–3960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Abu el-Asrar AM, Geboes K, Tabbara KF, al-Kharashi SA, Missotten L, et al. (1998) Immunopathogenesis of conjunctival scarring in trachoma. Eye 12 (Pt 3a) 453–460. [DOI] [PubMed] [Google Scholar]
- 49. Mabey DC, Bailey RL, Dunn D, Jones D, Williams JH, et al. (1991) Expression of MHC class II antigens by conjunctival epithelial cells in trachoma: implications concerning the pathogenesis of blinding disease. J Clin Pathol 44: 285–289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Natividad A, Freeman TC, Jeffries D, Burton MJ, Mabey DC, et al. (2010) Human conjunctival transcriptome analysis reveals the prominence of innate defense in Chlamydia trachomatis infection. Infect Immun 78: 4895–4911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Gall A, Horowitz A, Joof H, Natividad A, Tetteh K, et al. (2011) Systemic effector and regulatory immune responses to chlamydial antigens in trachomatous trichiasis. Front Microbiol 2: 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Rank RG, Soderberg LS, Barron AL (1985) Chronic chlamydial genital infection in congenitally athymic nude mice. Infect Immun 48: 847–849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Ramsey KH, Rank RG (1991) Resolution of chlamydial genital infection with antigen-specific T-lymphocyte lines. Infect Immun 59: 925–931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Morrison SG, Farris CM, Sturdevant GL, Whitmire WM, Morrison RP (2011) Murine Chlamydia trachomatis genital infection is unaltered by depletion of CD4+ T cells and diminished adaptive immunity. J Infect Dis 203: 1120–1128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Brunham RC, Rey-Ladino J (2005) Immunology of Chlamydia infection: implications for a Chlamydia trachomatis vaccine. Nat Rev Immunol 5: 149–161. [DOI] [PubMed] [Google Scholar]
- 56. Thomas SM, Garrity LF, Brandt CR, Schobert CS, Feng GS, et al. (1993) IFN-gamma-mediated antimicrobial response. Indoleamine 2,3-dioxygenase-deficient mutant host cells no longer inhibit intracellular Chlamydia spp. or Toxoplasma growth. J Immunol 150: 5529–5534. [PubMed] [Google Scholar]
- 57. Rapoza PA, Tahija SG, Carlin JP, Miller SL, Padilla ML, et al. (1991) Effect of interferon on a primary conjunctival epithelial cell model of trachoma. Invest Ophthalmol Vis Sci 32: 2919–2923. [PubMed] [Google Scholar]
- 58. Ramsey KH, Miranpuri GS, Sigar IM, Ouellette S, Byrne GI (2001) Chlamydia trachomatis persistence in the female mouse genital tract: inducible nitric oxide synthase and infection outcome. Infect Immun 69: 5131–5137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Igietseme JU, Ananaba GA, Candal DH, Lyn D, Black CM (1998) Immune control of Chlamydial growth in the human epithelial cell line RT4 involves multiple mechanisms that include nitric oxide induction, tryptophan catabolism and iron deprivation. Microbiol Immunol 42: 617–625. [DOI] [PubMed] [Google Scholar]
- 60. Bailey RL, Holland MJ, Whittle HC, Mabey DC (1995) Subjects recovering from human ocular chlamydial infection have enhanced lymphoproliferative responses to chlamydial antigens compared with those of persistently diseased controls. Infect Immun 63: 389–392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Burton MJ, Bailey RL, Jeffries D, Mabey DC, Holland MJ (2004) Cytokine and fibrogenic gene expression in the conjunctivas of subjects from a Gambian community where trachoma is endemic. Infect Immun 72: 7352–7356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Faal N, Bailey RL, Jeffries D, Joof H, Sarr I, et al. (2006) Conjunctival FOXP3 expression in trachoma: do regulatory T cells have a role in human ocular Chlamydia trachomatis infection? PLoS Med 3: e266 doi:10.1371/journal.pmed.0030266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Holland MJ, Faal N, Jeffries D, Joof H, Sarr I, et al.. (2010) Human ocular Chlamydia trachomatis disease, infection and immune responses examine using quantitative conjunctival gene expression with intensive longitudianl follow-up and gene co-expression analysis. Salzburg. pp. 145–148.
- 64. Jawetz E, Rose L, Hanna L, Thygeson P (1965) Experimental inclusion conjunctivitis in man: measurements of infectivity and resistance. JAMA 194: 620–632. [PubMed] [Google Scholar]
- 65. Dawson C, Jawetz E, Hanna L, Rose L, Wood TR, et al. (1966) Experimental inclusion conjunctivitis in man. II. Partial resistance to reinfection. Am J Epidemiol 84: 411–425. [DOI] [PubMed] [Google Scholar]
- 66. Tarizzo ML, Nataf R, Nabli B (1967) Experimental inoculation of thirteen volunteers with agent isolated from inclusion conjunctivitis. Am J Ophthalmol 63 Suppl 1120–1128. [DOI] [PubMed] [Google Scholar]
- 67. Nichols RL, Bell SD Jr, Haddad NA, Bobb AA (1969) Studies on trachoma. VI. Microbiological observations in a field trial in Saudi Arabia of bivalent rachoma vaccine at three dosage levels. Am J Trop Med Hyg 18: 723–730. [PubMed] [Google Scholar]
- 68. Sowa S, Sowa J, Collier LH, Blyth WA (1969) Trachoma vaccine field trials in The Gambia. J Hyg (Lond) 67: 699–717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Grassly NC, Ward ME, Ferris S, Mabey DC, Bailey RL (2008) The natural history of trachoma infection and disease in a gambian cohort with frequent follow-up. PLoS Negl Trop Dis 2: e341 doi: 10.1371/journal.pntd.0000341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Rank RG, Whittum-Hudson JA (2010) Protective immunity to chlamydial genital infection: evidence from animal studies. J Infect Dis 201 Suppl 2: S168–S177. [DOI] [PubMed] [Google Scholar]
- 71. Cohen CR, Koochesfahani KM, Meier AS, Shen C, Karunakaran K, et al. (2005) Immunoepidemiologic profile of Chlamydia trachomatis infection: importance of heat-shock protein 60 and interferon- gamma. J Infect Dis 192: 591–599. [DOI] [PubMed] [Google Scholar]
- 72. Bailey RL, Kajbaf M, Whittle HC, Ward ME, Mabey DC (1993) The influence of local antichlamydial antibody on the acquisition and persistence of human ocular chlamydial infection: IgG antibodies are not protective. Epidemiol Infect 111: 315–324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Holland MJ, Bailey RL, Hayes LJ, Whittle HC, Mabey DC (1993) Conjunctival scarring in trachoma is associated with depressed cell-mediated immune responses to chlamydial antigens. J Infect Dis 168: 1528–1531. [DOI] [PubMed] [Google Scholar]
- 74. Lu C, Holland MJ, Gong S, Peng B, Bailey RL, et al. (2012) Genome-wide identification of Chlamydia trachomatis antigens associated with trachomatous trichiasis. Invest Ophthalmol Vis Sci 53: 2551–2559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Ramsey KH, Soderberg LS, Rank RG (1988) Resolution of chlamydial genital infection in B-cell-deficient mice and immunity to reinfection. Infect Immun 56: 1320–1325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Su H, Feilzer K, Caldwell HD, Morrison RP (1997) Chlamydia trachomatis genital tract infection of antibody-deficient gene knockout mice. Infect Immun 65: 1993–1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Morrison SG, Su H, Caldwell HD, Morrison RP (2000) Immunity to murine Chlamydia trachomatis genital tract reinfection involves B cells and CD4(+) T cells but not CD8(+) T cells. Infect Immun 68: 6979–6987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Morrison SG, Morrison RP (2001) Resolution of secondary Chlamydia trachomatis genital tract infection in immune mice with depletion of both CD4+ and CD8+ T cells. Infect Immun 69: 2643–2649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Morrison SG, Morrison RP (2005) A predominant role for antibody in acquired immunity to chlamydial genital tract reinfection. J Immunol 175: 7536–7542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. Crane DD, Carlson JH, Fischer ER, Bavoil P, Hsia RC, et al. (2006) Chlamydia trachomatis polymorphic membrane protein D is a species-common pan-neutralizing antigen. Proc Natl Acad Sci U S A 103: 1894–1899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Silverstein AM (1974) The immunologic modulation of infectious disease pathogenesis. Friedenwald Lecture, 1973. Invest Ophthalmol 13: 560–574. [PubMed] [Google Scholar]
- 82. Burton MJ, Holland MJ, Faal N, Aryee EA, Alexander ND, et al. (2003) Which members of a community need antibiotics to control trachoma? Conjunctival Chlamydia trachomatis infection load in Gambian villages. Invest Ophthalmol Vis Sci 44: 4215–4222. [DOI] [PubMed] [Google Scholar]
- 83. Woolridge RL, Grayston JT, Chang IH, Cheng KH, Yang CY, et al. (1967) Field trial of a monovalent and of a bivalent mineral oil adjuvant trachoma vaccine in Taiwan school children. Am J Ophthalmol 63 Suppl 1645–1650. [DOI] [PubMed] [Google Scholar]
- 84. Taylor HR, Johnson SL, Schachter J, Prendergast RA (1983) An animal model of trachoma: IV. The failure of local immunosuppression to reveal inapparent infection. Invest Ophthalmol Vis Sci 24: 647–650. [PubMed] [Google Scholar]
- 85. Wang SP, Grayston JT, Alexander ER (1967) Trachoma vaccine studies in monkeys. Am J Ophthalmol 63 Suppl 1615–1630. [DOI] [PubMed] [Google Scholar]
- 86. Stephens RS (2003) The cellular paradigm of chlamydial pathogenesis. Trends Microbiol 11: 44–51. [DOI] [PubMed] [Google Scholar]
- 87. Watkins NG, Hadlow WJ, Moos AB, Caldwell HD (1986) Ocular delayed hypersensitivity: a pathogenetic mechanism of chlamydial-conjunctivitis in guinea pigs. Proc Natl Acad Sci U S A 83: 7480–7484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Morrison RP, Belland RJ, Lyng K, Caldwell HD (1989) Chlamydial disease pathogenesis. The 57-kD chlamydial hypersensitivity antigen is a stress response protein. J Exp Med 170: 1271–1283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Morrison RP, Lyng K, Caldwell HD (1989) Chlamydial disease pathogenesis. Ocular hypersensitivity elicited by a genus-specific 57-kD protein. J Exp Med 169: 663–675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Patton DL, Sweeney YT, Kuo CC (1994) Demonstration of delayed hypersensitivity in Chlamydia trachomatis salpingitis in monkeys: a pathogenic mechanism of tubal damage. J Infect Dis 169: 680–683. [DOI] [PubMed] [Google Scholar]
- 91. Lichtenwalner AB, Patton DL, Van Voorhis WC, Sweeney YT, Kuo CC (2004) Heat shock protein 60 is the major antigen which stimulates delayed-type hypersensitivity reaction in the macaque model of Chlamydia trachomatis salpingitis. Infect Immun 72: 1159–1161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Rank RG, Dascher C, Bowlin AK, Bavoil PM (1995) Systemic immunization with Hsp60 alters the development of chlamydial ocular disease. Invest Ophthalmol Vis Sci 36: 1344–1351. [PubMed] [Google Scholar]
- 93. Peeling RW, Bailey RL, Conway DJ, Holland MJ, Campbell AE, et al. (1998) Antibody response to the 60-kDa chlamydial heat-shock protein is associated with scarring trachoma. J Infect Dis 177: 256–259. [DOI] [PubMed] [Google Scholar]
- 94. Eckert LO, Hawes SE, Wolner-Hanssen P, Money DM, Peeling RW, et al. (1997) Prevalence and correlates of antibody to chlamydial heat shock protein in women attending sexually transmitted disease clinics and women with confirmed pelvic inflammatory disease. J Infect Dis 175: 1453–1458. [DOI] [PubMed] [Google Scholar]
- 95. Toye B, Laferriere C, Claman P, Jessamine P, Peeling R (1993) Association between antibody to the chlamydial heat-shock protein and tubal infertility. J Infect Dis 168: 1236–1240. [DOI] [PubMed] [Google Scholar]
- 96. Wynn TA (2004) Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat Rev Immunol 4: 583–594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Holland MJ, Bailey RL, Conway DJ, Culley F, Miranpuri G, et al. (1996) T helper type-1 (Th1)/Th2 profiles of peripheral blood mononuclear cells (PBMC); responses to antigens of Chlamydia trachomatis in subjects with severe trachomatous scarring. Clin Exp Immunol 105: 429–435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Holland MJ, Jeffries D, Pattison M, Korr G, Gall A, et al. (2010) Pathway-focused arrays reveal increased matrix metalloproteinase-7 (matrilysin) transcription in trachomatous trichiasis. Invest Ophthalmol Vis Sci 51: 3893–3902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99. Burton MJ, Rajak SN, Bauer J, Weiss HA, Tolbert SB, et al. (2011) Conjunctival transcriptome in scarring trachoma. Infect Immun 79: 499–511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Hu VH, Weiss HA, Ramadhani AM, Tolbert SB, Massae P, et al. (2012) Innate immune responses and modified extracellular matrix regulation characterize bacterial infection and cellular/connective tissue changes in scarring trachoma. Infect Immun 80: 121–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Skwor TA, Atik B, Kandel RP, Adhikari HK, Sharma B, et al. (2008) Role of secreted conjunctival mucosal cytokine and chemokine proteins in different stages of trachomatous disease. PLoS Negl Trop Dis 2: e264 doi:10.1371/journal.pntd.0000264 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Natividad A, Holland M, Mabey D, Bailey R (2010) Expression profiling of cyokines, chemokines and their receptors during human ocular chlamydial infection. Salzburg. pp. 149–152.
- 103. Burton MJ, Ramadhani A, Weiss HA, Hu V, Massae P, et al. (2011) Active trachoma is associated with increased conjunctival expression of IL17A and profibrotic cytokines. Infect Immun 79: 4977–4983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Kelly KA (2003) Cellular immunity and Chlamydia genital infection: induction, recruitment, and effector mechanisms. Int Rev Immunol 22: 3–41. [DOI] [PubMed] [Google Scholar]
- 105. Frazer LC, O'Connell CM, Andrews CW Jr, Zurenski MA, Darville T (2011) Enhanced neutrophil longevity and recruitment contribute to the severity of oviduct pathology during C. muridarum infection. Infect Immun 79 (10) 4029–4041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106. Roan NR, Starnbach MN (2008) Immune-mediated control of Chlamydia infection. Cell Microbiol 10: 9–19. [DOI] [PubMed] [Google Scholar]
- 107. Darville T, O'Neill JM, Andrews CW Jr, Nagarajan UM, Stahl L, et al. (2003) Toll-like receptor-2, but not Toll-like receptor-4, is essential for development of oviduct pathology in chlamydial genital tract infection. J Immunol 171: 6187–6197. [DOI] [PubMed] [Google Scholar]
- 108. Karimi O, Ouburg S, de Vries HJ, Pena AS, Pleijster J, et al. (2009) TLR2 haplotypes in the susceptibility to and severity of Chlamydia trachomatis infections in Dutch women. Drugs Today (Barc) 45 Suppl B: 67–74. [PubMed] [Google Scholar]
- 109. den Hartog JE, Lyons JM, Ouburg S, Fennema JS, de Vries HJ, et al. (2009) TLR4 in Chlamydia trachomatis infections: knockout mice, STD patients and women with tubal factor subfertility. Drugs Today (Barc) 45 Suppl B: 75–82. [PubMed] [Google Scholar]
- 110. Ouburg S, Lyons JM, Land JA, den Hartog JE, Fennema JS, et al. (2009) TLR9 KO mice, haplotypes and CPG indices in Chlamydia trachomatis infection. Drugs Today (Barc) 45 Suppl B: 83–93. [PubMed] [Google Scholar]
- 111. Hu VH, Massae P, Weiss HA, Cree IA, Courtright P, et al. (2010) In vivo confocal microscopy of trachoma in relation to normal tarsal conjunctiva. Ophthalmol 118 (4) 747–754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Natividad A, Hull J, Luoni G, Holland M, Rockett K, et al. (2009) Innate immunity in ocular Chlamydia trachomatis infection: contribution of IL8 and CSF2 gene variants to risk of trachomatous scarring in Gambians. BMC Med Genet 10: 138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Conway DJ, Holland MJ, Bailey RL, Campbell AE, Mahdi OS, et al. (1997) Scarring trachoma is associated with polymorphism in the tumor necrosis factor alpha (TNF-alpha) gene promoter and with elevated TNF-alpha levels in tear fluid. Infect Immun 65: 1003–1006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Natividad A, Hanchard N, Holland MJ, Mahdi OS, Diakite M, et al. (2007) Genetic variation at the TNF locus and the risk of severe sequelae of ocular Chlamydia trachomatis infection in Gambians. Genes Immun 8: 288–295. [DOI] [PubMed] [Google Scholar]
- 115. Atik B, Skwor TA, Kandel RP, Sharma B, Adhikari HK, et al. (2008) Identification of novel single nucleotide polymorphisms in inflammatory genes as risk factors associated with trachomatous trichiasis. PLoS ONE 3: e3600 doi:10.1371/journal.pone.0003600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Bobo L, Novak N, Mkocha H, Vitale S, West S, et al. (1996) Evidence for a predominant proinflammatory conjunctival cytokine response in individuals with trachoma. Infect Immun 64: 3273–3279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Burton MJ, Bowman RJ, Faal H, Aryee EA, Ikumapayi UN, et al. (2005) Long term outcome of trichiasis surgery in the Gambia. Br J Ophthalmol 89: 575–579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Burton MJ, Hu VH, Massae P, Burr SE, Chevallier C, et al. (2011) What is causing active trachoma? The role of nonchlamydial bacterial pathogens in a low prevalence setting. Invest Ophthalmol Vis Sci 52: 6012–6017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Hu VH, Massae P, Weiss HA, Chevallier C, Onyango JJ, et al. (2011) Bacterial infection in scarring trachoma. Invest Ophthalmol Vis Sci 52: 2181–2186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Burton MJ, Bailey RL, Jeffries D, Rajak SN, Adegbola RA, et al. (2010) Conjunctival expression of matrix metalloproteinase and proinflammatory cytokine genes after trichiasis surgery. Invest Ophthalmol Vis Sci 51: 3583–3590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Onishi RM, Gaffen SL (2010) Interleukin-17 and its target genes: mechanisms of interleukin-17 function in disease. Immunology 129: 311–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Reynolds JM, Angkasekwinai P, Dong C (2010) IL-17 family member cytokines: regulation and function in innate immunity. Cytokine Growth Factor Rev 21: 413–423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Khader SA, Gaffen SL, Kolls JK (2009) Th17 cells at the crossroads of innate and adaptive immunity against infectious diseases at the mucosa. Mucosal Immunol 2: 403–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Torrado E, Cooper AM (2010) IL-17 and Th17 cells in tuberculosis. Cytokine Growth Factor Rev 21: 455–462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Kolls JK, Khader SA (2010) The role of Th17 cytokines in primary mucosal immunity. Cytokine Growth Factor Rev 21: 443–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Mi S, Li Z, Yang HZ, Liu H, Wang JP, et al. (2011) Blocking IL-17A promotes the resolution of pulmonary inflammation and fibrosis via TGF-{beta}1-dependent and -independent mechanisms. J Immunol 187: 3003–3014. [DOI] [PubMed] [Google Scholar]
- 127. Cruz A, Fraga AG, Fountain JJ, Rangel-Moreno J, Torrado E, et al. (2010) Pathological role of interleukin 17 in mice subjected to repeated BCG vaccination after infection with Mycobacterium tuberculosis. J Exp Med 207: 1609–1616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Ramsey KH (2006) Alternative mechanisms of pathogenesis. Chlamydia: genomics and pathogenesis. Norfolk, UK: Horizon Bioscience. pp. 435–474.
- 129. Hu J, Van den Steen PE, Sang QX, Opdenakker G (2007) Matrix metalloproteinase inhibitors as therapy for inflammatory and vascular diseases. Nat Rev Drug Discov 6: 480–498. [DOI] [PubMed] [Google Scholar]
- 130. Chakrabarti S, Patel KD (2005) Matrix metalloproteinase-2 (MMP-2) and MMP-9 in pulmonary pathology. Exp Lung Res 31: 599–621. [DOI] [PubMed] [Google Scholar]
- 131. Wang JC (2005) Importance of plasma matrix metalloproteinases (MMP) and tissue inhibitors of metalloproteinase (TIMP) in development of fibrosis in agnogenic myeloid metaplasia. Leuk Lymphoma 46: 1261–1268. [DOI] [PubMed] [Google Scholar]
- 132. Wynn TA (2008) Cellular and molecular mechanisms of fibrosis. J Pathol 214: 199–210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Parks WC, Wilson CL, Lopez-Boado YS (2004) Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat Rev Immunol 4: 617–629. [DOI] [PubMed] [Google Scholar]
- 134. Wong TT, Sethi C, Daniels JT, Limb GA, Murphy G, et al. (2002) Matrix metalloproteinases in disease and repair processes in the anterior segment. Surv Ophthalmol 47: 239–256. [DOI] [PubMed] [Google Scholar]
- 135. Wong TT, Mead AL, Khaw PT (2005) Prolonged antiscarring effects of ilomastat and MMC after experimental glaucoma filtration surgery. Invest Ophthalmol Vis Sci 46: 2018–2022. [DOI] [PubMed] [Google Scholar]
- 136. Imtiaz MT, Distelhorst JT, Schripsema JH, Sigar IM, Kasimos JN, et al. (2007) A role for matrix metalloproteinase-9 in pathogenesis of urogenital Chlamydia muridarum infection in mice. Microbes Infect 9: 1561–1566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137. El-Asrar AM, Geboes K, Al-Kharashi SA, Al-Mosallam AA, Missotten L, et al. (2000) Expression of gelatinase B in trachomatous conjunctivitis. Br J Ophthalmol 84: 85–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Natividad A, Cooke G, Holland MJ, Burton MJ, Joof HM, et al. (2006) A coding polymorphism in matrix metalloproteinase 9 reduces risk of scarring sequelae of ocular Chlamydia trachomatis infection. BMC Med Genet 7: 40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Abu El-Asrar AM, Al-Kharashi SA, Missotten L, Geboes K (2006) Expression of growth factors in the conjunctiva from patients with active trachoma. Eye 20: 362–369. [DOI] [PubMed] [Google Scholar]
- 140. Thylefors B, Negrel AD, Pararajasegaram R (1992) [Epidemiological surveillance of trachoma: evaluation and perspective]. Rev Int Trach Pathol Ocul Trop Subtrop Sante Publique 69: 107–114. [PubMed] [Google Scholar]
- 141. Leach AJ, Shelby-James TM, Mayo M, Gratten M, Laming AC, et al. (1997) A prospective study of the impact of community-based azithromycin treatment of trachoma on carriage and resistance of Streptococcus pneumoniae. Clin Infect Dis 24: 356–362. [DOI] [PubMed] [Google Scholar]
- 142. Brunham RC, Rekart ML (2008) The arrested immunity hypothesis and the epidemiology of chlamydia control. Sex Transm Dis 35: 53–54. [DOI] [PubMed] [Google Scholar]
- 143. Woodall JP (2003) Proceedings of the Chlamydia Vaccine Development Colloquim. The Albert B Sabin Vaccine Institute [Google Scholar]
- 144. Frick KD, Colchero MA, Dean D (2004) Modeling the economic net benefit of a potential vaccination program against ocular infection with Chlamydia trachomatis. Vaccine 22: 689–696. [DOI] [PubMed] [Google Scholar]
- 145. Grayston JT, Woolridge RL, Wang SP, Yen CH, Yang CY, et al. (1963) Field studies of protection from infection by experimental trachoma virus vaccine in preschool-aged children on Taiwan. Proc Soc Exp Biol Med 112: 589–595. [DOI] [PubMed] [Google Scholar]
- 146. Grayston JT, Wang SP, Woolridge RL, Alexander ER (1964) Prevention of trachoma with vaccine. Arch Environ Health 8: 518–526. [DOI] [PubMed] [Google Scholar]
- 147. Nichols RL, Bell SD Jr, Murray ES, Haddad NA, Bobb AA (1966) Studies on trachoma. V. Clinical observations in a field trial of bivalent trachoma vaccine at three dosage levels in Saudi Arabia. Am J Trop Med Hyg 15: 639–647. [PubMed] [Google Scholar]
- 148. Clements C, Dhir SP, Grayston JT, Wang SP (1979) Long term follow-up study of a trachoma vaccine trial in villages of Northern India. Am J Ophthalmol 87: 350–353. [DOI] [PubMed] [Google Scholar]
- 149. Kari L, Whitmire WM, Olivares-Zavaleta N, Goheen MM, Taylor LD, et al. (2011) A live-attenuated chlamydial vaccine protects against trachoma in nonhuman primates. J Exp Med 208: 2217–2223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150. Wang Y, Kahane S, Cutcliffe LT, Skilton RJ, Lambden PR, et al. (2011) Development of a transformation system for Chlamydia trachomatis: restoration of glycogen biosynthesis by acquisition of a plasmid shuttle vector. PLoS Pathog 7: e1002258 doi:10.1371/journal.ppat.1002258 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Wolle MA, Munoz B, Mkocha H, West SK (2009) Age, sex, and cohort effects in a longitudinal study of trachomatous scarring. Invest Ophthalmol Vis Sci 50: 592–596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152. Bowman RJ, Jatta B, Cham B, Bailey RL, Faal H, et al. (2001) Natural history of trachomatous scarring in The Gambia: results of a 12-year longitudinal follow-up. Ophthalmology 108: 2219–2224. [DOI] [PubMed] [Google Scholar]
- 153. Bowman RJ, Faal H, Myatt M, Adegbola R, Foster A, et al. (2002) Longitudinal study of trachomatous trichiasis in the Gambia. Br J Ophthalmol 86: 339–343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Munoz B, Aron J, Turner V, West S (1997) Incidence estimates of late stages of trachoma among women in a hyperendemic area of central Tanzania. Trop Med Int Health 2: 1030–1038. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1: Human trachoma genetic studies; Table S2: Histology and immunohistochemistry studies using human tarsal conjunctival biopsies; Table S3: Quantitative gene expression and microarray studies from human tarsal conjunctival swab samples; Table S4: Lymphoproliferative and cytokine studies using human PBMCs; Table S5: Antibody/B-cell responses from human serum, conjunctival, and tear samples; Table S6: Miscellaneous human trachoma studies.
(DOCX)