

Abstract
Food-storing corvids use many cache-protection and pilfering strategies. We tested whether Eurasian jays (Garrulus glandarius) reduce the transfer of auditory information to a competitor when caching and pilfering. We gave jays a noisy and a quiet substrate to cache in. Compared with when alone, birds cached less in the noisy substrate when with a conspecific that could hear but could not see them caching. By contrast, jays did not change the amount cached in the noisy substrate when they were with a competitor that could see and hear them caching compared with when they were alone. Together, these results suggest that jays reduce auditory information during caching as a cache-protection strategy. By contrast, as pilferers, jays did not attempt to conceal their presence from a cacher and did not prefer a silent viewing perch over a noisy one when observing caching. However, birds vocalized less when watching caching compared with when they were alone, when they were watching a non-caching conspecific or when they were watching their own caches being pilfered. Pilfering jays may therefore attempt to suppress some types of auditory information. Our results raise the possibility that jays both understand and can attribute auditory perception to another individual.
Keywords: Eurasian jay, Garrulus glandarius, auditory information, cache-protection, pilfering, perception attribution
1. Introduction
Many animals cache food for later consumption. However, a large proportion of caches may be lost to both conspecific and heterospecific pilferers [1]. In corvids, the risk of conspecifics pilfering caches is particularly high because they remember the locations of caches that they have seen another individual make [2–6]. This observational spatial memory may have acted as a catalyst in an evolutionary arms race, leading to the development of sophisticated cache-protection and pilfering strategies [6,7]. Corvid cache-protection strategies limit opportunities for conspecifics to witness caching events [8,9]. In the western scrub-jay (Aphelocoma californica), for example, strategies include recovering items and moving them to new locations [10], caching at a distance from competitors or behind barriers [8] and caching in the shade [11].
Cache-protection strategies are not limited to the visual domain. When given a noisy (small pebbles) and a quiet (soil) substrate to cache in, western scrub-jays avoided the noisy substrate when caching in the presence of a competitor that could hear but could not see them, but did not do so when alone or when with a competitor that could both see and hear them [12]. Noises associated with caching, such as the movement of substrate, may alert potential pilferers to the presence of a caching individual and increase the likelihood of caches being stolen. Reducing the amount of auditory information that is available to eavesdropping conspecifics by caching in a quiet substrate therefore represents a cache-protection strategy.
To date, most studies of cache-protection and pilfering have tested western scrub-jays and ravens (Corvus corax) [7,13]. However, comparative studies of species that differ in their socio-ecology and caching behaviour are crucial to further investigate cache-protection and pilfering strategies, and their underlying cognition [7,14]. Ravens and western scrub-jays are moderate generalist cachers [15] that are also semi-social. Ravens routinely form non-kin groups and alliances when foraging [16,17], whereas western scrub-jays will tolerate non-breeding individuals on their territories and forage in flocks [18]. By contrast, territorial Eurasian jays (Garrulus glandarius) defend a territory with a mate and do not form closely associated groups to forage [19]. In the jays' preferred forest habitat [20], visual signals attenuate more rapidly than acoustic signals. Given their territorial nature, Eurasian jays are therefore likely to be sensitive to auditory information that indicates the presence of intruders in their territory. Additionally, the Eurasian jay is a highly specialized and intense cacher, with caches forming an important part of the jay's diet [15,21]. Despite jays being a territorial species, their caches are pilfered by both conspecifics and heterospecifics [21], and there is evidence that jays flexibly use a range of cache-protection behaviours in the visual domain [22]. In this study, we investigate whether Eurasian jays use strategies to limit the transfer of auditory information to potential competitors when caching and when pilfering.
2. Experiment 1
(a). Hypothesis and predictions
We gave Eurasian jays two caching substrates, one of which was noisy (fine gravel) and one which was quiet (sand). We tested the jays' preferences for caching in each substrate in three conditions: alone with no competitor present, with a competitor that could both see and hear caching, and with a competitor that could hear, but could not see caching. If jays reduce auditory information available to conspecifics as a cache-protection strategy, then compared with when alone individuals should avoid caching in the noisy substrate when a competitor can hear but cannot see them. If a competitor has visual access to a caching event, there is potentially little benefit in suppressing noises associated with caching. We therefore predicted that similar to western scrub-jays [12], compared with when caching in the presence of a competitor that can both see and hear them, Eurasian jays should preferentially avoid caching in the noisy substrate when with a competitor that can hear but cannot see them.
(b). Methods
Subjects came from a group of 12 (six male, six female) sexually mature, hand-raised Eurasian jays. Birds had a daily maintenance diet of soaked dog biscuits, bread, cheese, egg, vegetables, seeds, nuts and fruit. We tested subjects outside of breeding season when they were housed in an outdoor aviary complex (figure 1a). Five compartments in the complex were used for testing (A, B, C, D and E as shown in figure 1a). Compartments were separated by a wooden wall from the rest of the aviary and had wire mesh walls between them. Compartments B–D contained wooden platforms positioned 1 m above the floor, preventing access to the ground. Each compartment had a single perch positioned 2 m above the floor (1 m above the wooden platform in compartments B–D). The walls between compartments A and B, and D and E were covered with white sheets to create opaque barriers.
Figure 1.
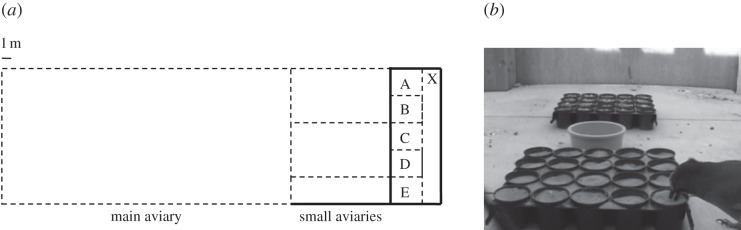
(a) A scale (1 : 300) aerial view of the main aviary (3 × 20 × 6 m), three small aviaries (top two: 3 × 6 × 2 m; bottom: 3 × 6 × 1 m) and five compartments (A, B, C, D, E; each measuring 3 × 1 × 1 m). Dashed lines represent wire mesh walls, and solid lines represent wooden walls. The floor consisted of large gravel throughout the aviary complex. The main aviary connected to the smaller aviaries via doors (0.7 × 2 m). Each compartment could be accessed from the small aviaries through opaque trap-doors (0.4 × 0.6 m) set 1.5 m off the ground. The experimenter, when present during a trial in experiment 2, stood in the 1.5 m wide corridor at the position marked X. (b) A video still of compartment B taken from the corridor, showing the arrangement of the trays and food bowl during a trial in experiment 1. The jay is caching in the ‘quiet’ substrate (sand). The noisy substrate (gravel) is in the far tray. The food bowl is positioned an equal distance between the two trays.
In total, 11 jays (six males, five females) were used in experiment 1. Trials took place from 10.00 to 12.30 between 18 October 2010–16 November 2010 and 27 September 2011–08 December 2011. To ensure that subjects were mildly hungry, the experimenter (R.C.S.) removed the maintenance diet approximately 2 h before a trial began. The experimenter enclosed the subject (n = 8; four males, four females) in compartment B, C or D (figure 1a) and gave them a bowl containing 30 peanuts and two caching trays (figure 1b). Individuals not being tested were excluded from the small aviaries and testing compartments (figure 1a). Caching trays were 20 × 25 cm seedling trays containing 20 pots (diameter = 5 cm; figure 1b); each was painted with a unique colour and pattern to create visually distinct locations. One tray contained sand (the ‘quiet’ substrate) and the other contained fine gravel (the ‘noisy’ substrate). The jays had not experienced caching in either substrate prior to this experiment. Between trials, we randomized the location of the ‘noisy’ and ‘quiet’ trays and gave the subject novel tray combinations. Trials lasted 30 min, after which the subject was released and the food bowl and trays were removed from the compartment. The location and number of caches made in the trays and in the surrounding compartment were recorded. Caches made in out-of-tray locations were considered to be quiet [12].
We tested subjects in three conditions. In the ‘alone’ condition, no conspecific was present. In the ‘seen and heard’ condition, the subject was tested in compartment B, C or D, with a conspecific in an adjacent compartment (B, C or D) separated by a wire mesh wall. In the ‘heard but not seen’ condition, compartments A and B, or D and E were used: the subject was tested in B or D and, a conspecific was in the adjacent compartment and therefore not visible to the subject. As Eurasian jays suppress caching in the presence of a dominant conspecific [22], the conspecific was socially subordinate to the subject when possible. We used four birds (two males, two females) with no prior experience of caching in either substrate as conspecifics. Only one of these (Purchas, table 1) was later tested as a subject.
Table 1.
The number of items cached by subjects in each location within each condition in experiment 1. In the alone condition, the data are shown for trials 1 and 2. Caches made in out-of-tray locations were considered to be quiet.
| condition: | alone |
seen and heard |
heard but not seen |
|||||||
|---|---|---|---|---|---|---|---|---|---|---|
| subject | substrate: | sand | gravel | out | sand | gravel | out | sand | gravel | out |
| Wilson | 5, 1 | 2, 0 | 7, 1 | 3 | 1 | 8 | 6 | 1 | 7 | |
| Ohuruogu | 1, 3 | 0, 0 | 19, 11 | 1 | 0 | 12 | 4 | 0 | 10 | |
| Pendleton | 2, 0 | 7, 3 | 6, 5 | 0 | 2 | 6 | 3 | 2 | 10 | |
| Hoy | 1, 21 | 0, 0 | 0, 2 | 11 | 2 | 1 | 25 | 0 | 0 | |
| Hunter | 4, 3 | 1, 0 | 0, 1 | 3 | 0 | 0 | 0 | 0 | 3 | |
| Romero | 15, 5 | 2, 2 | 6, 8 | 11 | 0 | 3 | 4 | 0 | 3 | |
| Wiggins | 6, 5 | 4, 0 | 1, 2 | 2 | 0 | 0 | 4 | 0 | 0 | |
| Purchas | 5, 5 | 10, 8 | 8, 10 | 5 | 1 | 5 | 4 | 1 | 10 | |
Initially, we gave subjects ‘alone’ trials to establish a baseline for their caching preferences. After an ‘alone’ trial, individuals were allowed to recover caches for 15 min. Recovery took place either 90 min after the end of a trial (while subjects were still deprived), or the following morning after approximately 90 min of deprivation. Subjects received three, four or five ‘alone’ trials, depending on their motivation to cache in their initial trials. Following the ‘alone’ trials, we gave four subjects a ‘seen and heard’ trial and four subjects a ‘heard but not seen’ trial. We then tested subjects in the condition that they had not yet experienced. To rule out the possibility of birds being either punished or rewarded for caching in a particular location or substrate [12], in the ‘seen and heard’ and the ‘heard but not seen’ conditions, subjects received only one trial and were not given the opportunity to recover their caches.
Data were analysed using Genstat v. 13.1, whereas all non-parametric statistics were calculated by hand [23]. Mean data from the last two trials of the ‘alone’ condition were analysed, as five birds only cached in their last two trials of this condition. As the assumptions of sphericity and normally distributed residuals were met, a repeated-measures analysis of variance (ANOVA) was used to analyse the effect of condition on the total number of items cached. As these assumptions were violated in the analysis of the effect of condition on the proportion of caches made in the ‘noisy’ substrate, a Friedman ANOVA was used. Within each condition, the proportion cached in the noisy tray was compared with the proportion cached in the quiet tray using Wilcoxon signed ranks tests. Following Stulp et al. [12] and to reflect the relative expenditure of caching effort, we calculated the amount cached in each substrate as a proportion out of all caches made (noisy tray + quiet tray + out-of-tray caches), with caches made in out-of-tray caches locations considered to be quiet. Wilcoxon signed ranks tests were used for pair-wise planned contrasts comparing the proportion of caches made in the noisy substrate between the ‘alone’ and the ‘heard but not seen’ condition, and between the ‘alone’ and the ‘seen and heard’ condition. Alpha was 0.05 for all tests. Wilcoxon test significance was evaluated using a non-directional table of critical values in all tests except the planned contrasts, for which one-tailed tables were used.
(c). Results and discussion
The total amount that jays cached did not differ between the conditions (repeated measures ANOVA: F2,14 = 1.247, p = 0.32; individual data in table 1). Eurasian jays had a clear preference for caching in the quiet substrate in both conditions where a conspecific was present (‘seen and heard’: n = 8, t = 3.00, p < 0.05; ‘heard but not seen’: n = 8, (ties = 1), t = 0.00, p < 0.02; figure 2), while this was a trend in the ‘alone’ condition (n = 8, t = 4.00, 0.1 > p > 0.05). Despite this general preference for caching in the quiet substrate, jays nevertheless altered the proportion of caches that were made in the noisy substrate across conditions (Friedman ANOVA: n = 8, 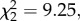 p = 0.01; figure 2).
p = 0.01; figure 2).
Figure 2.
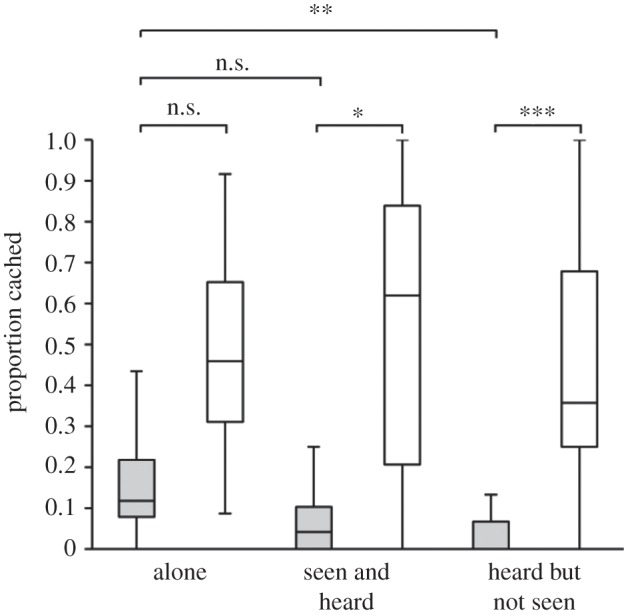
The proportion of caches made by subjects in the ‘noisy’ tray (grey) and in the ‘quiet’ tray (white) within each condition (calculated as a proportion of noisy+quiet+out-of-tray caches; n.s.: p > 0.05, *p < 0.05, **p < 0.025, ***p < 0.02).
In support of the hypothesis that jays conceal auditory information as a cache-protection strategy, relative to when caching in the ‘alone’ condition, subjects reduced the proportion cached in the noisy substrate in the ‘heard but not seen’ condition (n = 8 (ties = 2), t = 0.00, p < 0.025; figure 2). By contrast, no difference was found between the ‘alone’ and the ‘seen and heard’ conditions (n = 8 (ties = 1), t = 5.00, p > 0.05). Four jays cached nothing in the noisy substrate in both the ‘seen and heard’ and the ‘heard but not seen’ conditions; so the difference between these conditions could not be tested using Wilcoxon signed ranks. Nevertheless, the four subjects that cached in the noisy substrate in the ‘seen and heard’ condition all made a smaller proportion of caches in the noisy substrate in the ‘heard but not seen’ condition. Taken together, these results potentially indicate that subjects attended to the specific state, rather than the general presence of a competitor. The reduction in the proportion of caches made in the noisy substrate when a competitor could hear but could not see them suggests that Eurasian jays reduce the amount of auditory information available to competitors as a cache-protection strategy.
A coevolutionary arms race between caching and pilfering behaviour predicts that as cachers use strategies to conceal caching from potential thieves, pilferers should adopt counter-tactics that maximize the likelihood that they will witness caching [6,14]. Accordingly, ravens observing caching adjust their position to gain a better view, while maintaining their distance from the cacher [6]. Experiment 1 provides evidence that jays reduce the transfer of auditory information to competitors as a cache-protection strategy. It is therefore possible that when observing a conspecific cache, jays will also avoid providing any auditory cues that may reveal their presence to the cacher. We tested this hypothesis in experiment 2.
3. Experiment 2
(a). Hypothesis and predictions
We gave jays the opportunity to look into an adjacent compartment through two peep-holes in an otherwise opaque barrier. At one peep-hole was a noisy perch with bells attached to it, while at the other was quiet perch with non-functional bells. The jays' perching, looking and vocalizing behaviours were tested when jays were (i) alone, (ii) observing a caching conspecific, (iii) observing a non-caching conspecific and (iv) observing a conspecific pilfering caches that they had made. A fifth condition (v) tested perching and vocalizing behaviour when there was no opaque barrier, such that birds were visible to the cacher and had an unrestricted view of caching. If pilfering jays reduce the amount of noise they make to avoid alerting a cacher to their presence, then compared with conditions (i, iii–v), individuals should prefer the quiet perch and (when applicable) quiet peep-hole and suppress vocalizations when watching caching from behind the opaque barrier (ii).
(b). Methods
General housing and husbandry was the same as for experiment 1. We tested subjects (n = 6; three males, three females) between 14 January 2011–10 March 2011 and 19 September 2011–21 March 2012. Compartments C, D and E (figure 1a) were modified for this experiment (figure 3). Compartment D contained a single, fixed perch, whereas compartments C and E each contained two straight tree branches with wire loops at either end that were hooked over nails, creating perches that swung freely back and forth. At both ends of each swinging perch, a cluster of three small red bells were attached with cable ties. Compartment C and E each contained one ‘noisy’ perch and one ‘quiet’ perch. The ‘noisy’ perch had bells that had been coated in glue but still rang, whereas the ‘quiet’ perch had bells that had been glued together and did not ring when moved. Two peep-holes (3 cm diameter, spaced 50 cm apart) were cut into the opaque wall separating compartments D and E, and each was aligned with one of the perches in E.
Figure 3.
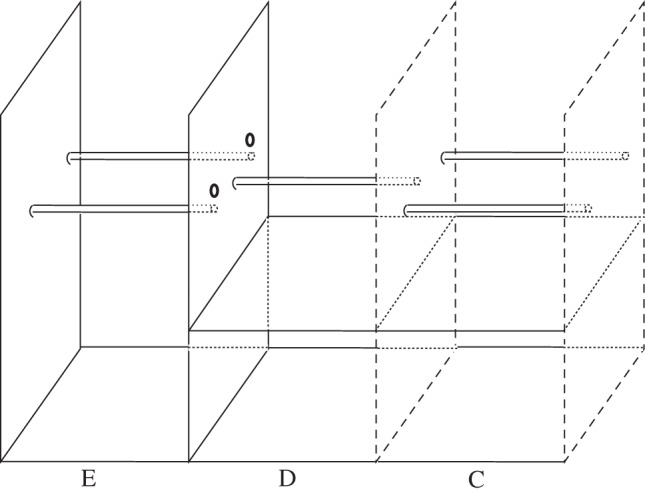
The location of perches, peep-holes (bold circles) and the wooden platforms in compartments C and D. Long-dashed lines represent wire mesh barriers and solid lines opaque structures.
Before testing, subjects experienced using each perch and peep-hole in two 15 min trials. In each trial, either the noisy or the quiet perch was present in compartment E and a conspecific, caching tray (as used in experiment 1, containing sand) and food bowl containing 20 wax moth (Achroia grisella) larvae and 20 peanuts were present in compartment D. The order in which each perch type was experienced, and the location of the noisy and quiet perches was randomized between individuals. In a subject's remaining trials, the noisy and quiet perch remained in the same position as during the forced choice trials.
Subjects were tested for 15 min in each of the five conditions. In the (i) ‘alone’ condition, subjects were tested in compartment E with no caching trays or conspecific present in D or C. In the (ii) ‘watching caching’ condition, subjects were tested in compartment E and a dominant conspecific (the ‘cacher’), caching tray and food bowl were present in D. In the (iii) ‘social control’ condition, the subject was tested in E, a caching tray and food bowl were in D and the cacher was in C (where the swinging perches had been replaced by a single, fixed perch), such that it could be seen by the subject but could not cache or eat during the trial. In the (iv) ‘watching pilfering’ condition, we gave subjects a caching tray and food bowl in compartment D and allowed them to cache for 15 min in view of a conspecific in compartment C. We then removed the food bowl from compartment D, moved the subject into compartment E and moved the conspecific into compartment D to act as a pilferer. Finally, in the (v) ‘open wall’ condition the cacher, caching tray and food bowl were present in compartment D while subjects were tested in C, such that their view was un-obscured and they were in full view of the cacher. Between subjects we randomized, the order of conditions (i), (ii), (iii) and (v). The experience of caching in compartment D could have had carry-over effects on the jays' motivation to observe compartment D in subsequent trials, which may have differed greatly from the trials in which subjects did not cache in D [24]. All subjects therefore underwent condition (iv) last.
Three individuals (two males, one female) acted as cachers. One male and the female acted as cachers after being tested as subjects. In all conditions except the ‘alone’ condition, R.C.S., with whom all birds were familiar and accustomed to caching in front of, stood at the far end of the corridor (figure 1a) and quietly dictated the locations of any caches made during the trial using a Nokia C3-00 phone with a voice recorder application. The subject and the cacher were filmed, using Sony DCR-PC110E digital video cameras.
Behaviour was coded from the videos using The Observer XT (v. 9.0). Landing and taking off from perches and peep-hole use was recorded using frame-by-frame analysis (30 frames s−1). A subject was deemed to be looking through a peep-hole when they were within 10 cm of it and their head was either aligned with it, inclined towards it or they were oriented facing towards it. Videos were played at normal speed to count to number of vocalizations. For each subject, the video of a randomly chosen trial was analysed by an independent observer (E.L.) who was unfamiliar with the experimental conditions and subject identity. Inter-observer agreement was reasonable for each behavioural category (perching: 87% agreement, Cohen's κ = 0.73; looking: 86% agreement, Cohen's κ = 0.75; vocalization: 90% agreement).
As for experiment 1, data were analysed using Genstat (v. 13.1), and exact non-parametric statistics were calculated by hand [23]. For the analysis of total number of looks, the assumptions of repeated-measures ANOVA were met and so this was used to test for the effect of condition on the total number of times that subjects looked into the adjacent compartment. For the remaining analyses, the repeated-measures ANOVA assumptions were violated and so Friedman ANOVAs were used to analyse the effect of condition on perching, looking and vocalizing behaviour. Exact Wilcoxon signed ranks for matched pairs tests were used for the planned contrasts stated in §3a for experiment 2. The significance of these predictions was evaluated using one-tailed tables. Alpha was set at 0.05.
(c). Results and discussion
Throughout the experiment, jays were consistent in their use of the peep-holes. Accordingly, there was no effect of condition found in the total number of times subjects looked into the adjacent compartment (F3,15 = 2.004, p = 0.16) or the total amount of time subjects spent looking (n = 6, 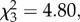 p = 0.19). Moreover, when observing caching from behind an opaque barrier, Eurasian jays did not attempt to conceal their presence from the cacher by preferentially using the quiet perch and peep-hole and their preference for the quiet perch and peep-hole did not differ across conditions (table 2; data available in electronic supplementary material).
p = 0.19). Moreover, when observing caching from behind an opaque barrier, Eurasian jays did not attempt to conceal their presence from the cacher by preferentially using the quiet perch and peep-hole and their preference for the quiet perch and peep-hole did not differ across conditions (table 2; data available in electronic supplementary material).
Table 2.
Medians and inter-quartile ranges for the proportion of time spent perching and proportion of hops using the quiet perch and for the proportion of time spent looking and proportion of looks using the quiet peep hole in each condition in experiment 2.
| condition |
Friedman ANOVA |
||||||||
|---|---|---|---|---|---|---|---|---|---|
| behaviour | alone | watching caching | social control | watching pilfering | open wall | n | χ2 | d.f. | p |
| perching time | 0.75 | 0.64 | 0.61 | 0.66 | 0.51 | 6 | 2.27 | 4 | 0.69 |
| 0.64–0.80 | 0.59–0.71 | 0.32–0.70 | 0.61–0.87 | 0.32–0.82 | |||||
| no. of hops | 0.65 | 0.56 | 0.61 | 0.67 | 0.50 | 6 | 5.82 | 4 | 0.21 |
| 0.51–0.74 | 0.51–0.69 | 0.55–0.67 | 0.60–0.67 | 0.40–0.63 | |||||
| looking time | 0.69 | 0.76 | 0.64 | 0.73 | — | 6 | 3.96 | 3 | 0.26 |
| 0.46–0.80 | 0.59–0.85 | 0.48–0.93 | 0.61–0.84 | — | |||||
| no. of looks | 0.66 | 0.60 | 0.63 | 0.67 | — | 6 | 0.66 | 3 | 0.88 |
| 0.46–0.67 | 0.51–0.73 | 0.52–0.92 | 0.61–0.67 | — | |||||
Despite the jays' potential indifference towards the amount of noise created by their movements, there was evidence that subjects suppressed vocalizations when watching caching. The number of vocalizations that jays made differed between the five conditions (n = 6, 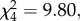 p = 0.044; figure 4). In the ‘watching caching’ condition, subjects vocalized less compared with when they were alone (n = 6, t = 0.00, p < 0.025) and when they were watching a non-caching conspecific (n = 6, t = 2.00, p < 0.05), while there was no difference in the vocalization rate whether the jays were fully visible to a caching conspecific or not (n = 6, t = 3.00, p > 0.05; figure 4). Potentially, the vocalizing rate is a function of attention, and so the jay's suppression of vocalizations when watching caching may have been because birds were more attentive in these conditions. However, the jays' overall looking behaviour did not differ across conditions. Moreover, compared with when using the peep-holes to observe caching, jays vocalized more frequently when they were watching their own caches being pilfered (n = 6, t = 2.00, p < 0.05; figure 4), potentially to intimidate or deter the pilferer. Watching caches being pilfered likely engaged the subjects' attention, further making it unlikely that vocalization suppression when watching caching was only due to the jay's attentiveness differing between conditions. Instead, these results suggest that when observing caching, Eurasian jays may avoid disturbing the cacher by suppressing vocalizations.
p = 0.044; figure 4). In the ‘watching caching’ condition, subjects vocalized less compared with when they were alone (n = 6, t = 0.00, p < 0.025) and when they were watching a non-caching conspecific (n = 6, t = 2.00, p < 0.05), while there was no difference in the vocalization rate whether the jays were fully visible to a caching conspecific or not (n = 6, t = 3.00, p > 0.05; figure 4). Potentially, the vocalizing rate is a function of attention, and so the jay's suppression of vocalizations when watching caching may have been because birds were more attentive in these conditions. However, the jays' overall looking behaviour did not differ across conditions. Moreover, compared with when using the peep-holes to observe caching, jays vocalized more frequently when they were watching their own caches being pilfered (n = 6, t = 2.00, p < 0.05; figure 4), potentially to intimidate or deter the pilferer. Watching caches being pilfered likely engaged the subjects' attention, further making it unlikely that vocalization suppression when watching caching was only due to the jay's attentiveness differing between conditions. Instead, these results suggest that when observing caching, Eurasian jays may avoid disturbing the cacher by suppressing vocalizations.
Figure 4.
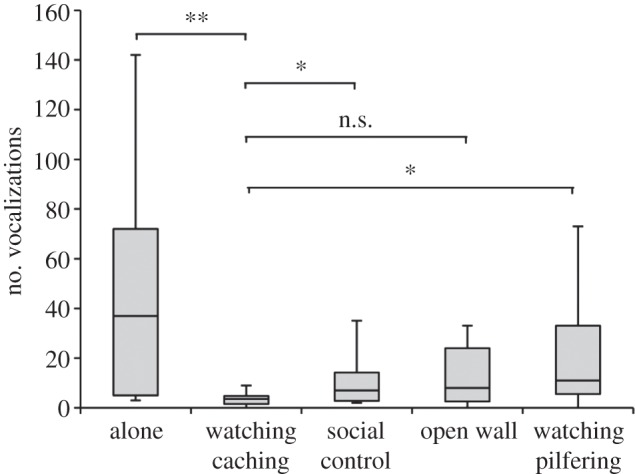
The number of vocalizations made by subjects in each condition of experiment 2 (n.s.: p > 0.05, *p < 0.05, **p < 0.025).
4. General discussion
This is the first study to experimentally evaluate both caching (experiment 1) and pilfering (experiment 2) strategies in the auditory domain. Before discussing the implications of our results, we note that the small sample sizes and large amount of within and between subject variation in experiments 1 and 2 did not allow us to take into account the order in which subjects underwent test conditions. This limitation potentially inflated type II error in experiment 1 and may have biased results in experiment 2 [24]. Unfortunately, we were unable to test additional subjects to allow the effect of trial order to be included in the analyses. Nonetheless, our conservative, non-parametric analyses revealed an effect of condition on the jay's behaviour in both experiments. As cachers, Eurasian jays were sensitive to the noise that they produced when caching, favouring quiet substrates in general. Despite this general preference, jays responded to the presence and perceptual state of a competitor. Compared with when alone, jays reduced the proportion of items cached in the noisy substrate when in the presence of a conspecific that could hear but could not see them, but did not do so when with a competitor that could see and hear caching. By contrast, as potential pilferers observing a caching conspecific, jays did not attend to the amount of noise created by their movements, but did suppress vocalizations in the presence of a cacher.
In contrast to previously tested western scrub-jays [12], Eurasian jays did not prefer to cache in the noisy substrate when visible to a conspecific. This contrast in the amount of noise produced by cachers when in the presence of a competitor that can both see and hear caching could be influenced by differences in the ecology of these two corvid species. In the wild, western scrub-jays are relatively conspicuous, whereas Eurasian jays are secretive and frequently difficult to detect even when in close proximity [21]. This secretive disposition may result in Eurasian jays being averse to drawing attention to themselves whenever a competitor (which for wild jays may be either conspecific or heterospecific [21]) is present, resulting in a general preference for quiet substrates. Testing more species will elucidate the extent and distribution of cache-protection and pilfering strategies and how these interact with the ecology of species to create such behavioural differences.
There is evidence that corvids can attribute visual perception to others. Western scrub-jays remember the specific identity of potential pilferers [9,10], whereas pilfering ravens remember what a conspecific has seen when competing over caches they have seen a human experimenter make [25]. In experiment 2, the subjects' response may have used a rule such as ‘suppress vocalization in the presence of a cacher’ that was learned through experience while interacting with conspecifics in the aviary. By contrast, in experiment 1, the jays had no prior experience of the acoustic properties of the caching substrates. Although the out-of-tray locations were not novel to the jays, five of the eight subjects only began to cache in out-of-tray locations after at least one alone trial, suggesting that the majority of subjects were unfamiliar with caching in the testing compartments. It therefore remains possible that the subjects' response in experiment 1 was not based on learned rules about where to cache in the presence of competitors with differing perceptual states. Instead, the reduction in the relative amount cached in noisy substrates in response to the specific state of a competitor may have been a cache-protection tactic that was based on the jay's understanding that their competitors can perceive auditory information.
Together, our experiments reveal that Eurasian jays attempt to limit the transfer of some types of auditory information to competitors both as cachers and as pilferers, potentially using cognitive strategies for the former and learned rules for the latter. A coevolutionary arms race, therefore, may shape cache-protection and pilfering tactics in the auditory domain in Eurasian jays. Ultimately, such an arms race could result in the emergence of the ability to attribute knowledge gained through auditory perception to other individuals [7]. Investigating the extent of the jay's use of auditory information and determining the distribution of similar abilities in other corvids therefore remain exciting avenues for future research.
Acknowledgements
The work was supported by grants to N.S.C. from the Biotechnology and Biological Sciences Research Council, the Royal Society and the University of Cambridge. R.C.S. was funded by a Cambridge-Rutherford Memorial Scholarship from the Rutherford Foundation of the Royal Society of New Zealand and the Cambridge Commonwealth Trust. The experiments were conducted under the UK Home Office project licences no. PPL 80/1975 and PPL 80/2519. We thank Edward Legg for assistance with inter-observer reliability and comments on the manuscript, three anonymous reviewers for their comments, Ljerka Ostojić and Andrew Bateman for assistance with statistical analyses and discussion, and Ivan Vakrilov and Charmaine Donovan for avian husbandry.
References
- 1.Vander Wall SB, Jenkins SH. 2003. Reciprocal pilferage and the evolution of food-hoarding behavior. Behav. Ecol. 14, 656–667 10.1093/beheco/arg064 (doi:10.1093/beheco/arg064) [DOI] [Google Scholar]
- 2.Scheid C, Bugnyar T. 2008. Short-term observational spatial memory in jackdaws (Corvus monedula) and ravens (Corvus corax). Anim. Cogn. 11, 691–698 10.1007/s10071-008-0160-5 (doi:10.1007/s10071-008-0160-5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Watanabe S, Clayton NS. 2007. Observational visuospatial encoding of the cache locations of others by western scrub-jays (Aphelocoma californica). J. Ethol. 25, 271–279 10.1007/s10164-006-0023-y (doi:10.1007/s10164-006-0023-y) [DOI] [Google Scholar]
- 4.Bednekoff PA, Balda RP. 1996. Observational spatial memory in Clark's nutcrackers and Mexican jays. Anim. Behav. 52, 833–839 10.1006/anbe.1996.0228 (doi:10.1006/anbe.1996.0228) [DOI] [PubMed] [Google Scholar]
- 5.Bednekoff PA, Balda RP. 1996. Social caching and observational spatial memory in pinyon jays. Behaviour 133, 807–826 10.1163/156853996X00251 (doi:10.1163/156853996X00251) [DOI] [Google Scholar]
- 6.Bugnyar T, Kotrschal K. 2002. Observational learning and the raiding of food caches in ravens, Corvus corax: is it ‘tactical’ deception? Anim. Behav. 64, 185–195 10.1006/anbe.2002.3056 (doi:10.1006/anbe.2002.3056) [DOI] [Google Scholar]
- 7.Grodzinski U, Clayton NS. 2010. Problems faced by food-caching corvids and the evolution of cognitive solutions. Phil. Trans. R. Soc. B 365, 977–987 10.1098/rstb.2009.0210 (doi:10.1098/rstb.2009.0210) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dally JM, Emery NJ, Clayton NS. 2005. Cache protection strategies by western scrub-jays, Aphelocoma californica: implications for social cognition. Anim. Behav. 70, 1251–1263 10.1016/j.anbehav.2005.02.009 (doi:10.1016/j.anbehav.2005.02.009) [DOI] [Google Scholar]
- 9.Dally JM, Emery NJ, Clayton NS. 2006. Food-caching western scrub-jays keep track of who was watching when. Science 312, 1662–1665 10.1126/science.1126539 (doi:10.1126/science.1126539) [DOI] [PubMed] [Google Scholar]
- 10.Emery NJ, Clayton NS. 2001. Effects of experience and social context on prospective caching strategies by scrub jays. Nature 414, 443–446 10.1038/35106560 (doi:10.1038/35106560) [DOI] [PubMed] [Google Scholar]
- 11.Dally JM, Emery NJ, Clayton NS. 2004. Cache protection strategies by western scrub-jays (Aphelocoma californica): hiding food in the shade. Proc. R. Soc. Lond. B 271, S387–S390 10.1098/rsbl.2004.0190 (doi:10.1098/rsbl.2004.0190) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Stulp G, Emery NJ, Verhulst S, Clayton NS. 2009. Western scrub-jays conceal auditory information when competitors can hear but cannot see. Biol. Lett. 5, 583–585 10.1098/rsbl.2009.0330 (doi:10.1098/rsbl.2009.0330) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Clayton NS, Dally JM, Emery NJ. 2007. Social cognition by food-caching corvids. The western scrub-jay as a natural psychologist. Phil. Trans. R. Soc. B 362, 507–522 10.1098/rstb.2006.1992 (doi:10.1098/rstb.2006.1992) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dally JM, Clayton NS, Emery NJ. 2006. The behaviour and evolution of cache protection and pilferage. Anim. Behav. 72, 13–23 10.1016/j.anbehav.2005.08.020 (doi:10.1016/j.anbehav.2005.08.020) [DOI] [Google Scholar]
- 15.de Kort SR, Clayton NS. 2006. An evolutionary perspective on caching by corvids. Proc. R. Soc. B 273, 417–423 10.1098/rspb.2005.3350 (doi:10.1098/rspb.2005.3350) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Braun A, Bugnyar T. 2012. Social bonds and rank acquisition in raven nonbreeder aggregations. Anim. Behav. 1–9 10.1016/j.anbehav.2012.09.024 (doi:10.1016/j.anbehav.2012.09.024) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bugnyar T, Kotrschal K. 2002. Scrounging tactics in free-ranging ravens, Corvus corax. Ethology 108, 993–1009 10.1046/j.1439-0310.2002.00832.x (doi:10.1046/j.1439-0310.2002.00832.x) [DOI] [Google Scholar]
- 18.Curry RL, Peterson AT, Langen TA. 2002. Western scrub-jay. In The birds of North America (eds Poole A, Gill F.), pp. 1–36 Philadelphia, PA: The Birds of North America, Inc [Google Scholar]
- 19.Goodwin D. 1986. Crows of the world, 2nd edn Suffolk, UK: British Museum (Natural History) [Google Scholar]
- 20.Andrén H. 1992. Corvid density and nest predation in relation to forest fragmentation: a landscape perspective. Ecology 73, 794–804 10.2307/1940158 (doi:10.2307/1940158) [DOI] [Google Scholar]
- 21.Bossema I. 1979. Jays and oaks: an eco-ethological study of a symbiosis. Behaviour 70, 1–117 10.1163/156853979X00016 (doi:10.1163/156853979X00016) [DOI] [Google Scholar]
- 22.Shaw RC, Clayton NS. 2012. Eurasian jays, Garrulus glandarius, flexibly switch caching and pilfering tactics in response to social context. Anim. Behav. 84, 1191–1200 10.1016/j.anbehav.2012.08.023 (doi:10.1016/j.anbehav.2012.08.023) [DOI] [Google Scholar]
- 23.Mundry R, Fischer J. 1998. Use of statistical programs for nonparametric tests of small samples often leads to incorrect P-values: examples from Animal Behaviour. Anim. Behav. 56, 256–259 10.1006/anbe.1998.0756 (doi:10.1006/anbe.1998.0756) [DOI] [PubMed] [Google Scholar]
- 24.Díaz-Uriarte R. 2002. Incorrect analysis of crossover trials in animal behaviour research. Anim. Behav. 63, 815–822 10.1006/anbe.2001.1950 (doi:10.1006/anbe.2001.1950) [DOI] [Google Scholar]
- 25.Bugnyar T. 2011. Knower–guesser differentiation in ravens: others’ viewpoints matter. Proc. R. Soc. B 278, 634–640 10.1098/rspb.2010.1514 (doi:10.1098/rspb.2010.1514) [DOI] [PMC free article] [PubMed] [Google Scholar]