

Abstract
Anthropogenic climate change is predicted to be a major cause of species extinctions in the next 100 years. But what will actually cause these extinctions? For example, will it be limited physiological tolerance to high temperatures, changing biotic interactions or other factors? Here, we systematically review the proximate causes of climate-change related extinctions and their empirical support. We find 136 case studies of climatic impacts that are potentially relevant to this topic. However, only seven identified proximate causes of demonstrated local extinctions due to anthropogenic climate change. Among these seven studies, the proximate causes vary widely. Surprisingly, none show a straightforward relationship between local extinction and limited tolerances to high temperature. Instead, many studies implicate species interactions as an important proximate cause, especially decreases in food availability. We find very similar patterns in studies showing decreases in abundance associated with climate change, and in those studies showing impacts of climatic oscillations. Collectively, these results highlight our disturbingly limited knowledge of this crucial issue but also support the idea that changing species interactions are an important cause of documented population declines and extinctions related to climate change. Finally, we briefly outline general research strategies for identifying these proximate causes in future studies.
Keywords: climate change, extinction, physiological tolerances, species interactions
1. Introduction
Anthropogenic climate change is recognized as a major threat to global biodiversity, one that may lead to the extinction of thousands of species over the next 100 years [1–7]. Climate change is an especially pernicious threat, as it may be difficult to protect species from its effects, even within reserves [8,9]. Furthermore, climate change may have important interactions with other anthropogenic impacts (e.g. habitat loss [2,6]). Given this, understanding the responses of species to modern climate change is one of the most pressing issues facing biologists today.
But what do we actually know about how climate change causes extinction? It might seem that limited physiological tolerances to high temperatures should be the major factor that causes climate change to threaten the persistence of populations and species, and many studies have justifiably focused on these tolerances [10–13]. However, there may be many other proximate causes of extinction, even when anthropogenic climate change is the ultimate cause. These proximate factors include negative impacts of heat-avoidance behaviour [14], the climate-related loss of host and pollinator species [15,16] and positive impacts of climate change on pathogens and competitors [17,18], among others. The relative importance of these factors is unclear and has not, to our knowledge, previously been reviewed, despite increasing interest in mechanisms underlying the impacts of climate change [19].
Identifying these proximate causes may be critical for many reasons. For example, different proximate factors may call for different conservation strategies to ameliorate their effects [20]. These different proximate factors may also influence the accuracy with which the impacts of climate change are predicted and may drive populations to extinction at different rates.
In this paper, we address three topics related to how anthropogenic climate change causes extinction. First, we briefly review and categorize the many proposed factors that potentially lead to extinction from climate change. Second, we argue that there is already abundant evidence for current local extinctions as a result of climate change, based on the widespread pattern of range contractions at the warm edges of species' ranges (low latitude and low elevation). Third, and most importantly, we perform to the best of our knowledge, the first large-scale review of empirical studies that have addressed the proximate causes of local extinctions related to climate change. This review reveals some unexpected results. We find that despite intensive research on the impacts of climate change, only a handful of studies have demonstrated a proximate cause of local extinctions. Further, among those studies that have identified a proximate cause, very few implicate limited physiological tolerance to high temperatures as the main, direct cause. Instead, a diverse set of factors are supported, with species interactions being particularly important. Finally, we outline some of the research approaches that can be used to examine the proximate factors causing extinction from climate change.
2. Proximate factors causing extinction from climate change
We briefly review and categorize the diverse proximate factors that may cause extinctions due to climate change. We organize these factors by distinguishing between abiotic and biotic factors (following the literature on species range limits [21]). However, all factors are ultimately related to abiotic climate change.
We make several caveats about this classification. First, we emphasize broad categories of factors, so some specific factors may not be included. Second, some factors are presently hypothetical and have not yet been demonstrated as causes of extinction. Third, we recognize that these factors are not mutually exclusive and may act synergistically to drive extinction. They may also interact with other, non-climatic factors (e.g. habitat modification [2,6]) and many different ecological and demographic factors may come into play as populations approach extinction [22]. Finally, we do not address factors that impede climate-induced dispersal.
(a). Abiotic factors
(i). Temperature (physiological tolerances)
Many effects of anthropogenic climate change follow from an increase in temperature. The most obvious proximate factor causing extinction is temperatures that exceed the physiological tolerance of the species [10,12]. This factor may be most important in sessile organisms and those with limited thermoregulatory ability, and in regions and time scales in which temperature increase is greatest.
The impacts of temperature may also be more indirect, but still related to physiological tolerances. For example, in spiny lizards (Sceloporus), local extinctions seem to occur because higher temperatures restrict surface activity during the spring breeding season to a daily time window that is overly short [23]. Similarly, increased air temperatures may both decrease activity time and increase energy maintenance costs, leading organisms to die from starvation rather than from overheating [14]. In aquatic organisms, increased water temperatures may lead to increased metabolic demand for oxygen while reducing the oxygen content of the water [24]. Variability in temperature may also be an important proximate cause of extinction [25], including both extreme events and large differences over the course of a year. In temperate and polar latitudes, a mismatch between photoperiod cues and temperature may be important, with fixed photoperiod responses leading to activity patterns that are inappropriate for the changed climate [26]. Here, both low and high temperatures could increase mortality rates and lead to population extinction.
(ii). Precipitation (physiological tolerances)
Anthropogenic changes are also modifying precipitation patterns [27], and these changes may drive extinction in a variety of ways. For example, decreasing precipitation may lead directly to water stress, death and local extinction for terrestrial species [28], and loss of habitat for freshwater species or life stages [29,30]. There may also be synergistic effects between heat and drought stress (e.g. in trees [31]). Changing precipitation may be more important to some species than changing temperature, sometimes leading to range shifts in the direction opposite to those predicted by rising temperatures [32].
(iii). Other abiotic factors
Other abiotic, non-climatic factors may drive extinctions that are ultimately caused by climate change. For example, climate change can increase fire frequency, and these fires may be proximate causes of extinction (e.g. in South African plants [33]). Similarly, increases in temperature lead to melting icecaps and rising sea levels [27], which may eliminate coastal habitats and modify the salinity of freshwater habitats [34].
(b). Biotic factors
The biotic factors that are the proximate causes of extinction from climate change can be placed in three general categories.
(i). Negative impacts on beneficial species
Climate change may cause local extinction of a given species by causing declines in a species upon which it depends. These may include prey for predators [35], hosts for parasites and specialized herbivores [16], species that create necessary microhabitats [36] and species that are essential for reproduction (e.g. pollinators [15]).
(ii). Positive impacts on harmful species
Alternately, climate change may cause extinction through positive effects on species that have negative interactions with a focal species, including competitors [37,38], predators [39,40] and pathogens [41–43]. Warming temperatures can also benefit introduced species, exacerbating their negative effects on native flora and fauna [44].
(iii). Temporal mismatch between interacting species
Climate change may also create incongruence between the activity times of interacting species [45]. These phenological mismatches may occur when interacting species respond to different environmental cues (e.g. temperature versus photoperiod for winter emergence) that are not congruently influenced by climate change [46]. We consider this category to be distinct from the other two because the differences in activity times are not necessarily negative or positive impacts on the species that are interacting with the focal species.
3. Are there current extinctions due to climate change?
Our goal is to understand which proximate factors cause extinctions due to climate change. However, we first need to establish that such extinctions are presently occurring. Few global species extinctions are thought to have been caused by climate change. For example, only 20 of 864 species extinctions are considered by the International Union for Conservation of Nature (IUCN) [47] to potentially be the result of climate change, either wholly or in part (using the same search criteria as a recent review [9]), and the evidence linking them to climate change is typically very tenuous (see the electronic supplementary material, table S1). However, there is abundant evidence for local extinctions from contractions at the warm edges of species' ranges. A pattern of range shifts (generally polewards and upwards) has been documented in hundreds of species of plants and animals [48,49], and is one of the strongest signals of biotic change from global warming. These shifts result from two processes: cold-edge expansion and warm-edge contraction (see the electronic supplementary material, figure S1). Much has been written about cold-edge expansions [21,50], and these may be more common than warm-edge contractions [51]. Nevertheless, many warm-edge contractions have been documented [52–58], including large-scale review studies spanning hundreds of species [48,59]. These warm-edge populations are a logical place to look for the causes of climate-related extinctions, especially because they may already be at the limits of their climatic tolerances [60]. Importantly, this pattern of warm-edge contraction provides evidence that many local extinctions have already occurred as a result of climate change.
We generally assume that the proximate factors causing local extinction from climate change are associated with the death of individuals. However, others factors may be involved as well. These include emigration of individuals into adjacent localities, declines in recruitment, or a combination of these and other factors. The question of whether climate-related local extinctions occur through death, dispersal or other processes has received little attention (but see [61,62]), and represents another important but poorly explored area in climate-change research.
4. What causes extinction due to climate change? current evidence
Given that there are many different potential causes of extinction as a result of climate change, and given that many populations have already gone extinct (as evidenced by warm-edge range contractions), what proximate causes of climate-related extinction have actually been documented? We conducted a systematic review of the literature to address this question.
(a). Causes of extinction: methods
We conducted three searches in the ISI Web of Science database, using the following keywords: (i) ((‘locally extinct’ OR ‘local extinction’ OR ‘extinc*’) AND (caus*) AND (‘climate change’ OR ‘global warming’)); (ii) ((‘locally extinct’ OR ‘local extinction’) AND (‘climate change’ OR ‘global warming’)); and (iii) ((‘extinc*’ OR ‘extirpat*’) AND (‘climate change’ OR ‘global warming’ OR ‘changing climate’ OR ‘global change’)). The first two were conducted on 7 December 2011 and the third on 4 February 2012. Each search identified a partially overlapping set of studies (687 unique studies overall). We then reduced this to 136 studies which suggested that climate change is associated with local extinctions or declines (see the electronic supplementary material, appendix S1).
Among these 136 studies, we then identified those that reported an association between local extinction and climatic variables and that also identified a specific proximate cause for these extinctions (see the electronic supplementary material, appendix S1). The evidence linking these proximate causes to anthropogenic climate change varied considerably, but included studies integrating experimental and correlative results [23,63], and those that also accounted for factors unrelated to climate change [64]. Although we did not perform a separate, comprehensive search for all studies of climate-related declines, we also include studies of population declines that were connected to potential local extinctions as a second category of studies. Studies of declines should also be informative, given that the factors causing population declines may ultimately lead to extinctions [65]. All studies reported declines in abundance but some also considered declines in other parameters (e.g. fecundity). We also included studies of impacts from natural oscillations (such as the El Niño-Southern Oscillation, ENSO) as a third category of results.
(b). Causes of extinction: results
(i). Proximate causes of local extinctions
Of 136 studies focusing on local extinctions associated with climate change (see the electronic supplementary material, appendix S1), only seven identified the proximate causes of these extinctions (table 1 and figure 1a). Surprisingly, none of the seven studies shows a straightforward relationship between local extinction and limited tolerances to high temperature. For example, for the two studies that relate extinctions most directly to changing temperatures, the proximate factor is related either to how temperature limits surface activity time during the breeding season [23] or to a complex relationship between extreme temperatures (both cold and hot), precipitation and physiology [25,63]. Most studies (four of seven) implicate species interactions as the proximate cause, especially decreases in food availability [35,64,66]. Many authors have predicted that altered species interactions may be an important cause of extinction resulting from climate change (e.g. [67,68]), and our results empirically support the importance of these interactions (relative to other factors) among documented cases of local extinction.
Table 1.
Studies documenting the proximate causes of local extinction due to anthropogenic climatic change.
| species | location | hypothesized proximate cause of local extinction | reference |
|---|---|---|---|
| American pika (Ochotona princeps) | Great Basin region, USA | limited tolerance to temperature extremes (both high and low) | [25,63] |
| planarian (Crenobia alpina) | Wales, UK | loss of prey as result of increasing stream temperatures | [35] |
| desert bighorn sheep (Ovis canadensis) | California, USA | decrease in precipitation leading to altered plant community (food) | [64] |
| checkerspot butterfly (Euphydryas editha bayensis) | San Francisco Bay area, CA, USA | increase in variability of precipitation corresponding with reduction of temporal overlap between larvae and host plants | [66] |
| fish (Gobiodon sp. A) | New Britain, Papua New Guinea | destruction of obligate coral habitat due to coral bleaching caused by increasing water temperatures | [36] |
| 48 lizard species (genus Sceloporus) | Mexico | increased maximum air temperature approaches physiological limit, seemingly causing decreased surface activity during the reproductive season | [23] |
| Adrar Mountain fish species | Mauritania | loss of water bodies due to drought | [30] |
Figure 1.
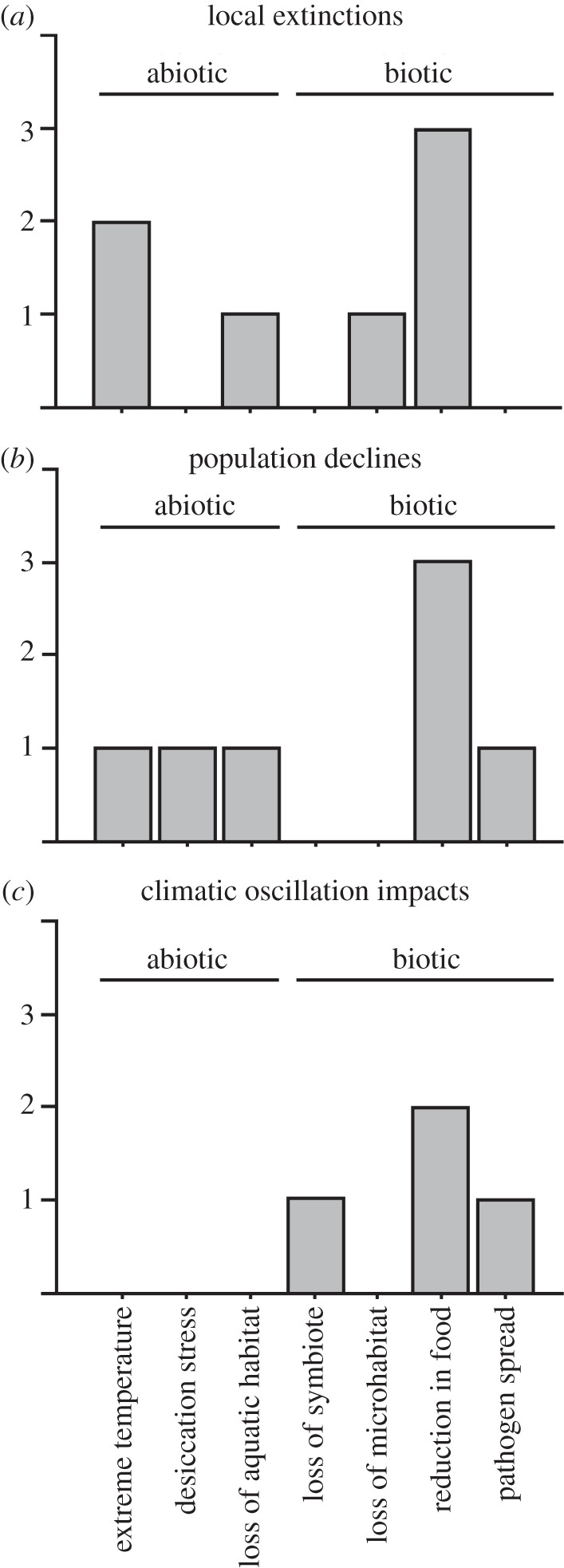
Summary of the frequency of different proximate causes of extinction due to climate change, among published studies. (a) ‘local extinctions’ refers to studies of local extinctions related to anthropogenic climate change (table 1), (b) ‘population declines’ refers to studies of declines in population abundance related to anthropogenic climate change (table 2), whereas (c) ‘climatic oscillation impacts’ refers to studies showing declines related to natural climatic oscillations (table 3) (but these oscillations may also be influenced by human factors, see relevant text). We note that there is some ambiguity in assigning some studies to a single, simple category.
(ii). Proximate causes of population declines
Seven studies identified proximate causes of population declines (table 2). The frequency of different proximate causes is intriguingly similar to those for population extinctions (figure 1a,b). Specifically, species interactions are the proximate cause of declines in the majority of studies, with declines in food availability being the most common cause [69,71,72], along with disease [70]. Drying of aquatic habitats is the cause in one study [29]. Two studies show physiological tolerances to abiotic factors as responsible for declines, with the declines being due to desiccation stress in desert trees [28], and due to oxygen limitation at high temperatures in a fish [24]. However, we find again that no studies show a straightforward relationship between population declines and temperatures exceeding the critical thermal limits of physiological tolerance.
Table 2.
Studies documenting the proximate causes of declines in abundance due to anthropogenic climatic change.
| species | location | hypothesized proximate cause of decline | reference |
|---|---|---|---|
| aloe tree (Aloe dichotoma) | Namib desert | desiccation stress owing to decreasing precipitation | [28] |
| four species of amphibians | Yellowstone National Park, USA | increasing temperature and decreasing precipitation cause a decline in habitat availability (pond drying) | [29] |
| plover (Pluvialis apricaria) | United Kingdom | high summer temperatures reduce abundance of craneflies (prey) | [69] |
| eelpout (Zoarces viviparus) | Baltic Sea | oxygen limitation at high temperatures | [24] |
| frogs (genus Atelopus) | Central and South America | climate change facilitates spread of pathogen (chytrid fungus) | [70] |
| grey jay (Perisoreus canadensis) | Ontario, Canada | warm autumns cause rotting in hoarded food, compromising overwinter survival and breeding success in the following year | [71] |
| Cassin's auklet (Ptychoramphus aleuticus) | California, USA | changes in upwelling timing and strength lower both adult survival and breeding success by changing food availability | [72] |
(iii). Proximate causes of extinction due to ‘natural’ climatic oscillations
Among the 136 studies, four documented proximate causes of climate-change related extinctions that were associated with climatic oscillations (table 3). These oscillations may increase in frequency and severity due to anthropogenic impacts ([77], but see [78]). All four studies reinforce the importance of species interactions as the proximate cause of many extinctions attributable to climate change (figure 1c), including climate-related losses of food resources [73,75], loss of an algal symbiont (‘coral bleaching’; [74]) and pathogen infection [76].
Table 3.
Studies that report proximate causes of declines in abundance or fitness associated with El Niño-Southern Oscilliation (ENSO) events.
| species | location | hypothesized proximate cause of decline | reference |
|---|---|---|---|
| fig wasps (Hymenoptera: Agonidae) | Borneo | ENSO event causes obligate host trees (Ficus sp.) to fail to produce inflorescences, resulting in local extinction of pollinating wasps | [73] |
| corals | Panama and Ecuador | high sea surface temperatures cause bleaching and mortality | [74] |
| butterflyfish | Indian Ocean | climate-related loss of coral food source | [75] |
| toad (Bufo boreas) | Western USA | warming reduces water depth in ponds, which increases ultraviolet-B exposure of embryos, which in turn increases risk of fungal infection | [76] |
Two of the most widely discussed examples of climate-change related extinctions involve chytrid fungus in amphibians and coral bleaching (including many examples given above [36,70,74,75]). In both cases, local extinctions are strongly connected to natural climatic oscillations (e.g. [74]), but the links to anthropogenic climate change are still uncertain. For example, Pounds et al. [42] concluded that chytrid-related declines and extinctions in the frog genus Atelopus are related to anthropogenic warming, but Rohr & Raffel [70] subsequently suggested that chytrid spread in Atelopus was largely due to El Niño events. The link between anthropogenic climate change and local extinction of coral populations through bleaching also remains speculative [79]. For example, severe climate anomalies can cause bleaching and coral mortality [80], but bleaching itself does not always lead to mass mortality [81].
(c). Proximate causes of extinction: synthesis
Our review of the proximate causes of population extinctions and declines due to climate change reveals three main results, which are concordant across the three categories of studies (extinctions, declines and climatic oscillations). First, very few studies have documented proximate factors (18 of 136). Second, a diversity of proximate causes are empirically supported. Third, changing interspecific interactions are the most commonly demonstrated causes of extinctions and declines (figure 1). Specifically, changes in biotic interactions leading to reduced food availability are the single most common proximate factor (figure 1). In contrast, limited physiological tolerances to high temperatures are supported only infrequently and indirectly (figure 1). Interestingly, the impacts of species interactions may be particularly difficult to document, inviting underestimation. However, we caution that these generalizations are based on few studies. For example, all three datasets (tables 1–3) are dominated by vertebrates, with only one plant study represented. Thus, the frequencies of documented proximate causes may change as the pool of studies becomes more taxonomically representative.
Finally, we note that we did not specifically address global species extinctions associated with climate change in our review. However, IUCN lists 20 species as extinct or extinct in the wild that potentially declined because of climate change (see the electronic supplementary material, table S1). Of these 20 species, seven are frogs that were possibly infected by chytrid fungus, which may be facilitated by climate change (see above). Four are snails, which may have become extinct as a result of drought. Two are freshwater fishes that lost their habitats because of drought. Among the six birds, two were also potentially affected by drought. The other four birds are island species possibly impacted by storms (the severity of which may be related to climate change), but these all had clear non-climatic threats. A similar pattern occurs in one island rodent species. In almost all cases, the links between extinction and anthropogenic climate change are speculative (but see [82]), which is why these cases were not included previously in our review. Intriguingly, none of the 20 is clearly related to limited tolerances to high temperatures (see the electronic supplementary material, table S1).
5. Approaches for finding the proximate causes of climate-related extinction
Our review demonstrates that disturbingly little is known about the proximate causes of extinctions due to recent climate change. How can this important gap be filled? Many approaches are possible, and we very briefly summarize two general frameworks that are beginning to be used. One focuses on individual species at multiple localities [23,25,63], the other on species assemblages at a particular locality [83–85]. These approaches are summarized graphically in the electronic supplementary material, figure S2.
Focusing on individual species (see the electronic supplementary material, figure S2), one must first document local extinctions or declines. To test whether populations have gone extinct, the present and past geographical ranges of the species can be compared. These analyses need not require surveying the entire species range, but could focus on a more limited series of transects (e.g. near the lowest latitudes and elevations, where ranges may already be limited by climatic factors [69,86]). The historical range can be determined from literature records and/or museum specimen localities [87]. These latter data are becoming increasingly available through online databases (e.g. GBIF; http://www.gbif.org/). Next, the species range (or select transects) should be resurveyed to document which populations are extant [23,56]. Evaluating whether populations persist is not trivial, and recent studies [56,88] have applied specialized approaches (e.g. occupancy modelling [89]). Furthermore, resurveys should account for false absences that may be misinterpreted as extinctions and for biases created by unequal sampling effort in space and time [87,90,91].
Documenting climate-related declines presents different challenges than documenting extinctions, given that most species lack data on population parameters over time. Some populations have been the focus of long-term monitoring, facilitating detailed studies of climate change impacts [86,92]. Large-scale databases on population dynamics through time are now becoming available. For example, the Global Population Dynamics Database [93] contains nearly 5000 time-series datasets. However, for many species, resurveying ranges to document local extinctions may be a necessary first step instead.
Given demonstrable local extinctions or declines, the next step is to determine whether these are related to large-scale trends in global climate change. Peery et al. [94] summarize six approaches that can be used to relate environmental factors to population declines [95]. These same approaches can be applied to connect global climate change and local extinctions. Relationships between changes in climate over time and population extinction versus persistence can be tested using GIS-based climatic data for relatively fine time scales (e.g. each month and year; PRISM; [96]). These analyses should preferably include data on other potential causes of local extinction not directly related to climate change, such as human habitat modification [64]. These analyses should help establish whether the observed local extinctions or declines are indeed due to climate change. If so, the next step is to understand their proximate causes.
Correlative analyses can be carried out to generate and test hypotheses about which proximate causes may be involved. Biophysical modelling [97] may be especially useful for these analyses, as it can incorporate many important factors, such as microclimate [98] and related variables (e.g. shade, wind speed, cloudiness, humidity) and relevant behavioural, ecological, demographic and physiological parameters [14,23]. Dissecting the specific aspects of climate that are most strongly associated with local extinctions may be important (e.g. is it warmer temperatures in the hottest part of the year, or the coldest?). Correlative studies can also test potential biotic factors, including the association between population extinctions or declines and the abundance of other species with negative impacts on the species in question (e.g. competitors, pathogens) or reductions in species necessary for persistence (e.g. prey, hosts). Two-species occupancy models [99] could be applied to test for the impacts of these and other types of interspecific interactions. Identifying the particular interactions that are responsible for climate-related extinctions may be challenging, given the diversity of interactions and species that may be involved. However, our results suggest that changing biotic interactions may be the most common proximate causes of climate-related extinction (figure 1).
Once potential factors are identified with correlative studies, these can be tested with mechanistic analyses. These could include experimental tests of physiological tolerances to relevant temperature and precipitation regimes [10,24,86,100], and laboratory and field tests of species interactions [39]. Transplant experiments that move individuals from extant populations into nearby localities where the species has recently gone extinct [100] may be particularly useful (for species in which this is practical). In many ways, experimental analyses can provide the strongest tests of the hypothesized causes of local extinctions. However, these should be informed by broader correlative studies. For example, simply testing the physiological tolerances of a species to extremely high temperatures may say little about the causes of climate-associated local extinction in that species if those extinctions are actually caused by warmer temperatures in winter or the spread of a competitor.
The second major approach (see the electronic supplementary material, figure S2) is to focus on species assemblages at single localities over time [83–85], rather than analysing multiple localities across the range of one or more species. Given data on species composition at different points in time, the local extinctions or declines of certain species can be tested for association with temporal changes in climate. These losses can then be related to specific biological traits (e.g. greater loss of species with temperature-cued flowering times versus those using photoperiod, or species for which the site is near their southern versus northern range limits [84]). These relationships can then point the way to more mechanistic and experimental studies.
6. Questions for future research
Understanding the proximate factors that cause climate-related extinctions should be an urgent priority for future research and should open the door to many additional applied and basic questions. Are there specific conservation and management strategies that can be matched to specific extinction causes? Are there phylogenetic trends or life-history correlates [20] of these factors that may allow researchers to predict which factors will be important in a species without having to conduct lengthy studies within that species? Do different factors influence the ability of niche models to accurately predict range shifts and extinctions due to climate change (e.g. physiological tolerances versus species interactions)? Can species adapt to some potential causes of extinction and not others?
7. Conclusions
Climate change is now recognized as a major threat to global biodiversity, and one that is already causing widespread local extinctions. However, the specific causes of these present and future extinctions are much less clear. Here, we have reviewed the presently available evidence for the proximate causes of extinction from climate change. Our review shows that only a handful of studies have focused specifically on these factors, and very few suggest a straightforward relationship between limited tolerance to high temperatures and local extinction. Instead, a diverse set of factors is implicated, including effects of precipitation, food abundance and mismatched timing with host species. Overall, we argue that understanding the proximate causes of extinction from climate change should be an urgent priority for future research. For example, it is hard to imagine truly effective strategies for species conservation that ignore these proximate causes. We also outline some general approaches that may be used to identify these causes. However, we make the important caveat that the relative importance of different proximate causes may change radically over the next 100 years as climate continues to change, and limited physiological tolerances to high temperatures may become the dominant cause of extinction. Nevertheless, our review suggests the disturbing possibility that there may be many extinctions due to other proximate causes long before physiological tolerances to high temperatures become predominant.
Acknowledgements
We thank H. Resit Akçakaya, Amy Angert, Steven Beissinger, Doug Futuyma, Spencer Koury, Javier Monzón, Juan Parra and anonymous reviewers for discussion and helpful comments on the manuscript.
References
- 1.Thomas C. D., et al. 2004. Extinction risk from climate change. Nature 427, 145–148 10.1038/nature02121 (doi:10.1038/nature02121) [DOI] [PubMed] [Google Scholar]
- 2.Jetz W., Wilcove D. S., Dobson A. P. 2007. Projected impacts of climate and land-use change on the global diversity of birds. PLoS Biol. 5, 1211–1219 10.1371/journal.pbio.0050157 (doi:10.1371/journal.pbio.0050157) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Leadley P., et al. Biodiversity scenarios: projections of 21st century change in biodiversity and associated ecosystem services. 2010. Secretariat of the Convention on Biological Diversity, Montreal, 2010.
- 4.Pereira H. M., et al. 2010. Scenarios for global biodiversity in the 21st century. Science 330, 1496–1501 10.1126/science.1196624 (doi:10.1126/science.1196624) [DOI] [PubMed] [Google Scholar]
- 5.Dawson T. P., Jackson S. T., House J. I., Prentice I. C., Mace G. M. 2011. Beyond predictions: biodiversity conservation in a changing climate. Science 332, 53–58 10.1126/science.1200303 (doi:10.1126/science.1200303) [DOI] [PubMed] [Google Scholar]
- 6.Hof C., Araujo M. B., Jetz W., Rahbek C. 2011. Additive threats from pathogens, climate and land-use change for global amphibian diversity. Nature 480, 516–519 10.1038/nature10650 (doi:10.1038/nature10650) [DOI] [PubMed] [Google Scholar]
- 7.Bellard C., Bertelsmeier C., Leadley P., Thuiller W., Courchamp F. 2012. Impacts of climate change on the future of biodiversity. Ecol. Lett. 15, 5–377 10.1111/j.1461-0248.2011.01736.x (doi:10.1111/j.1461-0248.2011.01736.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Loarie S. R., Duffy P. B., Hamilton H., Asner G. P., Field C. B., Ackerly D. D. 2009. The velocity of climate change. Nature 462, 1052–1055 10.1038/nature08649 (doi:10.1038/nature08649) [DOI] [PubMed] [Google Scholar]
- 9.Monzón J., Moyer-Horner L., Palamar M. B. 2011. Climate change and species range dynamics in protected areas. BioScience 61, 752–761 10.1525/bio.2011.61.10.5 (doi:10.1525/bio.2011.61.10.5) [DOI] [Google Scholar]
- 10.Deutsch C. A., Tewksbury J. J., Huey R. B., Sheldon K. S., Ghalambor C. K., Haak D. C., Martin P. R. 2008. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl Acad. Sci. USA 105, 6668–6672 10.1073/pnas.0709472105 (doi:10.1073/pnas.0709472105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Huey R. B., Deutsch C. A., Tewksbury J. J., Vitt L. J., Hertz P. E., Perez H. J. A., Garland T. 2009. Why tropical forest lizards are vulnerable to climate warming. Proc. R. Soc. B 276, 1939–1948 10.1098/rspb.2008.1957 (doi:10.1098/rspb.2008.1957) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Somero G. N. 2010. The physiology of climate change: how potentials for acclimatization and genetic adaptation will determine ‘winners’ and ‘losers’. J. Exp. Biol. 213, 912–920 10.1242/jeb.037473 (doi:10.1242/jeb.037473) [DOI] [PubMed] [Google Scholar]
- 13.Somero G. N. 2011. Comparative physiology: a ‘crystal ball’ for predicting consequences of global change. Am. J. Physiol. Regul. Integr. Comp. Physiol. 301, R1–R14 10.1152/ajpregu.00719.2010 (doi:10.1152/ajpregu.00719.2010) [DOI] [PubMed] [Google Scholar]
- 14.Kearney M., Shine R., Porter W. P. 2009. The potential for behavioral thermoregulation to buffer ‘cold-blooded’ animals against climate warming. Proc. Natl Acad. Sci. USA 106, 3835–3840 10.1073/pnas.0808913106 (doi:10.1073/pnas.0808913106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Memmott J., Craze P., Waser N., Price M. 2007. Global warming and the disruption of plant–pollinator interactions. Ecol. Lett. 10, 710–717 10.1111/j.1461-0248.2007.01061.x (doi:10.1111/j.1461-0248.2007.01061.x) [DOI] [PubMed] [Google Scholar]
- 16.Schweiger O., Heikkinen R.K., Harpke A., Hickler T., Klotz S., Kudrna O., Kühn I., Pöyry J., Settele J. 2012. Increasing range mismatching of interacting species under global change is related to their ecological characteristics. Glob. Ecol. Biogeogr. 21, 88–99 10.1111/j.1466-8238.2010.00607.x (doi:10.1111/j.1466-8238.2010.00607.x) [DOI] [Google Scholar]
- 17.Tylianakis J. M., Didham R. K., Bascompte J., Wardle D. A. 2008. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 11, 1351–1363 10.1111/j.1461-0248.2008.01250.x (doi:10.1111/j.1461-0248.2008.01250.x) [DOI] [PubMed] [Google Scholar]
- 18.Bonelli S., Cerrato C., Loglisci N., Balletto E. 2011. Population extinctions in the Italian diurnal Lepidoptera: an analysis of possible causes. J. Insect Conserv. 15, 879–890 10.1007/s10841-011-9387-6 (doi:10.1007/s10841-011-9387-6) [DOI] [Google Scholar]
- 19.Beever J. A., Belant J. L. (eds) 2012. Ecological consequences of climate change. Mechanisms, conservation, and management. Boca Raton, FL: CRC Press [Google Scholar]
- 20.Beever J. A., Belant J. L. 2012. Ecological consequences of climate change: synthesis and research needs. In Ecological consequences of climate change. Mechanisms, conservation, and management (eds Beever J. A., Belant J. L.), pp. 285–294 Boca Raton, FL: CRC Press [Google Scholar]
- 21.Sexton J. P., McIntyre P. J., Angert A. L., Rice K. J. 2009. Evolution and ecology of species range limits. Annu. Rev. Ecol. Evol. Syst. 40, 415–436 10.1146/annurev.ecolsys.110308.120317 (doi:10.1146/annurev.ecolsys.110308.120317) [DOI] [Google Scholar]
- 22.Brook B. W., Sodhi N.S., Bradshaw C. J. A. 2008. Synergies among extinction risks under global change. Trends Ecol. Evol. 23, 453–460 10.1016/j.tree.2008.03.011 (doi:10.1016/j.tree.2008.03.011) [DOI] [PubMed] [Google Scholar]
- 23.Sinervo B., et al. 2010. Erosion of lizard diversity by climate change and altered thermal niches. Science 328, 894–899 10.1126/science.1184695 (doi:10.1126/science.1184695) [DOI] [PubMed] [Google Scholar]
- 24.Pörtner H. O., Knust R. 2007. Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 315, 95–97 10.1126/science.1135471 (doi:10.1126/science.1135471) [DOI] [PubMed] [Google Scholar]
- 25.Beever E. A., Ray C., Wilkening J. L., Brussard P. F., Mote P. W. 2011. Contemporary climate change alters the pace and drivers of extinction. Glob. Change Biol. 17, 2054–2070 10.1111/j.1365-2486.2010.02389.x (doi:10.1111/j.1365-2486.2010.02389.x) [DOI] [Google Scholar]
- 26.Bradshaw W. E., Holzapfel C. M. 2010. Light, time, and the physiology of biotic response to rapid climate change in animals. Annu. Rev. Physiol. 72, 147–166 10.1146/annurev-physiol-021909-135837 (doi:10.1146/annurev-physiol-021909-135837) [DOI] [PubMed] [Google Scholar]
- 27.IPCC 2007. Climate change 2007, synthesis report. In Contributions of Working Groups I, II and III to the Fourth Assessment Report of the Intergovernmental Panel on Climage Change (eds Core Writing Team, Pachauri R. K., Reisinger A.), pp. 104 Geneva, Switzerland: IPCC [Google Scholar]
- 28.Foden W., et al. 2007. A changing climate is eroding the geographical range of the Namib Desert tree aloe through population declines and dispersal lags. Divers. Distrib. 13, 645–653 10.1111/j.1472-4642.2007.00391.x (doi:10.1111/j.1472-4642.2007.00391.x) [DOI] [Google Scholar]
- 29.McMenamin S. K., Hadley E. A., Wright C. K. 2008. Climatic change and wetland desiccation cause amphibian decline in Yellowstone National Park. Proc. Natl Acad. Sci. USA 105, 16 988–16 993 10.1073/pnas.0809090105 (doi:10.1073/pnas.0809090105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Trape S. 2009. Impact of climate change on the relict tropical fish fauna of Central Sahara: threat for the survival of Adrar Mountains fishes, Mauritania. PLoS ONE 4, e4400. 10.1371/journal.pone.0004400 (doi:10.1371/journal.pone.0004400) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Adams H. D., Guardiola-Claramonte M., Barron-Gafford G. A., Villegas J. C., Breshears D. D., Zou C. B., Troch P. A., Huxman T. E. 2009. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global change-type drought. Proc. Natl Acad. Sci. USA 106, 7063–7066 10.1073/pnas.0901438106 (doi:10.1073/pnas.0901438106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Crimmins S. M., Dobrowski S. Z., Greenberg J. A., Abatzoglou J. T., Mynsberge A. R. 2011. Changes in climatic water balance drive downhill shifts in plant species’ optimum elevations. Science 331, 324–327 10.1126/science.1199040 (doi:10.1126/science.1199040) [DOI] [PubMed] [Google Scholar]
- 33.Keith D. A., Akçakaya H. R., Thuiller W., Midgley G. F., Pearson R. G., Phillips S. J., Regan H. M., Araújo M. B., Rebelo T. G. 2008. Predicting extinction risks under climate change: coupling stochastic population models with dynamic bioclimatic habitat models. Biol. Lett. 4, 560–563 10.1098/rsbl.2008.0049 (doi:10.1098/rsbl.2008.0049) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Jones A. R. 2012. Climate change and sandy beach ecosystems. In Ecological consequences of climate change. Mechanisms, conservation, and management (eds Beever J. A., Belant J. L.), pp. 133–159 Boca Raton, FL: CRC Press [Google Scholar]
- 35.Durance I., Ormerod S. J. 2010. Evidence for the role of climate in the local extinction of a cool-water triclad. J. N. Am. Benthol. Soc. 29, 1367–1378 10.1899/09-159.1 (doi:10.1899/09-159.1) [DOI] [Google Scholar]
- 36.Munday P. L. 2004. Habitat loss, resource specialization, and extinction on coral reefs. Glob. Change Biol. 10, 1642–1647 10.1111/j.1365-2486.2004.00839.x (doi:10.1111/j.1365-2486.2004.00839.x) [DOI] [Google Scholar]
- 37.Wethey D. S. 2002. Biogeography, competition, and microclimate: the barnacle Chthamalus fragilis in New England. Integr. Comp. Biol. 42, 872–880 10.1093/icb/42.4.872 (doi:10.1093/icb/42.4.872) [DOI] [PubMed] [Google Scholar]
- 38.Suttle K. B., Thomsen M. A., Power M. E. 2007. Species interactions reverse grassland responses to changing climate. Science 315, 640–642 10.1126/science.1136401 (doi:10.1126/science.1136401) [DOI] [PubMed] [Google Scholar]
- 39.Goddard J. H. R., Gosliner T. M., Pearse J. S. 2011. Impacts associated with the recent range shift of the aeolid nudibranch Phidiana hiltoni (Mollusca, Opisthobranchia) in California. Mar. Biol. 158, 1095–1109 10.1007/s00227-011-1633-7 (doi:10.1007/s00227-011-1633-7) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Harley C. D. G. 2011. Climate change, keystone predation, and biodiversity loss. Science 334, 1124–1127 10.1126/science.1210199 (doi:10.1126/science.1210199) [DOI] [PubMed] [Google Scholar]
- 41.Benning T. L., LaPointe D., Atkinson C. T., Vitousek P. M. 2002. Interactions of climate change with biological invasions and land use in the Hawaiian Islands: modeling the fate of endemic birds using a geographic information system. Proc. Natl Acad. Sci. USA 99, 14 246–14 249 10.1073/pnas.162372399 (doi:10.1073/pnas.162372399) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Pounds J. A., et al. 2006. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 439, 161–167 10.1038/nature04246 (doi:10.1038/nature04246) [DOI] [PubMed] [Google Scholar]
- 43.Ytrehus B., Bretten T., Bergsjø B., Isaksen K. 2008. Fatal pneumonia epizootic in musk ox (Ovibos moschatus) in a period of extraordinary weather conditions. Ecohealth 5, 213–223 10.1007/s10393-008-0166-0 (doi:10.1007/s10393-008-0166-0) [DOI] [PubMed] [Google Scholar]
- 44.Stachowicz J. J., Terwin J. R., Whitlach R. B., Osman R. W. 2002. Linking climate change and biological invasions: ocean warming facilitates nonindigenous species invasions. Proc. Natl Acad. Sci. USA 99, 15 497–15 500 10.1073/pnas.242437499 (doi:10.1073/pnas.242437499) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Visser M. E., van Noordwijk A. J., Tinbergen J. M., Lessells C. M. 1998. Warmer springs lead to mistimed reproduction in great tits (Parus major). Proc. R. Soc. Lond. B 265, 1867–1870 10.1098/rspb.1998.0514 (doi:10.1098/rspb.1998.0514) [DOI] [Google Scholar]
- 46.Visser M. E., Holleman L. J. M. 2001. Warmer springs disrupt the synchrony of oak and winter moth phenology. Proc. R. Soc. Lond. B 268, 289–294 10.1098/rspb.2000.1363 (doi:10.1098/rspb.2000.1363) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.IUCN 2012. The IUCN red list of threatened species. Version 2012.1 See http://www.iucnredlist.org (accessed 19 June 2012) [Google Scholar]
- 48.Parmesan C., Yohe G. 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37–42 10.1038/nature01286 (doi:10.1038/nature01286) [DOI] [PubMed] [Google Scholar]
- 49.Thomas C. D. 2010. Climate, climate change, and range boundaries. Divers. Distrib. 16, 488–495 10.1111/j.1472-4642.2010.00642.x (doi:10.1111/j.1472-4642.2010.00642.x) [DOI] [Google Scholar]
- 50.Angert A. L., Crozier L. G., Rissler L. J., Gilman S. E., Tewksbury J. J., Chunco A. J. 2011. Do species’ traits predict recent shifts at expanding range edges? Ecol. Lett. 14, 677–689 10.1111/j.1461-0248.2011.01620.x (doi:10.1111/j.1461-0248.2011.01620.x) [DOI] [PubMed] [Google Scholar]
- 51.Parmesan C., Gaines S., Gonzalez L., Kaufman D. M., Kingsolver J., Peterson A. T., Sagarin R. 2005. Empirical perspectives on species borders: from traditional biogeography to global change. Oikos 108, 58–75 10.1111/j.0030-1299.2005.13150.x (doi:10.1111/j.0030-1299.2005.13150.x) [DOI] [Google Scholar]
- 52.Hickling R., Roy D. B., Hill J. K., Thomas C. D. 2005. A northward shift of range margins in the British Odonata. Glob. Change Biol. 11, 502–506 10.1111/j.1365-2486.2005.00904.x (doi:10.1111/j.1365-2486.2005.00904.x) [DOI] [Google Scholar]
- 53.Perry A. L., Low P. J., Ellis J. R., Reynolds J. D. 2005. Climate change and distribution shifts in marine species. Science 308, 1912–1915 10.1126/science.1111322 (doi:10.1126/science.1111322) [DOI] [PubMed] [Google Scholar]
- 54.Wilson J. W., Gutierrez D., Martinez D., Agudo R., Monserrat V. J. 2005. Changes to the elevational limits and extent of species ranges associated with climate change. Ecol. Lett. 8, 1138–1146 10.1111/j.1461-0248.2005.00824.x (doi:10.1111/j.1461-0248.2005.00824.x) [DOI] [PubMed] [Google Scholar]
- 55.Thomas C. D., Franco A. M. A., Hill J. K. 2006. Range retractions and extinctions in the face of climate warming. Trends Ecol. Evol. 21, 415–416 10.1016/j.tree.2006.05.012 (doi:10.1016/j.tree.2006.05.012) [DOI] [PubMed] [Google Scholar]
- 56.Moritz C., Patton J. L., Conroy C. J., Parra J. L., White G. C., Beissinger S. R. 2008. Impact of a century of climate change on small-mammal communities in Yosemite National Park, USA. Science 322, 261–264 10.1126/science.1163428 (doi:10.1126/science.1163428) [DOI] [PubMed] [Google Scholar]
- 57.Hawkins S. J., et al. 2009. Consequences of climate-driven biodiversity changes for ecosystem functioning of North European rocky shores. Mar. Ecol. Prog. Ser. 396, 245–259 10.3354/meps08378 (doi:10.3354/meps08378) [DOI] [Google Scholar]
- 58.Jones S. J., Lima F. P., Wethey D. S. 2010. Rising environmental temperatures and biogeography: poleward range contraction of the blue mussel, Mytilus edulis L., in the western Atlantic. J. Biogeogr. 37, 2243–2259 10.1111/j.1365-2699.2010.02386.x (doi:10.1111/j.1365-2699.2010.02386.x) [DOI] [Google Scholar]
- 59.Chen I. C., Hill J. K., Ohlemuller R., Roy D. B., Thomas C. D. 2011. Rapid range shifts of species associated with high levels of climate warming. Science 333, 1024–1026 10.1126/science.1206432 (doi:10.1126/science.1206432) [DOI] [PubMed] [Google Scholar]
- 60.Anderson B. J., Akçakaya H. R., Araújo M. B., Fordham D. A., Martinez-Meyer E., Thuiller W., Brook B. W. 2009. Dynamics of range margins for metapopulations under climate change. Proc. R. Soc. B 276, 1415–1420 10.1098/rspb.2008.1681 (doi:10.1098/rspb.2008.1681) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Gilchrist H. G., Mallory M. L. 2005. Declines in abundance and distribution of the ivory gull (Pagophila eburnea) in Arctic Canada. Biol. Conserv. 121, 303–309 10.1016/j.biocon.2004.04.021 (doi:10.1016/j.biocon.2004.04.021) [DOI] [Google Scholar]
- 62.Tyler N. J. C. 2010. Climate, snow, ice, crashes, and declines in populations of reindeer and caribou (Rangifer tarandus L.). Ecol. Monogr. 80, 197–219 10.1890/09-1070.1 (doi:10.1890/09-1070.1) [DOI] [Google Scholar]
- 63.Beever E. A., Ray C., Mote P. W., Wilkening J. L. 2010. Testing alternative models of climate-mediated extirpations. Conserv. Biol. 20, 164–178 10.1890/08-1011.1 (doi:10.1890/08-1011.1) [DOI] [PubMed] [Google Scholar]
- 64.Epps C. W., McCullough D., Wehausen J. D., Bleich V. C., Rechel J. L. 2004. Effects of climate change on population persistence of desert-dwelling mountain sheep in California. Conserv. Biol. 18, 102–113 10.1111/j.1523-1739.2004.00023.x (doi:10.1111/j.1523-1739.2004.00023.x) [DOI] [Google Scholar]
- 65.Caughley G. 1994. Directions in conservation biology. J. Anim. Ecol. 63, 215–244 10.2307/5542 (doi:10.2307/5542) [DOI] [Google Scholar]
- 66.McLaughlin J. F., Hellmann J. J., Boggs C. L., Ehrlich P. R. 2002. Climate change hastens population extinctions. Proc. Natl Acad. Sci. USA 99, 6070–6074 10.1073/pnas.052131199 (doi:10.1073/pnas.052131199) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gilman S. E., Urban M. C., Tewksbury J., Gilchrist G. W., Holt R. D. 2010. A framework for community interactions under climate change. Trends Ecol. Evol. 25, 325–331 10.1016/j.tree.2010.03.002 (doi:10.1016/j.tree.2010.03.002) [DOI] [PubMed] [Google Scholar]
- 68.Urban M., Tewksbury J. J., Sheldon K. S. 2012. On a collision course: competition and dispersal differences create no-analogue communities and cause extinctions during climate change. Proc. R. Soc. B 279, 2072–2080 10.1098/rspb.2011.2367 (doi:10.1098/rspb.2011.2367) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Pearce-Higgins J. W., Dennis P., Whittingham M. J., Yalden D. W. 2010. Impacts of climate on prey abundance account for fluctuations in a population of a northern wader at the southern edge of its range. Glob. Change Biol. 16, 12–23 10.1111/j.1365-2486.2009.01883.x (doi:10.1111/j.1365-2486.2009.01883.x) [DOI] [Google Scholar]
- 70.Rohr J. R., Raffel T. R. 2010. Linking global climate and temperature variability to widespread amphibian declines putatively caused by disease. Proc. Natl Acad. Sci. USA 107, 8269–8274 10.1073/pnas.0912883107 (doi:10.1073/pnas.0912883107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Waite T. A., Strickland D. 2006. Climate change and the demographic demise of a hoarding bird living on the edge. Proc. R. Soc. B 273, 2809–2813 10.1098/rspb.2006.3667 (doi:10.1098/rspb.2006.3667) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Wolf S. G., Snyder M. A., Doak D. F., Croll D. A. 2010. Predicting population consequences of ocean climate change for an ecosystem sentinel, the seabird Cassin's auklet. Glob. Change Biol. 16, 1923–1935 10.1111/j.1365-2486.2010.02194.x (doi:10.1111/j.1365-2486.2010.02194.x) [DOI] [Google Scholar]
- 73.Harrison R. D. 2000. Repercussions of El Niño: drought causes extinction and the breakdown of mutualism in Borneo. Proc. R. Soc. Lond. B 267, 911–915 10.1098/rspb.2000.1089 (doi:10.1098/rspb.2000.1089) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Glynn P. W., Mate J. L., Baker A. C., Calderon M. O. 2001. Coral bleaching and mortality in Panama and Ecuador during the 1997-1998 El Niño-Southern Oscillation event: spatial/temporal patterns and comparisons with the 1982–1983 event. Bull. Mar. Sci. 69, 79–109 [Google Scholar]
- 75.Graham N. A. J., Wilson S. K., Pratchett M. S., Polunin N. V. C., Spalding M. D. 2009. Coral mortality versus structural collapse as drivers of corallivorous butterflyfish decline. Biodivers. Conserv. 18, 3325–3336 10.1007/s10531-009-9633-3 (doi:10.1007/s10531-009-9633-3) [DOI] [Google Scholar]
- 76.Kiesecker J. M., Blaustein A. R., Belden L. K. 2001. Complex causes of amphibian population declines. Nature 410, 681–684 10.1038/35070552 (doi:10.1038/35070552) [DOI] [PubMed] [Google Scholar]
- 77.Timmermann A., Oberhuber J., Bacher A., Esch M., Latif M., Roeckner E. 1999. Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature 398, 694–697 10.1038/19505 (doi:10.1038/19505) [DOI] [Google Scholar]
- 78.Collins M., et al. 2010. The impact of global warming on the tropical Pacific Ocean and El Niño. Nat. Geosci. 3, 391–397 10.1038/ngeo868 (doi:10.1038/ngeo868) [DOI] [Google Scholar]
- 79.Pandolfi J. M., Connolly S. R., Marshall D. J., Cohen A. L. 2011. Projecting coral reef futures under global warming and ocean acidification. Science 333, 418–422 10.1126/science.1204794 (doi:10.1126/science.1204794) [DOI] [PubMed] [Google Scholar]
- 80.Hoegh-Guldberg O. 1999. Climate change, coral bleaching and the future of the world's coral reefs. Mar. Freshwater Res. 50, 839–866 10.1071/MF99078 (doi:10.1071/MF99078) [DOI] [Google Scholar]
- 81.Eakin C. M., et al. 2010. Caribbean corals in crisis: record thermal stress, bleaching, and mortality in 2005. PLoS ONE 5, e13969. 10.1371/journal.pone.0013969 (doi:10.1371/journal.pone.0013969) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Gerlach J. 2007. Short-term climate change and the extinction of the snail Rhachistia aldabrae (Gastropoda: Pulmonata). Biol. Lett. 3, 581–584 10.1098/rsbl.2007.0316 (doi:10.1098/rsbl.2007.0316) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Sagarin R. D., Barry J. P., Gilman S. E., Baxter C. H. 1999. Climate-related change in an intertidal community over short and long time scales. Ecol. Monogr. 69, 465–490 10.1890/0012-9615(1999)069[0465:CRCIAI]2.0.CO;2 (doi:10.1890/0012-9615(1999)069[0465:CRCIAI]2.0.CO;2) [DOI] [Google Scholar]
- 84.Willis C. G., Ruhfel B., Primack R. B., Miller-Rushing A. J., Davis C. C. 2008. Phylogenetic patterns of species loss in Thoreau's woods are driven by climate change. Proc. Natl Acad. Sci. USA 105, 17 029–17 033 10.1073/pnas.0806446105 (doi:10.1073/pnas.0806446105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wethey D. S., Woodin S. A., Hilbish T. J., Jones S. J., Lima F. P., Brannock P. M. 2011. Response of intertidal populations to climate: effects of extreme events versus long term change. J. Exp. Mar. Biol. Ecol. 400, 132–144 10.1016/j.jembe.2011.02.008 (doi:10.1016/j.jembe.2011.02.008) [DOI] [Google Scholar]
- 86.Dahlhoff E. P., Fearnley S. L., Bruce D. A., Gibbs A. G., Stoneking R., McMillan D. M., Deiner K., Smiley J. T., Rank N. E. 2008. Effects of temperature on physiology and reproductive success of a montane leaf beetle: implications for persistence of native populations enduring climate change. Physiol. Biochem. Zool. 81, 718–732 10.1086/590165 (doi:10.1086/590165) [DOI] [PubMed] [Google Scholar]
- 87.Tingley M. W., Beissinger S. R. 2009. Detecting range shifts from historical species occurrences: new perspectives on old data. Trends Ecol. Evol. 24, 625–633 10.1016/j.tree.2009.05.009 (doi:10.1016/j.tree.2009.05.009) [DOI] [PubMed] [Google Scholar]
- 88.Tingley M. W., Monahan W. B., Beissinger S. R., Moritz C. 2009. Birds track their Grinnellian niche through a century of climate change . Proc. Natl Acad. Sci. USA 106, 19 637–19 643 10.1073/pnas.0901562106 (doi:10.1073/pnas.0901562106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.MacKenzie D. I., Nichols J. D., Royle J. A., Pollock K. H., Bailey L. L., Hines J. E. 2006. Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Burlington, MA: Academic Press [Google Scholar]
- 90.Charmantier A., McCleery R. H., Cole L. R., Perrins C., Kruuk L. E. B., Sheldon B. C. 2008. Adaptive phenotypic plasticity in response to climate change in a wild bird population . Science 320, 800–803 10.1126/science.1157174 (doi:10.1126/science.1157174) [DOI] [PubMed] [Google Scholar]
- 91.Boakes E. H., McGowan P. J. K., Fuller R. A., Chang-qing D., Clark N. E., O'Connor K., Mace G. M. 2010. Distorted views of biodiversity: spatial and temporal bias in species occurrence data. PLoS Biol. 8, e1000385. 10.1371/journal.pbio.1000385 (doi:10.1371/journal.pbio.1000385) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Botts E. A., Erasmus B. F. N., Alexander G. J. 2011. Geographic sampling bias in the South African Frog Atlas Project: implications for conservation planning. Biodivers. Conserv. 20, 119–139 10.1007/s10531-010-9950-6 (doi:10.1007/s10531-010-9950-6) [DOI] [Google Scholar]
- 93.NERC Centre for Population Biology, Imperial College 2010. The global population dynamics database version 2. See http://www3.imperial.ac.uk/cpb/research/patternsandprocesses/gpdd. [Google Scholar]
- 94.Peery M. Z., Beissinger S. R., Newman S. H., Burkett E. B., Williams T. D. 2004. Applying the declining population paradigm: diagnosing causes of poor reproduction in the marbled murrelet. Conserv. Biol. 18, 1088–1098 10.1111/j.1523-1739.2004.00134.x (doi:10.1111/j.1523-1739.2004.00134.x) [DOI] [Google Scholar]
- 95.Beissinger S. R., Wunderle J. M., Jr, Meyers J. M., Saether B. E., Engen S. 2008. Anatomy of a bottleneck: diagnosing factors limiting population growth in the Puerto Rican parrot. Ecol. Monogr. 78, 185–203 10.1890/07-0018.1 (doi:10.1890/07-0018.1) [DOI] [Google Scholar]
- 96.Daly C., Gibson W. P., Taylor G. H., Johnson G. L., Pasteris P. 2002. A knowledge-based approach to the statistical mapping of climate. Clim. Res. 22, 99–113 10.3354/cr022099 (doi:10.3354/cr022099) [DOI] [Google Scholar]
- 97.Buckley L. B., Urban M. C., Angilletta M. J., Crozier L. G., Rissler L. J., Sears M. W. 2010. Can mechanism inform species distribution models? Ecol. Lett. 13, 1041–1054 10.1111/j.1461-0248.2010.01479.x (doi:10.1111/j.1461-0248.2010.01479.x) [DOI] [PubMed] [Google Scholar]
- 98.Helmuth B., et al. 2006. Mosaic patterns of thermal stress in the rocky intertidal zone: implications for climate change. Ecol. Monogr. 76, 461–479 10.1890/0012-9615(2006)076[0461:MPOTSI]2.0.CO;2 (doi:10.1890/0012-9615(2006)076[0461:MPOTSI]2.0.CO;2) [DOI] [Google Scholar]
- 99.Richmond O. M. W., Hines J. E., Beissinger S. R. 2010. Two-species occupancy models: a new parameterization applied to co-occurrence of secretive rails. Ecol. Appl. 20, 2036–2046 10.1890/09-0470.1 (doi:10.1890/09-0470.1) [DOI] [PubMed] [Google Scholar]
- 100.Jones S. J., Mieszkowska N., Wethey D. S. 2009. Linking thermal tolerances and biogeography: Mytilus edulis (L.) at its southern limit on the east coast of the United States. Biol. Bull. 217, 73–85 [DOI] [PubMed] [Google Scholar]