

Abstract
Land-use change is one of the main drivers of current and likely future biodiversity loss. Therefore, understanding how species are affected by it is crucial to guide conservation decisions. Species respond differently to land-use change, possibly related to their traits. Using pan-tropical data on bird occurrence and abundance across a human land-use intensity gradient, we tested the effects of seven traits on observed responses. A likelihood-based approach allowed us to quantify uncertainty in modelled responses, essential for applying the model to project future change. Compared with undisturbed habitats, the average probability of occurrence of bird species was 7.8 per cent and 31.4 per cent lower, and abundance declined by 3.7 per cent and 19.2 per cent in habitats with low and high human land-use intensity, respectively. Five of the seven traits tested affected the observed responses significantly: long-lived, large, non-migratory, primarily frugivorous or insectivorous forest specialists were both less likely to occur and less abundant in more intensively used habitats than short-lived, small, migratory, non-frugivorous/insectivorous habitat generalists. The finding that species responses to land use depend on their traits is important for understanding ecosystem functioning, because species' traits determine their contribution to ecosystem processes. Furthermore, the loss of species with particular traits might have implications for the delivery of ecosystem services.
Keywords: birds, functional traits, land-use change, likelihood-based model, tropical forest
1. Introduction
Anthropogenic conversion of natural habitats presents one of the greatest threats to biodiversity: land-use change will probably remain a greater threat than climate change over the coming decades, especially in the tropics [1–3]. Habitat loss caused by land-use change limits species' ability to move in response to climate change, and hence will probably exacerbate the effects of climate change by increasing species extinctions [4]. Moreover, with global human population expected to reach approximately 9.3 billion by 2050 [5], coupled with increased wealth and demand for more varied diets, the extent of land-use change is likely to increase rapidly. In order to reduce biodiversity loss and to ensure the continued functioning of ecosystems, we need to understand how present and future land-use change will impact species.
While the densities of most species are reduced in human-dominated landscapes [6,7], different species show widely varying responses to environmental change, and the extent and form of the response often depends on species' ecological and morphological traits. For example, the responses of a number of taxa—measured by extinction risk [8–11], population trends [12,13], range shifts [14,15] or sensitivity to specific drivers, such as land-use change [16], habitat fragmentation [17–21], disturbance [22] and infrastructure [23]—to recent environmental changes correlate with a variety of traits, including body size, fecundity, diet, diet breadth, habitat breadth, natural abundance and range size. However, we are still a long way from a general understanding of how traits determine the response of species to land-use change.
For birds, the best understood class of organisms, extinction risk correlates with body size, habitat and diet specialization, migratory status and generation length [24,25]. For understanding changes in the structure and function of ecosystems, it will be important to understand how the structure of local communities responds to environmental change. Local and regional studies have shown that sensitivity to habitat loss and habitat fragmentation is associated variously with body size, reproductive rate, diet and diet breadth, range size and population size [7,20,26–28]. An informal comparison among these studies suggests that species which are large-bodied, slow-breeding, sedentary, habitat and dietary specialists—especially frugivores and insectivores—with small ranges and small population sizes have shown the greatest declines in modified and fragmented habitats. These studies typically focused on species composition, measured as the presence or the absence of species, and relatively few have considered changes in the abundance of species that persist in modified habitats [7]. However, abundance changes are known to have important effects on the structure of communities and on the functioning of ecosystems [29]. Furthermore, these studies generally focused on particular regions or on a relatively small number of traits.
We present a pan-tropical meta-analysis of the occurrence and abundance of birds under different anthropogenic land-use intensities, to our knowledge the first such to consider the effect traits on observed responses. Using data from studies spanning five continents (North and South America, Africa, Asia and Oceania; figure 1), we simultaneously test the effect of seven traits: body mass, generation length, range size, trophic level, diet, migratory status and forest habitat specialization. We focus on tropical and sub-tropical forest biomes as these contain a large proportion of all bird species [30], and because human populations are projected to increase rapidly in these biomes during the rest of this century. The result is a set of conclusions about whether, and how, different traits affect the response of bird species to land-use change; a quantification of the magnitude of unexplained variation in response that remains after the effect traits have been accounted for; and a simple model that could be used in future studies to make projections of the potential effects of land-use change in bird communities at local, regional or pan-tropical scales.
Figure 1.
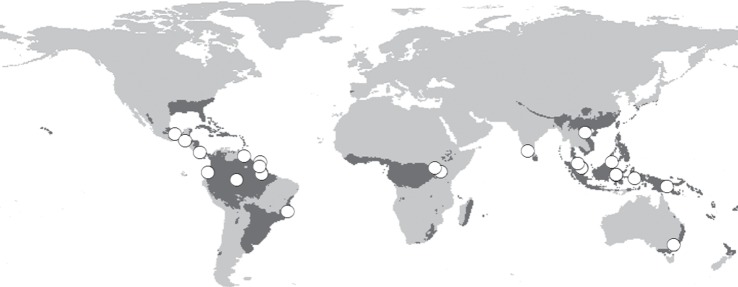
Location of studies included in analyses (white circles) based on geographical coordinates provided in the publication or georeferenced as accurately as possible from the description of the study areas provided. Therefore, these provide an overview of the geographical coverage of studies included here, rather than accurate locations. The extent of tropical and sub-tropical forest biomes is shown (grey shading) as used in the IMAGE [31], based on the BIOME model [32]. All gridded data are at 0.5° resolution. Shown in the World Geodetic System 1984 projection.
2. Material and methods
(a). Study area
The meta-analysis included studies from all tropical and sub-tropical areas (40° N–40° S) where the potential vegetation is forest. Potentially forested areas were defined according to the classification used in the Integrated Model to Assess the Global Environment (IMAGE [31]), which is based on the BIOME model [32] (figure 1).
(b). Abundance data
Estimates of local population density (4685 records for 1317 species, 22% of 6041 bird species whose range maps [33] intersected the study area) were collated from 24 studies in 23 peer-reviewed publications covering sites distributed throughout tropical and sub-tropical forests (figure 1). These were found using a Thomson Reuters Web of Knowledge search in April 2008 using the following search term: species diversity, biodiversity, richness or abundance; land use or habitat conversion; and pristine, primary, undisturbed or original; and supplemented by an additional search focused on the tropics in January 2010 using the following search term: [species] AND [tropic*] AND [primary forest OR mature forest OR intact forest OR old growth forest OR virgin forest OR pristine forest] AND [conversion OR degradation OR land use OR habitat type OR agro-forestry OR secondary forest OR plantation OR cropland OR urban area] AND [diversity OR biodiversity OR richness OR abundance] (a full list of references is given in the electronic supplementary material, appendix S1; for details of the methods used to select the papers, see [6]). To check that the one study from sub-tropical forests in Australia did not bias the results, since it is likely to be environmentally distinct from the other studies, we repeated the models excluding all data from this study.
Records consisted of direct estimates of population density, e.g. using distance sampling methods (194 records, in four studies), counts of individuals or of groups (2917 records, in 17 studies), or as the frequency of occurrence across a number of samples of a site (1574 records, in four studies). The latter two measures of density depend on sampling effort, which in some cases differed among samples in the same study. We adjusted records that used these two measures according to the relative sampling effort, assuming that perceived population density would increase linearly with sampling effort. The implications of using frequency of occurrence, which is based on the presence and absence of species, as a proxy for abundance warrants further testing with a larger dataset. However, for the main purpose of this study, which was to test the effect of species’ traits on responses to land-use intensity, a problem will only arise if traits have very different effects on occurrence and abundance of species.
Each record was assigned to one of three broad land-use intensity classes, based on human impacts on the vegetation at the site, as described in the paper from which the data were taken: ‘undisturbed habitat’ for sites in primary vegetation with very little human disturbance; ‘lightly used habitat’ for sites with intact natural vegetation but impacted to some degree by human activities, such as selective logging or harvesting of non-timber products, and including secondary vegetation; or ‘intensively used habitat’ for sites where the vegetation had been cleared for human use, including sites used for timber plantations, perennial tree crops, agroforestry (agriculture intercropped with trees), low-input arable agriculture, high-input arable agriculture, irrigated arable agriculture and pasture. A list of the specific local habitat types included in each of the classes is given in the electronic supplementary material, appendix S2.
The land-use intensity classes used were necessarily coarse given the size of the dataset. One of the distinctions excluded by this classification was between vegetation where vertical structure remained intact, for example, in forest plantations, and vegetation with no vertical structure, such as some agricultural land uses. Vertical structure may play a role in determining the abundance of species and the dominant traits in communities. In the future, when larger datasets are available, including the effect of vertical structure on species with different traits may enable a better understanding of community responses to land-use change.
Where species were sampled several times in different locations within one land-use intensity class within one study, we took the average of all non-zero densities across these samples. If a species was not recorded in a sampled land-use intensity class then it was assigned a density of zero. Differences among species in absolute recorded densities may be at least partly owing to differences in detectability [34]. To avoid this source of bias, we compared the relative abundance of each species among any land-use intensity classes sampled within each study.
(c). Trait data
We compiled data on seven traits: body mass, generation length, range size, migratory status, habitat affinity, diet and trophic level. Data on the first five of these came from BirdLife International's World Bird Database (available online at http://www.birdlife.org/datazone). The data on body mass were compiled primarily from Dunning [35], supplemented from a wide range of field guides, family monographs and other ornithological literature. Data on generation length were based on published and unpublished estimates for age at first breeding, survival and longevity, applied to the formulae recommended by the International Union for Conservation of Nature (IUCN) Standards and Petitions Subcommittee [36]. Where species-specific values of body mass and generation length were not available, for 36 and 26 species, respectively, we used estimates based on mean values for congeners.
Species were assigned to four migratory classes: non-migrants, nomads, altitudinal migrants and latitudinal/longitudinal migrants, using data from the ornithological literature applied to definitions developed by BirdLife International, as follows. Nomadic species move in response to resources that are sporadic and unpredictable in distribution and timing, and may congregate, but not predictably in terms of location and timing. Altitudinal migrants regularly or seasonally make cyclical movements to higher or lower elevations with predictable timing and destinations. Latitudinal/longitudinal migrants are species for which a substantial proportion of the global or regional population makes regular or seasonal cyclical movements beyond the breeding range, with predictable timing and destinations. This includes species that may be migratory only in part of their range/population, short-distance migrants and migrants that occasionally respond to unusual conditions in a semi-nomadic way.
Habitat preferences for each species were coded using the IUCN Habitats Classification Scheme (http://www.iucnredlist.org/technical-documents/classification-schemes/habitats-classification-scheme-ver3), based on information in the ornithological literature and reviewed by relevant experts. The upper level of the scheme, as used in the analyses, classifies habitats as: forest, savannah, shrubland, grassland, inland wetlands, rocky areas, caves, desert, marine neritic, marine oceanic, marine coastal, marine intertidal, artificial terrestrial/aquatic and introduced/exotic vegetation. The importance of each habitat inhabited by a species was classified as major, suitable, marginal or unknown (see http://www.birdlife.org/datazone/info/spchabalt). For the purposes of this study, species were considered as forest specialists, if forest was recorded as being of major importance to the species, or as generalists otherwise.
Range sizes were extracted from global distribution maps of bird species [33]. Diet data were compiled by one of us (C.H.S.) from the literature with most of the data from the Handbook of the birds of the world ([37]; for a detailed description and a complete list of sources see [38]). Species diet was classified into six groups according to their primary diet: fruit, nectar, other plant material, invertebrates, vertebrates, and mixed. We also pooled these classes by trophic level: herbivores (fruit-, nectar- and plant-eaters), carnivores (invertebrate- and vertebrate-eaters) or omnivores.
(d). Statistical analysis
We analysed the response of bird species to land use by using the data described above to parametrize simple models, using a likelihood-based model. The models all had the same overall structure (figure 2), defining: first, presence (and detection) in each land-use intensity class; and second, given presence, relative abundance, defined as local abundance relative to the abundance in undisturbed habitat (i.e. no change in abundance compared with undisturbed means a relative abundance of 1; a reduction in abundance meant relative abundance of less than 1; increased abundance meant a relative abundance above 1). These relative abundance ratios were log-transformed in all analyses. Observed relative abundance ratios were assumed to follow a lognormal distribution. The models were used to estimate seven parameters (figure 2): probabilities of presence in primary, light and intensive land use (PU, PL and PI); and the mean (mL and mI) and s.d. (sL and sI) of abundance ratios in light and intensive land use.
Figure 2.
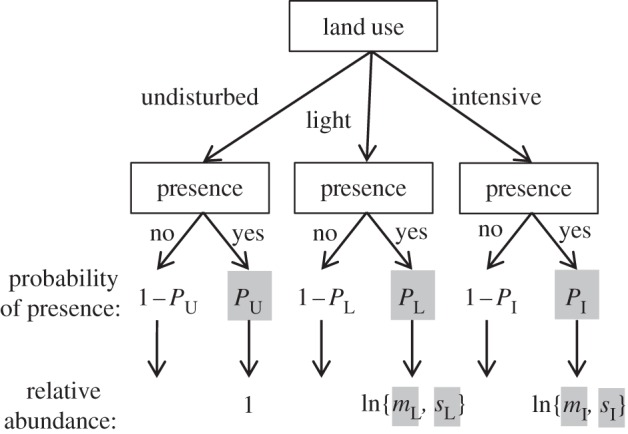
Model structure relating the presence or the absence, and given presence local abundance, of a species in three classes of land-use intensity (undisturbed, light use and intensive use). In each class, the species was defined as being present with probability PU, PL and PI. Abundances were normalized to 1 in the undisturbed land-use class. Observed abundances in the light and intensive land-use intensity classes were assumed to follow a lognormal distribution (denoted ln{}) with mean (mL and mI) and s.d. (sL and sI). Thus, the basic model estimated seven parameters, highlighted in grey.
To analyse the effect of species traits on responses to land use, an additional set of parameters was included to modify the probabilities of presence and abundance ratios according to species' traits. The effects of mass and generation length were treated as continuous functions having either a linear or quadratic effect (quadratic effects required more parameters but did not improve model fit, and so are not discussed further here). Trophic level, diet, migratory status and habitat specificity were treated as categorical variables.
We parametrized models containing every possible combination of traits separately against the data using standard methods based on Markov Chain Monte Carlo (MCMC) sampling. This in turn allowed us to use a standard method based on Akaike Information Criterion (AIC) values to: (i) calculate an overall measure of explanatory power for each trait, by weighting across the set of models according to their level of support given the data, following [39]; (ii) select the single best model from the set of models; and (iii) inspect parameter estimates, and confidence intervals on parameter estimates, in order to draw conclusions about the direction and magnitude of the effects of traits on occurrence and abundance.
The results of analyses comparing responses across species may be biased because of phylogenetic non-independence. Closely related species may respond similarly to land-use intensity owing to some factor not captured in the model. Including an effect of phylogeny in the models was complicated by the multi-factorial likelihood-based analysis used, and because there is not currently a fully resolved species-level phylogeny for birds. Instead, we tested whether a phylogenetic signal in the response of the bird species to land-use change remained unaccounted for in our models, by estimating Pagel's λ statistic for the residuals of each of the parameters in our best model (see the electronic supplementary material, appendix S3 for full details).
The response of species to land-use intensity might vary among the different studies included in the meta-analysis owing to factors not accounted for in the models, and this may bias the perceived effect of traits on observed responses. To check that this was not the case, we repeated the best model and models fitting each of the traits individually, with the addition of study as a factor. We did not run the full set of models with study as a factor, because the large number of free parameters required (up to 182) was larger than justified by the number of occurrence and abundance data and resulted in long convergence times for the models.
The data and code used to construct the models in this paper are available on request from the corresponding author.
3. Results
The probability of presence declined on average by 7.8 per cent and 31.4 per cent in lightly and intensively used habitat, respectively, compared with undisturbed habitat (figure 3a). Given presence, abundance was on average 3.7 per cent lower in lightly used habitat and 19.2 per cent lower in intensively used habitat compared with undisturbed habitat (figure 3b). However, there was considerable variation among species in observed responses to land-use intensity.
Figure 3.
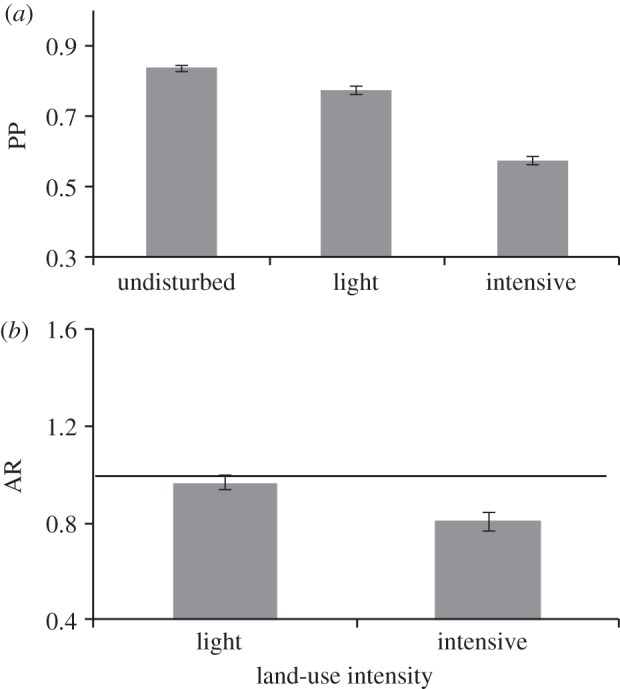
For 1317 tropical and sub-tropical bird species, modelled estimates of: (a) the probability of presence (and detection, PP) of any species in each of the three land-use intensities considered; and (b) given presence, the ratio of abundance (AR) in light and intensive land use relative to undisturbed. The horizontal line (y = 1) indicates no difference in abundance compared with undisturbed land use. Probabilities of presence and ratios of abundance were estimated using Bayesian analysis with a MCMC sampling process and uninformative priors. Error bars show 67% credible intervals.
Including traits of species improved the ability to explain observed changes in probability of presence and abundance. Of the 98 models with different combinations of traits considered, the null model (including no traits) ranked 88th in terms of its fit to the data (measured using the AIC). Except range size and trophic level, all traits had a substantial effect (sum of AIC weights > 0.3) on observed responses to land-use intensity (figure 4 and table 1) and were contained in at least one of the four highest ranked models in terms of fit to the data (table 2). Bird species that are non-migratory, dependent on forest habitat, and/or that have diets primarily of fruits and invertebrates occur with lower probability and at lower abundances in intensively used habitats than migratory habitat generalists with primary diets other than fruit and invertebrates (figure 4a–f). Furthermore, probability of occurrence and abundance were relatively lower in intensively used habitats compared with lightly used and undisturbed habitats for species with larger body masses and/or longer generation lengths compared with small and/or fast-breeding species (figure 4g–j).
Figure 4.

For 1317 tropical and sub-tropical bird species, modelled estimates of probabilities of presence (PP) in each of three land-use intensities (a,c,e,g,i) and, given presence, ratios of abundance (AR) in light relative to undisturbed, and in intensive relative to undisturbed land use (b,d,f,h,j). Probabilities of presence and ratios of abundance were modelled as functions of species' traits. Traits shown here are those for which the sum of AIC weights of all models containing them was greater than 0.3 (see [39] for more details): (a,b) forest habitat specificity (AIC weight sum ≈ 1), (c,d) migratory status (AIC weight sum ≈ 1), (e,f) diet (AIC weight sum = 0.997), (g,h) generation length (AIC weight sum = 0.689) and (i,j) body mass (AIC weight sum = 0.336). Methods were the same as used in figure 3.
Table 1.
The relative ability of each of six functional traits to explain observed responses of 1317 tropical and sub-tropical bird species to land-use intensity. Explanatory power was estimated as the sum of AIC weights [1] of all models containing a given trait, from a set of models fitting all possible combinations of traits. Response to land-use intensity was modelled as the probability of presence of species in each of three land-use intensity classes—undisturbed, light and intensive—and, given presence, the abundance ratio in light relative to undisturbed and, intensive relative to undisturbed land use. AIC weights were calculated following [1].
| trait | sum of AIC weights |
|---|---|
| mass | 0.336 |
| generation length | 0.689 |
| range size | 0.150 |
| migratory status | ≈1.000 |
| trophic level | 0.003 |
| diet | 0.997 |
| forest specialization | ≈1.000 |
Table 2.
The four best models for explaining the response of 1317 tropical and sub-tropical bird species to land-use intensity. (The fit of each model to the data was measured using the AIC, shown here as the difference between the AIC of the model in question and lowest AIC of all models. Traits included in these models were: GL, generation length; MIGR, migratory status; DIET, dietary guild; FORSPEC, whether the species was a forest specialist; MASS, body mass. Response to land-use intensity was modelled as the probability of presence of species in each of three land-use intensities—undisturbed, light and intensive—and, given presence, the abundance ratio in light relative to undisturbed and, intensive relative to undisturbed land use. AIC weights reflect the relative support for each model in terms of AIC values, and were calculated following [1].)
| rank | model | AIC difference | AIC weight |
|---|---|---|---|
| 1 | GL+MIGR+DIET+FORSPEC | 0 | 0.455 |
| 2 | MASS+GL+MIGR+DIET+FORSPEC | −2.35 | 0.131 |
| 3 | MASS+MIGR+DIET+FORSPEC | −2.53 | 0.124 |
| 4 | MIGR+DIET+FORSPEC | −2.60 | 0.057 |
Even in the best model—which included the effects of generation length, migratory behaviour, diet and habitat specialization—unexplained variation in observed responses to land-use intensity remained substantial. The parameters describing the standard deviation in observed log-transformed abundance ratios were 0.96 and 1.21 in light and intensively used habitat, respectively. An s.d. of 1 in log-space corresponds to variation in abundance change from a 65 per cent decline to a 2.7-fold increase.
Including the study from which data were taken as a factor in the models substantially improved the fit, but did not qualitatively alter the effect of traits on observed responses (see the electronic supplementary material, appendix S4). Dropping data from the Australian sub-tropical study similarly did not alter the results (see the electronic supplementary material, appendix S5). The phylogenetic signal in the residuals of the model was weak (λ < 0.09) and significant (p < 0.05) only for two of the five residual types (see the electronic supplementary material, appendix S3 for full details).
4. Discussion
Overall, the results show a substantial global impact of land use on the local abundance of bird species in tropical forests, consistent with the results of several previous studies on a wide range of species, including birds [1,3,6,40,41]. Compared with undisturbed land use, both the probability of species occurrence and abundance given occurrence, was slightly lower in land-use classes impacted by humans but where the native vegetation remains largely intact. In land-use classes involving clearance of the natural vegetation, species occurrence and abundance were both markedly reduced.
The effect of land-use intensity on tropical forest bird species differed among species with different ecological traits: species that are large-bodied, slower-breeding, non-migratory, that depend on forest habitat, and/or that feed primarily on fruit and invertebrates showed greater declines than other species. The effect of habitat specialization on the response to land-use intensity has been shown previously to affect the extinction risk of birds [24,25] as well as their response to habitat fragmentation [21]. Species that are dependent on forest habitat are likely to be more adversely impacted by forest clearance or degradation of forest habitat than species with broader habitat preferences. Reproductive rate, for which generation length was used here as a proxy, probably affects the ability of populations to recover after perturbations and probably also correlates with population abundance, which might affect species' vulnerability. Reproductive rate is known to correlate with extinction risk [24] and minimum habitat requirements of bird species [27], but is only available for a small minority of tropical bird species. Although migratory bird species are 2.6 times less likely to be threatened or near threatened with extinction [25], the effect of migratory status on response to land-use change has not been shown before. In fact, it is often migratory bird species that decline the most in response to habitat fragmentation [18]. In our analysis, migratory species tend also to be habitat generalists (χ2 = 633.6, p < 0.0001), being adapted to cope with a broader suite of habitats on migration.
It has been suggested that species at higher trophic levels should be more vulnerable to environmental changes because they occur at lower densities [18]. However, we found no evidence of an effect of trophic position on the response of birds to land-use change. On the other hand, diet had a very strong effect, with insectivores and frugivores declining most consistently in human-impacted land-use classes. The sensitivity of frugivorous and insectivorous birds to habitat fragmentation and to land-use change has been shown before (and may result from a reduction in the abundance of fruiting trees and invertebrates [21,26]), but this is, to our knowledge, the first analysis to show this effect using a global dataset. Large body size often correlates with increased extinction risk and sensitivity to human activities in a wide variety of animal taxonomic groups, including birds [11,24,26,28,38,42]. This is probably because body size correlates with traits that directly affect species' vulnerability, such as reproductive rate, abundance and trophic level [18,23]. In this study, the effect of body mass on response to land-use intensity was weak and was not included in the best model. Bird species with small range sizes have been shown to be more sensitive to habitat fragmentation than more wide-ranging species [42]. However, we failed to detect an effect of range size on the response to land-use intensity. Range size probably affects vulnerability indirectly because species with small range sizes typically occur at lower densities [43] and probably have more specific habitat requirements than more widespread species. It is also possible that species with small ranges and low density were missed in the studies included here, and that an effect of range size was thus missed. Range size may also correlate with other traits that we considered here, such as reproductive rate and body mass, which might account for the failure to detect an effect of range size on observed responses.
The results of analyses like this one can be biased by sampling artefacts and issues of detectability [34]. Abundance will differ among species and will also vary geographically for a single species. Furthermore, reported abundance will differ among species because of differences in detectability [34]. We avoided most of these issues by comparing the abundance among land-use classes of each species within each study, thus comparing abundances for the same species in a very similar geographical location. Nevertheless, issues of detectability may have remained. For example, it is likely that birds are more easily detected where vegetation has been cleared than in intact natural vegetation. As a result, the estimated overall response of bird species to land-use intensity would be an underestimate. On the other hand, there may have been a bias towards the publication of papers that showed a significant effect of land use on bird abundance; although species that did not respond to land use will also have been included, because most studies sampled whole bird assemblages. The perceived association between traits and responses to land-use intensity will only have been biased if the effect of traits and land use on the detectability of species covary with one another.
This study focused on the direct effects that local land-use intensity has on local bird communities. However, at landscape and regional scales, land use can result in fragmentation and reduction in area of remaining natural habitat (see [19] for a review). In future analyses, the effect of habitat fragmentation could be considered, assuming that accurate locations and associated environmental data for the individual sites sampled were available. Abundance could then be related to the size of habitat patches and the nature of the habitat in the surrounding landscape, derived, for example, from remotely sensed data.
Although traits improved our ability to explain observed changes in the occurrence and abundance of tropical bird species, considerable unexplained variation remained in the model. The four most obvious sources of this variability are: (i) an effect of traits of the species not captured in the models; (ii) a degree of subjectivity in the assignment of records to land-use classes because these were based on the assessments of individual authors; (iii) changes in abundance in response to other pressures, including habitat fragmentation and vegetation structure; and (iv) local and regional differences in the impact of different land-use regimes. The latter explanation is supported by the fact that fitting study as a factor in the models substantially improved the fit of the models. The effect of traits was consistent in spite of this spatial variation in observed responses, and the causes of the variation would make an interesting topic for future study. In practical terms, the consequence of unexplained variation in the models is that it will not be possible to make a precise prediction about the occurrence and abundance of a specific species in a specific location. However, the models can be used to make predictions of which species are more or less likely to be vulnerable to land-use change, and of change in the overall structure of bird communities in response to changes in land-use intensity.
More closely related species could be expected to respond more similarly to land-use intensity than more distantly related species, which could bias the apparent effect of traits on response to land-use intensity, since the traits are also likely to covary with relatedness. We tested for phylogenetic signal in the residuals of the best-fitting model, finding a significant signal for only two of the five residual types, namely for the probability of occurrence in lightly and intensively used habitats. The effect of traits was strong and consistent for both probabilities of occurrence and abundances. Therefore, the main result of our study was not affected by phylogenetic non-independence.
Overall, land-use change presents a serious threat to forest birds, especially for species with particular combinations of ecological traits. As well as allowing the identification of species of particular conservation concern, differences in the response of species with different sets of traits also have implications for the functioning of ecosystems and for the delivery of ecosystem services [38]. Disproportionate declines in the abundance of species with particular traits will reduce the functional diversity of biological communities, which will probably impair the functioning of ecosystems and make them less resilient to other environmental changes [44]. The loss of species with particular traits will also affect the delivery of specific ecosystem services. For example, insectivorous birds play an important role in pest control in ecosystems [45], while frugivores are essential for seed dispersal, especially in the tropics [46,47]. We show that both of these guilds show consistently negative responses to forest clearance in sites throughout the tropics, potentially having serious implications for agriculture. The novel likelihood-based approach to analysing responses to land use allows uncertainty in the observed responses to be quantified, which will be crucial when projecting future changes to biological communities.
Acknowledgements
We thank the volunteers and students (Felicity Bedford, Vicki Powell, Julie Day, Alexandra Robinson and Jake Simpson) who assisted with the compilation of the data, and Andy Purvis for invaluable advice on the project and comments on a draft of the manuscript. C.H.S. thanks the University of Utah for its support, and is grateful to Sherron Bullens, Debbie Fisher, David Hayes and especially Beth Karpas and Kathleen McMullen for their dedicated help with the world bird ecology database. We also thank two anonymous reviewers for valuable comments on an earlier draft of this paper. The data used in this study are available on request from T.N.
References
- 1.Jetz W, Wilcove DS, Dobson AP. 2007. Projected impacts of climate and land-use change on the global diversity of birds. PLoS Biol. 5, e157. 10.1371/journal.pbio.0050157 (doi:10.1371/journal.pbio.0050157) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Green RE, Cornell SJ, Scharlemann JPW, Balmford A. 2005. Farming and the fate of wild nature. Science 307, 550–555 10.1126/science.1106049 (doi:10.1126/science.1106049) [DOI] [PubMed] [Google Scholar]
- 3.Gibson L, et al. 2011. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 478, 378–381 10.1038/nature10425 (doi:10.1038/nature10425) [DOI] [PubMed] [Google Scholar]
- 4.Sekercioglu CH, Schneider SH, Fay JP, Loarie SR. 2008. Climate change, elevational range shifts, and bird extinctions. Conserv. Biol. 22, 140–150 10.1111/j.1523-1739.2007.00852.x (doi:10.1111/j.1523-1739.2007.00852.x) [DOI] [PubMed] [Google Scholar]
- 5.United Nations Population Division 2011. World Population Prospects: The 2010 Revision Population Database. See http://www.un.org/esa/population/. [Google Scholar]
- 6.Alkemade R, van Oorschot M, Miles L, Nellemann C, Bakkenes M, ten Brink B. 2009. GLOBIO3: a framework to investigate options for reducing global terrestrial biodiversity loss. Ecosystems 12, 374–390 10.1007/s10021-009-9229-5 (doi:10.1007/s10021-009-9229-5) [DOI] [Google Scholar]
- 7.Phalan B, Onial M, Balmford A, Green RE. 2011. Reconciling food production and biodiversity conservation: land sharing and land sparing compared. Science 333, 1289–1291 10.1126/science.1208742 (doi:10.1126/science.1208742) [DOI] [PubMed] [Google Scholar]
- 8.Koh LP, Sodhi NS, Brook BW. 2004. Ecological correlates of extinction proneness in tropical butterflies. Conserv. Biol. 18, 1571–1578 10.1111/j.1523-1739.2004.00468.x (doi:10.1111/j.1523-1739.2004.00468.x) [DOI] [Google Scholar]
- 9.Jones KE, Purvis A, Gittleman JL. 2003. Biological correlates of extinction risk in bats. Am. Nat. 161, 601–614 10.1086/368289 (doi:10.1086/368289) [DOI] [PubMed] [Google Scholar]
- 10.Kotiaho JS, Kaitala V, Komonen A, Päivinen J. 2005. Predicting the risk of extinction from shared ecological characteristics. Proc. Natl Acad. Sci. USA 102, 1963–1967 10.1073/pnas.0406718102 (doi:10.1073/pnas.0406718102) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Davidson AD, Hamilton MJ, Boyer AG, Brown JH, Ceballos G. 2009. Multiple ecological pathways to extinction in mammals. Proc. Natl Acad. Sci. USA 106, 10 702–10 705 10.1073/pnas.0901956106 (doi:10.1073/pnas.0901956106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kotze DJ, O'Hara RB. 2003. Species decline—but why? Explanations of carabid beetle (Coleoptera, Carabidae) declines in Europe. Oecologia 135, 138–148 10.1007/s00442-002-1174-3 (doi:10.1007/s00442-002-1174-3) [DOI] [PubMed] [Google Scholar]
- 13.Murray KA, Rosauer D, McCallum H, Skerratt LF. 2011. Integrating species traits with extrinsic threats: closing the gap between predicting and preventing species declines. Proc. R. Soc. B 278, 1515–1523 10.1098/rspb.2010.1872 (doi:10.1098/rspb.2010.1872) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Mattila N, Kaitala V, Komonen A, Kotiaho JS, Päivinen J. 2006. Ecological determinants of distribution decline and risk of extinction in moths. Conserv. Biol. 20, 1161–1168 10.1111/j.1523-1739.2006.00404.x (doi:10.1111/j.1523-1739.2006.00404.x) [DOI] [PubMed] [Google Scholar]
- 15.Mattila N, Kaitala V, Komonen A, Päivinen J, Kotiaho JS. 2011. Ecological correlates of distribution change and range shift in butterflies. Insect Conserv. Divers. 4, 239–246 10.1111/j.1752-4598.2011.00141.x (doi:10.1111/j.1752-4598.2011.00141.x) [DOI] [Google Scholar]
- 16.Barragán F, Moreno CE, Escobar F, Halffter G, Navarrete D. 2011. Negative impacts of human land use on dung beetle functional diversity. PLoS ONE 6, e17976. 10.1371/journal.pone.0017976 (doi:10.1371/journal.pone.0017976) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Davies KF, Margules CR, Lawrence JF. 2000. Which traits of species predict population declines in experimental forest fragments? Ecology 81, 1450–1461 10.1890/0012-9658(2000)081[1450:WTOSPP]2.0.CO;2 (doi:10.1890/0012-9658(2000)081[1450:WTOSPP]2.0.CO;2) [DOI] [Google Scholar]
- 18.Henle K, Davies KF, Kleyer M, Margules C, Settele J. 2004. Predictors of species sensitivity to fragmentation. Biodivers. Conserv. 13, 207–251 10.1023/B:BIOC.0000004319.91643.9e (doi:10.1023/B:BIOC.0000004319.91643.9e) [DOI] [Google Scholar]
- 19.Ewers RM, Didham RK. 2006. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 81, 117–142 10.1017/S1464793105006949 (doi:10.1017/S1464793105006949) [DOI] [PubMed] [Google Scholar]
- 20.Barbaro L, van Halder I. 2009. Linking bird, carabid beetle and butterfly life-history traits to habitat fragmentation in mosaic landscapes. Ecography 32, 321–333 10.1111/j.1600-0587.2008.05546.x (doi:10.1111/j.1600-0587.2008.05546.x) [DOI] [Google Scholar]
- 21.Vetter D, Hansbauer MM, Végvári Z, Storch I. 2011. Predictors of forest fragmentation sensitivity in Neotropical vertebrates: a quantitative review. Ecography 34, 1–8 10.1111/j.1600-0587.2010.06453.x (doi:10.1111/j.1600-0587.2010.06453.x) [DOI] [Google Scholar]
- 22.Bernhardt-Römermann M, et al. 2011. Functional traits and local environment predict vegetation responses to disturbance: a pan-European multi-site experiment. J. Ecol. 99, 777–787 10.1111/j.1365-2745.2011.01794.x (doi:10.1111/j.1365-2745.2011.01794.x) [DOI] [Google Scholar]
- 23.Rytwinski T, Fahrig L. 2011. Reproductive rate and body size predict road impacts on mammal abundance. Ecol. Appl. 21, 589–600 10.1890/10-0968.1 (doi:10.1890/10-0968.1) [DOI] [PubMed] [Google Scholar]
- 24.Owens IPF, Bennett PM. 2000. Ecological basis of extinction risk in birds: habitat loss versus human persecution and introduced predators. Proc. Natl Acad. Sci. USA 97, 12 144–12 148 10.1073/pnas.200223397 (doi:10.1073/pnas.200223397) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sekercioglu CH. 2007. Conservation ecology: area trumps mobility in fragment bird extinctions. Curr. Biol. 17, R283–R286 10.1016/j.cub.2007.02.018 (doi:10.1016/j.cub.2007.02.018) [DOI] [PubMed] [Google Scholar]
- 26.Cleary DFR, Boyle TJB, Setyawati T, Anggraeni CD, Van Loon EE, Menken SBJ. 2007. Bird species and traits associated with logged and unlogged forest in Borneo. Ecol. Appl. 17, 1184–1197 10.1890/05-0878 (doi:10.1890/05-0878) [DOI] [PubMed] [Google Scholar]
- 27.Vance MD, Fahrig L, Flather CH. 2003. Effect of reproductive rate on minimum habitat requirements of forest-breeding birds. Ecology 84, 2643–2653 10.1890/02-0159 (doi:10.1890/02-0159) [DOI] [Google Scholar]
- 28.Tscharntke T, Sekercioglu CH, Dietsch TV, Sodhi NS, Hoehn P, Tylianakis JM. 2008. Landscape constraints on functional diversity of birds and insects in tropical agroecosystems. Ecology 89, 944–951 10.1890/07-0455.1 (doi:10.1890/07-0455.1) [DOI] [PubMed] [Google Scholar]
- 29.Gaston KJ, Fuller RA. 2008. Commonness, population depletion and conservation biology. Trends Ecol. Evol. 23, 14–19 10.1016/j.tree.2007.11.001 (doi:10.1016/j.tree.2007.11.001) [DOI] [PubMed] [Google Scholar]
- 30.Sekercioglu CH. 2012. Bird functional diversity and ecosystem services in tropical forests, agroforests and agricultural areas. J. Ornithol. 153, S153–S161 10.1007/s10336-012-0869-4 (doi:10.1007/s10336-012-0869-4) [DOI] [Google Scholar]
- 31.Bouwman AF, Kram T, Klein Goldwijk K. (eds) 2006. Integrated modelling of global environmental change: an overview of IMAGE 2.4. Bilthoven, The Netherlands: Netherlands Environmental Assessment Agency [Google Scholar]
- 32.Prentice IC, Cramer W, Harrison SP, Leemans R, Monserud RA, Solomon AM. 1992. A global biome model based on plant physiology and dominance, soil properties and climate. J. Biogeogr. 19, 117–134 10.2307/2845499 (doi:10.2307/2845499) [DOI] [Google Scholar]
- 33.Birdlife International and NatureServe 2011. Bird species distribution maps of the world. See http://www.birdlife.org/datazone/info/spcdownload.
- 34.Thornton D, Branch L, Sunquist M. 2011. Passive sampling effects and landscape location alter associations between species traits and response to fragmentation. Ecol. Appl. 21, 817–829 10.1890/10-0549.1 (doi:10.1890/10-0549.1) [DOI] [PubMed] [Google Scholar]
- 35.Dunning JB. 1993. CRC handbook of avian body masses. Boca Raton, FL: CRC Press [Google Scholar]
- 36.IUCN Standards and Petitions Subcommittee 2010. Guidelines for Using the IUCN Red List Categories and Criteria, v. 8.0. See http://intranet.iucn.org/webfiles/doc/SSC/RedList/RedListGuidelines.pdf. [Google Scholar]
- 37.Del Hoyo J, Elliott A, Sargatal J. 1992. Handbook of the birds of the world. Barcelona, Spain: Lynx Edicions [Google Scholar]
- 38.Şekercioğlu ÇH, Daily GC, Ehrlich PR. 2004. Ecosystem consequences of bird declines. Proc. Natl Acad. Sci. USA 101, 18 042–18 047 10.1073/pnas.0408049101 (doi:10.1073/pnas.0408049101) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Burnham KP, Anderson DR. 2002. Model selection and multimodel inference: a practical information-theoretic approach. New York, NY: Springer [Google Scholar]
- 40.Sala OE, et al. 2000. Global biodiversity scenarios for the year 2100. Science 287, 1770–1774 10.1126/science.287.5459.1770 (doi:10.1126/science.287.5459.1770) [DOI] [PubMed] [Google Scholar]
- 41.Pimm S, Raven P, Peterson A, Şekercioğlu ÇH, Ehrlich PR. 2006. Human impacts on the rates of recent, present, and future bird extinctions. Proc. Natl Acad. Sci. USA 103, 10 941–10 946 10.1073/pnas.0604181103 (doi:10.1073/pnas.0604181103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Renjifo LM. 1999. Composition changes in a Subandean avifauna after long-term forest fragmentation. Conserv. Biol. 13, 1124–1139 10.1046/j.1523-1739.1999.98311.x (doi:10.1046/j.1523-1739.1999.98311.x) [DOI] [Google Scholar]
- 43.Gaston KJ, Blackburn TM, Lawton JH. 1997. Interspecific abundance-range size relationships: an appraisal of mechanisms. J. Anim. Ecol. 66, 579–601 10.2307/5951 (doi:10.2307/5951) [DOI] [Google Scholar]
- 44.Hooper DU, et al. 2005. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecol. Monogr. 75, 3–35 10.1890/04-0922 (doi:10.1890/04-0922) [DOI] [Google Scholar]
- 45.Van Bael SA, Brawn JD, Robinson SK. 2003. Birds defend trees from herbivores in a Neotropical forest canopy. Proc. Natl Acad. Sci. USA 100, 8304–8307 10.1073/pnas.1431621100 (doi:10.1073/pnas.1431621100) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sekercioglu CH. 2006. Increasing awareness of avian ecological function. Trends Ecol. Evol. 21, 464–471 10.1016/j.tree.2006.05.007 (doi:10.1016/j.tree.2006.05.007) [DOI] [PubMed] [Google Scholar]
- 47.Wenny DG, DeVault TL, Johnson MD, Kelly D, Sekercioglu CH, Tomback DF, Whelan CJ. 2011. The need to quantify ecosystem services provided by birds. Auk 128, 1–14 10.1525/auk.2011.10248 (doi:10.1525/auk.2011.10248) [DOI] [Google Scholar]