

Summary
Metabolism is influenced by age, food intake, and conditions such as diabetes and obesity. How do physiological or pathological metabolic changes influence stem cells, which are crucial for tissue homeostasis? This Commentary reviews recent evidence that stem cells have different metabolic demands than differentiated cells, and that the molecular mechanisms that control stem cell self-renewal and differentiation are functionally connected to the metabolic state of the cell and the surrounding stem cell niche. Furthermore, we present how energy-sensing signaling molecules and metabolism regulators are implicated in the regulation of stem cell self-renewal and differentiation. Finally, we discuss the emerging literature on the metabolism of induced pluripotent stem cells and how manipulating metabolic pathways might aid cellular reprogramming. Determining how energy metabolism regulates stem cell fate should shed light on the decline in tissue regeneration that occurs during aging and facilitate the development of therapies for degenerative or metabolic diseases.
Key words: Metabolism, Stem cells, ESCs, Reprogramming, iPSCs, HSCs, NSCs, AMPK, FOXO, mTOR, SIRT1, Insulin, Hypoxia, Aging
Introduction
Stem cells serve as the origin for all tissues during embryonic and postnatal development, and contribute to tissue homeostasis and repair throughout adult life. Stem cells hold great promise for replacement therapies for degenerative diseases and age-related disorders. Embryonic, postnatal and adult stem cells share two crucial characteristics: the ability to produce at least one daughter stem cell upon division (self-renewal) and the ability to generate differentiated cells (potency). Stem cell potency varies depending on the type of stem cell. For example, embryonic stem cells (ESCs) are pluripotent and can generate all three germ layers (endoderm, ectoderm and mesoderm) (Thomson et al., 1998). Stem cells that are present in adult tissues can be either multipotent or unipotent (i.e. giving rise to multiple differentiated cell types, or only one cell type, respectively) (Nakada et al., 2011). Interestingly, induced pluripotent stem cells (iPSCs) can be generated from either embryonic or adult differentiated cells upon expression of specific combinations of transcription factors (Takahashi and Yamanaka, 2006) (Box 1). As iPSCs can be generated from a specific patient, the use of these cells avoids potential medical or ethical issues when considering their application in regenerative medicine.
Box 1. Induced pluripotent stem cells and reprogramming.
In a landmark, Nobel prize-winning study, Takahashi and Yamanaka showed that the introduction of four transcriptional regulators (Oct4, Sox2, Klf4, Myc) was sufficient to convert mouse differentiated cells into ESC-like cells (Takahashi and Yamanaka, 2006). This cellular reprogramming was subsequently demonstrated in human cells (Takahashi et al., 2007; Yu et al., 2007). These pluripotent and self-renewing cells, termed induced pluripotent stem cells (iPSCs) appear to have most of the cellular and molecular properties of bona fide ESCs. Like ESCs, iPSCs are able to differentiate into all three germ layers (endoderm, mesoderm and ectoderm). When injected into a blastocyst, mouse iPSCs can produce viable chimeras that contribute to germline production (Okita et al., 2007). In the past few years, great strides have been made in generating iPSCs from a variety of mouse and human differentiated cell types using different combinations of reprogramming factors, chemicals and delivery methods (Feng et al., 2009; Wang and Na, 2011). Importantly, iPSCs have been derived from patients that are affected by various diseases (Grskovic et al., 2011), leading to patient-specific in vitro disease modeling. Patient-derived iPSCs facilitate the exploration of the genetic and molecular bases of human diseases and enable in vitro drug screening for these diseases. iPSCs also have benefits over ESCs for therapeutic applications, including eliminating the possibility of graft-versus-host disease and avoiding ethical concerns of human embryo-derived cells.
Emerging evidence suggests that pluripotent stem cells and certain adult stem cells are metabolically distinct from their differentiated counterparts and that these metabolic properties are important for stem cell identity. Furthermore, molecular regulators of energy metabolism have essential roles in stem cell fate, in particular, the decision to self-renew or differentiate. Finally, stem cells respond to fluctuations in organismal energy states in vivo. This Commentary will discuss the connections between stem cells and energy metabolism, focusing on human and mouse stem cells. The influence of metabolism on stem cells in other species has been described elsewhere (Jasper and Jones, 2010). The main stem cell types that will be discussed are ESCs and iPSCs as examples of pluripotent stem cells, and neural stem cells (NSCs) and hematopoietic stem cells (HSCs) as examples of adult tissue-specific stem cells.
Metabolic properties of stem cells
Stem cells appear to depend mostly on glycolysis for production of ATP
In contrast to differentiated cells, many stem cells appear to rely to a greater extent on glycolysis than on oxidative phosphorylation to generate adenosine-5′-triphosphate (ATP). Bioenergetics studies have revealed that human ESCs (Zhang et al., 2011b; Zhou et al., 2012) depend, in a large part, on glycolysis for ATP production (Fig. 1). Consistently, mitochondria are less complex and fewer in number in human ESCs than in their differentiated progeny (Cho et al., 2006; Facucho-Oliveira et al., 2007; St John et al., 2005; Varum et al., 2011; Zhang et al., 2011b). Furthermore, studies analyzing mitochondrial respiration, glycolytic flux or proteomic profiles of purified adult HSCs have shown that these adult stem cells rely primarily on glycolysis to generate ATP (Miharada et al., 2011; Simsek et al., 2010; Unwin et al., 2006). The dependency of stem cells on glycolysis for ATP generation is reminiscent of that of cancer cells (Hsu and Sabatini, 2008; Warburg, 1956). Unlike oxidative phosphorylation, glycolysis can proceed anaerobically, raising the possibility that the dependency of a stem cell on glycolysis is an adaptation to the low oxygen levels that are present in vivo during development and in an adult stem cell microenvironment or ‘niche’ (see below) (Fig. 1).
Fig. 1.

Energy sources in stem and differentiated cells. Many stem cell niches exhibit low oxygen concentrations. Stem cells appear to generate ATP mainly through glycolysis, which is independent of oxygen. Under low oxygen (<9% O2), the hypoxia-inducible factor 1α (HIF1α) is stabilized and binds to its partner HIF1β. The HIF1 heterodimer binds to hypoxia response elements to control the expression of genes involved in glucose metabolism and transport, the cell cycle and cell death. HIF1 activity appears to have an active role in the regulation of stem cell metabolism, as it can induce stem cells to shift towards a predominantly anaerobic glycolytic metabolism. Conversely, differentiated cells generate ATP largely through oxidative phosphorylation, which requires oxygen. ETC, electron transport chain; TCA, tricarboxylic acid.
Interestingly, metabolic differences between stem cells and differentiated cells might even promote the ‘stemness’ state. A glycolytic engagement is a crucial step in the conversion of terminally differentiated cells into iPSCs (Folmes et al., 2011; Panopoulos et al., 2012; Suhr et al., 2010; Varum et al., 2011). But do metabolic shifts precede pluripotency acquisition or does a stem cell state promote changes in metabolism? Evidence exists for both possibilities. Chemically inhibiting mitochondrial activity in human ESCs promotes the expression of pluripotency markers (Varum et al., 2009). However, the reprogramming factor OCT4 (also known as POU5F1) and the related factor OCT1 (also known as POU2F1) have many metabolic transcriptional targets, and loss of OCT1 leads to a shift away from glycolysis (Kang et al., 2009; Shakya et al., 2009), suggesting that transcriptional regulators of pluripotency and self-renewal coordinate the metabolism of stem cells. It is possible that during reprogramming a positive feedback loop is initiated, in which transcription factors first elicit a metabolic shift that is then necessary to induce additional endogenous pluripotency factors to complete the reprogramming into a stem cell state. The shift toward glycolysis might also minimize the production of reactive oxidative species (ROS), which could indirectly affect stem cell function, as described by Kobayashi and Suda (Kobayashi and Suda, 2012).
However, stem cell metabolism is likely to be more complex than a simple reliance on glycolysis. For example, one study reported that human ESCs generate the majority of ATP from oxidative phosphorylation, not from glycolysis (Birket et al., 2011). Such a difference to other studies is not completely understood yet, but could arise from the sensitivity of the detection technology used, or the fact that there might be slightly different stages of pluripotency – and thus metabolism – among ESCs (Zhou et al., 2012). Mouse ESCs also depend on the amino acid threonine as a crucial source of energy, whereby the citric acid cycle metabolite acetyl-coenzyme A is generated through the action of threonine dehydrogenase (Shyh-Chang et al., 2013; Wang et al., 2011; Wang et al., 2009). Interestingly, lipid metabolism is also emerging as a key regulator of stem cell maintenance and differentiation (Ito et al., 2012; Knobloch et al., 2013). A more in-depth analysis of the metabolism of stem cells is needed to clarify the circumstances under which the different components of respiration are utilized (Box 2).
Box 2. Technologies enabling studies of stem cell metabolism.
The comprehensive identification of potentially new metabolic states in stem cells will require unbiased methods. Advances in detection technologies, together with a developing excitement for understanding how metabolic state can influence cellular properties, has enabled the unbiased profiling of large numbers of metabolites in a single experiment, a field of study termed metabolomics. Metabolites are typically profiled by using liquid chromatography coupled with mass spectrometry or by using nuclear magnetic resonance. Emerging studies suggest that metabolomics studies can be performed on small numbers of cells (or even single cells) directly after their isolation and purification from tissue using fluorescence-activated cell sorting (FACS) methods (Rubakhin et al., 2011), which will aid our understanding of the metabolite profile of stem cells in vivo. Metabolic profiling of pluripotent stem cells has already determined that a bias towards a glycolytic metabolism is conducive to the acquisition of a pluripotency state (Folmes et al., 2011; Panopoulos et al., 2012). It has also been demonstrated that mouse ESCs possess more unsaturated molecules (containing double- and triple-bonded carbons) than differentiated cell types (Yanes et al., 2010). Specific unsaturated signaling molecules, such as the lipid second messengers arachidonic acid and diacylglyercol, appear to be crucial for ESC properties and subsequent multilineage differentiation (Yanes et al., 2010). Performing these types of metabolomic studies in additional contexts, for example, in adult stem cells, should allow the unbiased identification of ‘metabolic signatures’ of stem versus differentiated cells. Such metabolic signatures could then be coupled with other types of signatures (e.g. transcriptional or epigenetic signatures) to help to truly define ‘stemness’. Another technique called multi-isotope imaging mass spectrometry should also aid the characterization of stem cell metabolism through the high-resolution tracking of heavy isotope-labeled molecules as they are being utilized by the cell (Steinhauser et al., 2012). Knowing the differences in metabolic profiles as a function of stem cell type or external stimuli will be a key step in determining how metabolic properties of stem cells, in particular adult stem cells, are connected to quiescence and proliferation, differentiation potential and age-related changes (Rando, 2006).
It is worth noting that metabolic information obtained from cultured stem cells might not reflect the metabolism of stem cells in their natural niche for two main reasons (Joseph and Morrison, 2005). First, culture systems can differ quite dramatically from stem cell niches in terms of nutrient availability, cell–cell and cell–extracellular matrix contacts, and oxygen availability (Mohyeldin et al., 2010). These differences might be particularly significant for cells that are normally relatively quiescent, but are triggered to divide rapidly once they are exposed to growth factors in vitro, such as adult NSCs (Morshead et al., 1994). Second, stem cell cultures are often a heterogeneous mixture of stem cells, progenitors and differentiating cells. Future metabolic studies performed on small numbers of cells immediately after their isolation from tissue will aid our understanding of the metabolic profile of stem cells in vivo (Box 2).
Regardless of technical limitations, it is becoming increasingly clear that stem cells harbor different energy metabolism requirements compared with differentiating progeny. The unique metabolic properties of stem cells could be harnessed to facilitate the development of stem-cell-targeted therapies, in which stem cells are selectively directed to self-renew or differentiate by manipulating their metabolic state.
Oxygen availability directs metabolic and stem cell states
The dependency of stem cells on anaerobic glycolysis for energy production might be an adaptation to the low levels of oxygen that are present where these cells reside in vivo. For example, the uterus of rodents has particularly low levels of oxygen (3.5–5%) during the period of late blastocyst development and implantation (Fischer and Bavister, 1993), a period of intense stem cell proliferation. Adult HSCs and NSCs are also thought to reside in niches that are characterized by low oxygen levels (<1–6%) (Eliasson and Jönsson, 2010; Silver and Erecińska, 1998), and it is likely that most stem cell niches experience low oxygen although further investigation of other stem cell compartments is needed.
Accumulating evidence supports the idea that the fates of embryonic and adult stem cells are controlled by oxygen signaling (Mannello et al., 2011; Mohyeldin et al., 2010; Rafalski and Brunet, 2011; Suda et al., 2011). For example, lower levels of oxygen (3–5% compared with 20% atmospheric O2) promote human ESC self-renewal in vitro by preventing premature differentiation (Ezashi et al., 2005). Low oxygen concentrations (5%) also help the reprogramming of fibroblasts into iPSCs (Ezashi et al., 2005; Yoshida et al., 2009). In a similar manner, NSCs that are isolated from embryonic rodent or neonatal human brains show enhanced proliferation and reduced levels of cell death when cultured in low oxygen (Chen et al., 2007; Pistollato et al., 2007; Studer et al., 2000). Oxygen gradients in the niche might even help to direct stem cells to differentiate into specific cell lineages. Indeed, low oxygen (3–5%) can specify the fate of differentiating NSCs, promoting the production of dopaminergic neurons and oligodendrocytes (Pistollato et al., 2007; Studer et al., 2000).
The hypoxia-inducible transcription factors (HIF), which are stabilized and activated under low oxygen (<9% O2), are crucial for relaying the effect that oxygen has on stem cell fate. For example, HIF2α (also known as EPAS1) is necessary for mouse ESC self-renewal and the upregulation of pluripotency genes, such as Oct4 (Covello et al., 2006; Mathieu et al., 2011). Mice lacking HIF1α have substantial reductions in the number of HSCs in bone marrow transplantation assays and during normal aging (Takubo et al., 2010). The expression of Hif1α in HSCs is, in part, controlled by MEIS1, a transcription factor that is expressed in HSCs and downregulated during differentiation (Simsek et al., 2010), suggesting that HSCs are programmed to thrive under low oxygen. In a similar manner, HIF1 signaling is also important for normal mouse brain development (Tomita et al., 2003), and gene expression analyses comparing adult NSCs with differentiated neural cells indicate that adult NSCs have higher expression levels of Hif1a, emphasizing the importance of hypoxia signaling for NSCs (Bonnert et al., 2006; Ramalho-Santos et al., 2002).
Collectively, these studies suggest that low oxygen concentrations trigger alterations in HIF signaling, which, in turn, affects the metabolic and the ‘stemness’ networks of the cell, both of which might be linked by reciprocal amplification loops. Dissecting the interactions and feedback loops in this ‘metabolic–stemness’ network and understanding how it responds to external oxygen availability will be crucial in order to develop better methods to maintain stemness and enhance reprogramming.
Nutrient-sensing pathways coordinate energy metabolism with stem cell function
Nutrient-sensing signaling pathways orchestrate cellular and organismal metabolism in response to dietary changes. Accordingly, a number of these signaling pathways and molecules, including the insulin-forkhead box O factors (FOXO) pathway, mammalian target of rapamycin (mTOR), AMP-activated protein kinase (AMPK), and Sirtuins, have been implicated in the regulation of lifespan and healthspan (Greer and Brunet, 2008). This section reviews how nutrient-sensing pathways affect stem cell fate, thereby connecting energy metabolism with tissue regeneration and homeostasis.
The insulin–FOXO and mTOR pathways in stem cell quiescence and oxidative stress resistance
The insulin–FOXO pathway
The insulin–FOXO pathway regulates aging in a conserved manner from worms to mammals (Kenyon, 2010). Insulin and insulin growth factor 1 (IGF1) signaling leads to the phosphorylation of FOXO transcription factors (FOXOs) by the protein kinase AKT and serum-glucocorticoid regulated kinase (SGK) and the subsequent inactivation of these transcription factors by sequestration in the cytoplasm (Greer and Brunet, 2005). Conversely, FOXOs translocate to the nucleus when insulin and IGF1 signaling is reduced, that is, under conditions that lead to longevity. There are four FOXO family members in mammals (FOXO1, FOXO3, FOXO4, FOXO6). The isoform FOXO3 has been linked to exceptional longevity in several independent studies (Anselmi et al., 2009; Flachsbart et al., 2009; Pawlikowska et al., 2009; Willcox et al., 2008).
FOXOs have recently been shown to be essential for both adult and embryonic stem cells (Table 1). FOXO1, FOXO3 and FOXO4 are important for the long-term homeostasis of HSCs and NSCs in adult mice, as deletion of FOXO family members leads to the premature depletion of these adult stem cells (Miyamoto et al., 2007; Paik et al., 2009; Renault et al., 2009; Tothova et al., 2007; Yalcin et al., 2008). FOXO3, which is associated with exceptional human longevity, appears to be particularly important for the maintenance of HSCs and NSCs, as its deletion is sufficient to result in the depletion of HSCs and NSCs (Miyamoto et al., 2007; Paik et al., 2009; Renault et al., 2009; Tothova et al., 2007). FOXO1 is also crucial for the homeostasis of spermatogonial stem cells (SSCs) in adult mice (Goertz et al., 2011). Finally, FOXO1 has recently been found to be important for the pluripotency of both human and mouse ESCs (Zhang et al., 2011c), and FOXO4 upregulates proteasome activity, which is pivotal for human ESC pluripotency (Vilchez, et al., 2012). Although FOXO3 does not appear to control proteasome activity in human ESCs (Vilchez et al., 2012), it can regulate pluripotency in mouse ESCs (Zhang et al., 2011c). Thus, FOXO family members are important for the maintenance of adult stem cells and the pluripotency of ESCs, but the respective contribution of different FOXO isoforms can differ as a function of stem cell type.
Table 1. Effects of metabolic regulators on stem cell fate.
| Metabolic regulator | Stem cell type | Method of perturbation | Main phenotypes | References |
| FOXO transcription factors | ESCs | FOXO1 shRNA-mediated knockdown. | Loss of pluripotency markers, spontaneous differentiation into mesoderm and endoderm lineages, impaired teratoma formation. | (Zhang et al., 2011c) |
| ESCs | FOXO4 shRNA-mediated knockdown | Reduced proteasome activity | (Vilchez et al., 2012) | |
| HSCs | Conditional FOXO1, 3 and 4 deletion in adult hematopoietic lineage. | Premature depletion due to excessive proliferation, impaired self-renewal and increased apoptosis. High levels of ROS. | (Tothova et al., 2007) | |
| HSCs | Deletion of FOXO3. | Premature depletion due to impaired self-renewal and loss of quiescence. Myeloproliferative-like syndrome. High levels of ROS. | (Miyamoto et al., 2007; Yalcin et al., 2008, Yalcin et al., 2010) | |
| NSCs | Conditional FOXO1, 3, and 4 deletion in the brain. | Premature depletion due to excessive proliferation, impaired self-renewal and increased apoptosis. High levels of ROS. | (Paik et al., 2009) | |
| NSCs | Deletion of FOXO3. | Premature depletion due to excessive proliferation, impaired self-renewal and increased apoptosis. | (Renault et al., 2009) | |
| Muscle stem cells | Overexpression of FOXO3. | Decreased cell proliferation. | (Rathbone et al., 2008) | |
| Spermatogonial stem cells | Conditional FOXO1, 3, and 4 deletion in male germ line. | Increased cell death. | (Goertz et al., 2011) | |
| mTOR kinase | ESCs | Deletion of mTOR. | Aborted embryonic development, impaired cell proliferation and small cell size. | (Murakami et al., 2004) |
| ESCs | shRNA-mediated knockdown of mTOR or inhibition by rapamycin. | Decreased cell proliferation, loss of pluripotency markers, and increased mesoderm and endoderm differentiation. | (Schieke et al., 2008; Zhou et al., 2009) | |
| ESCs | Expression of constitutively active S6 kinase 1 (target of mTOR). | Increased differentiation. | (Easley et al., 2010) | |
| iPSCs | Inhibition by rapamycin. | Increased reprogramming efficiency. | (Chen et al., 2011) | |
| iPSCs | Hyperactivation by TSC knockdown or knockout. | Decreased reprogramming efficiency. | (He et al., 2012) | |
| HSCs | Hyperactivation of mTOR by deletion of TSC1 (negative regulator of mTOR). | Loss of cellular quiescence, increased apoptosis, elevated mitochondrial biogenesis and ROS levels; defective hematopoiesis in transplant assays. | (Gan et al., 2008; Chen et al., 2008) | |
| HSCs | Inhibition by rapamycin (treatment of old mice). | Rescue of age-related decline in self-renewal and hematopoiesis. | (Chen et al., 2009) | |
| NSCs | Hyperactivation of mTOR in dorsal telenchephalic neuroepithelium by deletion of TSC1 (negative regulator of mTOR). | Abnormal CNS development due to transient increase in cell proliferation, followed by deregulated differentiation and neuronal migration. | (Magri et al., 2011) | |
| Epidermal stem cells | Inhibition by rapamycin (treatment of Wnt1-overexpressing mice). | Rescue of Wnt1-associated cellular senescence. | (Castilho et al., 2009) | |
| AMPK | ESCs | Activation by AICAR. | Decreased cell proliferation, loss of pluripotency markers and enhanced erythroid lineage differentiation. | (Chae et al., 2012) |
| iPSCs | Activation by metformin or A-769662. | Decreased reprogramming efficiency. | (Vazquez-Martin et al., 2012) | |
| HSCs | Deletion of α1 and α2 subunits of AMPK. | Reduced numbers of HSCs but normal reconstitution capacity in transplant assays; decreased levels of ATP and mitochondrial DNA. | (Nakada et al., 2010) | |
| NSCs | Deletion of β1 subunit of AMPK. | Smaller brain size due to mitotic defects and increased apoptosis of differentiating progeny. | (Dasgupta and Milbrandt, 2009) | |
| SIRT1 | ESCs | Deletion or knockdown of SIRT1. | Largely normal but impaired DNA damage repair and apoptosis induction in response to oxidative stress. | (McBurney et al., 2003; Han et al., 2008b; Oberdoerffer et al., 2008; Chae and Broxmeyer, 2011) |
| iPSCs | Activation by reservatrol or fisetin. | Increased reprogramming efficiency. | (Chen et al., 2011) | |
| HSCs | Deletion of SIRT1. | Normal hematopoiesis in adult mice despite increased proliferation, anemia and leukocytosis. | (Narala et al., 2008; Leko et al., 2012) | |
| HSCs | Deletion of SIRT1. | Impaired hematopoietic differentiation from embryoid bodies; decreased in vitro survival of progenitors cultured from adult mice. | (Ou et al., 2011) | |
| HSCs | Deletion of SIRT1. | Impaired self-renewal of embryonic HSCs. | (Matsui et al., 2012) | |
| NSCs | Deletion or knockdown of SIRT1. | Increased differentiation of astrocytes at the expense of neurons from embryonic NSCs cultured under oxidative conditions. | (Prozorovski et al., 2008) | |
| NSCs | Knockdown or overexpression of SIRT1. | Impaired neuronal differentiation upon knockdown; enhanced neuronal differentiation upon overexpression. | (Hisahara et al., 2008) | |
| Muscle stem cells | Knockdown or overexpression of SIRT1. | Enhanced differentiation into myocytes upon knockdown; repressed differentiation into myocytes upon overexpression. | (Fulco et al., 2003) | |
| Muscle stem cells | Overexpression of SIRT1. | Increased cell proliferation. | (Rathbone et al., 2009) | |
| Spermatogonial stem cells | Deletion of SIRT1. | Reduced numbers probably due to increased levels of apoptosis; abnormal sperm differentiation. | (McBurney et al., 2003; Coussens et al., 2008) |
In NSCs and HSCs, FOXOs act by maintaining stem cell quiescence (infrequent cell cycle entry), a key feature of adult stem cells (Fig. 2). In the absence of FOXOs, more committed progenitors overproliferate, thereby exhausting the pool of quiescent stem cells. FOXOs upregulate the expression of proteins that are involved in cell cycle arrest, including p27KIP1, p57KIP2 and cyclin G2 (encoded by Cdkn1b, Cdkn1c and Ccng2, respectively), which are likely to contribute to maintaining quiescence (Paik et al., 2009; Renault et al., 2009; Tothova et al., 2007). FOXOs also directly repress the expression of abnormal spindle-like microcephaly-associated protein (ASPM), which is required for proliferative divisions of neural stem and progenitor cells (Paik et al., 2009), an activity that can also promote NSC quiescence. Interestingly, although FOXOs tend to promote quiescence in stem cells, FOXO function in non-cycling, differentiated cell types, such as neurons, appears to be largely to promote apoptosis in response to cellular stress (Salih and Brunet, 2008).
Fig. 2.

Nutrient-sensing pathways in stem cells. Nutrient and energy-responsive signaling pathways impact stem cells in a variety of ways. Shown here is a schematic illustration of the cellular components that respond to energy availability to influence stem cell metabolism and fate. Blue shading highlights molecules that are active in a high-energy state. Red shading highlights molecules that are inactive in a high-energy state, or that are active in response to cellular stresses, such as low oxygen and low energy. AKT, protein kinase B; AMPK, AMP-activated protein kinase; ETC, electron transport chain; FOXO, Forkhead Box O; HIF1, hypoxia-inducible factor 1; Insulin–IGF1R:insulin–insulin-like growth factor 1 receptor; LKB1, liver kinase B1; mTOR, mammalian target of rapamycin; PI3K, phosphatidylinositol 3-kinase; PTEN, phosphatase and tensin homolog; SIRT1, Sirtuin 1; SGK1, serum-glucocorticoid regulated kinase 1; S6K1, ribosomal S6 protein kinase 1; TCA, tricarboxylic acid cycle (also known as Krebs cycle or citric acid cycle); 4E-BP1, eukaryotic translation initiation factor 4E-binding protein 1.
Stem cell self-renewal is sensitive to oxidative stress (Ito et al., 2004) and FOXOs help maintain stem cells by preventing the accumulation of ROS, which can disrupt genomic and protein integrity. Indeed, some, but not all, of the consequences of FOXO loss on the HSC and NSC compartments can be rescued by the antioxidant N-acetyl cysteine (NAC) (Paik et al., 2009; Tothova et al., 2007; Yalcin et al., 2010). FOXOs are known to regulate several genes involved in the resistance to oxidative stress in many cell types (Dansen and Burgering, 2008). This activity not only helps to preserve the stem cell pool, but is also likely to minimize the incorporation of abnormal stem cell progeny into tissues and to avoid the transition from stem cell to cancer cell.
Interestingly, in NSCs, FOXO3 also regulates the expression of genes that are involved in hypoxia signaling (e.g. Ddit4, a known target of HIF1) (Renault et al., 2009). Accordingly, the pro-proliferative response of FOXO3-deficient NSCs to low oxygen (2%) is impaired in vitro (Renault et al., 2009). Like HIF1, FOXO3 regulates genes that are part of a ‘molecular signature’ for glycolysis and fructose metabolism (Renault et al., 2009). It is also worth noting that the phenotypes of FOXO3-deficient and HIF1-deficient HSCs are similar, particularly in terms of loss of cellular quiescence and premature depletion of the HSC pool (Miyamoto et al., 2007; Takubo et al., 2010). Together, these observations argue for a network of overlapping nutrient-sensing (through FOXO3) and oxygen-sensing (through HIF1) signaling.
Finally, in human ESCs, FOXO1 appears to regulate the expression of two crucial transcription factors for the stemness program, OCT4 and SOX2 (Zhang et al., 2011c). These data suggest that FOXO transcription factors are part of a transcriptional network that connects energy metabolism and ROS responses with the stem cell properties of self-renewal and potency.
mTOR
An essential component of cellular and organismal metabolism in mammals is the protein kinase mTOR. mTOR is activated by amino acids or the phosphatidylinositol 3-kinase (PI3K)–AKT pathway and is part of two distinct complexes, mTORC1 and mTORC2, which coordinate cell growth, proliferation and survival (Laplante and Sabatini, 2012) (Fig. 2). At the organismal level, mTOR is involved in energy metabolism regulation, cancer and aging (Kapahi et al., 2010; Laplante and Sabatini, 2012). For example, blocking mTOR activity with the pharmacological drug rapamycin can extend the lifespan of mice (Harrison et al., 2009).
Multiple recent studies have revealed vital, yet different, roles for mTOR signaling in ESCs and adult tissue-specific stem cells (Table 1). In ESCs, mTOR is important for self-renewal; inhibition of mTOR by rapamycin or by knockdown in mouse and human ESCs reduces the expression of pluripotency markers and impairs the self-renewal of ESCs in vitro (Schieke et al., 2008; Zhou et al., 2009). Furthermore, deletion of mTOR inhibits proliferation in early mouse embryos (Murakami et al., 2004). Somewhat surprisingly, mTOR also appears to promote ESC differentiation. For example, the transition from ESCs to more differentiated cell types is associated with increased activation of mTORC1 signaling (Easley et al., 2010; Sampath et al., 2008). Furthermore, overexpression of a constitutively active form of S6 kinase 1, a substrate of mTORC1, promotes differentiation of ESCs (Easley et al., 2010). It will be important to understand the circumstances under which TOR promotes either ESC self-renewal and pluripotency or differentiation, and whether different mTOR substrates are involved in mediating these diverse functions of mTOR.
In contrast to ESCs, in which mTOR activity is necessary for self-renewal, adult stem cell self-renewal might actually benefit from reduced levels of mTOR signaling, perhaps because adult stem cells are relatively quiescent. Deletion of tuberous sclerosis protein 1 (TSC1), a negative regulator of mTORC1, leads to overproliferation of HSCs as well as defects in multilineage differentiation and the capacity for bone marrow reconstitution (Chen et al., 2008). Consistently, treatment of old mice with the mTORC1 inhibitor rapamycin confers ‘youthful’ phenotypes to old HSCs and improves the function of their immune system (Chen et al., 2009). Because mTOR signaling increases dramatically in HSCs with age (Chen et al., 2009), mTOR might be a primary effector of HSC aging, although it remains unknown what causes this age-dependent increase in mTOR signaling in this stem cell compartment. The observation that maintaining lower mTOR activity prevents adult stem cell exhaustion is not limited to HSCs. In the mouse epidermis, mTOR activation by chronic Wnt signaling leads to a premature depletion of epidermal stem cells, which manifests itself as premature loss of hair (Castilho et al., 2009). Although maintaining lower TOR activity is beneficial for adult stem cells, a complete loss of mTOR activity is detrimental. Indeed, deletion of Raptor, a key component of the mTORC1 complex, leads to defects in HSC function upon transplantation (Kalaitzidis et al., 2012). Finally, although mTOR appears to affect stem cells mostly by acting in a cell-autonomous manner (Kalaitzidis et al., 2012; Magri et al., 2011), it can also influence adult stem cells (e.g. intestinal stem cells) in a non-cell-autonomous manner by acting on the stem cell niche (Yilmaz et al., 2012).
How does mTOR hyper- or hypoactivation affect stem cell metabolism? mTOR hyperactivation is associated with increased numbers of mitochondria and higher ROS levels in adult HSCs (Chen et al., 2008). Lowering ROS levels by treatment with the antioxidant NAC partially rescues these HSC defects (Chen et al., 2008), which are reminiscent of HSC phenotypes that lack FOXOs (Tothova et al., 2007). Hypoactivation of mTOR owing to loss of Raptor is associated with changes in metabolism, in particular, the lipid and cholesterol metabolism in HSCs (Kalaitzidis et al., 2012). The relative contributions of mTOR targets, such as ribosomal S6 protein kinase 1 (S6K1, also known as RPS6KB1), eukaryotic translation initiation factor 4E-binding protein 1 [(4E-BP1), also known as EIF4EBP1] and SGK1 (Fig. 2), to these metabolic phenotypes in stem cells remain to be determined.
Together, these studies highlight that mTOR signaling is essential for ESC growth and proliferation, yet excessive mTOR activation can be ultimately detrimental for adult stem cell pools and lead to early aging phenotypes. Both mTOR and FOXO are downstream mediators of insulin signaling, which leads to mTOR activation and inhibition of FOXOs. Consistently, FOXOs and mTOR have largely antagonistic functions in stem cells; FOXOs promote the maintenance of stem cells in adulthood and help minimize oxidative stress, whereas overactive mTOR signaling leads to premature stem cell depletion and accumulation of oxidative stress. Collectively, these observations support the idea that a certain degree of cellular quiescence is required to preserve the pool of adult stem cells (Kippin et al., 2005; Morshead et al., 1994). Although stem cell populations are considered ‘immortal’, there might be a limit in the number of times a stem cell can undergo cell division before losing its self-renewal capacity in vivo. These findings also support the idea that high levels of oxidative stress contribute to stem cell dysfunction. Minimizing oxidative damage is likely to be more important in stem cells than in differentiated cells, as stem cells give rise to daughter stem cells and therefore must maintain overall cellular integrity, including genomic, protein and organelle content in order to ensure normal tissue function and avoid tumor development.
AMPK in stem cell mitosis and mitochondrial homeostasis
AMPK is a central ‘fuel gauge’ that is activated by a wide range of stimuli, including low energy or cellular stress. AMPK becomes active when intracellular levels of AMP or ADP are higher than that of ATP, and its activation requires the presence of one of several upstream kinases, including the tumor suppressor liver kinase B1 (LKB1, also known as STK11) and Ca2+-calmodulin-dependent protein kinase kinase β (CAMKKβ) (Kahn et al., 2005) (Fig. 2). AMPK switches off energy-consuming pathways and triggers energy-producing pathways by phosphorylating many substrates that are involved in glucose and lipid metabolism, autophagy and mitophagy, transcription, and cell cycle regulation (Banko et al., 2011; Mihaylova and Shaw, 2011). AMPK has been shown to be crucial for longevity (Greer et al., 2007; Mair et al., 2011), the prevention of type II diabetes (Li et al., 2011) and tumor suppression (Shaw et al., 2004).
Although the importance of AMPK in stem cells has not yet been extensively studied, it is known that AMPK functions in adult HSCs to influence mitochondrial homeostasis. HSCs that lack both catalytic α-subunits of AMPK have decreased levels of ATP and mitochondrial DNA (Nakada et al., 2010). Numbers of HSCs in the bone marrow are substantially reduced several months after AMPK activity is abrogated (Nakada et al., 2010), suggesting that the regulation of mitochondrial homeostasis by AMPK contributes to the control of HSC proliferation. However, AMPK activity is not necessary for HSCs to reconstitute the blood in bone marrow transplantation assays (Nakada et al., 2010). Furthermore, AMPK does not appear to mediate the majority of the actions of its upstream regulator LKB1 in the maintenance of HSCs, suggesting that AMPK-related kinases are important for the function of LKB1 in this context (Gan et al., 2010; Gurumurthy et al., 2010; Nakada et al., 2010). Given that AMPK activity relies on a high ratio between [AMP] and [ATP], the functional relevance of the observed AMPK-dependent phenotypes might depend on the nutrient status of the organism.
AMPK can also regulate stem cell mitosis in the nervous system. For example, in the developing mouse brain, abrogating normal AMPK activity leads to defective mitosis of neural progenitor cells and abnormal brain development (Dasgupta and Milbrandt, 2009). This possible effect is consistent with the recent identification of a role for AMPK substrates in mitosis (Banko et al., 2011). Because AMPK is active under low energy conditions, it appears counterintuitive that it promotes stem cell proliferation given the high-energy demands of cellular division. It is possible that under low energy conditions, AMPK activity ensures the completion of mitosis, because cell cycle arrest at this stage could have disastrous consequences for the genomic stability of a cell. Much still remains to be determined with regard to the roles of AMPK in stem cells, the metabolic conditions under which it is most important, and the substrates that mediate its actions in stem cells.
SIRT1 in the proliferation, differentiation and genomic integrity of stem cells
The protein deacetylase Sirtuin 1 (SIRT1) is one of seven mammalian Class III deacetylases (also called Sirtuins) that become active when the ratio between [NAD+] and [NADH] is high, a state associated with low energy as well as oxidative stress (Imai and Guarente, 2010; Webster et al., 2012). SIRT1 is the Sirtuin with the greatest homology to the yeast Sir2 deacetylase, which is known to extend the replicative lifespan of yeast (Kaeberlein et al., 1999). In mammals, SIRT1 is a crucial regulator of cellular and organismal metabolism (Yu and Auwerx, 2009), improves various markers of health (Bordone et al., 2007; Herranz et al., 2010), provides neuroprotection in a variety of neurodegenerative diseases (Zhang et al., 2011a), and has also been shown to have tumor-suppressive functions in some, but not all, contexts (Fang and Nicholl, 2011).
The effects of SIRT1 perturbation are not identical in all stem cells (Table 1). SIRT1 is highly expressed in human and mouse ESCs, and its expression declines during multilineage differentiation, suggesting that SIRT1 has a more important function in ESCs than in differentiated progeny (Calvanese et al., 2010; Saunders et al., 2010). Yet, under basal conditions, mouse ESCs that lack SIRT1 show no obvious defects (McBurney et al., 2003). However, exposure to mild oxidative stress activates SIRT1 to both promote apoptosis (Chae and Broxmeyer, 2011; Han et al., 2008) and minimize the accumulation of chromosomal abnormalities in mouse ESCs (Oberdoerffer et al., 2008). Together, these studies indicate that SIRT1 carries out anti-tumorigenic activities by regulating nuclear translocation of FOXO and p53, as well as promoting the repair of DNA damage. These findings contrast with previously demonstrated roles for SIRT1 in preventing apoptosis in response to oxidative stress in mouse embryonic fibroblasts, cerebellar granule neurons and human cancer cell lines (Brunet et al., 2004; Motta et al., 2004). It is possible that the ability of SIRT1 to induce or prevent apoptosis depends on the amount and source of oxidative stress present, or the specific cell type (stem versus differentiated).
The function of SIRT1 in HSCs is also age and context-specific. In vitro differentiation of mouse ESCs into the hematopoietic lineage is defective in the absence of SIRT1 (Ou et al., 2011), yet adult mice lacking SIRT1 display virtually normal hematopoiesis (Leko et al., 2012; Narala et al., 2008). It is possible that other histone deacetylases compensate for loss of SIRT1 in adulthood, allowing the blood constituents to form properly after development, even in the absence of SIRT1. Intriguingly, HSC maintenance in old mice is not affected by the deletion of SIRT1 despite increased proliferation levels, suggesting that HSC self-renewal can be maintained in the absence of SIRT1 (Leko et al., 2012; Narala et al., 2008). The mechanisms and specific substrates (histones or non-histone proteins) underlying the ability of SIRT1 to either inhibit or promote the expansion of a cell population in the blood lineage remain to be elucidated.
SIRT1 activity is highly responsive to changes in oxidative state given its dependence on a high [NAD+] to [NADH] ratio, and this has large implications for stem cell fate under varying oxidative conditions. For example, SIRT1 promotes proliferation of adult rat muscle stem cells (Rathbone et al., 2009) and represses their differentiation into myocytes (Fulco et al., 2008; Fulco et al., 2003). Indeed, a reduction in [NAD+]:[NADH], a state inhibitory for SIRT1 activity, is associated with mouse muscle cell differentiation and, by this means, is likely to relieve SIRT1-mediated repression of genes that promote muscle differentiation and maturation (Fulco et al., 2003). In addition, under oxidative conditions, SIRT1 skews the fate of embryonic neural progenitors towards astrocytes at the expense of neurons by repressing transcription of the proneural transcription factor ASCL1 (also known as MASH1) (Prozorovski et al., 2008). Such an activity might be relevant in neurodegenerative conditions that are associated with high levels of oxidative stress, such as amyotrophic lateral sclerosis (ALS), Parkinson's Disease and Alzheimer's Disease (Emerit et al., 2004).
Interestingly, pathways that respond to nutrient availability, such as those involving insulin–FOXO, mTOR, SIRT1 and AMPK, interact with each other and with other factors that are involved in metabolic regulation, forming a ‘signaling and metabolic network’ (Fig. 2). For example, FOXO and the oxygen-sensor HIF1 share common target genes (Renault et al., 2009), and HIF1 translation is regulated by mTOR (Wouters and Koritzinsky, 2008). Additionally, AMPK can activate SIRT1 by upregulating the biosynthesis of NAD+ (Cantó and Auwerx, 2009; Cantó et al., 2009; Cantó et al., 2010; Price et al., 2012). These studies raise the intriguing question of how the activity of metabolic regulators is altered by their interactions with niche constituents, such as oxygen concentration and secreted signaling molecules. Finally, given that FOXO, mTOR, SIRT1 and AMPK all have important functions in the metabolic homeostasis of the whole organism (Gross et al., 2008; Hardie et al., 2012; Laplante and Sabatini, 2012; Satoh et al., 2011), it remains to be explored how the regulation of organismal metabolism by these energy-responsive molecules can have non-cell-autonomous effects on stem cells.
Metabolic regulators in somatic cell reprogramming
The identification of important roles for metabolic regulators, such as FOXOs, mTOR, AMPK and SIRT1, in affecting stem cell fate has motivated the investigation of their influence on the transition from differentiated cell into iPSCs. One could expect that experimental manipulations that promote ESC self-renewal and pluripotency would also enhance reprogramming into iPSCs. However, despite the fact that mTOR is important for maintaining ESC pluripotency and proliferation (Table 1), mTOR inhibitors actually positively affect cell reprogramming (Chen et al., 2011). Conversely, elevating mTOR activity inhibits reprogramming of differentiated cells into iPSCs (He et al., 2012). What are the molecular mechanisms by which mTOR activity regulates cellular reprogramming? Because mTOR is necessary for mitochondrial oxidative function (Cunningham et al., 2007), it is probable that excessive mTOR activity inhibits reprogramming by preventing the switch to a glycolytic metabolism (Menendez et al., 2011). Surprisingly, even though mTOR and AMPK usually act antagonistically, activation of AMPK by metformin or its specific activator A-769662 also represses cellular reprogramming (Vazquez-Martin et al., 2012), potentially by shifting metabolic dependence towards oxidative phosphorylation. These studies contribute to the emerging idea that metabolic state, in particular, the balance between glycolysis and oxidative phosphorylation, crucially impacts the establishment of stem cell characteristics. However, it is still unclear why the activation of either AMPK or mTOR has similar effects on reprogramming, given the normally antagonistic role of these two pathways. More consistent with such opposing functions of mTOR and AMPK, it has been shown that resveratol and fisetin, two compounds known to activate Sirtuins and the AMPK pathway, among other pathways, increase the reprogramming efficiency of mouse embryonic fibroblasts sixfold (Chen et al., 2011), although the mechanism underlying this enhanced reprogramming remains unclear. It is possible that mTOR, AMPK and SIRT1 contribute both to the metabolic changes that occur in the transition between a differentiated cell and a stem cell, and to stem cell properties per se. Thus, understanding the function of these pathways in cellular reprogramming will require more detailed metabolic, gene expression, chromatin and proteomic analyses over the reprogramming time course.
Concluding remarks: implications of fluctuations in organismal metabolism for stem cells
The numerous observations that stem cell properties are affected by energy-responsive molecules and signaling pathways raise questions about the fate of stem cells under conditions when metabolic homeostasis is perturbed (Fig. 3). Studies suggest that abnormal endocrine signaling in organisms with extreme metabolic states has a substantial impact on proliferation and differentiation of multiple stem cell populations throughout the body (Fig. 3). It is also possible that variations in metabolism during gestation could contribute to observable phenotypes in offspring through their effects on stem cells. For example, low energy levels and the associated hormonal signals that occur in the pregnant mother could be directly transmitted to the offspring through the placenta, resulting in transient or permanent changes to stem cells in the embryo (Fig. 3). Further exploration into how stem cells are affected by systemic metabolic states might reveal exciting new roles for metabolism during development, and might also help explain complex diseases and phenotypes, such as obesity, diabetes and aging.
Fig. 3.
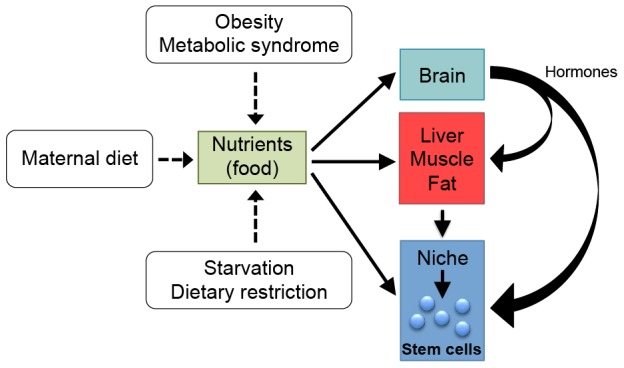
The impact of organismal metabolism on stem cell fate. Schematic diagram of how energy availability and metabolic state of a whole organism can influence stem cells, either directly or indirectly. Nutrient availability can be influenced by food consumption, metabolic disorders, or, in the case of the fetus, maternal diet. Obesity and dietary restriction are two examples of extremes of nutrient availability, the former is closely associated with type II diabetes, a widespread disease of insulin resistance, whereas the latter is associated with health benefits in many species (Fontana et al., 2010). Studies examining how diabetes and other diseases of metabolism alter stem cell function are beginning to emerge. For example, mobilization of HSCs by granulocyte colony-stimulating factor (G-CSF) is impaired in both diabetic human patients and mouse models of type I or type II diabetes (Ferraro et al., 2011). In the brains of rodents with either type I or type II diabetes, NSC proliferation and neurogenesis are reduced in the hippocampus in a corticosterone-dependent manner (Rafalski and Brunet, 2011; Stranahan et al., 2008). Conversely, dietary restriction (30% reduction in calorie intake without malnutrition) enhances the survival of newborn neurons in the adult rodent hippocampus (Lee et al., 2000). These studies highlight how stem cells can respond dramatically to organismal changes in metabolic homeostasis and argue that more studies need to be conducted to characterize not only how stem cell populations are affected by disordered metabolism, but also how pharmaceutical drugs that are used to treat these metabolic conditions affect stem cells. Maternal nutrition also has the potential to impact the fetus through changes in stem cell fate. The Dutch famine during World War II is an example of how starvation in pregnant mothers can result in glucose intolerance, cognitive dysfunctions, and greater risk for breast cancer and heart disease in offspring that were developing embryos during the time of maternal starvation (de Rooij et al., 2010; Roseboom et al., 2006). Although the effects of maternal starvation on stem cells of the fetus are not known, in utero changes in metabolism are likely to have a key function in the regulation of stem cells and the tissues that develop from them.
Supplementary Material
Footnotes
Funding
This work is supported by a National Institute on Aging grant [grant number P01 AG036695 to A.B.]; a California Institute for Regenerative Medicine New Faculty Award; an Ellison Medical Foundation Senior Scholar Award; the Glenn Foundation for Medical Research (to A.B.); a National Institute of Neurological Disorders and Stroke (NINDS) Graduate Fellowship [grant number 5F31NS064600 to V.A.R.]; and a Stanford University Dean's Post-doctoral Fellowship (to E.M.). Deposited in PMC for release after 12 months.
References
- Anselmi C. V., Malovini A., Roncarati R., Novelli V., Villa F., Condorelli G., Bellazzi R., Puca A. A. (2009). Association of the FOXO3A locus with extreme longevity in a southern Italian centenarian study. Rejuvenation Res. 12, 95–104 10.1089/rej.2008.0827 [DOI] [PubMed] [Google Scholar]
- Banko M. R., Allen J. J., Schaffer B. E., Wilker E. W., Tsou P., White J. L., Villén J., Wang B., Kim S. R., Sakamoto K.et al. (2011). Chemical genetic screen for AMPKα2 substrates uncovers a network of proteins involved in mitosis. Mol. Cell 44, 878–892 10.1016/j.molcel.2011.11.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birket M. J., Orr A. L., Gerencser A. A., Madden D. T., Vitelli C., Swistowski A., Brand M. D., Zeng X. (2011). A reduction in ATP demand and mitochondrial activity with neural differentiation of human embryonic stem cells. J. Cell Sci. 124, 348–358 10.1242/jcs.072272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonnert T. P., Bilsland J. G., Guest P. C., Heavens R., McLaren D., Dale C., Thakur M., McAllister G., Munoz–Sanjuan I. (2006). Molecular characterization of adult mouse subventricular zone progenitor cells during the onset of differentiation. Eur. J. Neurosci. 24, 661–675 10.1111/j.1460-9568.2006.04912.x [DOI] [PubMed] [Google Scholar]
- Bordone L., Cohen D., Robinson A., Motta M. C., van Veen E., Czopik A., Steele A. D., Crowe H., Marmor S., Luo J.et al. (2007). SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell 6, 759–767 10.1111/j.1474-9726.2007.00335.x [DOI] [PubMed] [Google Scholar]
- Brunet A., Sweeney L. B., Sturgill J. F., Chua K. F., Greer P. L., Lin Y., Tran H., Ross S. E., Mostoslavsky R., Cohen H. Y.et al. (2004). Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science 303, 2011–2015 10.1126/science.1094637 [DOI] [PubMed] [Google Scholar]
- Calvanese V., Lara E., Suárez–Alvarez B., Abu Dawud R., Vázquez–Chantada M., Martínez–Chantar M. L., Embade N., López–Nieva P., Horrillo A., Hmadcha A.et al. (2010). Sirtuin 1 regulation of developmental genes during differentiation of stem cells. Proc. Natl. Acad. Sci. USA 107, 13736–13741 10.1073/pnas.1001399107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantó C., Auwerx J. (2009). PGC-1alpha, SIRT1 and AMPK, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 20, 98–105 10.1097/MOL.0b013e328328d0a4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantó C., Gerhart–Hines Z., Feige J. N., Lagouge M., Noriega L., Milne J. C., Elliott P. J., Puigserver P., Auwerx J. (2009). AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature 458, 1056–1060 10.1038/nature07813 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantó C., Jiang L. Q., Deshmukh A. S., Mataki C., Coste A., Lagouge M., Zierath J. R., Auwerx J. (2010). Interdependence of AMPK and SIRT1 for metabolic adaptation to fasting and exercise in skeletal muscle. Cell Metab. 11, 213–219 10.1016/j.cmet.2010.02.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castilho R. M., Squarize C. H., Chodosh L. A., Williams B. O., Gutkind J. S. (2009). mTOR mediates Wnt-induced epidermal stem cell exhaustion and aging. Cell Stem Cell 5, 279–289 10.1016/j.stem.2009.06.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chae H. D., Broxmeyer H. E. (2011). SIRT1 deficiency downregulates PTEN/JNK/FOXO1 pathway to block reactive oxygen species-induced apoptosis in mouse embryonic stem cells. Stem Cells Dev. 20, 1277–1285 10.1089/scd.2010.0465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen H. L., Pistollato F., Hoeppner D. J., Ni H. T., McKay R. D., Panchision D. M. (2007). Oxygen tension regulates survival and fate of mouse central nervous system precursors at multiple levels. Stem Cells 25, 2291–2301 10.1634/stemcells.2006-0609 [DOI] [PubMed] [Google Scholar]
- Chen C., Liu Y., Liu R., Ikenoue T., Guan K. L., Liu Y., Zheng P. (2008). TSC-mTOR maintains quiescence and function of hematopoietic stem cells by repressing mitochondrial biogenesis and reactive oxygen species. J. Exp. Med. 205, 2397–2408 10.1084/jem.20081297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C., Liu Y., Liu Y., Zheng P. (2009). mTOR regulation and therapeutic rejuvenation of aging hematopoietic stem cells. Sci. Signal. 2, ra75 10.1126/scisignal.2000559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen T., Shen L., Yu J., Wan H., Guo A., Chen J., Long Y., Zhao J., Pei G. (2011). Rapamycin and other longevity-promoting compounds enhance the generation of mouse induced pluripotent stem cells. Aging Cell 10, 908–911 10.1111/j.1474-9726.2011.00722.x [DOI] [PubMed] [Google Scholar]
- Cho Y. M., Kwon S., Pak Y. K., Seol H. W., Choi Y. M., Park J., Park K. S., Lee H. K. (2006). Dynamic changes in mitochondrial biogenesis and antioxidant enzymes during the spontaneous differentiation of human embryonic stem cells. Biochem. Biophys. Res. Commun. 348, 1472–1478 10.1016/j.bbrc.2006.08.020 [DOI] [PubMed] [Google Scholar]
- Covello K. L., Kehler J., Yu H., Gordan J. D., Arsham A. M., Hu C. J., Labosky P. A., Simon M. C., Keith B. (2006). HIF-2alpha regulates Oct-4: effects of hypoxia on stem cell function, embryonic development, and tumor growth. Genes Dev. 20, 557–570 10.1101/gad.1399906 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cunningham J. T., Rodgers J. T., Arlow D. H., Vazquez F., Mootha V. K., Puigserver P. (2007). mTOR controls mitochondrial oxidative function through a YY1-PGC-1alpha transcriptional complex. Nature 450, 736–740 10.1038/nature06322 [DOI] [PubMed] [Google Scholar]
- Dansen T. B., Burgering B. M. (2008). Unravelling the tumor-suppressive functions of FOXO proteins. Trends Cell Biol. 18, 421–429 10.1016/j.tcb.2008.07.004 [DOI] [PubMed] [Google Scholar]
- Dasgupta B., Milbrandt J. (2009). AMP-activated protein kinase phosphorylates retinoblastoma protein to control mammalian brain development. Dev. Cell 16, 256–270 10.1016/j.devcel.2009.01.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Rooij S. R., Wouters H., Yonker J. E., Painter R. C., Roseboom T. J. (2010). Prenatal undernutrition and cognitive function in late adulthood. Proc. Natl. Acad. Sci. USA 107, 16881–16886 10.1073/pnas.1009459107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Easley C. A., 4th, Ben–Yehudah A., Redinger C. J., Oliver S. L., Varum S. T., Eisinger V. M., Carlisle D. L., Donovan P. J., Schatten G. P. (2010). mTOR-mediated activation of p70 S6K induces differentiation of pluripotent human embryonic stem cells. Cell Reprogram. 12, 263–273 10.1089/cell.2010.0011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eliasson P., Jönsson J. I. (2010). The hematopoietic stem cell niche: low in oxygen but a nice place to be. J. Cell. Physiol. 222, 17–22 10.1002/jcp.21908 [DOI] [PubMed] [Google Scholar]
- Emerit J., Edeas M., Bricaire F. (2004). Neurodegenerative diseases and oxidative stress. Biomed. Pharmacother. 58, 39–46 10.1016/j.biopha.2003.11.004 [DOI] [PubMed] [Google Scholar]
- Ezashi T., Das P., Roberts R. M. (2005). Low O2 tensions and the prevention of differentiation of hES cells. Proc. Natl. Acad. Sci. USA 102, 4783–4788 10.1073/pnas.0501283102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Facucho–Oliveira J. M., Alderson J., Spikings E. C., Egginton S., St John J. C. (2007). Mitochondrial DNA replication during differentiation of murine embryonic stem cells. J. Cell Sci. 120, 4025–4034 10.1242/jcs.016972 [DOI] [PubMed] [Google Scholar]
- Fang Y., Nicholl M. B. (2011). Sirtuin 1 in malignant transformation: friend or foe? Cancer Lett. 306, 10–14 10.1016/j.canlet.2011.02.019 [DOI] [PubMed] [Google Scholar]
- Feng B., Ng J. H., Heng J. C., Ng H. H. (2009). Molecules that promote or enhance reprogramming of somatic cells to induced pluripotent stem cells. Cell Stem Cell 4, 301–312 10.1016/j.stem.2009.03.005 [DOI] [PubMed] [Google Scholar]
- Ferraro F., Lymperi S., Mendez–Ferrer S., Saez B., Spencer J. A., Yeap B. Y., Masselli E., Graiani G., Prezioso L., Rizzini E. L.et al. (2011). Diabetes impairs hematopoietic stem cell mobilization by altering niche function. Sci. Transl. Med. 3, 104ra101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer B., Bavister B. D. (1993). Oxygen tension in the oviduct and uterus of rhesus monkeys, hamsters and rabbits. J. Reprod. Fertil. 99, 673–679 10.1530/jrf.0.0990673 [DOI] [PubMed] [Google Scholar]
- Flachsbart F., Caliebe A., Kleindorp R., Blanché H., von Eller–Eberstein H., Nikolaus S., Schreiber S., Nebel A. (2009). Association of FOXO3A variation with human longevity confirmed in German centenarians. Proc. Natl. Acad. Sci. USA 106, 2700–2705 10.1073/pnas.0809594106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folmes C. D., Nelson T. J., Martinez–Fernandez A., Arrell D. K., Lindor J. Z., Dzeja P. P., Ikeda Y., Perez–Terzic C., Terzic A. (2011). Somatic oxidative bioenergetics transitions into pluripotency-dependent glycolysis to facilitate nuclear reprogramming. Cell Metab. 14, 264–271 10.1016/j.cmet.2011.06.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fontana L., Partridge L., Longo V. D. (2010). Extending healthy life span–from yeast to humans. Science 328, 321–326 10.1126/science.1172539 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fulco M., Schiltz R. L., Iezzi S., King M. T., Zhao P., Kashiwaya Y., Hoffman E., Veech R. L., Sartorelli V. (2003). Sir2 regulates skeletal muscle differentiation as a potential sensor of the redox state. Mol. Cell 12, 51–62 10.1016/S1097-2765(03)00226-0 [DOI] [PubMed] [Google Scholar]
- Fulco M., Cen Y., Zhao P., Hoffman E. P., McBurney M. W., Sauve A. A., Sartorelli V. (2008). Glucose restriction inhibits skeletal myoblast differentiation by activating SIRT1 through AMPK-mediated regulation of Nampt. Dev. Cell 14, 661–673 10.1016/j.devcel.2008.02.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gan B., Hu J., Jiang S., Liu Y., Sahin E., Zhuang L., Fletcher–Sananikone E., Colla S., Wang Y. A., Chin L.et al. (2010). Lkb1 regulates quiescence and metabolic homeostasis of haematopoietic stem cells. Nature 468, 701–704 10.1038/nature09595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goertz M. J., Wu Z., Gallardo T. D., Hamra F. K., Castrillon D. H. (2011). Foxo1 is required in mouse spermatogonial stem cells for their maintenance and the initiation of spermatogenesis. J. Clin. Invest. 121, 3456–3466 10.1172/JCI57984 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greer E. L., Brunet A. (2005). FOXO transcription factors at the interface between longevity and tumor suppression. Oncogene 24, 7410–7425 10.1038/sj.onc.1209086 [DOI] [PubMed] [Google Scholar]
- Greer E. L., Brunet A. (2008). Signaling networks in aging. J. Cell Sci. 121, 407–412 10.1242/jcs.021519 [DOI] [PubMed] [Google Scholar]
- Greer E. L., Dowlatshahi D., Banko M. R., Villen J., Hoang K., Blanchard D., Gygi S. P., Brunet A. (2007). An AMPK-FOXO pathway mediates longevity induced by a novel method of dietary restriction in C. elegans. Curr. Biol. 17, 1646–1656 10.1016/j.cub.2007.08.047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross D. N., van den Heuvel A. P., Birnbaum M. J. (2008). The role of FoxO in the regulation of metabolism. Oncogene 27, 2320–2336 10.1038/onc.2008.25 [DOI] [PubMed] [Google Scholar]
- Grskovic M., Javaherian A., Strulovici B., Daley G. Q. (2011). Induced pluripotent stem cells–opportunities for disease modelling and drug discovery. Nat. Rev. Drug Discov. 10, 915–929 [DOI] [PubMed] [Google Scholar]
- Gurumurthy S., Xie S. Z., Alagesan B., Kim J., Yusuf R. Z., Saez B., Tzatsos A., Ozsolak F., Milos P., Ferrari F.et al. (2010). The Lkb1 metabolic sensor maintains haematopoietic stem cell survival. Nature 468, 659–663 10.1038/nature09572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han M. K., Song E. K., Guo Y., Ou X., Mantel C., Broxmeyer H. E. (2008). SIRT1 regulates apoptosis and Nanog expression in mouse embryonic stem cells by controlling p53 subcellular localization. Cell Stem Cell 2, 241–251 10.1016/j.stem.2008.01.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hardie D. G., Ross F. A., Hawley S. A. (2012). AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 13, 251–262 10.1038/nrm3311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison D. E., Strong R., Sharp Z. D., Nelson J. F., Astle C. M., Flurkey K., Nadon N. L., Wilkinson J. E., Frenkel K., Carter C. S.et al. (2009). Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 460, 392–395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He J., Kang L., Wu T., Zhang J., Wang H., Gao H., Zhang Y., Huang B., Liu W., Kou Z.et al. (2012). An elaborate regulation of Mammalian target of rapamycin activity is required for somatic cell reprogramming induced by defined transcription factors. Stem Cells Dev. 21, 2630–2641 10.1089/scd.2012.0015 [DOI] [PubMed] [Google Scholar]
- Herranz D., Muñoz–Martin M., Cañamero M., Mulero F., Martinez–Pastor B., Fernandez–Capetillo O., Serrano M. (2010). Sirt1 improves healthy ageing and protects from metabolic syndrome-associated cancer. Nat Commun. 1, 3 10.1038/ncomms1001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu P. P., Sabatini D. M. (2008). Cancer cell metabolism: Warburg and beyond. Cell 134, 703–707 10.1016/j.cell.2008.08.021 [DOI] [PubMed] [Google Scholar]
- Imai S., Guarente L. (2010). Ten years of NAD-dependent SIR2 family deacetylases: implications for metabolic diseases. Trends Pharmacol. Sci. 31, 212–220 10.1016/j.tips.2010.02.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito K., Hirao A., Arai F., Matsuoka S., Takubo K., Hamaguchi I., Nomiyama K., Hosokawa K., Sakurada K., Nakagata N.et al. (2004). Regulation of oxidative stress by ATM is required for self-renewal of haematopoietic stem cells. Nature 431, 997–1002 10.1038/nature02989 [DOI] [PubMed] [Google Scholar]
- Ito K., Carracedo A., Weiss D., Arai F., Ala U., Avigan D. E., Schafer Z. T., Evans R. M., Suda T., Lee C. H.et al. (2012). A PML-PPARdelta pathway for fatty acid oxidation regulates hematopoietic stem cell maintenance. Nat. Med. 18, 1350–1358 10.1038/nm.2882 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jasper H., Jones D. L. (2010). Metabolic regulation of stem cell behavior and implications for aging. Cell Metab. 12, 561–565 10.1016/j.cmet.2010.11.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joseph N. M., Morrison S. J. (2005). Toward an understanding of the physiological function of Mammalian stem cells. Dev. Cell 9, 173–183 10.1016/j.devcel.2005.07.001 [DOI] [PubMed] [Google Scholar]
- Kaeberlein M., McVey M., Guarente L. (1999). The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes Dev. 13, 2570–2580 10.1101/gad.13.19.2570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahn B. B., Alquier T., Carling D., Hardie D. G. (2005). AMP-activated protein kinase: ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 1, 15–25 10.1016/j.cmet.2004.12.003 [DOI] [PubMed] [Google Scholar]
- Kalaitzidis D., Sykes S. M., Wang Z., Punt N., Tang Y., Ragu C., Sinha A. U., Lane S. W., Souza A. L., Clish C. B.et al. (2012). mTOR complex 1 plays critical roles in hematopoiesis and Pten-loss-evoked leukemogenesis. Cell Stem Cell 11, 429–439 10.1016/j.stem.2012.06.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang J., Shakya A., Tantin D. (2009). Stem cells, stress, metabolism and cancer: a drama in two Octs. Trends Biochem. Sci. 34, 491–499 10.1016/j.tibs.2009.06.003 [DOI] [PubMed] [Google Scholar]
- Kapahi P., Chen D., Rogers A. N., Katewa S. D., Li P. W., Thomas E. L., Kockel L. (2010). With TOR, less is more: a key role for the conserved nutrient-sensing TOR pathway in aging. Cell Metab. 11, 453–465 10.1016/j.cmet.2010.05.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenyon C. J. (2010). The genetics of ageing. Nature 464, 504–512 10.1038/nature08980 [DOI] [PubMed] [Google Scholar]
- Kippin T. E., Martens D. J., van der Kooy D. (2005). p21 loss compromises the relative quiescence of forebrain stem cell proliferation leading to exhaustion of their proliferation capacity. Genes Dev. 19, 756–767 10.1101/gad.1272305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knobloch M., Braun S. M., Zurkirchen L., von Schoultz C., Zamboni N., Arauzo–Bravo M. J., Kovacs W. J., Karalay O., Suter U., Machado R. A.et al. (2013). Metabolic control of adult neural stem cell activity by Fasn-dependent lipogenesis. Nature 493, 226–230 10.1038/nature11689 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi C. I., Suda T. (2012). Regulation of reactive oxygen species in stem cells and cancer stem cells. J. Cell. Physiol. 227, 421–430 10.1002/jcp.22764 [DOI] [PubMed] [Google Scholar]
- Laplante M., Sabatini D. M. (2012). mTOR signaling in growth control and disease. Cell 149, 274–293 10.1016/j.cell.2012.03.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J., Duan W., Long J. M., Ingram D. K., Mattson M. P. (2000). Dietary restriction increases the number of newly generated neural cells, and induces BDNF expression, in the dentate gyrus of rats. J. Mol. Neurosci. 15, 99–108 10.1385/JMN:15:2:99 [DOI] [PubMed] [Google Scholar]
- Leko V., Varnum–Finney B., Li H., Gu Y., Flowers D., Nourigat C., Bernstein I. D., Bedalov A. (2012). SIRT1 is dispensable for function of hematopoietic stem cells in adult mice. Blood 119, 1856–1860 10.1182/blood-2011-09-377077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y., Xu S., Mihaylova M. M., Zheng B., Hou X., Jiang B., Park O., Luo Z., Lefai E., Shyy J. Y.et al. (2011). AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 13, 376–388 10.1016/j.cmet.2011.03.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magri L., Cambiaghi M., Cominelli M., Alfaro–Cervello C., Cursi M., Pala M., Bulfone A., Garcìa–Verdugo J. M., Leocani L., Minicucci F.et al. (2011). Sustained activation of mTOR pathway in embryonic neural stem cells leads to development of tuberous sclerosis complex-associated lesions. Cell Stem Cell 9, 447–462 10.1016/j.stem.2011.09.008 [DOI] [PubMed] [Google Scholar]
- Mair W., Morantte I., Rodrigues A. P., Manning G., Montminy M., Shaw R. J., Dillin A. (2011). Lifespan extension induced by AMPK and calcineurin is mediated by CRTC-1 and CREB. Nature 470, 404–408 10.1038/nature09706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mannello F., Medda V., Tonti G. A. (2011). Hypoxia and neural stem cells: from invertebrates to brain cancer stem cells. Int. J. Dev. Biol. 55, 569–581 10.1387/ijdb.103186fm [DOI] [PubMed] [Google Scholar]
- Mathieu J., Zhang Z., Zhou W., Wang A. J., Heddleston J. M., Pinna C. M., Hubaud A., Stadler B., Choi M., Bar M.et al. (2011). HIF induces human embryonic stem cell markers in cancer cells. Cancer Res. 71, 4640–4652 10.1158/0008-5472.CAN-10-3320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McBurney M. W., Yang X., Jardine K., Bieman M., Th'ng J., Lemieux M. (2003). The absence of SIR2alpha protein has no effect on global gene silencing in mouse embryonic stem cells. Mol. Cancer Res. 1, 402–409 [PubMed] [Google Scholar]
- Menendez J. A., Vellon L., Oliveras–Ferraros C., Cufí S., Vazquez–Martin A. (2011). mTOR-regulated senescence and autophagy during reprogramming of somatic cells to pluripotency: a roadmap from energy metabolism to stem cell renewal and aging. Cell Cycle 10, 3658–3677 10.4161/cc.10.21.18128 [DOI] [PubMed] [Google Scholar]
- Miharada K., Karlsson G., Rehn M., Rörby E., Siva K., Cammenga J., Karlsson S. (2011). Cripto regulates hematopoietic stem cells as a hypoxic-niche-related factor through cell surface receptor GRP78. Cell Stem Cell 9, 330–344 10.1016/j.stem.2011.07.016 [DOI] [PubMed] [Google Scholar]
- Mihaylova M. M., Shaw R. J. (2011). The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 13, 1016–1023 10.1038/ncb2329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyamoto K., Araki K. Y., Naka K., Arai F., Takubo K., Yamazaki S., Matsuoka S., Miyamoto T., Ito K., Ohmura M.et al. (2007). Foxo3a is essential for maintenance of the hematopoietic stem cell pool. Cell Stem Cell 1, 101–112 10.1016/j.stem.2007.02.001 [DOI] [PubMed] [Google Scholar]
- Mohyeldin A., Garzón–Muvdi T., Quiñones–Hinojosa A. (2010). Oxygen in stem cell biology: a critical component of the stem cell niche. Cell Stem Cell 7, 150–161 10.1016/j.stem.2010.07.007 [DOI] [PubMed] [Google Scholar]
- Morshead C. M., Reynolds B. A., Craig C. G., McBurney M. W., Staines W. A., Morassutti D., Weiss S., van der Kooy D. (1994). Neural stem cells in the adult mammalian forebrain: a relatively quiescent subpopulation of subependymal cells. Neuron 13, 1071–1082 10.1016/0896-6273(94)90046-9 [DOI] [PubMed] [Google Scholar]
- Motta M. C., Divecha N., Lemieux M., Kamel C., Chen D., Gu W., Bultsma Y., McBurney M., Guarente L. (2004). Mammalian SIRT1 represses forkhead transcription factors. Cell 116, 551–563 10.1016/S0092-8674(04)00126-6 [DOI] [PubMed] [Google Scholar]
- Murakami M., Ichisaka T., Maeda M., Oshiro N., Hara K., Edenhofer F., Kiyama H., Yonezawa K., Yamanaka S. (2004). mTOR is essential for growth and proliferation in early mouse embryos and embryonic stem cells. Mol. Cell. Biol. 24, 6710–6718 10.1128/MCB.24.15.6710-6718.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakada D., Saunders T. L., Morrison S. J. (2010). Lkb1 regulates cell cycle and energy metabolism in haematopoietic stem cells. Nature 468, 653–658 10.1038/nature09571 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakada D., Levi B. P., Morrison S. J. (2011). Integrating physiological regulation with stem cell and tissue homeostasis. Neuron 70, 703–718 10.1016/j.neuron.2011.05.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narala S. R., Allsopp R. C., Wells T. B., Zhang G., Prasad P., Coussens M. J., Rossi D. J., Weissman I. L., Vaziri H. (2008). SIRT1 acts as a nutrient-sensitive growth suppressor and its loss is associated with increased AMPK and telomerase activity. Mol. Biol. Cell 19, 1210–1219 10.1091/mbc.E07-09-0965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oberdoerffer P., Michan S., McVay M., Mostoslavsky R., Vann J., Park S. K., Hartlerode A., Stegmuller J., Hafner A., Loerch P.et al. (2008). SIRT1 redistribution on chromatin promotes genomic stability but alters gene expression during aging. Cell 135, 907–918 10.1016/j.cell.2008.10.025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okita K., Ichisaka T., Yamanaka S. (2007). Generation of germline-competent induced pluripotent stem cells. Nature 448, 313–317 10.1038/nature05934 [DOI] [PubMed] [Google Scholar]
- Ou X., Chae H. D., Wang R. H., Shelley W. C., Cooper S., Taylor T., Kim Y. J., Deng C. X., Yoder M. C., Broxmeyer H. E. (2011). SIRT1 deficiency compromises mouse embryonic stem cell hematopoietic differentiation, and embryonic and adult hematopoiesis in the mouse. Blood 117, 440–450 10.1182/blood-2010-03-273011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paik J. H., Ding Z., Narurkar R., Ramkissoon S., Muller F., Kamoun W. S., Chae S. S., Zheng H., Ying H., Mahoney J.et al. (2009). FoxOs cooperatively regulate diverse pathways governing neural stem cell homeostasis. Cell Stem Cell 5, 540–553 10.1016/j.stem.2009.09.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panopoulos A. D., Yanes O., Ruiz S., Kida Y. S., Diep D., Tautenhahn R., Herrerías A., Batchelder E. M., Plongthongkum N., Lutz M.et al. (2012). The metabolome of induced pluripotent stem cells reveals metabolic changes occurring in somatic cell reprogramming. Cell Res. 22, 168–177 10.1038/cr.2011.177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pawlikowska L., Hu D., Huntsman S., Sung A., Chu C., Chen J., Joyner A. H., Schork N. J., Hsueh W. C., Reiner A. P., et al. (2009). Association of common genetic variation in the insulin/IGF1 signaling pathway with human longevity. Aging Cell 8, 460–472 10.1111/j.1474-9726.2009.00493.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pistollato F., Chen H. L., Schwartz P. H., Basso G., Panchision D. M. (2007). Oxygen tension controls the expansion of human CNS precursors and the generation of astrocytes and oligodendrocytes. Mol. Cell. Neurosci. 35, 424–435 10.1016/j.mcn.2007.04.003 [DOI] [PubMed] [Google Scholar]
- Price N. L., Gomes A. P., Ling A. J., Duarte F. V., Martin–Montalvo A., North B. J., Agarwal B., Ye L., Ramadori G., Teodoro J. S.et al. (2012). SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 15, 675–690 10.1016/j.cmet.2012.04.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prozorovski T., Schulze–Topphoff U., Glumm R., Baumgart J., Schröter F., Ninnemann O., Siegert E., Bendix I., Brüstle O., Nitsch R.et al. (2008). Sirt1 contributes critically to the redox-dependent fate of neural progenitors. Nat. Cell Biol. 10, 385–394 10.1038/ncb1700 [DOI] [PubMed] [Google Scholar]
- Rafalski V. A., Brunet A. (2011). Energy metabolism in adult neural stem cell fate. Prog. Neurobiol. 93, 182–203 10.1016/j.pneurobio.2010.10.007 [DOI] [PubMed] [Google Scholar]
- Ramalho–Santos M., Yoon S., Matsuzaki Y., Mulligan R. C., Melton D. A. (2002). “Stemness”: transcriptional profiling of embryonic and adult stem cells. Science 298, 597–600 10.1126/science.1072530 [DOI] [PubMed] [Google Scholar]
- Rando T. A. (2006). Stem cells, ageing and the quest for immortality. Nature 441, 1080–1086 10.1038/nature04958 [DOI] [PubMed] [Google Scholar]
- Rathbone C. R., Booth F. W., Lees S. J. (2009). Sirt1 increases skeletal muscle precursor cell proliferation. Eur. J. Cell Biol. 88, 35–44 10.1016/j.ejcb.2008.08.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Renault V. M., Rafalski V. A., Morgan A. A., Salih D. A., Brett J. O., Webb A. E., Villeda S. A., Thekkat P. U., Guillerey C., Denko N. C.et al. (2009). FoxO3 regulates neural stem cell homeostasis. Cell Stem Cell 5, 527–539 10.1016/j.stem.2009.09.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roseboom T., de Rooij S., Painter R. (2006). The Dutch famine and its long-term consequences for adult health. Early Hum. Dev. 82, 485–491 10.1016/j.earlhumdev.2006.07.001 [DOI] [PubMed] [Google Scholar]
- Rubakhin S. S., Romanova E. V., Nemes P., Sweedler J. V. (2011). Profiling metabolites and peptides in single cells. Nat. Methods 8 Suppl., S20–S29 10.1038/nmeth.1549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salih D. A., Brunet A. (2008). FoxO transcription factors in the maintenance of cellular homeostasis during aging. Curr. Opin. Cell Biol. 20, 126–136 10.1016/j.ceb.2008.02.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sampath P., Pritchard D. K., Pabon L., Reinecke H., Schwartz S. M., Morris D. R., Murry C. E. (2008). A hierarchical network controls protein translation during murine embryonic stem cell self-renewal and differentiation. Cell Stem Cell 2, 448–460 10.1016/j.stem.2008.03.013 [DOI] [PubMed] [Google Scholar]
- Satoh A., Stein L., Imai S. (2011). The role of mammalian sirtuins in the regulation of metabolism, aging, and longevity. Handb. Exp. Pharmacol. 206, 125–162 10.1007/978-3-642-21631-2_7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saunders L. R., Sharma A. D., Tawney J., Nakagawa M., Okita K., Yamanaka S., Willenbring H., Verdin E. (2010). miRNAs regulate SIRT1 expression during mouse embryonic stem cell differentiation and in adult mouse tissues. Aging (Albany NY) 2, 415–431 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schieke S. M., Ma M., Cao L., McCoy J. P., Jr, Liu C., Hensel N. F., Barrett A. J., Boehm M., Finkel T. (2008). Mitochondrial metabolism modulates differentiation and teratoma formation capacity in mouse embryonic stem cells. J. Biol. Chem. 283, 28506–28512 10.1074/jbc.M802763200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shakya A., Cooksey R., Cox J. E., Wang V., McClain D. A., Tantin D. (2009). Oct1 loss of function induces a coordinate metabolic shift that opposes tumorigenicity. Nat. Cell Biol. 11, 320–327 10.1038/ncb1840 [DOI] [PubMed] [Google Scholar]
- Shaw R. J., Bardeesy N., Manning B. D., Lopez L., Kosmatka M., DePinho R. A., Cantley L. C. (2004). The LKB1 tumor suppressor negatively regulates mTOR signaling. Cancer Cell 6, 91–99 10.1016/j.ccr.2004.06.007 [DOI] [PubMed] [Google Scholar]
- Shyh–Chang N., Locasale J. W., Lyssiotis C. A., Zheng Y., Teo R., Ratanasirintrawoot S., Zhang J., Onder T., Unternaehrer J. J., Zhu H.et al. (2013). Influence of Threonine Metabolism on S-Adenosylmethionine and Histone Methylation. Science 339, 222–226 10.1126/science.1226603 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silver I., Erecińska M. (1998). Oxygen and ion concentrations in normoxic and hypoxic brain cells. Adv. Exp. Med. Biol. 454, 7–16 10.1007/978-1-4615-4863-8_2 [DOI] [PubMed] [Google Scholar]
- Simsek T., Kocabas F., Zheng J., Deberardinis R. J., Mahmoud A. I., Olson E. N., Schneider J. W., Zhang C. C., Sadek H. A. (2010). The distinct metabolic profile of hematopoietic stem cells reflects their location in a hypoxic niche. Cell Stem Cell 7, 380–390 10.1016/j.stem.2010.07.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- St John J. C., Ramalho–Santos J., Gray H. L., Petrosko P., Rawe V. Y., Navara C. S., Simerly C. R., Schatten G. P. (2005). The expression of mitochondrial DNA transcription factors during early cardiomyocyte in vitro differentiation from human embryonic stem cells. Cloning Stem Cells 7, 141–153 10.1089/clo.2005.7.141 [DOI] [PubMed] [Google Scholar]
- Steinhauser M. L., Bailey A. P., Senyo S. E., Guillermier C., Perlstein T. S., Gould A. P., Lee R. T., Lechene C. P. (2012). Multi-isotope imaging mass spectrometry quantifies stem cell division and metabolism. Nature 481, 516–519 10.1038/nature10734 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stranahan A. M., Arumugam T. V., Cutler R. G., Lee K., Egan J. M., Mattson M. P. (2008). Diabetes impairs hippocampal function through glucocorticoid-mediated effects on new and mature neurons. Nat. Neurosci. 11, 309–317 10.1038/nn2055 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Studer L., Csete M., Lee S. H., Kabbani N., Walikonis J., Wold B., McKay R. (2000). Enhanced proliferation, survival, and dopaminergic differentiation of CNS precursors in lowered oxygen. J. Neurosci. 20, 7377–7383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suda T., Takubo K., Semenza G. L. (2011). Metabolic regulation of hematopoietic stem cells in the hypoxic niche. Cell Stem Cell 9, 298–310 10.1016/j.stem.2011.09.010 [DOI] [PubMed] [Google Scholar]
- Suhr S. T., Chang E. A., Tjong J., Alcasid N., Perkins G. A., Goissis M. D., Ellisman M. H., Perez G. I., Cibelli J. B. (2010). Mitochondrial rejuvenation after induced pluripotency. PLoS ONE 5, e14095 10.1371/journal.pone.0014095 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi K., Yamanaka S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663–676 10.1016/j.cell.2006.07.024 [DOI] [PubMed] [Google Scholar]
- Takahashi K., Tanabe K., Ohnuki M., Narita M., Ichisaka T., Tomoda K., Yamanaka S. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131, 861–872 10.1016/j.cell.2007.11.019 [DOI] [PubMed] [Google Scholar]
- Takubo K., Goda N., Yamada W., Iriuchishima H., Ikeda E., Kubota Y., Shima H., Johnson R. S., Hirao A., Suematsu M.et al. (2010). Regulation of the HIF-1alpha level is essential for hematopoietic stem cells. Cell Stem Cell 7, 391–402 10.1016/j.stem.2010.06.020 [DOI] [PubMed] [Google Scholar]
- Thomson J. A., Itskovitz–Eldor J., Shapiro S. S., Waknitz M. A., Swiergiel J. J., Marshall V. S., Jones J. M. (1998). Embryonic stem cell lines derived from human blastocysts. Science 282, 1145–1147 10.1126/science.282.5391.1145 [DOI] [PubMed] [Google Scholar]
- Tomita S., Ueno M., Sakamoto M., Kitahama Y., Ueki M., Maekawa N., Sakamoto H., Gassmann M., Kageyama R., Ueda N.et al. (2003). Defective brain development in mice lacking the Hif-1alpha gene in neural cells. Mol. Cell. Biol. 23, 6739–6749 10.1128/MCB.23.19.6739-6749.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tothova Z., Kollipara R., Huntly B. J., Lee B. H., Castrillon D. H., Cullen D. E., McDowell E. P., Lazo–Kallanian S., Williams I. R., Sears C.et al. (2007). FoxOs are critical mediators of hematopoietic stem cell resistance to physiologic oxidative stress. Cell 128, 325–339 10.1016/j.cell.2007.01.003 [DOI] [PubMed] [Google Scholar]
- Unwin R. D., Smith D. L., Blinco D., Wilson C. L., Miller C. J., Evans C. A., Jaworska E., Baldwin S. A., Barnes K., Pierce A.et al. (2006). Quantitative proteomics reveals posttranslational control as a regulatory factor in primary hematopoietic stem cells. Blood 107, 4687–4694 10.1182/blood-2005-12-4995 [DOI] [PubMed] [Google Scholar]
- Varum S., Momcilović O., Castro C., Ben–Yehudah A., Ramalho–Santos J., Navara C. S. (2009). Enhancement of human embryonic stem cell pluripotency through inhibition of the mitochondrial respiratory chain. Stem Cell Res. 3, 142–156 10.1016/j.scr.2009.07.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varum S., Rodrigues A. S., Moura M. B., Momcilovic O., Easley C. A., 4th, Ramalho–Santos J., Van Houten B., Schatten G. (2011). Energy metabolism in human pluripotent stem cells and their differentiated counterparts. PLoS ONE 6, e20914 10.1371/journal.pone.0020914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vazquez–Martin A., Vellon L., Quiros P. M., Cufi S., Ruiz de Galarreta E., Oliveras–Ferraros C., Martin A. G., Martin–Castillo B., Lopez–Otin C., Menendez J. A. (2012). Activation of AMP-activated protein kinase (AMPK) provides a metabolic barrier to reprogramming somatic cells into stem cells. Cell Cycle 11, 974–989 [DOI] [PubMed] [Google Scholar]
- Vilchez D., Boyer L., Morantte I., Lutz M., Merkwirth C., Joyce D., Spencer B., Page L., Masliah E., Berggren W. T.et al. (2012). Increased proteasome activity in human embryonic stem cells is regulated by PSMD11. Nature 489, 304–308 10.1038/nature11468 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang P., Na J. (2011). Mechanism and methods to induce pluripotency. Protein Cell 2, 792–799 10.1007/s13238-011-1107-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Alexander P., Wu L., Hammer R., Cleaver O., McKnight S. L. (2009). Dependence of mouse embryonic stem cells on threonine catabolism. Science 325, 435–439 10.1126/science.1173288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Alexander P., McKnight S. L. (2011). Metabolic specialization of mouse embryonic stem cells. Cold Spring Harb. Symp. Quant. Biol. 76, 183–193 10.1101/sqb.2011.76.010835 [DOI] [PubMed] [Google Scholar]
- Warburg O. (1956). On the origin of cancer cells. Science 123, 309–314 10.1126/science.123.3191.309 [DOI] [PubMed] [Google Scholar]
- Webster B. R., Lu Z., Sack M. N., Scott I. (2012). The role of sirtuins in modulating redox stressors. Free Radic. Biol. Med. 52, 281–290 10.1016/j.freeradbiomed.2011.10.484 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willcox B. J., Donlon T. A., He Q., Chen R., Grove J. S., Yano K., Masaki K. H., Willcox D. C., Rodriguez B., Curb J. D. (2008). FOXO3A genotype is strongly associated with human longevity. Proc. Natl. Acad. Sci. USA 105, 13987–13992 10.1073/pnas.0801030105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wouters B. G., Koritzinsky M. (2008). Hypoxia signalling through mTOR and the unfolded protein response in cancer. Nat. Rev. Cancer 8, 851–864 10.1038/nrc2501 [DOI] [PubMed] [Google Scholar]
- Yalcin S., Zhang X., Luciano J. P., Mungamuri S. K., Marinkovic D., Vercherat C., Sarkar A., Grisotto M., Taneja R., Ghaffari S. (2008). Foxo3 is essential for the regulation of ataxia telangiectasia mutated and oxidative stress-mediated homeostasis of hematopoietic stem cells. J. Biol. Chem. 283, 25692–25705 10.1074/jbc.M800517200 [DOI] [PubMed] [Google Scholar]
- Yalcin S., Marinkovic D., Mungamuri S. K., Zhang X., Tong W., Sellers R., Ghaffari S. (2010). ROS-mediated amplification of AKT/mTOR signalling pathway leads to myeloproliferative syndrome in Foxo3(-/-) mice. EMBO J. 29, 4118–4131 10.1038/emboj.2010.292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yanes O., Clark J., Wong D. M., Patti G. J., Sánchez–Ruiz A., Benton H. P., Trauger S. A., Desponts C., Ding S., Siuzdak G. (2010). Metabolic oxidation regulates embryonic stem cell differentiation. Nat. Chem. Biol. 6, 411–417 10.1038/nchembio.364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yilmaz O. H., Katajisto P., Lamming D. W., Gültekin Y., Bauer–Rowe K. E., Sengupta S., Birsoy K., Dursun A., Yilmaz V. O., Selig M.et al. (2012). mTORC1 in the Paneth cell niche couples intestinal stem-cell function to calorie intake. Nature 486, 490–495 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida Y., Takahashi K., Okita K., Ichisaka T., Yamanaka S. (2009). Hypoxia enhances the generation of induced pluripotent stem cells. Cell Stem Cell 5, 237–241 10.1016/j.stem.2009.08.001 [DOI] [PubMed] [Google Scholar]
- Yu J., Auwerx J. (2009). The role of sirtuins in the control of metabolic homeostasis. Ann. N. Y. Acad. Sci. 1173 Suppl. 1, E10–E19 10.1111/j.1749-6632.2009.04952.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu J., Vodyanik M. A., Smuga–Otto K., Antosiewicz–Bourget J., Frane J. L., Tian S., Nie J., Jonsdottir G. A., Ruotti V., Stewart R.et al. (2007). Induced pluripotent stem cell lines derived from human somatic cells. Science 318, 1917–1920 10.1126/science.1151526 [DOI] [PubMed] [Google Scholar]
- Zhang F., Wang S., Gan L., Vosler P. S., Gao Y., Zigmond M. J., Chen J. (2011a). Protective effects and mechanisms of sirtuins in the nervous system. Prog. Neurobiol. 95, 373–395 10.1016/j.pneurobio.2011.09.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J., Khvorostov I., Hong J. S., Oktay Y., Vergnes L., Nuebel E., Wahjudi P. N., Setoguchi K., Wang G., Do A.et al. (2011b). UCP2 regulates energy metabolism and differentiation potential of human pluripotent stem cells. EMBO J. 30, 4860–4873 10.1038/emboj.2011.401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X., Yalcin S., Lee D. F., Yeh T. Y., Lee S. M., Su J., Mungamuri S. K., Rimmelé P., Kennedy M., Sellers R.et al. (2011c). FOXO1 is an essential regulator of pluripotency in human embryonic stem cells. Nat. Cell Biol. 13, 1092–1099 10.1038/ncb2293 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou J., Su P., Wang L., Chen J., Zimmermann M., Genbacev O., Afonja O., Horne M. C., Tanaka T., Duan E.et al. (2009). mTOR supports long-term self-renewal and suppresses mesoderm and endoderm activities of human embryonic stem cells. Proc. Natl. Acad. Sci. USA 106, 7840–7845 10.1073/pnas.0901854106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou W., Choi M., Margineantu D., Margaretha L., Hesson J., Cavanaugh C., Blau C. A., Horwitz M. S., Hockenbery D., Ware C.et al. (2012). HIF1α induced switch from bivalent to exclusively glycolytic metabolism during ESC-to-EpiSC/hESC transition. EMBO J. 31, 2103–2116 10.1038/emboj.2012.71 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.