

Abstract
Objectives
We investigated the genotoxic effects of 40-59 nm silver nanoparticles (Ag-NPs) by bacterial reverse mutation assay (Ames test), in vitro comet assay and micronucleus (MN) assay. In particular, we directly compared the effect of cytochalasin B (cytoB) and rat liver homogenate (S9 mix) in the formation of MN by Ag-NPs.
Methods
Before testing, we confirmed that Ag-NPs were completely dispersed in the experimental medium by sonication (three times in 1 minute) and filtration (0.2 µm pore size filter), and then we measured their size in a zeta potential analyzer. After that the genotoxicity were measured and especially, S9 mix and with and without cytoB were compared one another in MN assay.
Results
Ames test using Salmonella typhimurium TA98, TA100, TA1535 and TA1537 strains revealed that Ag-NPs with or without S9 mix did not display a mutagenic effect. The genotoxicity of Ag-NPs was also evaluated in a mammalian cell system using Chinese hamster ovary cells. The results revealed that Ag-NPs stimulated DNA breakage and MN formation with or without S9 mix in a dose-dependent manner (from 0.01 µg/mL to 10 µg/mL). In particular, MN induction was affected by cytoB.
Conclusions
All of our findings, with the exception of the Ames test results, indicate that Ag-NPs show genotoxic effects in mammalian cell system. In addition, present study suggests the potential error due to use of cytoB in genotoxic test of nanoparticles.
Keywords: Bacterial reverse mutation test, Comet assay, Genotoxicity, Micronucleus assay, Silver nanoparticles
Introduction
Genetic toxicity testing is routinely performed to identify potential genotoxic carcinogens and germ cell mutagens. In vitro genotoxicity test batteries recommended by regulatory agencies to detect genotoxic carcinogens include at least two or three test procedures, such as bacterial reverse mutation test (Ames test), mammalian cell chromosome damage test and mammalian cell mutation assay [1]. Strategies for assessing the safety of nanoparticles have recently been proposed [2]. Several initiatives, for example, organization for economic cooperation and development (OECD) Working Party on Manufactured Nanomaterials and Nanogenotox Joint Action have been convened, resulting in the establishment of standardized testing methods to determine the genotoxicity of nanoparticles.
The Ames test is known as the most accurate and commonly used procedure to detect genotoxic carcinogens which cause two classes of gene mutation, base pair substitution and small frameshift [3]. It is an essential test within the current battery of assays required for genotoxicity evaluation and has also recently been the conspicuous one of the two assays recommended by the United Kingdom expert advisory Committee on Mutagenicity. Although this test has proven to be invaluable in the safety testing of chemical substances, it has been less commonly used with nanoparticles. The Ames tests of various kinds of nanoparticles have been predominantly negative for several reasons suggested by numerous studies [4,5]. Interestingly, even though many nanoparticles are negative in the Ames test, they have largely been found to have positive genotoxic responses in other in vitro mammalian cell test systems including the comet assay and micronucleus (MN) assay.
A review by researchers [4,6] noted that the in vitro comet and in vitro MN assays in mammalian cell lines were more sensitive and frequently used to confirm the genotoxicity of nanoparticles than the well-known Ames test in bacterial systems. The comet assay is able to detect early DNA breakage with more sensitivity than conventional techniques such as 4',6-diamidino-2-phenylindole staining and DNA flow cytometry [7,8]. It is also one of the most widely used tests and gives the most positive outcome for determining nanoparticles genotoxicity, however, there is no regulatory agency-approved protocol. The in vitro MN assay, which is in support of the draft OECD test guideline (487), rapidly detects small membrane-bound DNA fragments in the cytoplasm of interphase cells. It is recommended as an in vitro genotoxicity testing method to characterize the genotoxicity of chemical and pharmaceutical agents as well as nanoparticles [9,10]. When the study was conducted using both comet and MN assays, many nanoparticles seemed to show positive results in both assays [11,12]. There are, however, examples with opposite results, i.e., an effect showing in only one of the tests [5]. This opposite result may be due to the use of cytochalasin B (cytoB) in in vitro MN assay for some particles, which is used to prevent cytogenesis so that the divided cells stay binucleated. CytoB may inhibit phagocytosis, which indeed may lower the intracellular dose and thus the genotoxicity [13]. Moreover, it is reported that rat liver homogenate (S9 mix) can form micelles of nanoparticles or coat the nanoparticles [14]. This may interfere with the cellular uptake of nanoparticles. Even though there are many concerns about using cytoB and S9 mix in in vitro MN assay for nanoparticles, few papers have directly compared the formation of MN with or without them in mammalian cells.
Among various kinds of nanoparticles, silver nanoparticles (Ag-NPs) are the most commercialized nanoparticles according to the Woodrow-Wilson database which is a data source for information on products based on nanotechnology. The purpose of this study was to investigate the potential genotoxic effects of Ag-NPs using the Ames test in four different bacterial strains and the in vitro comet and MN assays in Chinese hamster ovary (CHO-K1) cells. In particular, we directly compared the effect of cytoB and S9 mix in the formation of MN by Ag-NPs.
Materials and Methods
Preparation and Characterization of Ag-NPs
Ag-NPs (Sigma-Aldrich, St. Louis, MO, USA) were homogeneously dispersed in sterile distilled water (DW) or RPMI1640 media (GIBCO Invitrogen, Grand Island, NY, USA) by sonication for 30 minutes (Bioruptor UCD-200T, Cosmobio Corp., Tokyo, Japan), and were filtered through a cellulose membrane (200 nm pore size; Advantec, Tokyo, Japan). A scanning electron microscope equipped with an energy dispersive spectroscopy (SEM; JEOL, Tokyo, Japan) and a transmission electron microscope (TEM; JEOL, Tokyo, Japan) were used to estimate the size, shape and composition of the Ag-NPs. The Ag-NPs were coated with platinum/palladium and then viewed using SEM. One drop of Ag-NP suspension was dried onto a 400-mesh carbon-coated copper grid and imaged with a TEM at 200 kV. The distribution of Ag-NP size was evaluated using dynamic light scattering (DLS) on the ELSZ-2 (Otsuka Electronics Co., Ltd. Osaka, Japan).
Cell Culture
CHO-K1 cells were obtained from the Korean Cell Line Bank (Seoul, Korea). CHO-K1 cells were grown in RPMI1640 supplemented with 5% fetal bovine serum, penicillin (100 units/mL) and streptomycin (100 µg/mL) at 37℃ in an atmosphere of 5% CO2/95% air under saturated humidity.
Ames Test
Cells were treated with various concentrations of Ag-NPs as described in the toxicological study. Mutagenicity was assessed by the pre-incubation assay as described by Maron and Ames [15]. Briefly, 100 µL of overnight cultures (1-2×108 cfu/mL) of four strains (TA98, TA100, TA1535 and TA1537) were treated separately for 30 minutes at 37℃ [16] with various concentrations of Ag-NPs in the absence or presence of S9 mix. The Ag-NPs were suspended in sterile DW and were used at a final concentration of less than 5% (v/v). For the Ames test, the controls and Ag-NP-treated cells were mixed with 2 mL of sterile top agar (0.6% agar and 0.5% NaCl containing 0.5 mM histidine and 0.5 mM biotin) and poured onto minimal glucose agar plates (1× Vogel-Bonner salts (0.2 g/L magnesium sulfate, 2 g/L citric acid monohydrate, 10 g/L dipotassium hydrogen phosphate, and 3.5 g/L sodium ammonium phosphate), 2% glucose, and 1.5% agar). The plates were then incubated at 37℃ for 48 hours, after which revertants and surviving colonies were counted. Three independent experiments were conducted and each experiment consisted of three replicate plates for each treatment. The positive control used in the absence of S9 mix was 2-nitrofluorene for the TA98 strain, sodium azide for the TA100 and TA1535 strains, and 9-aminoacridine hydrochloride for the TA1537 strain. The positive control used in the presence of S9 mix was 2-aminofluorene for all test strains.
In Vitro Comet Assay
To evaluate the DNA damage induced by Ag-NPs in CHO-K1 cells, we performed the comet assay as described by Singh et al. [17]. CHO-K1 cells were added to a six-well plate and the culture was maintained at 37℃ in a 5% CO2 atmosphere. After 24 hours of incubation, the cells were exposed to Ag-NPs for 24 hours. The treated cells were then trypsinized to produce a single-cell suspension, washed in ice-cold phosphate buffered saline (PBS) and resuspended in 0.7% low melting point agar (Quantum Biotechnologies Inc., Carlsbad, Canada) in PBS at a concentration of 5×104 cells/mL. A 150 µL aliquot of this suspension was pipetted onto precoated glass slides, covered with a glass coverslip, and then the agar was allowed to set at 4℃. The sample was then placed in lysing solution. In the alkaline comet assay, cells were lysed in pH 10 lysis solution (2.5 M NaCl, 100 mM Na2EDTA, 10 mM Tris-HCl, 1% Triton X-100, 10% (v/v) DMSO) at 4℃ for 60 minutes. The lysed cells were rinsed and allowed to unwind for 30 minutes in electrophoresis buffer (1 mM Na2EDTA, 300 mM NaOH, pH >13) before electrophoresis for 30 minutes at 25 V on ice. The gels were then neutralized with 0.4 M Tris-HCl (pH 7.5) for 10 minutes and stained with ethidium bromide (2 µg/mL) just prior to image analysis. DNA migration was assessed by fluorescence microscopy (excitation filter 515 to 560 nm and barrier filter of 590 nm; Leica DMLB, Leica, Wetzlar, Germany) in conjunction with a digital camera. The images were evaluated using an automatic image analysis software (Komet version 5.0; Kinetic Imaging Ltd., Liverpool, UK). The Olive Tail Moment (OTM; tail distance×percentage of DNA in tail) was used to quantify DNA damage, based on the random scoring of 100 nuclei per slide. A minimum of two replicate comet slides were prepared for each sample.
In Vitro Micronucleus assay
The MN assay for evaluating the genotoxic effect by Ag-NPs was performed with cytoB (cytokinesis-block micronucleus, CBMN assay) and without cytoB (MN assay). This is based on the previous studies showing that cytoB affects MN formation [18,19]. The cells were seeded in eight-well chamber slides at a density of 1.5×104 cells/well and cultured for 24 hours until reaching 60%-70% confluence. The CHO-K1 cells were treated with the Ag-NPs in four concentrations in culture medium with or without 3 µg/mL cytoB for 24 hours. After washing two times with PBS, 1% trisodium citrate was added for 5 minutes at 4℃. The slides were then placed in fresh fixative (99:1 methanol:acetic acid) at 4℃, followed by air-drying in a clean bench. Then ribonuclease A (10 µg/mL) dissolved in 2× saline-sodium citrate (SSC; 0.03 M trisodium citrate, 0.3 M NaCl) was added for 6 minutes at 30℃. The slides were rinsed in 2× SSC and dried in a clean bench. After thorough drying, the slides were stained with 5% Giemsa solution for 5 hours. Based on the MN scoring criterion, 1,000 binucleate cells were counted per independent culture. To assess the cytotoxicity induced by cytoB in the CBMN assay, the cytokinesis-block proliferation index (CBPI) was measured in CHO-K1 cells exposed to cytoB and Ag-NPs. CBPI is defined as [20] :
 |
When cytoB was not used, cytotoxicity in CHO-K1 cells exposed to Ag-NPs was measured using relative increase in cell counts (RICC). RICC is defined as [21] :
Data Analysis
Sigma Plot 10.0 (Jandel Science Software, San Rafael, CA, USA) and Excel 2007 (Microsoft, Redmond, WA, USA) were used to analyze the data. The data from each assay were expressed as mean±standard deviations. Statistical analysis was performed using SPSS version 18.0 (SPSS Inc., Chicago, IL, USA). The Shapiro-Wilk and Levene's tests were used to determine the normality of the data distributions and the homogeneity of variances, respectively. Differences between groups were determined by Duncan's post hoc test following one-way analysis of variance. Statistical significance was accepted at p<0.05 and 0.01.
Results
Characterization of Ag-NPs
In genotoxicity studies, Ag-NPs were characterized by SEM, TEM, and DLS, which showed spherical aggregates that were approximately 58.9 nm in size (Figure 1). In SEM and TEM analysis, the single particle size of Ag-NPs was 100 nm or less (Figure 1A, B). Although the size distribution of Ag-NPs by DLS ranged from 45 to 315 nm due to aggregation, 40-59 nm nanoparticles accounted for approximately 50% (Figure 1C). Energy dispersion spectrum (EDS) recorded from the Ag-NPs are shown in Figure 1D. EDS profile showed strong silver signal along with a weak oxygen and carbon peak, which may be derived from the partial oxidation.
Figure 1.

Characterization of silver nanoparticles using scanning electron microscope (A); transmission electron microscope (B); dynamic light scattering (C); and energy dispersion spectrum (D).
Ames Test
The Ag-NPs suspended in sterile DW were tested for bacterial mutagenicity using Salmonella typhimurium strains, TA98, TA100, TA1535 and TA1537, with and without S9 mix. The concentrations of Ag-NPs used were 100, 200, 300, 400, and 500 µg per plate. Growth inhibition by Ag-NPs was observed in all test strains with or without S9 mix at a concentration of 500 µg/plate or higher (Table 1). Large increases in the number of revertant colonies were seen for the positive controls in all cases, indicating that the test system responded appropriately. However, Ag-NP mutagenicity was not detected in any strain with or without S9 mix, as shown in Table 1.
Table 1.
Mutagenicity of silver nanoparticles in Salmonella typhimurium test strains
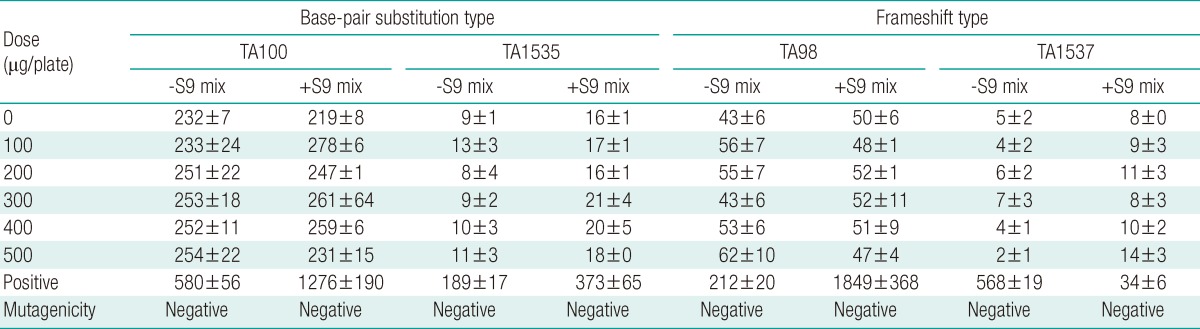
The positive control in the absence of S9 mix was 2-nitrofluorene for the TA98 strain, sodium azide for the TA100 and TA1535 strains, and 9-aminoacridine hydrochloride for the TA1537 strain. The positive control used in the presence of S9 mix was 2-aminofluorene for all tester strains.
DNA Breakage in CHO-K1 Cells Exposed to Ag-NPs
In order to identify DNA damage in mammalian cells, we carried out comet assays in CHO-K1 cells treated with Ag-NPs at non-cytotoxic concentrations (data not shown). DNA damage was quantified and expressed as the OTM. In the comet assay, CHO-K1 cells treated with Ag-NPs exhibited a dose-dependent increase in DNA breakage (Figure 2). The Ag-NPs (10 µg/mL) caused an approximately 450% increase in DNA breakage compared to the control (p<0.01).
Figure 2.

DNA strand damage in Chinese hamster ovary cells exposed to silver nanoparticles (Ag-NPs), as determined by the comet assay. Cells were treated with 0.01-10 µg/mL of Ag-NPs or with H2O2 (25 uM) as a positive control. Results are expressed as mean±standard deviation. of three separate experiments for each data point of the olive tail moment (% DNA in tail×distance between centers of mass). †Significantly different from the control at p<0.01.
Micronucleus Formation in CHO-K1 Cells Exposed to Ag-NPs
To identify chromosomal damage in CHO-K1 cells exposed to Ag-NPs, we carried out CBMN and MN assays (Figure 3). The CBMN assay was carried out in the presence or absence of S9 mix. In the CBMN assay, percentage of binucleated cells (%BN) and the CBPI was 80.6% and 1.95 for S9 mix (+) and 81.0% and 1.98 for S9 mix (-), respectively. Also, in the MN assay, the RICC was 85.9%. Treatment with Ag-NPs 10 µg/mL did not induce significant reductions in the %BN, CBPI or RICC, indicating that Ag-NPs did not induce cell cycle delay for 24 hours. In the CBMN assay with S9 mix, significantly greater MN formation was observed in cells treated with Ag-NPs (10 µg/mL; 3.0-fold) compared to the control (p<0.05). In the CBMN assay without S9 mix, MN formation was also significantly induced by Ag-NPs (10 µg/mL; 2.8-fold) compared to the control (p<0.05). However, we found that there was no significant difference in the results with or without S9 mix in the CBMN assay. Meanwhile, in the MN assay (without cytoB), MN formation was significantly induced by Ag-NPs (10 µg/mL; 3.8-fold) compared to the control (p<0.05). The results of the CBMN and MN assays without S9 mix were significantly different (p<0.05).
Figure 3.

Micronucleus (MN) formation induced by silver nanoparticles (Ag-NPs) in Chinese hamster ovary cells. Cells were treated with Ag-NPs (0.01-10 µg/mL) under conditions of +S9 mix/+cytochalasin B (white bar) or -S9 mix/+cytochalasin B (green bar) or -S9 mix/-cytochalasin B (red bar). The results are expressed as mean±standard deviation. of three separate experiments for each data point of MN frequency per 1,000 cells. MN frequencies per 1,000 cells of control are 12.2±0.8 (white bar), 13.5±2.1 (green bar) and 9.5±0.7 (red bar), respectively. The percentage of binucleated cells (%BN) and the cytokinesis-block proliferation index was 80.6% and 1.95 for white bar and 81.0% and 1.98 for green bar, respectively. The relative increase in cell counts of red bar was 85.9%. *Significantly different from the control at p<0.05. †p<0.05 between the cytokinesis-block micronucleus assay (with cytochalasin B) and MN assay (without cytochalasin B) values.
Discussion
Ag-NPs of different sizes, shapes and material properties have many applications in various areas such as in industrial, medical and cosmetic fields. However, there is no current data on the characteristics of Ag-NPs such as size and shape in the workplace atmosphere. Many researchers have suggested that size is a critical factor for nanoparticle-induced toxicity and biological responses [22,23]. Park et al. [24] reported that the potency of Ag-NPs to induce cell damage is size-dependent. Thus, the genotoxicity of Ag-NPs needs to be investigated as a function of particle size.
There have been many studies on the genotoxicity of Ag-NPs with different sizes. Dose-dependent genotoxicity caused by 6-20 nm Ag-NPs, as measured by the comet and CBMN assays, was noted in human glioblastoma cells and human lung fibroblast cells [11]. Exposure to 7-10 nm Ag-NPs increased the frequency of MN formation up to 47.9±3.2% of binucleated human hepatoma cells [25]. Li et al. [26] also reported that human lymphoblastoid cells exposed to 5 nm Ag-NPs showed a dose-dependent increase in MN frequency. Furthermore, Foldbjerg et al. [27] reported that DNA damage was detected as an increase in bulky DNA adducts by 32P post-labeling in human alveolar cells exposed to 120-150 nm Ag-NPs. On the contrary, no significant genotoxic responses were observed in an Ames test (5 nm) [26], a mouse lymphoma assay (<100 nm) [28] and an in vivo MN assay (60 nm) [29]. Jiang et al. [23] reported that the sizes at which gold and silver nanoparticles exhibit the greatest cellular responses are 40 and 50 nm. For this reason, we used 40-59 nm sized Ag-NPs in our previous study and found that they caused significant DNA and chromosomal damage in human bronchial epithelial cells [18]. In the current study, we also used 40-59 nm Ag-NPs and evaluated their in vitro genotoxicity in CHO-K1 cells by three test methodologies (Ames test, comet and MN assays) which are considered to be part of the standard in vitro genotoxicity test battery. Ag-NPs were observed as spherical aggregates (Figure 1), but 40-59 nm nanoparticles accounted for approximately 50% of the Ag-NPs, despite this aggregation.
In Vitro Genotoxicity of Ag-NPs Using the Ames Test, Comet Assay and Micronucleus Assay
The Ames test is generally used as a first screening method to assess chemical genotoxicity. In the database published by Kirkland et al. [1], 542 had published Ames results out of 756 rodent carcinogens. Although Ames test has proven to be invaluable in the safety testing of chemical substances, it has been less commonly used with nanoparticles. Doak et al. [30] noted that 17 studies reported negative mutagenicity among the published 19 Ames test studies. In this study, the Ag-NPs were negative in the Ames test with or without S9 mix (Table 1), and this result is consistent with previous reports that indicated Ames tests on nanoparticles were predominantly negative [4,5]. Many reasons suggested by numerous studies support that Ames test does not appear to be suitable for the assessment of nanoparticles.
We conducted the comet and MN assays to confirm the genotoxicity of Ag-NPs in CHO-K1 cells. A review by researchers [4,6] noted that the in vitro comet and in vitro MN assays in mammalian cell lines were more sensitive and frequently used to confirm the genotoxicity of nanoparticles than the well-known Ames test in bacterial systems. The Ag-NPs induced dose-dependent DNA breakage in the comet assay (Figure 2). This result is not surprising since many studies reported that nanoparticles showed positive genotoxic responses in other in vitro mammalian cell test systems but negative results in the Ames test [30]. The notable case is when two independent mammalian cell genotoxic tests showed different results. Single-walled carbon nanotubes (SWCNT) with 1 nm in diameter showed an increase of DNA breakage but negative results for mutagenicity. The MN assay simultaneously indicated some but not significant MN induction by SWCNT at the highest concentration tested. Moreover, nano-sized titanium dioxide (TiO2) rutile induced DNA breakage but did not cause significant MN induction [13]. They supposed that cytoB, which is used to prevent cytogenesis, may inhibit phagocytosis and decrease the genotoxicity by TiO2 rutile. We investigated MN induction by Ag-NPs in MN assay and CBMN assay with or without S9 mix. As a result, MN induction in all MN assays showed significant increase regardless of S9 mix or cytoB usage (Figure 3). In particular, S9 mix did not cause any difference in MN induction in the CBMN assay. Since Ag-NPs significantly increased genotoxic effects in in vitro mammalian cell test systems (in vitro comet and MN assays), we determined Ag-NPs as positive genotoxic substance. In vivo genotoxic tests of 40-59 nm Ag-NPs are required to carry out in the further studies.
Critical Considerations for Genotoxicity Evaluation of Ag-NPs
Most of nanoparticles on Ames test showed negative results including this study. There are several explanations for these negative Ames results. First, the negative results may be due to the inability of the nanoparticles to penetrate through the bacterial cell wall [4]. Prokaryotes (including bacterial cells) cannot perform endocytosis [30], which was evidenced in the uptake mechanism of Ag-NP aggregates in our previous study [18]. In addition, the Ag-NPs could be too large to be transported through the pores in the bacterial cell wall. It is therefore important to consider the fact that a lack of Ag-NP uptake in bacterial strains could potentially lead to false negative results. Second, the Ames test is not recommended for detecting genotoxins that induce large-scale DNA damage. Balasubramanyam et al. [31] reported that aluminum oxide nanoparticles were negative in the Ames test using Salmonella typhimurium TA97a, TA98, TA100, TA102 and TA1535 strains, although they caused size- and dose-dependent genotoxicity in other genotoxic tests [32,33]. They suggested that the results in the Ames test were negative because the nanoparticles generate large deletions including the histidine gene in the test strains. Consistent with Balasubramanyam et al. [31], report we observed negative Ames results even though Ag-NPs induced MN formation and DNA breakage in CHO-K1 cells. However, we were unable to determine whether Ag-NPs actually caused deletions in the histidine gene in all strains. Finally, it is also important to consider the fact that some nanoparticles, especially Ag-NPs, have antimicrobial activity [34]. There is a possibility that the inherent toxicity of Ag-NPs to the bacterial strains may have reduced the sensitivity of the test [26]. We also observed that all the strains were killed by Ag-NPs at concentrations at or above 500 µg/plate. Consequently, a combination of properties (including uptake mechanism and toxicity) of Ag-NPs may result in false negative results and an underestimation of their mutagenicity in the Ames test.
The possible factors which can affect to nanoparticles uptake into the cells in MN assay. First, Li et al. [26] expected that Ag-NPs were pure metal particles and were unlikely to be metabolized by S9 mix. However, S9 mix may have an effect on MN induction by Ag-NPs, because S9 mix can form micelles of nanoparticles or coat the nanoparticles [14]. Therefore, we compared MN induction in CBMN assay with or without S9 mix, and we confirmed that S9 mix did not cause any difference in MN induction in the CBMN assay (Figure 3; white and green bars). Second, we considered use of cytoB to stop dividing cells from performing cytokinesis. Many researchers reported that cytoB itself could interfere with the uptake of particles [18,35]. The blocking of cellular phagocytosis with cytoB dramatically reduced the mutagenecity of diesel exhausted particles (less than 1 µm) in mammalian cells [36]. Furthermore, the endocytosis process of aminosilane-coated Fe3O4 nanoparticles (approximately 30 nm) was effectively inhibited by cytoB [37]. We identified that 40-59 nm Ag-NPs may become compartmentalized within cells by phagocytosis of aggregates [18], and we assumed that gentoxocity of Ag-NPs could be affected by cytoB. Therefore, we compared MN induction by Ag-NPs in CBMN assay (with cytoB) and MN assay (without cytoB). Ag-NPs also stimulated a significant and dose-dependent increase of MN formation in the CBMN and MN assays (Figure 3; green and red). In particular, MN induction (10 µg/mL; 3.8-fold) in the MN assay was significantly greater than that (10 µg/mL; 2.8-fold) in the CBMN assay. These results are consistent with Falck et al. [13] report about the genotoxicity of TiO2 rutile decreased in the CBMN assay. In contrast, the frequency of MN observed in the MN assay on SWCNT was approximately half that detected by the sequential CBMN assay [38]. The SWCNT that were assessed had lengths of 400-800 nm, with a diameter of 1- to 2-nm. The authors suggested that endocytosis is not the only means for SWCNT uptake leading to DNA damage, as micronuclei were observed despite co-incubation with both SWCNT and cytoB. Therefore, we suggest that the in vitro CBMN assay may have underestimated the genotoxicity of nanoparticles, especially when they were taken up into the cell by phagocytosis.
In conclusion, Ag-NPs did not induce mutations in the Ames test. However, Ag-NPs exhibited dose-dependent genotoxicity in the comet and MN assays in CHO-K1 cells. The data indicated that the in vitro comet and MN assays may be more appropriate than the Ames test for evaluating the genotoxicity of Ag-NPs. Moreover, extra caution is required when using cytoB. Future studies are required to develop an in vitro test battery for the assessment of nanoparticle genotoxicity considering their distinctive characteristics including size, shape and uptake mechanism.
Acknowledgements
This study was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (MEST) (2010-0021587).
Footnotes
The authors have no conflict of interest to declare on this study.
This article is available from: http://e-eht.org/
References
- 1.Kirkland D, Aardema M, Henderson L, Müller L. Evaluation of the ability of a battery of three in vitro genotoxicity tests to discriminate rodent carcinogens and non-carcinogens I. Sensitivity, specificity and relative predictivity. Mutat Res. 20054;584(1-2):1–256. doi: 10.1016/j.mrgentox.2005.02.004. [DOI] [PubMed] [Google Scholar]
- 2.Warheit DB, Donner EM. Rationale of genotoxicity testing of nanomaterials: regulatory requirements and appropriateness of available OECD test guidelines. Nanotoxicology. 2010;4:409–413. doi: 10.3109/17435390.2010.485704. [DOI] [PubMed] [Google Scholar]
- 3.Ames BN, Lee FD, Durston WE. An improved bacterial test system for the detection and classification of mutagens and carcinogens. Proc Natl Acad Sci USA. 1973;70(3):782–786. doi: 10.1073/pnas.70.3.782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Landsiedel R, Kapp MD, Schulz M, Wiench K, Oesch F. Genotoxicity investigations on nanomaterials: methods, preparation and characterization of test material, potential artifacts and limitations--many questions, some answers. Mutat Res. 2009;681(2-3):241–258. doi: 10.1016/j.mrrev.2008.10.002. [DOI] [PubMed] [Google Scholar]
- 5.Kisin ER, Murray AR, Keane MJ, Shi XC, Schwegler-Berry D, Gorelik O, et al. Single-walled carbon nanotubes: geno- and cytotoxic effects in lung fibroblast V79 cells. J Toxicol Environ Health A. 2007;70(24):2071–2079. doi: 10.1080/15287390701601251. [DOI] [PubMed] [Google Scholar]
- 6.Oesch F, Landsiedel R. Genotoxicity investigations on nanomaterials. Arch Toxicol. 2012;86(7):985–994. doi: 10.1007/s00204-012-0838-y. [DOI] [PubMed] [Google Scholar]
- 7.Olive PL, Frazer G, Banáth JP. Radiation-induced apoptosis measured in TK6 human B lymphoblast cells using the comet assay. Radiat Res. 1993;136(1):130–136. [PubMed] [Google Scholar]
- 8.Godard T, Deslandes E, Lebailly P, Vigreux C, Poulain L, Sichel F, et al. Comet assay and DNA flow cytometry analysis of staurosporine-induced apoptosis. Cytometry. 1999;36(2):117–122. doi: 10.1002/(sici)1097-0320(19990601)36:2<117::aid-cyto5>3.0.co;2-#. [DOI] [PubMed] [Google Scholar]
- 9.Kirkland D, Reeve L, Gatehouse D, Vanparys P. A core in vitro genotoxicity battery comprising the Ames test plus the in vitro micronucleus test is sufficient to detect rodent carcinogens and in vivo genotoxins. Mutat Res. 2011;721(1):27–73. doi: 10.1016/j.mrgentox.2010.12.015. [DOI] [PubMed] [Google Scholar]
- 10.Gonzalez L, Sanderson BJ, Kirsch-Volders M. Adaptations of the in vitro MN assay for the genotoxicity assessment of nanomaterials. Mutagenesis. 2011;26(1):185–191. doi: 10.1093/mutage/geq088. [DOI] [PubMed] [Google Scholar]
- 11.AshaRani PV, Low Kah, Hande MP, Valiyaveettil S. Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano. 2009;3(2):279–290. doi: 10.1021/nn800596w. [DOI] [PubMed] [Google Scholar]
- 12.Lindberg HK, Falck GC, Suhonen S, Vippola M, Vanhala E, Catalán J, et al. Genotoxicity of nanomaterials: DNA damage and micronuclei induced by carbon nanotubes and graphite nanofibres in human bronchial epithelial cells in vitro. Toxicol Lett. 2009;186(3):166–173. doi: 10.1016/j.toxlet.2008.11.019. [DOI] [PubMed] [Google Scholar]
- 13.Falck GC, Lindberg HK, Suhonen S, Vippola M, Vanhala E, Catalán J, et al. Genotoxic effects of nanosized and fine TiO2. Hum Exp Toxicol. 2009;28(6-7):339–352. doi: 10.1177/0960327109105163. [DOI] [PubMed] [Google Scholar]
- 14.Kumar A, Pandey AK, Singh SS, Shanker R, Dhawan A. Cellular uptake and mutagenic potential of metal oxide nanoparticles in bacterial cells. Chemosphere. 2011;83(8):1124–1132. doi: 10.1016/j.chemosphere.2011.01.025. [DOI] [PubMed] [Google Scholar]
- 15.Maron DM, Ames BN. Revised methods for the Salmonella mutagenicity test. Mutat Res. 1983;113(3-4):173–215. doi: 10.1016/0165-1161(83)90010-9. [DOI] [PubMed] [Google Scholar]
- 16.Bajpayee M, Pandey AK, Zaidi S, Musarrat J, Parmar D, Mathur N, et al. DNA damage and mutagenicity induced by endosulfan and its metabolites. Environ Mol Mutagen. 2006;47(9):682–692. doi: 10.1002/em.20255. [DOI] [PubMed] [Google Scholar]
- 17.Singh NP, McCoy MT, Tice RR, Schneider EL. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp Cell Res. 1988;175(1):184–191. doi: 10.1016/0014-4827(88)90265-0. [DOI] [PubMed] [Google Scholar]
- 18.Kim HR, Kim MJ, Lee SY, Oh SM, Chung KH. Genotoxic effects of silver nanoparticles stimulated by oxidative stress in human normal bronchial epithelial (BEAS-2B) cells. Mutat Res. 2011;726(2):129–135. doi: 10.1016/j.mrgentox.2011.08.008. [DOI] [PubMed] [Google Scholar]
- 19.Papageorgiou I, Brown C, Schins R, Singh S, Newson R, Davis S, et al. The effect of nano- and micron-sized particles of cobalt-chromium alloy on human fibroblasts in vitro. Biomaterials. 2007;28(19):2946–2958. doi: 10.1016/j.biomaterials.2007.02.034. [DOI] [PubMed] [Google Scholar]
- 20.Kirsch-Volders M, Sofuni T, Aardema M, Albertini S, Eastmond D, Fenech M, et al. Report from the in vitro micronucleus assay working group. Mutat Res. 2003;540(2):153–163. doi: 10.1016/j.mrgentox.2003.07.005. [DOI] [PubMed] [Google Scholar]
- 21.Lorge E, Hayashi M, Albertini S, Kirkland D. Comparison of different methods for an accurate assessment of cytotoxicity in the in vitro micronucleus test. I. Theoretical aspects. Mutat Res. 2008;655(1-2):1–3. doi: 10.1016/j.mrgentox.2008.06.003. [DOI] [PubMed] [Google Scholar]
- 22.Nel A, Xia T, Mädler L, Li N. Toxic potential of materials at the nanolevel. Science. 2006;311(5761):622–627. doi: 10.1126/science.1114397. [DOI] [PubMed] [Google Scholar]
- 23.Jiang W, Kim BY, Rutka JT, Chan WC. Nanoparticle-mediated cellular response is size-dependent. Nat Nanotechnol. 2008;3(3):145–150. doi: 10.1038/nnano.2008.30. [DOI] [PubMed] [Google Scholar]
- 24.Park MV, Neigh AM, Vermeulen JP, de la Fonteyne LJ, Verharen HW, Briedé JJ, et al. The effect of particle size on the cytotoxicity, inflammation, developmental toxicity and genotoxicity of silver nanoparticles. Biomaterials. 2011;32(36):9810–9817. doi: 10.1016/j.biomaterials.2011.08.085. [DOI] [PubMed] [Google Scholar]
- 25.Kawata K, Osawa M, Okabe S. In vitro toxicity of silver nanoparticles at noncytotoxic doses to HepG2 human hepatoma cells. Environ Sci Technol. 2009;43(15):6046–6051. doi: 10.1021/es900754q. [DOI] [PubMed] [Google Scholar]
- 26.Li Y, Chen DH, Yan J, Chen Y, Mittelstaedt RA, Zhang Y, et al. Genotoxicity of silver nanoparticles evaluated using the Ames test and in vitro micronucleus assay. Mutat Res. 2012;745(1-2):4–10. doi: 10.1016/j.mrgentox.2011.11.010. [DOI] [PubMed] [Google Scholar]
- 27.Foldbjerg R, Dang DA, Autrup H. Cytotoxicity and genotoxicity of silver nanoparticles in the human lung cancer cell line, A549. Arch Toxicol. 2011;85(7):743–750. doi: 10.1007/s00204-010-0545-5. [DOI] [PubMed] [Google Scholar]
- 28.Kim YJ, Yang SI, Ryu JC. Cytotoxicity and genotoxicity of nano-silver in mammalian cell lines. Mol Cell Toxicol. 2010;6(2):119–125. [Google Scholar]
- 29.Kim YS, Kim JS, Cho HS, Rha DS, Kim JM, Park JD, et al. Twenty-eight-day oral toxicity, genotoxicity, and gender-related tissue distribution of silver nanoparticles in Sprague-Dawley rats. Inhal Toxicol. 2008;20(6):575–583. doi: 10.1080/08958370701874663. [DOI] [PubMed] [Google Scholar]
- 30.Doak SH, Manshian B, Jenkins GJ, Singh N. In vitro genotoxicity testing strategy for nanomaterials and the adaptation of current OECD guidelines. Mutat Res. 2012;745(1-2):104–111. doi: 10.1016/j.mrgentox.2011.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Balasubramanyam A, Sailaja N, Mahboob M, Rahman MF, Hussain SM, Grover P. In vitro mutagenicity assessment of aluminium oxide nanomaterials using the Salmonella/microsome assay. Toxicol In Vitro. 2010;24(6):1871–1876. doi: 10.1016/j.tiv.2010.07.004. [DOI] [PubMed] [Google Scholar]
- 32.Balasubramanyam A, Sailaja N, Mahboob M, Rahman MF, Hussain SM, Grover P. In vivo genotoxicity assessment of aluminium oxide nanomaterials in rat peripheral blood cells using the comet assay and micronucleus test. Mutagenesis. 2009;24(3):245–251. doi: 10.1093/mutage/gep003. [DOI] [PubMed] [Google Scholar]
- 33.Balasubramanyam A, Sailaja N, Mahboob M, Rahman MF, Misra S, Hussain SM, et al. Evaluation of genotoxic effects of oral exposure to aluminum oxide nanomaterials in rat bone marrow. Mutat Res. 2009;676(1-2):41–47. doi: 10.1016/j.mrgentox.2009.03.004. [DOI] [PubMed] [Google Scholar]
- 34.Kim JS, Kuk E, Yu KN, Kim JH, Park SJ, Lee HJ, et al. Antimicrobial effects of silver nanoparticles. Nanomedicine. 2007;3(1):95–101. doi: 10.1016/j.nano.2006.12.001. [DOI] [PubMed] [Google Scholar]
- 35.Haynes P, Lambert TR, Mitchell ID. Comparative in-vivo genotoxicity of antiviral nucleoside analogues; penciclovir, acyclovir, ganciclovir and the xanthine analogue, caffeine, in the mouse bone marrow micronucleus assay. Mutat Res. 1996;369(1-2):65–74. doi: 10.1016/s0165-1218(96)90049-x. [DOI] [PubMed] [Google Scholar]
- 36.Bao L, Chen S, Wu L, Hei TK, Wu Y, Yu Z, et al. Mutagenicity of diesel exhaust particles mediated by cell-particle interaction in mammalian cells. Toxicology. 2007;229(1-2):91–100. doi: 10.1016/j.tox.2006.10.007. [DOI] [PubMed] [Google Scholar]
- 37.Ma YJ, Gu HC. Study on the endocytosis and the internalization mechanism of aminosilane-coated Fe3O4 nanoparticles in vitro. J Mater Sci Mater Med. 2007;18(11):2145–2149. doi: 10.1007/s10856-007-3015-8. [DOI] [PubMed] [Google Scholar]
- 38.Doak SH, Griffiths SM, Manshian B, Singh N, Williams PM, Brown AP, et al. Confounding experimental considerations in nanogenotoxicology. Mutagenesis. 2009;24(4):285–293. doi: 10.1093/mutage/gep010. [DOI] [PubMed] [Google Scholar]