

Abstract
Water is a key resource, and the plant water transport system sets limits on maximum growth and drought tolerance. When plants open their stomata to achieve a high stomatal conductance (gs) to capture CO2 for photosynthesis, water is lost by transpiration1,2. Water evaporating from the airspaces is replaced from cell walls, in turn drawing water from the xylem of leaf veins, in turn drawing from xylem in the stems and roots. As water is pulled through the system, it experiences hydraulic resistance, creating tension throughout the system and a low leaf water potential (Ψleaf). The leaf itself is a critical bottleneck in the whole plant system, accounting for on average 30% of the plant hydraulic resistance3. Leaf hydraulic conductance (Kleaf = 1/ leaf hydraulic resistance) is the ratio of the water flow rate to the water potential gradient across the leaf, and summarizes the behavior of a complex system: water moves through the petiole and through several orders of veins, exits into the bundle sheath and passes through or around mesophyll cells before evaporating into the airspace and being transpired from the stomata. Kleaf is of strong interest as an important physiological trait to compare species, quantifying the effectiveness of the leaf structure and physiology for water transport, and a key variable to investigate for its relationship to variation in structure (e.g., in leaf venation architecture) and its impacts on photosynthetic gas exchange. Further, Kleaf responds strongly to the internal and external leaf environment3. Kleaf can increase dramatically with irradiance apparently due to changes in the expression and activation of aquaporins, the proteins involved in water transport through membranes4, and Kleaf declines strongly during drought, due to cavitation and/or collapse of xylem conduits, and/or loss of permeability in the extra-xylem tissues due to mesophyll and bundle sheath cell shrinkage or aquaporin deactivation5-10. Because Kleaf can constrain gs and photosynthetic rate across species in well watered conditions and during drought, and thus limit whole-plant performance they may possibly determine species distributions especially as droughts increase in frequency and severity11-14.
We present a simple method for simultaneous determination of Kleaf and gs on excised leaves. A transpiring leaf is connected by its petiole to tubing running to a water source on a balance. The loss of water from the balance is recorded to calculate the flow rate through the leaf. When steady state transpiration (E, mmol • m-2 • s-1) is reached, gs is determined by dividing by vapor pressure deficit, and Kleaf by dividing by the water potential driving force determined using a pressure chamber (Kleaf= E /- Δψleaf, MPa)15.
This method can be used to assess Kleaf responses to different irradiances and the vulnerability of Kleaf to dehydration14,16,17.
Keywords: Plant Biology, Issue 70, Molecular Biology, Physiology, Ecology, Biology, Botany, Leaf traits, hydraulics, stomata, transpiration, xylem, conductance, leaf hydraulic conductance, resistance, evaporative flux method, whole plant
Protocol
1. Measuring Leaf Hydraulic Conductance for Hydrated Leaves (Kmax)
Collect shoots preferably in the evening or night such that xylem tensions are low, cut in air and place in dark plastic bag filled with wet paper towels. In the lab, recut at least two nodes from the end of every shoot under water. Let shoot rehydrate overnight in pure water, or other solution to be used for Kleaf measurements. Ultrapure water is recommended as a flow solution unless the impacts of different solutions are to be investigated as a factor.
Cut the leaf from the shoot with a fresh razor blade under partially degassed flow solution. Ultrapure water is recommended as a flow solution unless the impacts of different solutions are to be investigated as a factor. We recommend degassing the solution overnight in a flask using a vacuum pump, but other methods may be employed.
Wrap pre-stretched parafilm around the petiole to ensure a good seal between petiole and tubing. This step is especially helpful for non-round petioles, as wrapping extra parafilm will help round the petiole to fit inside the tubing. Dental paste can be applied to the petiole before wrapping the parafilm to fill grooves in petioles.
Connect the leaf to silicone tubing under ultrapure water to prevent air entering the system. As the leaf is lifted above the source of water, the water in the tubing is under subatmospheric pressure, and thus the seal is good if no air is drawn into the tubing during the measurement. Alternatively to sealing the petiole directly into silicone tubing, a compression fitting with rubber gasket can be used (e.g., (Omnifit A2227 bore adaptor; Omnifit, Cambridge, UK); this approach is useful for petioles with especially complex shapes, or for grass blades, which can be wrapped around a glass rod and sealed with the compression fitting. When feasible, sealing the petiole directly into silicone tubing has the advantage of being rapid and leaving a petiole-tubing junction that is transparent, allowing bubbles to be more easily seen. The tubing connects the leaf to hard tubing running to a graduated cylinder on a balance (±10 μg resolution), which logs data every 30 sec to a computer (e.g. using "Balance Link" Software; Mettler-Toledo GmbH, Greifensee, Switzerland) for the calculation of flow rate through the leaf.
Leaves should be held adaxial surface upwards in wood frames strung with fishing line above a large box fan and under a light source ( > 1,000 μmol • m2 • s-1 photosynthetically active radiation for high-irradiance measurements), slightly above the level of the meniscus in the graduated cylinder. The leaf should be placed above the water in the graduated cylinder to ensure that water moving to the leaf is driven by transpiration, and not entering the leaf under pressure due to gravity, as would be the case if the leaf were placed below the meniscus of the water in the cylinder. Small containers filled with wet paper towel are placed inside the balance chamber to ensure the air in the balance is water-saturated, thus avoiding evaporation from the water source to the atmosphere in the balance. This possibility can be checked by halting flow into the leaf using a four-way stopcock valve in the tubing, and checking that the mass of the water cylinder on the balance does not decline.
Place a clear Pyrex container filled with water above the leaf to absorb the heat of the lamp.
Maintain leaf temperature between 23 °C and 28 °C throughout the experiment by adding more cold water to the Pyrex container such that the heat lamp does not heat the Pyrex container sufficiently to warm the air around the leaf below.
Allow the leaf to transpire on the apparatus for at least 30 min, thus giving sufficient time to acclimate to high irradiance, and additionally until flow rate has stabilized, with no upward or downward trend, and with a coefficient of variation <5% for at least 5-10 measurements made at 30 sec flow intervals. It is essential for the flow rate to reach a steady state because this method assumes a stable Ψleaf. Previous studies found these criteria to be sufficient for stabilization of flow rate (E), Ψleaf and Kleaf; tests with longer measurement periods after stable flow was established showed no relationship of Kleaf to measurement time for each of seven species of a wide range of leaf capacitance17,18 . Stabilization generally takes 30 min or less, but in some species may require more than an hour for some species.
Note: When flow rate is very low (<8 μg s-1), stability can be determined using the running average of the last five 30 sec intervals.
Discard measurements if the flow suddenly changes, either due to sudden stomatal closure, or apparent leakage from the seal or blockage in the system by particles or air bubbles.
When flow stabilizes, record leaf temperature with a thermocouple.
Quickly remove the leaf from the tubing, dab dry the petiole and place the leaf into a sealable bag which had been previously exhaled in, to halt transpiration.
Let the leaf equilibrate in its bag for at least 20 min.
Average the final 10 flow rate measurements. This will be your E measurement.
When the leaf is equilibrated, measure the final leaf water potential (Ψfinal) with a pressure chamber.
Measure leaf area with a flatbed scanner and image processing software, or with a leaf area meter.
Kleaf is calculated as E / -ΔΨleaf (where ΔΨleaf = Ψfinal - 0 MPa) and further normalized by leaf area. Units are in mmol m-2 s-1 MPa-1. Because the leaf is positioned just above the level of water in the graduated cylinder, the effect of gravity in reducing the pressure of water entering the leaf would be negligible, especially relative to the Ψfinal, which is always > -0.1 MPa.
To correct for changes in Kleaf induced by temperature dependence of water viscosity15,19,20 , standardize Kleaf values to 25 °C by using the following equation:
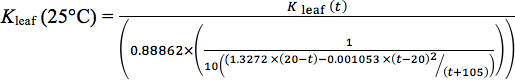
Stomatal conductance (gs) can be determined as E divided by the leaf-to-air vapor pressure difference, equivalent to the vapor pressure deficit (VPD), determined using a temperature and relative humidity sensor adjacent to the leaf. VPD is calculated using the following equation21
![]() where RH is the relative humidity, VP is the saturation vapor pressure of the air (in kPa; a function of temperature, according to the Arden Buck equation22), and AP the atmospheric pressure (in kPa).
where RH is the relative humidity, VP is the saturation vapor pressure of the air (in kPa; a function of temperature, according to the Arden Buck equation22), and AP the atmospheric pressure (in kPa).
2. Measuring Leaf Hydraulic Conductance for Dehydrated Leaves
Collect shoots preferably in the evening or night such that xylem tensions are low, cut in air and place in dark plastic bags filled with wet paper towels. In the lab, recut at least two nodes from the end of every shoot under water. Let shoot rehydrate overnight in pure water, or other solution to be used for Kleaf measurements. Ultrapure water is recommended as a flow solution unless the impacts of different solutions are to be investigated as a factor.
Cut shoots into segments with at least three leaves under ultrapure water and dehydrate shoots with a fan for different periods of time to a range of Ψleaf values.
Once shoots are dehydrated, place sealable bags (Whirl-Pak; Nasco, Fort Atkinson, WI, USA) which have been previously exhaled in) around each leaf of the shoot, and then place the whole shoot in a large sealable bag with wet paper towel.
Let the shoot equilibrate for at least 30 min (for strongly dehydrated shoots longer equilibration times may be necessary).
After equilibration, excise the top and bottom leaf and measure initial Ψleaf (=Ψo) using a pressure chamber.
Discard the shoot if the difference in the Ψleaf of those two leaves is greater than 0.1 MPa (for strongly dehydrated leaves, this range can be extended to 0.3 MPa).
The third leaf is then used to determine Kleaf and gs with the EFM, following steps 1.2 -1.18 above. Notably, steady state E can be obtained even for leaves that have been severely dehydrated leaves, as stomata still open and generate gs values substantially higher than cuticular conductance17.
To construct hydraulic vulnerability curves, calculate Kleaf using the Ψfinal as in step 1.16, and then plot Kleaf values against whichever is lowest, Ψo or Ψfinal, i.e., the strongest dehydration experienced by the leaf during the experiment (=Ψlowest). Similarly, the decline of gs in response to leaf dehydration can be determined by plotting gs against Ψlowest. Previous work has shown that dehydration induces at least partially irreversible declines in Kleaf and gs14,16.
We recommend obtaining at least 6 Kleaf values per 0.5 MPa interval of Ψlowest (this interval can be reduced to 0.25 MPa if the species you are working with is really vulnerable to dehydration).
Perform Dixon outlier test on Kleaf values for each 0.5 MPa water potential interval of Ψlowest to remove outliers from your vulnerability curve23.
Because species differ in their responses of Kleaf and gs to dehydration14,16, fit different functions for each species and select the maximum likelihood (lowest AIC value; see 14 for details). We recommend fitting at least four functions previously used in the literature: linear (
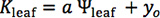 ), sigmoidal (
), sigmoidal (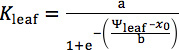 ), logistic (
), logistic (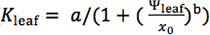 ) and exponential (
) and exponential (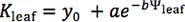 ).
).
3. Representative Results
The decline of leaf hydraulic conductance with dehydration varies strongly across species (Fig. 1). Drought sensitive species experience a stronger decline at less negative water potentials than more drought tolerant species, when measured under high irradiance (PAR > 1,000 μmol • m-2 • s-1). The water potential at 80% loss of hydraulic conductance (P80) for mesic herb Helianthus annuus was 3 MPa less negative than that of Heteromeles arbutifolia, native to California chaparral. Heteromeles arbutifolia responded linearly to the decline in water potential, whereas Helianthus annuus showed a non-linear decline.
Both stomatal conductance (gs) and leaf hydraulic conductance (Kleaf) can also respond dramatically to irradiance (Fig. 2). For Raphiolepis indica, the stomatal conductance declined strongly with dehydration for leaves measured under high irradiance (PAR > 1,000 μmol • m-2 • s-1) and under lab light (PAR < 6 μmol • m-2 • s-1). Under high irradiance, leaves measured with the EFM that had been dehydrated to the water potential that is typical for well-lit leaves on intact plants at mid-day showed similar gs as for leaves measured on intact plants with a porometer (star in Fig. 2). Notably, for leaves at maximum hydration, Kleaf was four-fold higher for leaves exposed to high than low irradiance (11 vs 2.8 mmol m-2 s-1 MPa-1), and therefore Kleaf showed a stronger light response in well hydrated leaves than gs. Further, under high irradiance, Kleaf declined more strongly with dehydration. The P80 was 0.93 MPa less negative under high than low irradiance.
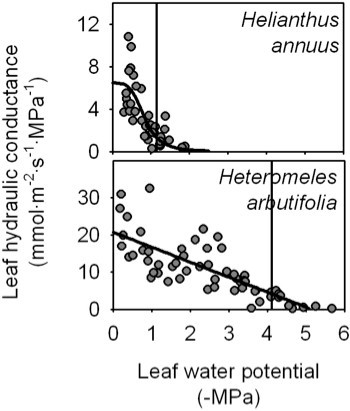 Figure 1. Leaf hydraulic vulnerability curves for a drought sensitive (top) and a drought tolerant (bottom) species (from 14,24 ). Each point represents a single measurement of leaf hydraulic conductance (Kleaf) on a detached leaf using the evaporative flux method (EFM). The vertical bars represent the water potential at which the species lost 80% of initial leaf hydraulic conductance (P80). Best fit functions using maximum likelihood are plotted for both species.
Figure 1. Leaf hydraulic vulnerability curves for a drought sensitive (top) and a drought tolerant (bottom) species (from 14,24 ). Each point represents a single measurement of leaf hydraulic conductance (Kleaf) on a detached leaf using the evaporative flux method (EFM). The vertical bars represent the water potential at which the species lost 80% of initial leaf hydraulic conductance (P80). Best fit functions using maximum likelihood are plotted for both species.
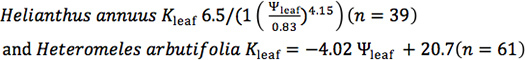
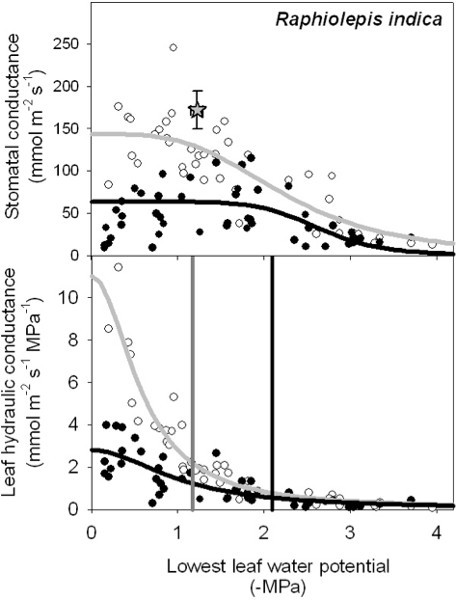 Figure 2. The response of stomatal conductance (gs) and of leaf hydraulic conductance (Kleaf) to declining leaf water potential for Raphiolepis indica measured under high vs low irradiance (from 16). (PAR > 1,000 and < 6 mmol • m-2 • s-1). Each point represents a single measurement on a detached leaf using the evaporative flux method (EFM), with white and black dots representing leaves measured under high and low irradiance respectively. Equations for stomatal conductance curves in high vs low irradiance:
Figure 2. The response of stomatal conductance (gs) and of leaf hydraulic conductance (Kleaf) to declining leaf water potential for Raphiolepis indica measured under high vs low irradiance (from 16). (PAR > 1,000 and < 6 mmol • m-2 • s-1). Each point represents a single measurement on a detached leaf using the evaporative flux method (EFM), with white and black dots representing leaves measured under high and low irradiance respectively. Equations for stomatal conductance curves in high vs low irradiance: ![]() and
and ![]() . Equations for vulnerability curves in high vs. low irradiance:
. Equations for vulnerability curves in high vs. low irradiance: ![]() and
and ![]() . The grey star in the upper panel represents the mean ± standard error for gs and leaf water potential measured on leaves of intact plants at mid-day with a porometer. Grey and black vertical bars in the lower panel represent the water potential at which the species lost 80% of initial leaf hydraulic conductance (P80) under high and low irradiance respectively.
. The grey star in the upper panel represents the mean ± standard error for gs and leaf water potential measured on leaves of intact plants at mid-day with a porometer. Grey and black vertical bars in the lower panel represent the water potential at which the species lost 80% of initial leaf hydraulic conductance (P80) under high and low irradiance respectively.
Discussion
The evaporative flux method presented here allows relatively rapid (30 min) determination of leaf hydraulic conductance in the laboratory with simultaneous measurement of stomatal conductance.
The EFM is so far the method that most closely follows the natural pathway of water in leaves, given that water evaporates in the leaf airspaces and diffuses from the stomata15. A number of other experimental methods for Kleaf determination exist25, with three being especially common. (1) In the high pressure flow method (HPFM), water is pushed through the leaf under high pressure15,26 . However, vulnerability curves cannot be obtained with this method because applied positive pressures may refill emboli and rehydrate mesophyll tissue. (2) In the rehydration kinetics method (RKM), Kleaf is calculated using the analogy between the rehydration of dehydrated leaves with the charging of a capacitor in series with a resistors 27,28. This method is rapid, but does not reproduce the entire system of natural transpirational pathways in the leaf and is not easily modified to allow measurement of leaves acclimated to high irradiance. (3) In the vacuum pump method (VPM) the leaf is placed in a chamber, its petiole connected to a water source on a balance, while vacuums of different intensities are applied to the leaf driving water loss from the leaf15,26 . This method can be time consuming. Measurements using all three methods have been found to give consistent results15,28,29 , and thus the EFM is frequently used given its being based on transpirational water movement, and its relative rapidity, and its modification for measuring Kleaf under different conditions.
All of the typical methods used for determining Kleaf involve a level of uncertainty in quantifying accurately the driving force corresponding to transpiration25. In the EFM, the ΔΨleaf used to calculate Kleaf is lower than the true driving force, as it is based on the bulk Ψleaf after the leaf is equilibrated for the pressure bomb. At this point, the bulk Ψleaf does not necessarily represent the water potential of the cells at the end of the hydraulic pathway, where the water evaporates, but rather a volume-weighted average for the relatively few cells of low water potential where water is moving and evaporating, and for the more numerous cells of high water potential that may be isolated from the transpiration stream. How much the Ψleaf differs from the true driving force thus depends on where the water is evaporating in the leaf. If water evaporates throughout the leaf, the bulk Ψleaf may be close to the true driving force, and if water evaporates only from relatively few cells near the stomata, the the bulk Ψleaf may substantially underestimate the true driving force, i.e., the water potential of those cells may in fact be much lower, and thus actual Kleaf would be lower than that determined with the EFM. Thus, the degree that Kleaf determined by the EFM represents the true Kleaf is sensitive to the water flow pathways, which are still not well understood. While the EFM provides data that match those of other methods for measuring Kleaf (see above), and is excellent for comparative use, the possibility of species-variation in water flow pathways through the mesophyll needs further investigation. Indeed, even for leaves of given species, these water pathways might vary under different conditions, e.g., under different irradiances and heat loads4,30 or of different water status (given tissue shrinkage)31. Further knowledge of the water transport pathways would thus allow the EFM to be used to resolve the effects of the changing pathways on overall leaf hydraulic transport.
The measurement of stomatal conductance in the EFM has the advantages of allowing determination of responses to irradiance and dehydration, and providing matched responses of gs with Kleaf. Additionally, the effect of previous leaf dehydration followed by rehydration on gs can be assessed with this method. However, this method has two potential drawbacks. First, this gs measurement is made on an excised leaf, rather than on an intact plant, and second, the determination of stomatal conductance is based on climate measurement some distance from the leaf, rather than close to the leaf as in a porometer chamber, though the ventilation of leaf with the fan equilibrates the leaf surface with surrounding air. For some species, excised leaves may show different stomatal responses than for leaves on an intact plant 16,32 . However, for several species gs estimated using the EFM was similar to values measured on intact plants with a porometer for leaves at mid-day Ψleaf16 (Fig. 2).
The EFM can be applied for physiological insight, i.e., investigating the dynamics of Kleaf, its basis in structure and anatomy, and its responses to environmental factors. Additionally, the method can be used to compare plants of given species grown under different conditions, or of different ages, or to compare plants of different species adapted to different environments.
By constructing leaf vulnerability curves, one can gain insight into the mechanisms of species tolerances to drought. During drought, cavitation and cell collapse have been shown to occur in the leaf xylem of some species, and cell shrinkage in the mesophyll during dehydration could also impact water movement (reviewed in 14). Future studies will be needed to determine the relative importance of each of these processes in driving the decline of Kleaf and stomatal closure.
Irradiance is also a strong factor influencing Kleaf and the EFM is highly suited for measurements under high irradiance. The combined response of aquaporins to irradiance and drought should gain much interest. High irradiance leads to increased expression and/or activation of aquaporins that permit faster water flow through living cells, increasing Kleaf, whereas dehydration leads to reduced expression and/or de-activatation of aquaporins, reducing Kleaf. The combined impacts of irradiance and water supply on Kleaf require further investigation4,10,16,33 .
Further, across species, Kleaf and its vulnerability has been related to species distributions with respect to the supply of light water and other resources3,13,24 . Species from dry areas tend to show greater resistance to hydraulic decline, facing more negative soil water potentials, and by maintaining a high Kleaf at more negative water potentials, they can maintain high gs and continue capturing CO2 for photosynthesis13,14. Thus, the study of leaf hydraulic properties provides insights at the level of cell, leaf, and whole plant and its responses to the environment, and is likely to yield numerous important new discoveries at all scales.
Disclosures
No conflicts of interest declared.
Acknowledgments
We are grateful to four anonymous reviewers for improving the manuscript, and to the National Science Foundation (Grant IOS-0546784) for support.
References
- Tyree MT, Zimmermann MH. Xylem Structure and the Ascent of Sap. Springer; 2002. [Google Scholar]
- Lawlor DW, Cornic G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002;25:275–294. doi: 10.1046/j.0016-8025.2001.00814.x. [DOI] [PubMed] [Google Scholar]
- Sack L, Holbrook NM. Leaf hydraulics. Annual Review of Plant Biology. 2006;57:361–381. doi: 10.1146/annurev.arplant.56.032604.144141. [DOI] [PubMed] [Google Scholar]
- Cochard H, et al. Putative role of aquaporins in variable hydraulic conductance of leaves in response to light. Plant Physiol. 2007;143:122–133. doi: 10.1104/pp.106.090092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson DM, Meinzer FC, Woodruff DR, McCulloh KA. Leaf xylem embolism, detected acoustically and by cryo-SEM, corresponds to decreases in leaf hydraulic conductance in four evergreen species. Plant Cell Environ. 2009;32:828–836. doi: 10.1111/j.1365-3040.2009.01961.x. [DOI] [PubMed] [Google Scholar]
- Cochard H, Froux F, Mayr FFS, Coutand C. Xylem wall collapse in water-stressed pine needles. Plant Physiol. 2004;134:401–408. doi: 10.1104/pp.103.028357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blackman CJ, Brodribb TJ, Jordan GJ. Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms. New Phytol. 2010;188:1113–1123. doi: 10.1111/j.1469-8137.2010.03439.x. [DOI] [PubMed] [Google Scholar]
- Nardini A, Salleo S, Raimondo F. Changes in leaf hydraulic conductance correlate with leaf vein embolism in Cercis siliquastrum L. Trees-- Structure and Function. 2003;17:529–534. [Google Scholar]
- Trifilo P, Nardini A, Lo Gullo HA, Salleo S. Vein cavitation and stomatal behaviour of sunflower (Helianthus annuus) leaves under water limitation. Physiol. Plant. 2003;119:409–417. [Google Scholar]
- Shatil-Cohen A, Attia Z, Moshelion M. Bundle-sheath cell regulation of xylem-mesophyll water transport via aquaporins under drought stress: a target of xylem-borne ABA. Plant J. 2011;67:72–80. doi: 10.1111/j.1365-313X.2011.04576.x. [DOI] [PubMed] [Google Scholar]
- Engelbrecht BMJ, Velez V, Tyree MT. Hydraulic conductance of two co-occuring neotropical understory shrubs with different habitat preferences. Ann. For. Sci. 2000;57:201–208. [Google Scholar]
- Sheffield J, Wood EF. Projected changes in drought occurrence under future global warming from multi-model multi-scenario, IPCC AR4 simulations. Climate Dynamics. 2008;31:79–105. [Google Scholar]
- Blackman CJ, Brodribb TJ, Jordan GJ. Leaf hydraulic vulnerability influences species' bioclimatic limits in a diverse group of woody angiosperms. Oecologia. 2012;168:1–10. doi: 10.1007/s00442-011-2064-3. [DOI] [PubMed] [Google Scholar]
- Scoffoni C, McKown AD, Rawls M, Sack L. Dynamics of leaf hydraulic conductance with water status: quantification and analysis of species differences under steady-state. J. Exp. Bot. 2012;63:643–658. doi: 10.1093/jxb/err270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sack L, Melcher PJ, Zwieniecki MA, Holbrook NM. The hydraulic conductance of the angiosperm leaf lamina: a comparison of three measurement methods. J. Exp. Bot. 2002;53:2177–2184. doi: 10.1093/jxb/erf069. [DOI] [PubMed] [Google Scholar]
- Guyot G, Scoffoni C, Sack L. Combined impacts of irradiance and dehydration on leaf hydraulic conductance: insights into vulnerability and stomatal control. Plant, Cell & Environment. 2012;35:857–871. doi: 10.1111/j.1365-3040.2011.02458.x. [DOI] [PubMed] [Google Scholar]
- Scoffoni C, Pou A, Aasamaa K, Sack L. The rapid light response of leaf hydraulic conductance: new evidence from two experimental methods. Plant Cell Environ. 2008;31:1803–1812. doi: 10.1111/j.1365-3040.2008.01884.x. [DOI] [PubMed] [Google Scholar]
- Pasquet-Kok J, Creese C. Turning over a new "leaf": multiple functional significances of leaves versus phyllodes in Hawaiian Acacia koa. Plant, Cell & Environment. 2010;33:2084–2100. doi: 10.1111/j.1365-3040.2010.02207.x. [DOI] [PubMed] [Google Scholar]
- Weast RC. Handbook of Chemistry and Physics. 54th. CRC Press; 1974. [Google Scholar]
- Yang SD, Tyree MT. Hydraulic resistance in Acer saccharum shoots and its influence on leaf water potential and transpiration. Tree Physiol. 1993;12:231–242. doi: 10.1093/treephys/12.3.231. [DOI] [PubMed] [Google Scholar]
- Pearcy RW, Schulze E-D, Zimmermann R. In: Plant Physiological Ecology: Field Methods and Instrumentation. Pearcy RW, Ehleringer JR, Mooney HA, Rundel PW, editors. Kluwer Academic; 2000. pp. 137–160. [Google Scholar]
- Buck AL. New equations for computing vapor-pressure and enhancement factor. Journal of Applied Meteorology. 1981;20:1527–1532. [Google Scholar]
- Sokal RR, Rohlf FJ, editors. Biometry. 3rd. Freeman, W.H; 1995. [Google Scholar]
- Scoffoni C, Rawls M, McKown A, Cochard H, Sack L. Decline of leaf hydraulic conductance with dehydration: relationship to leaf size and venation architecture. Plant Physiol. 2011;156:832–843. doi: 10.1104/pp.111.173856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sack L, Tyree MT. In: Vascular Transport in Plants. Holbrook NM, Zweiniecki MA, editors. Elsevier/Academic Press; 2005. [Google Scholar]
- Nardini A, Tyree MT, Salleo S. Xylem cavitation in the leaf of Prunus laurocerasus and its impact on leaf hydraulics. Plant Physiol. 2001;125:1700–1709. doi: 10.1104/pp.125.4.1700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodribb TJ, Holbrook NM. Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol. 2003;132:2166–2173. doi: 10.1104/pp.103.023879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blackman CJ, Brodribb TJ. Two measures of leaf capacitance: insights into the water transport pathway and hydraulic conductance in leaves. Funct. Plant Biol. 2011;38:118–126. doi: 10.1071/FP10183. [DOI] [PubMed] [Google Scholar]
- Nardini A, Raimondo F, Lo Gullo MA, Salleo S. Leafminers help us understand leaf hydraulic design. Plant Cell Environ. 2010;33:1091–1100. doi: 10.1111/j.1365-3040.2010.02131.x. [DOI] [PubMed] [Google Scholar]
- Sheriff DW. Evaporation sites and distillation in leaves. Ann. Bot. 1977;41:1081–1082. [Google Scholar]
- Canny MJ, Huang CX. Leaf water content and palisade cell size. New Phytol. 2006;170:75–85. doi: 10.1111/j.1469-8137.2005.01633.x. [DOI] [PubMed] [Google Scholar]
- Santiago LS, Mulkey SS. A test of gas exchange measurements on excised canopy branches of ten tropical tree species. Photosynthetica. 2003;41:343–347. [Google Scholar]
- Sack L, Cowan PD, Jaikumar N, Holbrook NM. The 'hydrology' of leaves: co-ordination of structure and function in temperate woody species. Plant Cell Environ. 2003;26:1343–1356. [Google Scholar]