

Abstract
EfeUOB-like tripartite systems are widespread in bacteria and in many cases they are encoded by genes organized into iron-regulated operons. They consist of: EfeU, a protein similar to the yeast iron permease Ftrp1; EfeO, an extracytoplasmic protein of unknown function and EfeB, also an extracytoplasmic protein with heme peroxidase activity, belonging to the DyP family. Many bacterial EfeUOB systems have been implicated in iron uptake, but a prefential iron source remains undetermined. Nevertheless, in the case of Escherichia coli, the EfeUOB system has been shown to recognize heme and to allow extracytoplasmic heme iron extraction via a deferrochelation reaction. Given the high level of sequence conservations between EfeUOB orthologs, we hypothesized that heme might be the physiological iron substrate for the other orthologous systems. To test this hypothesis, we undertook characterization of the Staphylococcus aureus FepABC system. Results presented here indicate: i) that the S. aureus FepB protein binds both heme and PPIX with high affinity, like EfeB, the E. coli ortholog; ii) that it has low peroxidase activity, comparable to that of EfeB; iii) that both FepA and FepB drive heme iron utilization, and both are required for this activity and iv) that the E. coli FepA ortholog (EfeO) cannot replace FepA in FepB-driven iron release from heme indicating protein specificity in these activities. Our results show that the function in heme iron extraction is conserved in the two orthologous systems.
Introduction
Exogenous heme is taken up by bacteria as an iron source. Once captured, however, iron must still be extracted from the tetrapyrrole heme molecule. For this purpose, bacteria use heme-degrading enzymes (either orthologs of the mammalian heme oxygenases (HO) or non-HO homologs) that cleave the tetrapyrrole ring [1]. The bound iron atom is released along with biliverdin and carbon monoxide. We have identified two E. coli K12 paralogs, YfeX and EfeB, which also facilitate the release of iron from heme, but in a manner which preserves the tetrapyrrol ring, generating a free iron atom and protoporphyrin IX (PPIX), by a reaction of heme deferrochelation. In an E. coli strain made competent for heme uptake by the expression of the heme outer membrane receptor HasR, either YfeX or EfeB are required for heme-iron extraction, the double mutant yfeX efeB being unable to use heme as an external iron source [2].
YfeX and EfeB proteins are ubiquitous in bacteria and fungi, but are absent in higher eukaryotes. They belong to the DyP heme peroxidase superfamily, which forms a separate family with low sequence similarities to classic fungal and plant heme peroxidases [3]. DyP enzymes oxidize various xenobiotic dyes and cleave dyes like anthraquinones. The bacterial enzymes can be grouped into two subfamilies. One group of bacterial DyP peroxidases consists of cytoplasmic proteins such as Bacteroides thetaiotaomicron BtDyP, Shewanella oneidensis TyrA and E. coli YfeX [4], [5]. Like some plant peroxidases, YfeX and possibly the other DyP peroxidases also oxidize protoporphyrinogen IX and coproporphyrinogen III. These results led to propose that the physiological function of YfeX is to oxidize porphyrins [5]. Be that as it may, plasmid encoded YfeX also complements the E. coli double mutant yfeX efeB and allows the use of heme as an iron source.
The second group is widespread in bacteria and consists of proteins secreted by the Tat pathway and having a twin arginine motif in their signal peptides. The best characterized enzymes of this group are EfeB of E. coli [6], FepB of S. aureus [7] and YwbN of Bacillus subtilis [8]. They are encoded by genes located in one operon repressed by the Fur repressor in the presence of iron. In B. subtilis and S. aureus, tat genes are contiguous to the ywbLMN and fepABC loci [7], [8]. E. coli EfeUOB, B. subtilis YwbLMN and S. aureus FepABC have been implicated in iron transport [7], [10]. These systems operate in an extracytoplasmic space. Although, in E. coli K12, the efeU gene is cryptic due to a single base pair deletion in codon 36, this gene is complete in many other E. coli strains such as E. coli O57:H7. The entire EfeU protein is necessary for iron Fe++ uptake in acidic medium.
What are the specific roles of each protein in iron uptake?
EfeU, FepC and YwbL are integral membrane proteins. They share 25% similarities with the S. cerevisiae Ftr1p ferric permease which constitutes the major yeast iron uptake system [11] (see Fig. 1 for cellular localization of the E. coli EfeUOB and S. aureus FepABC proteins). EfeO orthologs are proteins secreted by the Sec classical pathway. The E. coli EfeO protein is located in the periplasm [6]. B. subtilis YwbM and S. aureus FepA are predicted to be membrane-anchored lipoproteins. The E. coli EfeO consists of two domains, an N-terminal cupredoxin-like domain with a single potential copper binding site, and a C-terminal M75-peptidase domain which contains a conserved potential metal binding site HxxE [12]. FepA and YwbM are smaller, with the signal peptide fused to the M75-peptidase domain. [13]. They share 40% identity with the EfeO M75-peptidase domain. It was suggested that EfeO orthologs function like the multicopper oxidase Fet3p, which is required for Ftr1p activity, enabling ferrous iron oxidation and transfer to iron permease. However, the structures of EfeO-like proteins do not contain bound metal [14], [15]. EfeB orthologs FepB and YwbN share 31% and 36% identity respectively with EfeB. The E. coli EfeB protein binds heme and PPIX [16]. Four EfeB X-ray crystallographic structures have been resolved: apo-EfeB (pdb code: 2Y4D) and EfeB in complex with PPIX (pdb code: 2Y4E), and recently, EfeB bound to heme (pdb code: 2Y4F; 3O72). For other EfeB orthologs, binding to heme and PPIX was not tested. A role in electron shuttling for iron oxidation has been proposed [9].
Figure 1. General organization of the EfeUOB and FepABC systems.

This scheme represents both the genetic (left part of the figure) and envelope organization (right part of the figure) of the S. aureus fepABC and E. coli efeUOB. Orthologous proteins are shown with the same pictogram and the same color. Both operons are regulated by iron loaded Fur repressor. Fur repressor is represented by an arrow with a black circle for iron; FepB and EfeB by a hexagone; FepC and EfeU by a hollow barrel; EfeO by a N-terminal square corresponding to the cupredoxin domain (C) and a C-terminal hexagone corresponding to the M75 peptidase domain; FepA which has only a M75 peptidase domain by a hexagone; heme is represented by a ring and a black circle for iron; HasR, the outer membrane receptor for heme is represented by a barrel; OM for outer membrane; PG for peptidoglycan; IM for inner membrane. Their putative function and cellular localization are shown in the right part of the figure.
Thus, while EfeU orthologs are most likely Fe3+ permeases, the role of the other two proteins (EfeB and EfeO orthologs) in iron transport is less clear. Furthermore, physiological iron sources recognized by these systems have not been determined. Our previous study on the E. coli O57:H7 EfeUOB system, which has a non-cryptic EfeU protein, demonstrated that it promotes iron release from heme in the periplasm and iron transport to the cytoplasm in the absence of heme uptake to the cytoplasm, i.e. in the absence of DppABCDF heme permease [2]. The high number of sequence similarities and the conserved genetic organization of EfeUOB orthologs raised the hypothesis that heme could also be the physiological substrate for EfeUOB orthologs. As all three proteins are translocated across the cytoplasmic membrane, we propose that, in Gram-positive bacteria, these systems allow extraction of iron from heme at the cell surface, followed by uptake of iron. This would satisfy the need for iron without internalization of either heme or PPIX, both of which are toxic molecules. In the present work, we sought to address this question for the S. aureus FepABC system. However, the presence of many heme binding proteins at the cell surface, the absence of information on their interactions with the FepABC system, and the presence of two heme degrading enzymes complicate analysis of S. aureus fepABC mutants. Thus, our first attempt to characterize the S. aureus FepABC system involved the study of its activity in E. coli. We cloned three genes, fepA, fepB and fepC, separately or in various combinations, and we examined their ability to enable heme iron utilization in an E. coli mutant lacking its own systems for extracting iron from heme.
Results presented here indicate that, like EfeB, the S. aureus FepB protein binds both heme and PPIX and has low peroxidase activity. Together, FepA and FepB complement the E. coli ΔyfeX ΔefeUOB double mutant for heme iron utilization and both are required for this activity. The E. coli EfeO cannot replace FepA in FepB-driven iron release from heme. Similarilly, the S. aureus FepA protein cannot replace EfeO in EfeB-driven iron release from heme. Only homologous pairs were functional, indicating protein specificity in these activities. Although our results, in no way, disclose the role of the FepABC system in S. aureus, they nevertheless show that the function in heme iron extraction is conserved in these two orthologous systems.
Materials and Methods
Bacterial Strains and Plasmids
E. coli and S. aureus strains used in this study are listed in Table 1. E. coli K12 strains FB8 (wild type, F-), FB827 (entF::Tn10), JP313 and C600 were from laboratory collection. Strain SET 535 (FB827 pAM-hasR) has been described [2]. SET 536 (FB827 pAM-hasR ΔyfeX::Apra) anδ SET 537 (SET 535 ΔefeUOB::Cm) were constructed in the present work (see below). Strain SET 538 (SET 535 ΔyfeX::Apra ΔefeUOB ::Cm) was constructed in the present work by P1 transduction of ΔyfeX::Apra from SET 536 into SET 537. SET 539 was constructed by P1 transduction of dppF::Km from JWN 3513–1 (dppA::Km) obtained from the KO collection by means of the E. coli database “GenoBase” (http://ecoli.aist-nara.ac.jp/) into SET 537. S. aureus strain subsp. aureus NCTC 8325 (HG001) was used for S. aureus gene amplification. pAM238, pSU19, pAM238-hasR, pBAD24, pBAD24-efeB are from the laboratory collection. pDIA561 is described in [17] and pDIA561-efeO was constructed in our work. pSU19-fepB-6His and pET22b+ fepB-6His (pTE001) and pBAD24-fepA were also constructed here.
Table 1. Bacterial strains and plasmids.
| Strain | Genotype | Reference/source |
| S. aureus | ||
| HG001 | S. aureus strain subsp. aureus NCTC 8325 | [29] |
| E. coli | ||
| JP313 | araD139 ΔlacU169 LAM-flhD5301 fruA25relA1 rpsL150 (strR) rbsR22 deoC1 araΔ714 | [30] |
| BLI-5 | fhuA2 [lon] ompT gal λ(DE3) [dcm] ΔhsdSλ | NEB strain catalogue |
| FB8 | Wild type, F- | [31] |
| FB827 | FB8 entF::Tn10 | [31] |
| SET 535 | FB827 (pAM-hasR) | [22] |
| SET 536 | SET 535ΔyfeX::Apra | This work |
| SET537 | SET 535ΔefeUOB::Cm | This work |
| SET 538 | SET 535ΔyfeX::Apra ΔefeUOB::Cm | This work |
| SET 539 | SET 535ΔefeUOB::Cm dppF::Km | This work |
| Plasmid | ||
| pAM238-hasR | hasR under plac promoter SpcR | [32] |
| pTE001 | fepB under plac promoter in pET22b+ | This work |
| pET22b+ | Qiagen | |
| pSU19 | [33] | |
| pSU19-fepB-6His | fepB-6His under plac promoter in pSU19 | This work |
| pBAD24-fepA | fepA under para promoter | This work |
| pDIA561 | [17] | |
| pDIA561-efeO | efeO under T3 promoter CmR | This work |
| pTX15 | Shuttle vector used to clone fepABC genes under pxyl promoter | [18] |
| pMD1-fepB | fepB under pxyl promoter AmpR | This work |
| pMD1-fepAB | fepAB under pxyl promoter | This work |
| pMD1-fepA | fepA under pxyl promoter | This work |
| pMD1-fepABC | fepABC under pxyl promoter | This work |
pTX15 is an E. coli/Staphylococcus shuttle vector carrying Amp and Ery resistance genes [18]. It carries the XylR repressor and the XylA promoter (pxyl), along with the intergenic XylR operator. The cloned genes are repressed by XylR and transcription is induced upon xylose addition. pMD1-fepABC, pMD1-fepAB, pMD1-fepA and pMD1-fepB were constructed in the present work.
Media and Growth Conditions
Hemin (>90% pure), bovine hemoglobin (Hb) and 2,2′dipyridyl (Dip) were obtained from Sigma Chemical Company (Lyon, France). Protoporphyrin IX (PPIX) was purchased from Frontier Scientifc (Carnforth, United Kingdom). Bacteria were grown aerobically at 37°C or 30°C in LB medium, in M63 or M63 without added iron salt (M63*). All minimal media were supplemented with 0.4% glucose (glu) or glycerol (gly). For arabinose induction, 0.2% L-arabinose (ara) was added to induce the pBAD24 promoter. For xylose induction, 0.2% or 2% xylose (xyl) was added to induce the pxyl promoter. When required, Dip was added to a final concentration of 100 µM to M63* (M63* Dip). Antibiotics were added at the usual concentrations for E. coli.
Growth Promotion Assays
Cultures of strain SET 538 carrying various plasmids were grown in M63 gly to an OD600 of 1, and 100 µl aliquots were mixed with 3.5 ml of M63* soft agar (0.7% agar) and poured onto M63* Dip plates supplemented with xyl 0.2% or 2% or ara 0.2% to induce the pBAD24 and/or pMD1 encoded genes. Aliquots of 50 µl of bovine hemoglobin at various concentrations calculated on the basis of the heme monomer were provided in wells punched into solidified agar. Solutions of hemin and oxidized bovine hemoglobin (in which heme is weakly bound to globin) gave similar growth promotion results. Plates were incubated for 48 h at 37°C and photographed. The growth halo diameter in mm around the wells was measured.
Genetic and Molecular Biology Techniques
Mutants ΔefeUOB::Cm and ΔyfeX::Apra were constructed by replacing the entire efeUOB operon or the yfeX gene respectively with an antibiotic cassette as previously described [19]. These mutations were subsequently transduced into SET 535 derivative strains using the P1vir phage. Verification of deletions was done by PCR. S. aureus strain subsp. aureus NCTC 8325 (HG 001) was used for amplification of genes SAOUHSC 00325 (fepA), SAOUHSC 00326 (fepB), SAOUHSC 00327(fepC) with complementary oligonucleotides. Amplified fragments were inserted into pTX15 digested with BamH1 and EcoR1, giving plasmids pMD1-fepABC, pMD1-fepAB, pMD1-fepA and pMD1-fepB carrying the corresponding genes under the pxyl promoter and repressed by XylR. Full induction was obtained by addition of xyl 2% to the medium. The FepA potential iron binding sequence HKIE was mutagenized into AKIA on the plasmid pMD1-fepAB by a two-step PCR leading to plasmid pMD1-fepAAKIA-fepB. Verification of the DNA change was done by sequencing. The fepA gene was also cloned in pBAD24 under an arabinose promoter. The fepB gene without its signal sequence was amplified by PCR with genomic DNA from S. aureus NCTC 8325 using primer pairs fepB-PS-NdeI/XhoI-fepB. The PCR product was cloned at the NdeI and XhoI sites of vector pET22b+ (Novagen) containing a T7 promoter and a His-tag. Resulting plasmid pET22b-fepB-6His was used for transforming E. coli strain BLI5 for expression. The fepB-6His without its signal sequence was also recloned in pSU19. The plasmid carrying efeO was constructed by amplification of MG1655 genomic DNA using complementary oligonucleotides. The amplified fragment was inserted into pDIA561 cut with EcoR1 and BamH1, thus resulting in pDIA561-efeO. All amplified genes were checked by DNA sequencing. All plasmids used in this study are liste in Table 1. Oligonucleotides used for each cloning are available on demand.
FepB-6His Purification
Strain JP313 (pET22b-fepB-6His) was grown in M63 gly Amp medium at 30°C. When the culture reached OD600 = 0.6, 0.5 mM isopropylthiogalactopyranoside (IPTG) was added to the medium and growth was pursued for 2 h. Cells were harvested by centrifugation for 20 min at 5,000 g at 4°C. The cell pellet was resuspended in 50 mM Tris-HCl pH 8.0, 0.3 M NaCl and 15 mM imidazole and disrupted by sonication. After centrifugation at 20,000 g for 60 min at 4°C, soluble extract was purified by nickel-nitrilotriacetic acid affinity chromatography using the QIA express system. Fractions containing FepB were pooled and then subjected to Ultrogel AcA54 gel filtration chromatography (110 cm×1.1 cm) in 50 mM Tris-HCl pH 8.0, 0.2 M NaCl. The purified protein was analyzed by SDS polyacrylamide gel electrophoresis (SDS-PAGE) followed by Coomassie blue staining for purity testing and stored at −20°C. FepB-6His was also purified from cultures of SET538 strains carrying either pSU19-fepB-6His or pSU19-fepB-6His and pBAD24-fepA, as above, by nickel-nitrilotriacetic acid affinity chromatography.
Preparation of Membrane, Cytoplasmic and Periplasmic Fractions and Protein Analysis
SET 538 strains harboring pMD1-fepABC, pMD1-fepAB or pMD1-fepB were grown in M63 gly supplemented with xyl at two concentrations, 0.2 and 2% at 37°C. At OD600 = 1, cells were harvested by centrifugation for 10 min at 5,000 g at 4°C and were washed once in 50 mM Tris-HCl pH 7.5. Each pellet was resuspended in 500 µl of 50 mM Tris-HCl pH 7.5 and cells were broken by sonication. After centrifugation for 10 min at 5,000 g at 4°C to remove unbroken cells and aggregates, preparations corresponding to whole cell extracts were resuspended in SDS sample buffer for SDS-PAGE. The periplasmic fraction was prepared from washed intact cells by osmotic shock [20]. Spheroplasts resulting from osmotic shock were broken by sonication and centrifuged for 30 min at 15,000 g at 4°C to separate the cytoplasmic fraction from the whole membrane fraction [20]. Proteins present in the various samples were analyzed by SDS-PAGE followed by immunodetection with anti-FepB and anti-maltose binding protein (MBP) antibodies.
Antibody Preparation
Rabbit anti-MBP antibodies are from the laboratory collection and were used at a 1/1,000 dilution for immunoblot. For anti-FepB rabbit antibodies, the entire fepB gene (with its signal sequence) was cloned into pBAD24 digested with EcoR1 and Pst1 and expressed in JP313. After arabinose induction, overnight cultures of JP313 pBAD24-fepB were centrifuged to harvest cells. The cell pellet was disrupted by sonication. Inclusion bodies were prepared as previously described and solubilized with 50 mM Tris-HCl pH 8,0, 10 mM EDTA, 100 mM NaCl containing urea 8 M buffer. After SDS-PAGE, the band corresponding to FepB was cut and an aliquot was migrated on SDS-PAGE for determination of the N-terminal sequence by the Plateforme d’Analyse et de Microsequence des Protéines of the Institut Pasteur. It corresponded mainly to the N-terminus of the FepB precursor (MTNYE). The protein preparation was mixed with a Titer Max adjuvant and injected into a rabbit to raise antibodies. The serum was used at a 1/1,000 dilution for immunoblot.
Electrophoresis and Western Blot
SDS-PAGE and western blot were carried out according to standard protocols. Bound antibodies were detected with secondary anti-rabbit antibodies coupled with horseradish peroxidase revealed using a chemiluminescence system.
FepB Binding Studies
PPIX and hemin binding studies were determined as described in a previous report [2]. Experiments were carried out at room temperature. Apo-FepB was in 50 mM Tris-HCl pH 8.0, 0.2 M NaCl. Taking into account the expected molar extinction coefficient around 75000–100000 M−1 cm−1 of heme-FepB complexes (measured at 404 nm) or of PPIX-FepB complexes (measured at 407 nm), we used a protein concentration of 2.2 µM for both titration experiments. Hemin was dissolved in a solution of 0.1 M NaOH and then diluted to 100 µM with the Tris buffer. PPIX dissolved in DMSO was diluted to 500 µM in DMSO. Aliquots of either hemin or PPIX solutions were successively added to cell containing 0.5 ml of the apoprotein. Absorption spectra were recorded 5 min after each hemin or PPIX addition in a 1 cm path lengh cell on a Beckman DU 800 spectrophotometer and were followed by measuring the absorbance from 250 to 750 nm. Absorbance at the Soret bands (404 nm for hemin and 407 nm for PPIX) were reported as a function of hemin and PPIX concentration, corrected for absorbance of free heme and free PPIX.
To determine the Kd for hemin and PPIX, titration curves could be fitted according to a one-site binding model using FigP software (Biosoft Cambridge, U.K.) and the following equation:
where A is the absorbance, a M the molar extinction coefficient of bound ligand, [L]0 the total concentration of ligand, [P]t the total effective concentration of binding sites, and Kd the equilibrium dissociation constant.
Peroxidase Activity
The peroxidase activity of recombinant FepB was defined using 10 mM catechol or 1.5 mM ABTS (2,2′-azino-bis (3-ethylbenzthiazoline-6-sulfonic acid)) as a substrate in the reaction system containing 0.2 mM H2O2. The reaction was started by addition of H2O2 and followed spectrophotometrically at room temperature. One unit corresponds to 1 µmol of substrate oxidized per min in 50 mM citrate buffer pH 3.3 or 4.7. The wavelengths and absorption coefficients used were: catechol, 480 = 1 mM−1 cm−1 and ABTS, 436 = 29,3 mM−1 cm−1. Control reactions were included without enzyme, H2O2, or both.
Results
Expression, Purification and Binding Properties of Recombinant FepB-6His
The fepB gene encoding a protein lacking its N-terminal signal peptide was cloned into pET22b in frame with the sequence encoding six histidines. The recombinant FepB-6His was purified as described in Materials and Methods. The protein was 95% pure as evaluated by SDS PAGE. The UV-visible absorption spectrum of FepB solution, slightly reddish, showed the presence of a heme Soret band at 404 nm, indicating that the protein was at least partially loaded with heme. Next, gel filtration chromatography allowed separation into two peaks, one loaded with heme (dimeric) and a second containing the unloaded protein (apo-FepB, monomeric). The latter was used to determine affinity constants for hemin and PPIX. Addition of hemin or PPIX to FepB led different Soret band at 404 nm for hemin and at 407 nm for PPIX (Fig. 2A, 2C). Titration curves shown in Fig. 2B and 2D indicate that FepB binds hemin and PPIX each with stoechiometry of about 1 (0.9 and 0.7 respectively, suggesting that a part of FepB molecule is unfolded or denaturated) and with a Kd of 13.3±2.7 nM for heme and 21.1±5,2 nM for PPIX. Thus, like YfeX [2] and EfeB [16], FepB has high affinities for heme and PPIX.
Figure 2. FepB-6His characterization.

A and C: UV-visible spectra of apo-FepB (gray line) and FepB-hemin or FepB-PPIX complex of 2.2 mM (black line). B and D: Spectroscopic measure of hemin and PPIX binding to apo-FepB-6His. Aliquots of hemin or PPIX solutions were successively added to 0.5 ml of apo-FepB-6His (2.22 µM). Absorbance at the Soret bands (404 nm and 407 nm for hemin and PPIX respectively) were reported versus porphyrin concentration corrected for absorbance of free porphyrin at the same concentration. Experiments were done 5 times giving similar curves.
Peroxidase Activity of Recombinant FepB-6His
The peroxidase activity of FepB-6His was measured on two typical peroxidase substrates, ABTS and catechol, using the method described for EfeB [6], [16]. Table 2 shows that FepB has weak peroxidase activity on both substrates in comparison with HRP, the secreted plant peroxidase belonging to another group of heme peroxidases. FepB-6His peroxidase activity increases at acidic pH, as it is the case for EfeB [16].
Table 2. Peroxidase activity.
| Specific activity (U mg−1)a | |||||
| FepB | HRP | ||||
| Electron donor | Concentration (mM) | pH 3.3 | pH 4.7 | pH 3.3 | pH 4.7 |
| Catechol | 10 | 0.30±0.04 | nd | 179.1±5.0 | 413.1±35.6 |
| ABTS | 1.5 | 39.4±6.3 | 0.70±0.05 | 1.32±0.12 | 420±6.6 |
Substrate specificity of recombinant FepB and HRP.
Activity was calculated as specific activity in U mg−1 (1U = 1 µmol/min).
nd : not detectable.
Activity of the FepABC Cluster in E. coli
In order to determine whether the S. aureus fepABC cluster plays a role in heme iron extraction, we cloned it in the pTX15 vector, leading to plasmid pMD1 carrying fepABC genes under control of pxyl; we used the previously developed heme iron acquisition test [2]. The test is based on growth assay of the E. coli FB827 (pAM238-hasR) strain (SET 535) in iron-chelated medium in the presence of hemoglobin as the iron source. This strain lacks enterobactin, the major E. coli siderophore, and is thus easy to deprive of iron. It uptakes heme through the outer membrane receptor HasR and transports it through the inner membrane by the heme/dipeptide permease DppABCDF. The efeU gene is cryptic in E. coli K12, and iron extraction from heme takes place in the cytoplasm and is achieved by either YfeX or EfeB activity (though normally exported to the periplasm, a part of EfeB is active in the cytoplasm (17)). The E. coli mutant lacking these two activities, FB827 ΔyfeX::Apra.
ΔefeΥOB::Cm (pAM238-hasR) (strain SET 538), cannot grow in iron-chelated medium (M63 gly* Dip agar) in the presence of heme.
We tested whether expression of fepABC genes can complement ΔyfeX::Apra ΔefeUOB::Cm mutations of SET 538 for heme iron utilization. SET 538 strains harboring either pMD1-fepABC, pMD1-fepAB, pMD1-fepA or pMD1-fepB were grown in M63 gly supplemented with xyl at two concentrations, 0.2 and 2%. Expression of pMD1-fepB or pMD1-fepA in SET 538 did not restore the ability of the strain to grow on M63 gly* Dip agar around hemoglobin-containing wells (Fig. 3 and Table 3). In contrast, pMD1-fepABC and pMD1-fepAB restored the ability of the strain to grow around hemoglobin-containing wells only when xyl was added to induce the fepABC or fepAB genes (Fig. 3 and Table 3). Both xylose concentrations tested enabled similar growth.
Figure 3. Heme iron acquistion plates.

Growth around hemoglobin-containing wells on Petri dishes seeded with strain SET 538 carrying the indicated plasmids was photographed after 48 h incubation at 37°C. Wells were filled with 50 µl hemoglobin at the following concentrations in µM. Top left: 50; top right: 10; bottom left: 5 and bottom right: 1.
Table 3. FepABC role in heme acquisition.
| Growth halo diameter (mm) | |||||
| Strain name | Relevant mutation | 50 µM | 10 µM | 5 µM | 1 µM |
| SET 535 | none | 28±2 | 20±2 | 14±2 | 12±2 |
| SET 536 | ΔyfeX::Apra | 19±2 | 15±2 | ndg | ndg |
| SET 537 | ΔefeUOB::Cm | 19±2 | 13±2 | ndg | ndg |
| SET 538 | ΔyfeX::ApraΔefeUOB::Cm | ndg | ndg | ndg | ndg |
| SET 538 (pMD1) | ΔyfeX::ApraΔefeUOB::Cm | ndg | ndg | ndg | ndg |
| SET 538 (pMD1-fepB) | ΔyfeX::ApraΔefeUOB::Cm | ndg | ndg | ndg | ndg |
| SET 538 (pMD1-fepA) | ΔyfeX::ApraΔefeUOB::Cm | ndg | ndg | ndg | ndg |
| SET 538 (pMD1-fepAB) | ΔyfeX::ApraΔefeUOB::Cm | 25±2 | 22±2 | 17±2 | 11±2 |
| SET 538 (pMD1-fepABC) | ΔyfeX::ApraΔefeUOB::Cm | 23±2 | 22±2 | 17±2 | 11±2 |
| SET 538 (pSU19-fepB-6His), | ΔyfeX::ApraΔefeUOB::Cm | ndg | ndg | ndg | ndg |
| SET 538 (pSU19-fepB-6His)(pBAD24-fepA) | ΔyfeX::ApraΔefeUOB::Cm | 25±2 | 22±2 | 17±2 | 11±2 |
| SET 538 (pMD1-fepAAkiA-B) | ΔyfeX::ApraΔefeUOB::Cm | 25±2 | 22±2 | 17±2 | 11±2 |
| SET 538 (pBAD24-efeB) | ΔyfeX::ApraΔefeUOB::Cm | 17±2 | 15±2 | ndg | ndg |
| SET 538 (pBAD24-efeB)(pDIA561-efeO) | ΔyfeX::ApraΔefeUOB::Cm | 20±2 | 19±2 | 17±2 | 14±2 |
| SET 538 (pMD1-fepB)(pDIA561-efeO) | ΔyfeX::ApraΔefeUOB::Cm | ndg | ndg | ndg | ndg |
| SET 539 (pMD1-fepAB) | ΔefeUOB::CmΔdppF::Km | ndg | ndg | ndg | ndg |
| SET 539(pMD1- fepABC) | ΔefeUOB::CmΔdppF::Km | ndg | ndg | ndg | ndg |
(a) ndg: no detectable growth around wells.
Names of strains and plasmid-borne complementing genes carried on pDIA561, pBAD24 and pMD1 are indicated in the first column. Aliquots of 50 µl of bovine hemoglobin at various concentrations calculated on the basis of the heme monomer were provided in wells punched into solidified agar. Plates were incubated for 48 h at 37°C and the growth halo diameter in mm around the wells was measured. All experiments were repeated three times.
Activity of the FepABC System in an E. coli Strain Lacking Heme/Dipeptide Dpp Permease
The FepB protein has a typical twin-arginine signal peptide and is exported by the Tat translocon in S. aureus [7]. Sequence analysis indicates that FepA could be a membrane-anchored lipoprotein and FepC an integral membrane protein. In the three plasmids, fepA and fepB genes were cloned with their signal sequences. Since membrane export systems are conserved among bacteria [21], it was possible that S. aureus FepA, FepB and FepC proteins could be translocated across the cytoplasmic membrane when expressed in E. coli. The E. coli O157:H7 EfeUOB system (which has a complete efeU gene) extracts iron from heme in the periplasm, enabling use of heme iron in the absence of DppABCDF heme permease [2], [22], whereas E. coli K12 EfeUOB cannot work in the absence of the Dpp permease because of a mutation in efeU. We sought to determine whether FepABC allows the use of heme iron in the periplasm in the absence of heme uptake to the cytoplasm. Strain FB827 ΔefeUOB::Cm dppF::Km (pAM238-hasR) (strain SET 539) was transformed with pMD1-fepAB and pMD1-fepABC and tested as above for growth around hemoglobin-containing wells. None of the plasmids were able to rescue growth on hemoglobin as iron source (Table 3), indicating that FepA, FepB and FepC proteins were not exported, or else were not functional beyond the cytoplasm. In addition, fepB lacking its signal sequence was recloned in pSU19 (pSU19-fepB-6His) and fepA in pBAD24 a compatible plasmid (pBAD24-fepA). Complementation tests showed that pSU19-fepB-6His and pBAD24-fepA together, but not each plasmid alone, allowed the growth of strain SET 538 on M63 gly* Dip agar around hemoglobin-containing wells, confirming that FepB-6His lacking its signal peptide is active in the cytoplasm (Fig. 3 and Table 3).
Production, Stability and Cellular Localization of the Recombinant FepB Protein in E. coli
The absence of complementation by plasmid pMD1-fepB encoding the FepB protein alone could result from FepB inactivation due to lower production, aggregation or proteolytic degradation. To test this hypothesis, we quantified FepB production and stability in cells carrying the various plasmids. SET 538 strains harboring pMD1-fepB, pMD1-fepAB or pMD1-fepABC were grown under the same conditions as those used for complementation tests, i.e. M63 gly supplemented with xyl at the 2 concentrations, 0.2 and 2%. Total cell extracts were analyzed by western blot with polyclonal anti-FepB antibodies to test whether the three plasmids encoded stable FepB polypeptides. In all of these strains, the FepB protein was inducible by xyl and was immunodetected as a band doublet, which could correspond to precursor and mature forms (Fig. 4A). In fact, the higher form migrates at the same size as the overexpressed recombinant purified FepB protein carrying its entire signal peptide, as confirmed by N-terminal sequencing of this protein giving the MTNYE sequence. The strain harboring pMD1-fepB accumulated approximately 5 times more precursor and mature FepB polypeptides than strains carrying the other two constructs, while all three strains were induced with 2% xyl (Fig. 4A). The strain carrying pMD1-fepB induced with 0.2% xyl accumulated FepB amounts similar to those of strains carrying pMD1-fepAB and pMD1-fepABC induced with 2% xyl. In pMD1-fepB, the fepB gene moved closer to the pxyl promoter than in pMD1-fepAB and pMD1-fepABC, more likely leading to higher FepB expression in the pMD1-fepB plasmid. None of the total cell extracts exhibited cross-reacting material of lower molecular weight in immunoblots, indicating that FepB produced by the three constructs was stable (Fig. 4A). FepB was not immunodetected in pellets of these strains produced by low-speed centrifugation of cells broken by sonication, indicating that FepB did not form aggregates (data not shown). Thus, the absence of heme iron extraction activity of FepB alone was not the result of lower stability, expression or solubility.
Figure 4. FepB production and cellular localization in strain SET 538 carrying various plasmids.
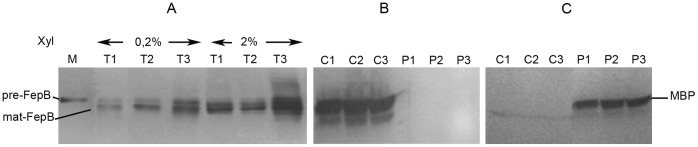
A: FepB immunodetection in total cell pellets: SET 538 strains harboring pMD1-fepABC, pMD1-fepAB or pMD1-fepB were grown at 37°C in M63 gly supplemented with xyl at two concentrations, 0.2 and 2% as indicated on the figure. Cells were harvested when they reached an OD600 = 1. M: FepB precursor proteins purified as inclusion bodies from strain JP313 pBAD24-fepB (the determined amino-terminal sequence was MTNYE); T1: total cell extract of strain SET 538 pMD1-fepABC; T2: total cell extract of strain SET 538 pMD1-fepAB; T3: total cell extract of strain SET 538 pMD1-fepB. Each lane was loaded with 0.5 OD600 cell culture equivalent. B: FepB immunodetection in cytoplasmic and periplasmic fractions: SET 538 strains harboring pMD1-fepABC, pMD1-fepAB or pMD1-fepB were grown in M63 gly supplemented with xyl at 2% for the first two strains and 0.2% concentration for the last strain. Cells were harvested when they reached OD600 = 1. C1: cytoplasmic fraction of strain SET 538 pMD1-fepABC; C2: cytoplasmic fraction of strain SET 538 pMD1-fepAB; C3: cytoplasmic fraction of strain SET 538 pMD1-fepB. P1: periplasmic fraction of strain SET 538 pMD1-fepABC; P2: periplasmic fraction of strain SET 538 pMD1-fepAB; P3: periplasmic fraction of strain SET 538 pMD1-fepB. FepB is not immunodetected in P1, P2 or P3 periplasmic fractions. Each lane was loaded with 0.5 OD600 cell culture equivalent. C: MBP immunodetection in cytoplasmic and periplasmic fractions: The same cytoplasmic (C1, C2, C3) and periplasmic fractions (P1, P2, P3) were immunodetected with anti-MBP antibodies. Each lane was loaded with 0.5 OD600 cell culture equivalent. MBP was not immunodetected in C1, C2 or C3 cytoplasmic fractions, but in P1, P2 and P3 periplasmic fractions of the 3 strains.
To determine the cellular localization of FepB, strain SET 538 carrying either pMD1-fepABC, pMD1-fepAB or pMD1-fepB was grown in M63 gly and induced with xyl 2% for the first two strains and with xyl 0.2% for the final one, to make sure that the various strains produced similar amounts of FepB. Cell fractions were prepared as described in Materials and Methods. FepB was not detected in shock fluid, whereas the maltose binding protein (MBP), a periplasmic marker [23], was entirely detected in the periplasmic fraction (Fig. 4B, C). FepB was not present in the total membrane fraction (data not shown) and was detected only in the cytoplasmic fraction. It migrated as a band doublet (Fig. 3A). Thus, FepB produced by the three plasmids accumulated in the cytoplasm, indicating that, in spite of the Tat signal sequence, it was not efficiently translocated to the periplasm. A significant part of this cytoplasmic FepB was mature in size.
Co-expression of FepA with FepB-6His Increases the apo-FepB to holo-FepB Ratio
In the E. coli strain expressing pSU19-fepB-6His, FepB purified by affinity chromatography was loaded primarily with heme. Since FepA is required for iron extraction activity, we tested whether co-expression of FepA might change the holo-FepB-6His concentration. FepB was purified from cultures of SET 538 strains carrying either pSU19-fepB-6His alone or pSU19-fepB-6His and pBAD24-fepA by nickel affinity chromatography. Absorption spectroscopy showed that FepB purified from a culture producing only FepB was 98% loaded with heme (Fig. S1A), whereas in the presence of FepA, only half of the purified FepB-6His was loaded with heme (Fig. S1B). These experiments were repeated three times giving similar ratios of apo-FepB versus holo-FepB. The remaining FepB did not contain any detectable pigment, as determined by fluorescence spectroscopy, and thus was in the apo form (data not shown). Addition of heme to the FepB protein purified from cells, whether expressing or not expressing FepA, led to its 100% saturation, indicating that the apo-protein is not denatured (data not shown). These results suggest that FepA is required for heme turnover on FepB. However, fluorescence spectroscopic analysis of FepB-6His purified from cells either expressing FepA or not did not reveal any detectable fluorescence. There was no increase in the PPIX concentration in soluble extracts of SET 538 (pSU19-fepB-6His pBAD24-fepA), as measured after pigment separation by HPLC and identification by fluorescence spectroscopy (data not shown). Thus, FepA-FepB activity was too low to drive detectable PPIX accumulation.
Activity of the FepA Mutant Protein and Hybrid Systems Combining FepB and EfeO
The requirement of both FepA and FepB for achieving extraction of iron from heme led us to hypothesize that FepA could be the iron collector. Since FepA has a unique potential metal binding HxxE motif [12], we tested whether this motif was involved in FepA activity. The HKIE sequence was changed into AKIA in pMD1-fepAB and the mutated plasmid was used to transform strain SET 538. The pMD1-fepAAKIA-fepB plasmid rescued growth on hemoglobin as iron source (Table 3), indicating that the HkiE motif is not essential for iron acquisition through FepB. To date, there is no experimental data indicating that the HxxE motif present in EfeO orthologs is involved in binding metal or EfeO functions.
Homologous pairs consisting either of FepA and FepB or of EfeO and EfeB are active in heme iron utilization in E. coli. We thus tested whether hybrid systems consisting of EfeB and FepA or FepB and EfeO are functional. SET 538 strain harboring PMD1-fepB and pDIA561-efeO was grown in M63 gly supplemented with 0.2 or 2% xyl. Expression of EfeO with FepB did not restore growth of strain SET 538 around hemoglobin-containing wells (Table 3), whereas pDIA561-efeO was able to stimulate growth of strain SET 538 carrying pBAD33-efeB, indicating that it is functional. Thus, EfeO cannot replace FepA in FepB-directed iron extraction from heme. This suggests that FepA and FepB are acting in a specific manner.
Discussion
In a first attempt to understand the role of the S. aureus FepABC system, we purified FepB and demonstrated that it binds heme and PPIX in the nanomolar range.
S. aureus fepABC genes were expressed in an E. coli strain deleted for genes allowing heme iron utilization (ΔyfeX ΔefeUOB). The fepABC cluster complemented the double mutant and conferred the ability to use heme as an iron source. We demonstrated that none of the three proteins can complement alone and that FepA and FepB are required for this complementation. FepC, the iron permease, is not required and the FepABC system did not allow heme iron extraction from the periplasm in the absence of the heme/dipeptide permease, suggesting that recombinant FepA and FepB proteins are active in the E. coli cytoplasm. Such activity in the inadequate cellular compartment was also observed for EfeB, which is able to extract iron from the cytoplasmic heme [2]. In a more general manner, Tat-dependent inefficient translocation has often been reported [24]. Cellular localization of the recombinant FepB protein produced in E. coli supported this hypothesis, as it was detected only in the cytoplasm. In addition, cytoplasmic FepB-6His lacking its signal peptide together with FepA were also able to achieve heme iron utilization.
In E. coli cells expressing FepB alone, purified FepB-6His is 98% loaded with heme. On the other hand, when coexpressed with FepA, Fep-6His is about 50% in the apo form, suggesting that FepA is required for heme turnover on FepB. FepB loaded with PPIX was not detected. Roughly similar results were obtained with EfeOB indicating that PPIX remains only weakly bound to the proteins (unpublished results). However, a fraction of EfeB is loaded with PPIX [16]. This fraction was not detected in the case of FepB. It is likely that the FepA-FepB activity is sufficient to meet iron requirements from heme, but not to drive detectable PPIX accumulation.
As a working model, we propose that FepB binds heme and proceeds to reduction of protoporphyrin iron. The molecular mechanism involved in this reaction is not yet elucidated. Nevertheless, the work of Liu et al [16] clearly establishes that several residues involved in peroxidase activity are also required for EfeB-driven protoporphyrin accumulation. Once iron is reduced, it could be captured by FepA. We tested whether the conserved potential metal binding site of FepA is involved in iron capture. The change in this HxxE site into AxxA did not abolish the activity of FepAB proteins in heme iron utilization. Further work is required to determine whether FepA and its orthologs are iron binding proteins. In fact, there exist no experimental data indicating that proteins belonging to the M75 peptidase family bind any metal. Three proteins of this family, AlgP7, EfeM and Bacteroides ovatus PIBO, were purified and crystallized, but no metal was detected in any protein [13], [15]. In addition, the PIBO protein was crystallized in the presence of Mg2+, but the divalent cation was not bound to the potential metal binding site HxxE [15].
We observed protein specificity in FepAB and EfeOB activities, as E. coli EfeO works with E. coli EfeB, but not with S. aureus FepB. This could indicate the existence of specific protein/protein interactions between FepA and FepB. Such complex between the two proteins has not been found biochemically indicating that it might be weak and or transient (unpublished results).
The DyP-type peroxidase family comprises cytoplasmic and extracytoplasmic proteins whose physiological functions remain poorly understood. The ability of YfeX to function as a deferrochelatase remains controversial [2], [5]. Our present work showing that FepA and FepB have the capacity of releasing iron from heme in E. coli indicate a conservation of this activity in DyP peroxidases of distantly related bacterial species.
Obviously, the physiological role of FepABC in S. aureus remains unclear. Results reported on heme acquisition in S. aureus [25], [26] and Listeria monocytogenes [27] indicate that mutations in genes involved in heme uptake have only a weak effect upon heme utilization. Mutations in isdI and isdG genes encoding the two S. aureus heme oxygenases do not eliminate growth on heme as an iron source. The B. subtilis triple mutant lacking hmoA and hmoB genes which encode orthologues of IsdI and IsdG and carrying a deletion of the ywbLMN operon had no observable phenotype on heme as an iron source, suggesting the existence of an additional functional redundancy for this pathway [28].
Thus, further work is required to determine the respective roles of heme transport and heme metabolism in iron acquisition from heme.
Supporting Information
UV-visible spectra of FepB-6His purified by Nickel-nitrilotriacetic acid chromatography. A: FepB-6His purified from SET538 (pSU19-fepB-6His); B: FepB-6His purified from SET538 (pSU19-fepB-6His pBAD24-fepA). Insert: FepB-6His SDS-PAGE analysis. After staining in the Coomassie Blue, gels were digitalized using a JX-330 scanner (Sharp) and the purity of FepB-6His was determined using the Quantity One 1-D software (BioRad).
(TIF)
Acknowledgments
We thank all members of C. W.’s laboratory for insightful discussions. We are grateful to Tarek Msadek and Alain Chaffotte for helpful discussion and critical reading of the manuscript.
Funding Statement
Institut Pasteur, CNRS ERL3526, Agence nationale de la Recherche (ANR Grabiron). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hammer N, Skaar E (2011) Molecular Mechanisms of Staphylococcus aureus Iron Acquisition. Annu Rev Microbiol 65: 129–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Létoffé S, Heuck G, Delepelaire P, Lange N, Wandersman C, et al. (2009) Bacteria capture iron from heme by keeping tetrapyrrol skeleton intact. Proc Natl Acad Sci U S A 106: 11719–11724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Sugano Y (2009) DyP-type peroxidases comprise a novel heme peroxidase family. Cell Mol Life Sci 66: 1387–1403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Zubieta C, Joseph R, Krishna S, McMullan D, Kapoor M, et al. (2007) Identification and structural characterization of heme binding in a novel dye-decolorizing peroxidase, TyrA. Proteins 69: 234–243. [DOI] [PubMed] [Google Scholar]
- 5. Dailey H, Septer A, Daugherty L, Thames D, Gerdes S, et al. (2011) The Escherichia coli protein YfeX functions as a porphyrinogen oxidase, not a heme dechelatase. MBio 2: e00248–00211 doi: 00210.01128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Sturm A, Schierhorn A, Lindenstrauss U, Lilie H, Brüser T (2006) YcdB from Escherichia coli reveals a novel class of Tat-dependently translocated hemoproteins. J Biol Chem 281: 13972–139728. [DOI] [PubMed] [Google Scholar]
- 7. Biswas L, Biswas R, Nerz C, Ohlsen K, Schlag M, et al. (2009) Role of the twin-arginine translocation pathway in Staphylococcus . J Bacteriol 191: 5921–5929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Van der Ploeg R, Mäder U, Homuth G, Schaffer M, Denham E, et al. (2011) Environmental salinity determines the specificity and need for Tat-dependent secretion of the YwbN protein in Bacillus subtilis . PLoS One 6: e18140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Cao J, Woodhall M, Alvarez J, Cartron M, Andrews S (2007) EfeUOB (YcdNOB) is a tripartite, acid-induced and CpxAR-regulated, low-pH Fe2+ transporter that is cryptic in Escherichia coli K12 but functional in E. coli O157: H7. Mol Microbiol 65: 857–875. [DOI] [PubMed] [Google Scholar]
- 10. Grosse C, Scherer J, Koch D, Otto M, Taudte N, et al. (2006) A new ferrous iron-uptake transporter, EfeU (YcdN), from Escherichia coli. . Mol Microbiol 62: 120–131. [DOI] [PubMed] [Google Scholar]
- 11.Severance S, Chakraborty S, Kosman D (2004) The Ftr1p iron permease in the yeast plasma membrane: orientation, topology and structure-function relationships. Biochem J 380 487–496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Rajasekaran M, Nilapwar S, Andrews S, Watson K (2010) EfeO-cupredoxins: major new members of the cupredoxin superfamily with roles in bacterial iron transport. Biometals 23: 1–17. [DOI] [PubMed] [Google Scholar]
- 13. Rajasekaran M, Mitchell S, Gibson T, Hussain R, Siligardi G, et al. (2010) Isolation and characterisation of EfeM, a periplasmic component of the putative EfeUOBM iron transporter of Pseudomonas syringae pv. syringae . Biochem Biophys Res Commun 398: 366–371. [DOI] [PubMed] [Google Scholar]
- 14. Maruyama Y, Ochiai A, Mikami B, Hashimoto W, Murata K (2011) Crystal structure of bacterial cell-surface alginate-binding protein with an M75 peptidase motif. Biochem Biophys Res Commun 405: 41141–41146. [DOI] [PubMed] [Google Scholar]
- 15. Xu Q, Rawlings N, Farr C, Chiu H, Grant J, et al. (2011) Structural and sequence analysis of imelysin-like proteins implicated in bacterial iron uptake. PLoS One 6: e21875 doi: 101371/journal.pone.0021875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Liu X, Du Q, Wang Z, Zhu D, Huang Y, et al. (2011) Crystal Structure and Biochemical Features of EfeB/YcdB from Escherichia coli O157: ASP235 plays divergent roles in different enzyme-catalysed processes. J Biol Chem 286: 14922–14931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Hommais F, Krin E, Laurent-Winter C, Soutourina O, Malpertuy A, et al. (2001) Large-scale monitoring of pleiotropic regulation of gene expression by the prokaryotic nucleoid-associated protein, H-NS. Mol Microbiol 40: 20–36. [DOI] [PubMed] [Google Scholar]
- 18. Peschel A, Götz F (1996) Analysis of the Staphylococcus epidermidis genes epiF, -E, and -G involved in epidermin immunity. J Bacteriol 178: 531–536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Mechold U, Ogryzko V, Ngo S, Danchin A (2006) Oligoribonuclease is a common downstream target of lithium-induced pAp accumulation in Escherichia coli and human cells. Nucleic Acids Res 34: 2364–2373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Osborn MJ, Munson R (1974) Separation of the inner (cytoplasmic) and outer membranes of Gram-negative bacteria. Methods Enzymol 31: 642–653. [DOI] [PubMed] [Google Scholar]
- 21. du Plessis D, Nouwen N, Driessen A (2011) The Sec translocase. Biochim Biophys Acta 1808: 851–865. [DOI] [PubMed] [Google Scholar]
- 22. Létoffé S, Delepelaire P, Wandersman C (2006) The housekeeping dipeptide permease is the Escherichia coli heme transporter and functions with two optional peptide binding proteins. Proc Natl Acad Sci U S A 103: 12891–12896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Kellermann O, Szmelcman S (1974) Active transport of maltose in Escherichia coli K12. Involvement of a “periplasmic” maltose binding protein. Eur J Biochem 47: 139–149. [DOI] [PubMed] [Google Scholar]
- 24. Barrett C, Ray N, Thomas D, Robinson C, Bolhuis A (2003.) Quantitative export of a reporter protein, GFP, by the twin-arginine translocation pathway in Escherichia coli. . Biochem Biophys Res Commun 30: 279–284. [DOI] [PubMed] [Google Scholar]
- 25.Hurd A, Garcia-Lara J, Rauter Y, Cartron M, Mohamed R, et al.. (2012) The Iron Regulated Surface Proteins IsdA, IsdB and IsdH Are Not Required For Haem Iron Utilisation In Staphylococcus aureus FEMS Microbiology Letters DOI: 10.1111/j.1574–6968.2012.02502.x. [DOI] [PubMed] [Google Scholar]
- 26. Torres V, Pishchany G, Humayun M, Schneewind O, Skaar E (2006) Staphylococcus aureus IsdB is a hemoglobin receptor required for heme iron utilization. J Bacteriol 188: 8421–8429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Xiao Q, Jiang X, Moore K, Shao Y, Pi H, et al. (2011) Sortase independent and dependent systems for acquisition of haem and haemoglobin in Listeria monocytogenes . Mol Microbiol 80: 1581–1597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Gaballa A, Helmann J (2011) Bacillus subtilis Fur represses one of two paralogous haem-degrading monooxygenases. Microbiology 157: 3221–3231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Herbert S, Ziebandt A, Ohlsen K, Schäfer T, Hecker M, et al. (2010) Repair of global regulators in Staphylococcus aureus 8325 and comparative analysis with other clinical isolates. Infect Immun 78: 2877–2889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Pogliano J, Lynch A, Belin D, Lin E, Beckwith J (1997) Regulation of Escherichia coli cell envelope proteins involved in protein folding and degradation by the Cpx two-component system. Genes Dev 11: 1169–1182. [DOI] [PubMed] [Google Scholar]
- 31. Perrotte-Piquemal M, Danchin A, Biville F (1999) Pyrophosphate increases the efficiency of enterobactin-dependent iron uptake in Escherichia coli. . Biochimie 81: 245–253. [DOI] [PubMed] [Google Scholar]
- 32. Létoffé S, Debarbieux L, Izadi N, Delepelaire N, Wandersman C (2003) Ligand delivery by heme carrier proteins: the binding of Serratia marcescens hemophore to its outer membrane receptor is mediated by two distinct peptide regions. Mol Microbiol 50: 77–88. [DOI] [PubMed] [Google Scholar]
- 33. Bartolome B, Jubete Y, Martınez E, de la Cruz F (1991) Construction and properties of a family of pACYC184-derived cloning vectors compatible with pBR322 and its derivates. Gene 102: 75–78. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
UV-visible spectra of FepB-6His purified by Nickel-nitrilotriacetic acid chromatography. A: FepB-6His purified from SET538 (pSU19-fepB-6His); B: FepB-6His purified from SET538 (pSU19-fepB-6His pBAD24-fepA). Insert: FepB-6His SDS-PAGE analysis. After staining in the Coomassie Blue, gels were digitalized using a JX-330 scanner (Sharp) and the purity of FepB-6His was determined using the Quantity One 1-D software (BioRad).
(TIF)