

Abstract
During the past century, few proteins have matched erythropoietin (Epo) in capturing the imagination of physiologists, molecular biologists, and, more recently, physicians and patients. Its appeal rests on its commanding role as the premier erythroid cytokine, the elegant mechanism underlying the regulation of its gene, and its remarkable impact as a therapeutic agent, arguably the most successful drug spawned by the revolution in recombinant DNA technology. This concise review will begin with a synopsis of the colorful history of this protein, culminating in its purification and molecular cloning. It then covers in more detail the contemporary understanding of Epo’s physiology as well as its structure and interaction with its receptor. A major part of this article focuses on the regulation of the Epo gene and the discovery of HIF, a transcription factor that plays a cardinal role in molecular adaptation to hypoxia. In the concluding section, a synopsis of Epo’s role in disorders of red blood cell production will be followed by an assessment of the remarkable impact of Epo therapy in the treatment of anemias, as well as concerns that provide a strong impetus for the development of even safer and more effective treatment.
Erythropoietin (Epo) is the primary mediator of red blood cell production under hypoxic conditions. Its gene is controlled by an elegant regulatory mechanism, and recombinant Epo is successfully used to treat anemias.
In 1890, Viault (1890) observed that 2 weeks after traveling in Peru from sea level (Lima) to the mountain area of Morococha (4200 m) his red blood cell count went from 5.0 million to 7.1 million/mm3. Values of five other sojourners in his party ranged from 7.1 to 8.0 million. These simple observations provided the first convincing demonstration of the robust burst of erythropoiesis in man soon after exposure to high altitude hypoxia. During the transition into the next millennium the mechanism underlying this phenomenon became a topic of heated debate. Friedrich Miescher (1893), well known for his discovery of DNA, proposed that a decrease in oxygen tension within the bone marrow provided a direct stimulus to erythroid cells. A half-century elapsed before this theory was disproven by carefully executed measurements of oxygen saturation in bone marrow specimens of patients with erythrocytosis, both primary (Berk et al. 1948) and secondary (Stohlman et al. 1954).
In 1906, Carnot and Deflandre (1906) proposed an alternate mechanism for hypoxic induction of erythropoiesis. They observed an increase in red blood cell counts following infusion of normal rabbits with serum from anemic animals and concluded that erythropoiesis is regulated by a humoral “factor” in the plasma. Attempts to reproduce this experiment over the ensuing decades yielded equivocal or negative results, thus casting doubt on this hypothesis. However, in the middle of the twentieth century, Krumdieck (1943) and Erslev (1953) modified the experimental design of Carnot and Deflandre by the addition of accurate measurements of reticulocytes and convincingly showed in rabbits the induction of new red cell production within 3–6 days following injection of anemic serum injected.
The notion that hypoxic stimulation of erythropoiesis involved an indirect humoral mechanism was strongly buttressed by experiments of Reissmann (1950) and Ruhenstroth-Bauer (1950). They used parabiotic pairs of rats whose circulations were connected at the capillary level by overlapping flaps of skin and elegantly showed that when one rat was exposed to low oxygen tension, whereas the other breathed room air and remained normoxic, both animals developed a surge of new red cell production and erythrocytosis.
Taken together, these studies led to the conception of a circulating erythroid-stimulating hormone, “erythropoietin” (Epo). Organ ablation studies in rats (Jacobson et al. 1957) and man (Nathan et al. 1964) firmly established that the kidney was the major site but not the sole site of Epo production. These findings led Eugene Goldwasser and his colleagues to undertake an intense and prolonged effort to isolate Epo. Initial attempts to obtain Epo from kidneys were unsuccessful owing to the release of proteolytic enzymes during tissue homogenization. In the search for a more tractable source of Epo, Goldwasser first turned to plasma of anemia sheep, then to urine from Argentinians with severe iron deficiency owing to hookworm infestation, and finally to urine from Japanese patients with aplastic anemia. This 15-year endeavor was greatly facilitated by the development of a sensitive and specific assay using incorporation of radio-labeled iron into newly produced red cells (Fried et al. 1956). By 1977, Goldwasser and his team were able to prepare 8 mg of highly purified human Epo (Miyake et al. 1977). Amino-terminal amino acid sequencing of this preparation enabled the synthesis of semidegenerate oligonucleotide probes that could then be used for the molecular cloning of the Epo gene (Jacobs et al. 1985; Lin et al. 1985). This advance opened up a new era in the exploration of the physiology and molecular biology of Epo and was exploited in the development of recombinant human Epo as a therapeutic agent for patients with various types of anemia.
For more detailed information on the history of Epo, see reviews by Grant and Root (1952), Erslev (1980, 1993), and Goldwasser (1996).
PHYSIOLOGY1
In humans and other mammals, erythropoiesis normally proceeds at a low basal rate, replacing senescent red blood cells with young reticulocytes. In humans, red cell production can be enhanced as much as eightfold the baseline rate in a variety of clinical settings including hemorrhage, hemolysis, and other types of stress that impair the oxygenation of arterial blood or the delivery of oxygen to the tissues. Epo is the primary, and probably the sole mediator of hypoxic induction of erythropoiesis. During fetal development, Epo is produced mainly in the liver. However, in keeping with the above-mentioned report of Jacobson et al. (1957), following birth, the kidney accounts for ∼80% of Epo production. Realizing that renal cortical blood flow closely matches oxygen consumption, Erslev et al. (1985) proposed that the proximal tubule is the ideal location for Epo production. In situ hybridization studies have indeed shown that Epo mRNA expression in the kidney is localized to a subset of peritubular fibroblasts in the cortex close to the boundary with the medulla (Koury et al. 1988, 1989; Lacombe et al. 1988; Bachmann et al. 1993; Maxwell et al. 1993a). In the liver, Epo is produced both in hepatocytes and in interstitial cells (Koury et al. 1991; Schuster et al. 1992). The latter have been shown to be Ito presinusoidal cells (Maxwell et al. 1994). In both Ito cells and the above-mentioned renal fibroblasts, in situ expression of Epo correlates with that of ecto-5′-nucleotidase, probably because this enzyme is also controlled by local oxygen tension (Schmid et al. 1994).
The notion that Epo production is markedly up-regulated by hypoxia and that it stimulates erythropoiesis in a dose-dependent manner led to the now well accepted paradigm of a negative feedback loop, similar to those underlying the regulation of blood glucose by insulin and of peripheral endocrine hormones by pituitary tropins. As shown in Figure 1A, hypoxia induces an increase in Epo hormone production in the kidney, which then circulates in the plasma and binds to receptors abundantly expressed on erythroid progenitor cells, thereby promoting the viability, proliferation, and terminal differentiation of erythroid precursors, and causing an increase in red blood cell mass. The oxygen-carrying capacity of the blood is thereby enhanced, increasing tissue oxygen tension, thus completing the feedback loop and suppressing further expression of Epo.
Figure 1.

Regulation of red cell production by Epo. (A) Decreased oxygen delivery to specialized cells in the kidney results in increased expression and secretion of Epo, which circulates in the plasma and stimulates marrow progenitors, thereby increasing red cell production. If the increase in red cell mass relieves the hypoxic signal, Epo expression is down-regulated. (B) Plasma Epo levels (milliunits/mL) in patients with different types of and degrees of anemia and in those with primary erythrocytosis and secondary erythrocytosis. HIF, hypoxia inducible factor; PCV, polycythemia vera.
Further insight into the physiology of Epo production has come from measurements of plasma levels in normal individuals as well as in those with different types of anemia and erythrocytosis (Erslev et al. 1987). These studies became more conclusive with the development of accurate immune assays. Figure 1B depicts plasma Epo levels in normal individuals as well as in those with different types of anemia of varying degree. Note that the plasma Epo levels are plotted on a logarithmic scale. In patients with very severe anemia, Epo may be increased up to 1000-fold. The striking inverse relationship between plasma Epo levels and hematocrit or hemoglobin is seen in many different types of anemia, including blood loss, impaired red cell production, and hemolysis.
BIOCHEMISTRY AND CELL BIOLOGY2
Structure
The molecular cloning of the Epo gene led straightaway to high-level production of recombinant human erythropoietin (rhEpo) in cell culture in sufficient purity and quantity for both definitive scientific pursuits and the development of therapy (discussed at the end of this article). Human Epo messenger RNA (mRNA) encodes a 193-residue polypeptide. Following cleavage of a canonical leader sequence in the endoplasmic reticulum and glycosylation in the Golgi, a 166-residue polypeptide is released (Jacobs et al. 1985; Lin et al. 1985). The primary structure of rhEpo was shown to be identical to that of the endogenous hormone except for the in vivo posttranslational cleavage of an arginine at the carboxyl terminus. Endogenous Epo and rhEpo have about 40% carbohydrate (w/w) of very similar structure, consisting of three N-linked polysaccharide groups and one O-linked group. Glycosylation of Epo slightly impedes its biological activity but is essential for ensuring prolonged circulation in the plasma (Goldwasser et al. 1974).
From its primary structure it was predicted that Epo, like other hematopoietic cytokines, folds into a globular three-dimensional structure consisting of a bundle of four amphipathic α helices connected by loops lacking secondary structure and stabilized by a critical disulphide bridge between the amino-terminal and carboxy-terminal helices (Bazan 1990; Boissel et al. 1993). This prediction was borne out by nuclear magnetic resonance spectroscopy of Epo in solution (Cheetham et al. 1998) as well as by X-ray crystallographic analysis of Epo in a complex with two extracellular domains of the Epo receptor (Syed et al. 1998).
Interaction with EpoR
Epo circulates in plasma with a plasma half-life of ∼7–8 h and binds to high-affinity (∼100 pM) receptors present in relatively small numbers (∼1000/cell) on the surface of erythroid progenitor cells (CFUe) in the bone marrow. The receptor-binding domains on the surface of Epo that were predicted from the properties of site-directed mutants (Wen et al. 1994; Elliott et al. 1996, 1997) have been confirmed and extended by structural analysis (Syed et al. 1998; Remy et al. 1999). EpoR is present on the surface of erythroid progenitors as a homodimer, even in the absence of ligand (Livnah et al. 1999). As shown in Figure 2, on binding to Epo, the receptor undergoes a conformational change that brings its intracellular domains into close apposition (Livnah et al. 1999; Remy et al. 1999), enabling cross phosphorylation via the binding of Jak2 kinase and the initiation of the signal transduction cascade.
Figure 2.

Epo-dependent signaling. When Epo binds to its dimeric receptor (EpoR) on erythroid progenitor cells, the two receptor monomers are pulled together allowing phosphorylation of JAK2 kinase, which initiates the signal transduction cascade.
The growth of CFUe in vitro was shown to be Epo dependent (McLeod et al. 1974). Ligand binding results in salvaging CFUe from apoptosis (Koury and Bondurant 1990) thereby enabling proliferation of erythroid cells as well as induction of terminal differentiation. The finding that both Epo mRNA and Epo protein are expressed in erythroid progenitors (Hermine et al. 1991; Stopka et al. 1998) has raised the intriguing possibility that tonic low-level erythropoiesis may be supported by autocrine stimulation, whereas circulating (hormonal) Epo provides a more robust stimulus to erythropoiesis during hypoxic stress.
In man, Epo’s hematopoietic role appears to be restricted to the erythron, whereas in rodents, Epo stimulates megakaryocyte proliferation and maturation as well (Clark and Dessypris 1986; McDonald et al. 1987; Berridge et al. 1988).
NONHEMATOPOIETIC EFFECTS OF Epo3
As stressed in the sections above, by far the most abundant sites of Epo production are the kidney and liver, whereas expression of EpoR is vastly more robust in marrow erythroid progenitors than in other cells. However, it has become increasingly evident that Epo has significant biological effects apart from regulation of red cell production. Attention has focused primarily on the central nervous system along with the heart, kidney, liver, and vasculature.
Within the brain, Epo and Epo-receptor mRNA are expressed widely (Tan et al. 1991; Masuda et al. 1994; Digicaylioglu et al. 1995; Sakanaka et al. 1998) and throughout development (Dame et al. 2000) in neurons, astrocytes, and endothelial cells (Yamaji et al. 1996). In contrast, demonstration of Epo and EpoR protein expression in situ has been considerably more challenging and less convincing. Oxygen-regulated expression of Epo has been observed in astrocytes both in vitro (Masuda et al. 1994; Marti et al. 1996) and in vivo (Tan et al. 1991; Marti et al. 1996), suggesting the possibility of a paracrine function for Epo in neural tissue.
The most compelling evidence of the importance of Epo signaling in the central nervous system comes from two independent sources. Targeted knockout of EpoR expression in the mouse brain has shown reduced neural cell proliferation and impaired poststroke neurogenesis (Tsai et al. 2006). Although global deletion of EpoR is lethal to mouse embryos, rescue by expression of erythroid-specific EpoR results in survival to adulthood (Suzuki et al. 2002) but impaired neural cell proliferation and viability (Chen et al. 2007). In addition, administration of Epo protects against experimental brain injury in vivo (Sakanaka et al. 1998; Brines et al. 2000). Moreover, structural derivatives of Epo that lack erythroid stimulating activity have been shown to confer neuroprotection in animal models of stroke, spinal chord compression, diabetic neuropathy, and autoimmume encephalomyelitis (Leist et al. 2004). In a preliminary trial in patients with acute stroke, recombinant human erythropoietin (rhEpo) therapy was well tolerated and readily penetrated the blood–brain barrier (Ehrenreich et al. 2002). The authors' cbservations of reduction in infarct size and clinical improvement have not been confirmed in a more extensive clinical trial (Ehrenreich et al. 2009).4
High doses of rhEpo (Parsa et al. 2004; Nishiya et al. 2006) and the above-mentioned Epo derivatives (Fiordaliso et al. 2005; Brines et al. 2008) confer protection in animal models of cardiac ischemia/reperfusion and myocardial infarction. Moreover rhEpo is also effective in protecting the kidney and liver from ischemia/reperfusion injury (Yang et al. 2003; Patel et al. 2004; Sharples et al. 2004; Sepodes et al. 2006; Schmeding et al. 2009) as well as other experimental threats to kidney (Vaziri et al. 1994; Bahlmann et al. 2004) or liver (Le Minh et al. 2007; Schmeding et al. 2008, 2009) integrity.
The salutary effects of rhEpo and derivatives thereof on experimental models of ischemia and other types of injury may be due in part to their effect on the vascular endothelium. In vitro rhEpo stimulates the proliferation, mobilization, and differentiation of endothelial progenitor and precursor cells (Bahlmann et al. 2003; Heeschen et al. 2003) and also enhances endothelial cell viability (Chen et al. 2008) and survival by blocking apoptosis (Bahlmann et al. 2004). Thus, exogenous Epo can significantly enhance neovascularization induced by inflammation and ischemia (Heeschen et al. 2003).
Perhaps the most puzzling aspect of nonerythroid effects of Epo is the nature of the receptor. In erythroid cells, EpoR mRNA is expressed at sufficiently high levels for abundant display of EpoR protein on cell surface and dimerization that is required for high-affinity Epo binding. In contrast, the mRNA expression in nonerythroid cells is so low that convincing demonstration of surface EpoR protein by immunological means has been problematic at best. It is very likely that such low-level expression would not be sufficient for the mass action needed for EpoR dimerization to occur. Accordingly, the low-affinity binding site for Epo may be a heterodimer composed of EpoR and another partner, perhaps CD131, the β common cytokine receptor (Brines et al. 2008; Brines 2010). This may explain the requirement of high levels of exogenous Epo for biological effects in nonerythroid cells and possibly how an Epo derivative could have activity only in nonerythroid cells.
REGULATION OF THE Epo GENE5
Initial investigation of the molecular basis of Epo gene regulation was greatly facilitated by the discovery that the human hepatoma cell lines Hep3B and HepG2 produce significant amounts of Epo constitutively, with marked induction in response to hypoxia (Goldberg et al. 1987). The magnitude and time course of the induction of Epo mRNA paralleled Epo protein production. More recently, a human renal cell line has also been shown to display oxygen-dependent regulation of Epo production (Frede et al. 2011).
Tissue-specific expression of the Epo gene and its induction by hypoxia are dependent on far upstream cis elements and an enhancer element downstream from the polyadenylation signal. Transgenic mice experiments indicate the presence of element(s) between 0.4 and 6 kb 5′ of the promoter, which suppress promiscuous expression, and element(s) 9.5 and 14 kb upstream, which are necessary for kidney-specific expression (Semenza et al. 1989, 1990, 1991a; Madan et al. 1995).
The Epo gene has a weak promoter, owing in part to suppression of transcription by a GATA motif (Imagawa et al. 1994, 1997). Mutation of this response element in a 180-kb Epo transgene resulted in constitutive in vivo expression in renal peritubular cells (Obara et al. 2008). Enhanced binding of GATA-2 to this site may contribute to the inhibition of Epo production by inflammatory cytokines (La Ferla et al. 2002). GATA-4 is expressed exclusively in hepatocytes and binds strongly to this GATA motif, enhancing Epo expression (Dame et al. 2004). The marked reduction in expression of hepatic GATA-4 following birth is likely an important contributor to the switch in Epo production from liver to kidney (Dame et al. 2004).
A crucial 3′ enhancer (Beck et al. 1991; Pugh et al. 1991; Semenza et al. 1991b; Semenza and Wang 1992) binds to two transcription factors, hypoxia-inducible factor 1 (HIF) and the nuclear receptor HNF-4. As shown in Figure 3, these two DNA-binding proteins interact with the transcriptional coactivator p300/CBP, triggering transcriptional activation.
Figure 3.
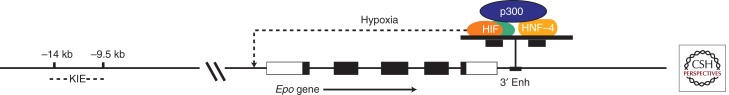
Diagram of the Epo gene. The five exons of the Epo gene are shown as rectangles with the coding regions in black. Far upstream of the Epo promoter is a kidney-inducible element (KIE) that is required for high-level up-regulation of Epo mRNA in the kidney. Just downstream from exon 5 is a critical enhancer that binds to HNF-4 and also to HIF when the Epo-producing cell is hypoxic. These two transcription factors bind to the transcriptional activator p300. This enhanceosome is a powerful inducer of Epo transcription.
HIF
Hypoxic induction of Epo depends in large part on the transcription factor HIF. HIF is activated in virtually all cells (Maxwell et al. 1993b; Wang and Semenza 1993b) by exposure to hypoxia, as well as cobalt or iron chelators. The activation of HIF enables it to bind to a consensus sequence (5′-TACGTGCT-3′) first identified in the Epo 3′ enhancer (Fig. 3) (Semenza et al. 1991b). Subsequently, this hypoxia response element has been identified in more than a hundred genes and shown to be required for hypoxic induction of transcription. In addition to Epo, an impressive number of other physiologically relevant genes are regulated by HIF (Wenger et al. 2005) including those involved in angiogenesis, intermediary metabolism, and, of particular interest to readers of this collection, iron homeostasis (Peyssonnaux et al. 2007).
Wang and Semenza (1995) purified HIF by its affinity for the Epo 3′ enhancer and showed that it is a heterodimer composed of a 120 kD α subunit and a 91–94 kD β subunit. Molecular cloning revealed that both subunits are basic helix-loop-helix proteins in the PAS family of transcription factors (Wang et al. 1995a). HIF-α, or HIF-1α, is a novel protein, whereas HIF-1β is the previously cloned and characterized aryl hydrocarbon receptor nuclear translocator (ARNT) (Hoffman et al. 1991).
The HIF-α subunit is widespread among animals, including not only mammals and fish but also invertebrates such as flies and worms. In man and other mammals, three homologous HIF-α genes have been identified: HIF-1α, HIF-2α (Tian et al. 1997), and HIF-3α (Gu et al. 1998). HIF-1α is the most abundant and present in most organs and tissues. In contrast, HIF-2α has more restricted expression and was originally thought to be limited to the vascular endothelium (Tian et al. 1997). Knockout of HIF-1α in mice is embryonic lethal (Iyer et al. 1998; Ryan et al. 1998), whereas HIF-2α null mice survive gestation but have impaired survival owing to phenotypic abnormalities that are strain-dependent. Most genes that are induced by hypoxia in an HIF-dependent manner require HIF-1α (Warnecke et al. 2004). In contrast, Epo production and its up-regulation by hypoxia require HIF-2α (Gruber et al. 2007) in the kidney (Scortegagna et al. 2005; Kapitsinou et al. 2010; Paliege et al. 2010) and an Epo-producing renal cell line (Frede et al. 2011), as well as in the fetal and adult liver (Rankin et al. 2007).
Oxygen Sensing and HIF Activation
To understand the mechanism by which HIF-1 is activated, it is necessary to determine how hypoxia impacts on the expression of its two subunits. The steady-state levels of HIF-α mRNA and ARNT mRNA are not significantly affected by oxygen tension (Gradin et al. 1996; Huang et al. 1996; Wood et al. 1996). At the protein level, the ARNT subunit remains abundant, irrespective of the oxygenation of the cell. In contrast, the HIF-α subunit cannot be detected in oxygenated cells (Wang et al. 1995b; Huang et al. 1996). HIF-α protein can only be detected in deoxygenated cells or in those exposed to iron chelators or certain transition-metal ions, most notably cobalt, all of which induce HIF-dependent gene transcription (Semenza and Wang 1992; Wang and Semenza 1993a). These observations suggest that the activation of HIF-1 depends on an increase in the amount of HIF-α protein in deoxygenated cells.
In cells exposed to 21% O2, HIF-α is remarkably unstable (Huang et al. 1996). It forms a complex with von Hippel–Lindau protein (vHL) (Maxwell et al. 1999), enabling it to undergo ubiquitination and subsequent degradation in proteasomes (Salceda and Caro 1997; Huang et al. 1998). Low oxygen tension abrogates this process, thereby allowing the HIF-α subunit to accumulate so that it can enter the nucleus and form a stable heterodimer that can participate in transcriptional regulation. Oxygen-dependent degradation of HIF-α depends on a large (∼200-residue) interior segment (Huang et al. 1998). When this domain is deleted, the protein is stable and capable of trans-activation, even in oxygenated cells. When this domain is fused to a foreign protein, it confers rapid oxygen-dependent degradation.
The oxygen-dependent degradation domain of HIF-1α contains two highly conserved LXXLAP sequences, which are recognition sites for an oxygen- and iron-dependent prolyl hydroxylase that converts the proline into a hydroxyproline (Ivan et al. 2001; Jaakkola et al. 2001; Masson et al. 2001; Yu et al. 2001). This posttranslational modification is necessary and sufficient for the binding of vHL to HIF-α. As shown in Figure 4, this interaction enables the ubiquitin E3 ligase to dock at this complex, thereby triggering the covalent addition of a number of small ubiquitin molecules (Cockman et al. 2000; Ohh et al. 2000). The polyubiquitinated HIF-α is then recognized by the proteasome where it is rapidly degraded. Ratcliffe and his colleagues (Epstein et al. 2001) showed that the hydroxylation of these two specific proline residues is mediated by a dioxygenase that is homologous to classic collagen prolyl hydroxylase. Shortly thereafter, a human homolog of HIF prolyl hydroxylase was biochemically purified and characterized (Ivan et al. 2002). These enzymes have an active center where an iron atom and a molecule of α-ketoglutarate interact with a conserved core of two histidines, an aspartate and an arginine residue. The two LXXLAP motifs within HIF-α each bind at this site, along with oxygen, catalyzing hydroxylation of the proline residue. This oxidative modification is inhibited by iron chelators as well as by cobaltous ion, which presumably displaces iron from the active site. The effect of the iron chelation and cobalt on the activity of HIF prolyl hydroxylases provides a satisfying explanation for how these agents mimic hypoxia in activating HIF. These enzymes are the proximate oxygen sensor for regulating HIF. Among the three HIF-α prolyl hyderoxylases, PHD2 plays the predominant role at most sites (Berra et al. 2003) including renal production of Epo (Minamishima et al. 2008; Takeda et al. 2008).
Figure 4.

Pathway by which the hypoxia-inducible transcription factor is up-regulated by low intracellular oxygen tension. In normally oxygenated cells, the α subunit (HIF-α) undergoes hydroxylation of two proline residues, one of which is shown here. In the presence of iron (Fe) and α-ketoglutarate, this oxygen-dependent posttranslational modification is catalyzed by an HIF-α-specific prolyl hydroxylase (PH). The von Hippel–Lindau protein (pVHL) binds to hydroxylated HIF-α. Subsequent docking of a ubiquitin ligase (UL) enables HIF-α to be polyubiquitinated.
Hemoglobin is encapsulated in circulating erythrocytes not only in man and other mammals but also in all other vertebrates. Oxygen homeostasis in these complex organisms is well served by HIF-dependent regulation of a cytokine that is required for erythropoiesis. In contrast, in many invertebrate phyla, oxygen transport depends on the presence of free soluble polymeric hemoglobins in circulating hemolymph, and in some, hemoglobin production is enhanced during hypoxic stress. One of the most dramatic observations in marine biology is the change in the fresh-water crustacean Daphnia magna from pale gray to bright red soon after transfer from normoxic to hypoxic water (Fox 1947, 1949). Two of their four tandem globin genes have been shown to have functional HIF response elements within their promoters (Gorr et al. 2004). Thus, in keeping with the relative simplicity of this organism, the regulation of oxygen-carrying capacity of the blood is direct, rather than mediated indirectly via a cytokine.
HNF-4
Downstream from the HIF-1 binding site, the Epo 3′ enhancer contains two tandem consensus steroid hormone response elements separated by 2 bp (Fig. 1). Mutations at these sites either abolish or markedly inhibit hypoxic induction of reporter genes (Blanchard et al. 1992; Semenza and Wang 1992; Pugh et al. 1994; Hu et al. 1997). These elements in the Epo enhancer interact with an orphan nuclear receptor, i.e., a DNA-binding protein that shares structural homology with hormone-binding nuclear receptors but lacks a known ligand (Blanchard et al. 1992). The orphan receptor HNF-4 plays a critical role in Epo gene regulation (Galson et al. 1995; Makita et al. 2001). The expression of HNF-4 is limited to renal cortex and liver, sites of Epo production, and also intestine. Thus the binding of HNF-4 to the Epo enhancer is likely to contribute to both the high-level induction of the Epo gene as well as its tissue specificity.
p300
As shown in Figure 1, the carboxy-terminal portion of HIF-1α binds specifically to p300 (Arany et al. 1996), a general transcriptional activator that participates in a number of biological functions such as induction of tissue-specific expression, regulation of cell cycle, and stimulation of differentiation pathways. This very large protein, which is closely homologous to CREB-binding protein (CBP), does not bind to DNA but does interact with a number of other proteins including HNF-4 and other nuclear receptors. Thus, as depicted in Figure 1, the HIF-1 heterodimer, activated by hypoxia, participates in a macromolecular assembly with p300 (or a related family member) and with HNF-4 to transduce a signal to the Epo promoter, enabling activation of transcription. It is very likely that such a combinatorial process applies not only to Epo but also to other genes that are induced by hypoxia (Ebert and Bunn 1998). Most, if not all, cells contain a protein factor called FIH (factor-inhibiting HIF) (Mahon et al. 2001) that hydroxylates an asparagine at the carboxyl terminus of HIF-α in an oxygen- and iron-dependent manner, thereby blocking binding to p300 and inhibiting transcription (Lando et al. 2002a,b).
IMPACT OF Epo PRODUCTION ON CLINICAL DISORDERS
In a variety of clinical settings, anemia can be caused by underproduction of Epo, whereas erythrocytosis can result from overproduction.
Underproduction of Epo
Patients with renal failure generally develop severe anemia, owing in part to suppression of erythropoiesis from a buildup of metabolic wastes and in part to a moderate reduction in red cell life span. However, the most important contributor to the anemia of uremia is insufficient Epo production (Fig. 1B) (Caro et al. 1979; Cotes 1989). The degree of anemia correlates roughly with the extent of renal functional impairment. In renal failure owing to a wide variety of etiologies, insufficient Epo production results either from direct damage to Epo-producing cells in the kidney or from the suppression of Epo production by inflammatory cytokines.
Patients with inflammatory disorders have a less marked impairment of Epo production. Examples include rheumatoid arthritis (Means 1994), cancer (Miller et al. 1990; Spivak 1994), and AIDS (Fischl et al. 1990). It is likely that inflammatory cytokines suppress Epo gene expression in these disorders.
Exposure to certain metals may result in disordered structure and function of the renal proximal tubule, resulting in suppression of Epo production, out of proportion to impairment of global renal function. Such a pathogenetic process is likely the basis for the marked anemia often encountered in cancer patients who have been treated with cis-platinum or individuals suffering from chronic cadmium intoxication (Horiguchi et al. 2000). Epo production is also inhibited in patients with increased plasma viscosity owing to monoclonal dysproteinemias (Singh et al. 1993).
Overproduction of Epo
Measurement of plasma Epo is useful in distinguishing primary from secondary erythrocytosis (Erslev and Caro 1984; Messinezy et al. 2002). Individuals with autonomous red cell overproduction due to polycythemia vera (or rarely to mutations at the carboxyl terminus of EpoR) have low plasma Epo levels, whereas individuals with various forms of chronic arterial hypoxemia have erythrocytosis owing to increased plasma Epo levels. The highest documented hematocrit levels have been observed in patients with right to left cardiac shunts. The erythropoietic response in patients with hypoxemia owing to chronic obstructive pulmonary disease is variable, depending in part on whether there is coexisting infection, which, as explained above, can suppress erythropoietin expression.
Increased oxygen affinity of red cells causes impaired delivery of oxygen to tissues and therefore hypoxia at the cellular level, which triggers increased Epo expression and therefore secondary erythrocytosis (Prchal and Prchal 1999). Such a “shift to the left” in the oxyhemoglobin dissociation curve can be caused by mutations in α- or β-globin subunits or in two red cell enzymes. Deficiency in cytochrome b5 reductase causes congenital mehemoglobinemia and mild erythrocytosis. A more pronounced increase in red cell mass has been reported in very rare families with deficiency of bisphosphoglycerate mutase that causes a deficit in red cell 2,3-BPG, a critical modulator of intracellular hemoglobin function (Rosa et al. 1978). These individuals with left-shifted O2-binding curves are generally asymptomatic because impaired oxygen delivery is balanced by increased oxygen-carrying capacity.
Occasional individuals have chronic elevation of plasma Epo that is inappropriate and autonomous, i.e., not due to arterial hypoxemia or cellular hypoxia. These individuals usually have either sporadic or germline mutation in genes responsible for oxygen sensing and HIF regulation.
Erythrocytosis due to inappropriate overproduction of Epo can be encountered in various neoplasms, particularly those arising in kidney, liver, and cerebellum, organs that are physiologic sites of erythropoietin expression. In particular, secondary erythrocytosis occurs in patients with renal carcinomas, Wilms tumor, hepatomas, and cerebellar hemangioblastomas. In some cases, the tumor cells secrete Epo (Jelkmann 1992). In other cases, the surrounding normal kidney or liver tissue secretes Epo, presumably because of local ischemia.
Highly vascular renal and central nervous system tumors can arise in families and sporadically owing to mutations in the von Hippel–Lindau (VHL) gene. As explained earlier in this article, this protein is required for the oxygen-dependent degradation of HIF-α. Inactivating mutations lead to constitutive activation of HIF-α and therefore overexpression of HIF responsive genes (Gnarra et al. 1996; Iliopoulos et al. 1996), particularly vascular endothelial growth factor (VEGF), which causes the tumors to have enhanced vascularity. In a subset of affected individuals, overexpression of Epo leads to erythrocytosis.
A number of families with congenital erythrocytosis have been encountered in Chuvashia, a circumscribed region in the Russian Federation in which there may have been inbreeding and a high likelihood of a founder effect (Sergeyeva et al. 1997; Gordeuk and Prchal 2006). The erythrocytosis (mean hemoglobin levels of 23 g/dL and hematocrit of 67%) follows an autosomal recessive pattern of inheritance. Affected individuals have elevated erythropoietin levels, and increased incidence of thrombotic and hemorrhagic complications. These individuals are homozygous for a missense mutation at the carboxyl terminus of the VHL gene (R200W) leading to HIF activation (Ang et al. 2002). The clinical phenotype is similar in a R200W “knock-in” mouse model (Hickey et al. 2007). Isolated families with congenital erythrocytosis have been encountered elsewhere that harbor the R200W mutation (Pastore et al. 2003; Cario et al. 2005; Perrotta et al. 2006). Affected individuals are usually homozygotes but some are compound heterozygotes, having R200W along with another VHL mutation (Pastore et al. 2003; Cario et al. 2005).
Families as well as isolated individuals have been encountered in which erythrocytosis owing to elevated plasma Epo levels is explained by heterozygosity for an activating mutation in HIF-2α (Gale et al. 2008; Martini et al. 2008; Percy et al. 2008a,b, 2012; Furlow et al. 2009; van Wijk et al. 2010). These mutations generally impair the ability of HIF-2α to bind to VHL or PHD2. In other families, affected individuals are heterozygous for loss-of-function mutations in PHD2 (Percy et al. 2006, 2007; Ladroue et al. 2008). Genome-wide association studies comparing lowland Tibetans to highlanders with erythrocytosis have shown highly significant polymorphisms in the vicinity of HIF-2α (Beall et al. 2010; Yi et al. 2010) and PHD2 (Simonson 2010). Both these selected “experiments of nature” and the broader population studies, attest to the primacy of HIF-2α and PHD2 in regulating the Epo gene.
THERAPY WITH rhEpo AND ERYTHROID- STIMULATING AGENTS6
By far the most common use of rhEpo has been in patients with chronic renal failure. Over one million patients worldwide have responded effectively to either rhEpo or a derivative, darbepoietin alfa (Aranesp), which has an enhanced plasma half-life (∼22 h) owing to the introduction of two additional N-linked glycosylation sites.
The importance of erythropoietin in the pathogenesis of the anemia of renal failure has been affirmed by the dramatic efficacy of rhEpo therapy. Figure 5 shows one of the first cases treated with rhEpo. Within a few days after initiation of rhEpo therapy, the hematocrit approached normal, necessitating a reduction in dose. The marked increase in red cell mass following treatment was accompanied by enhanced utilization of iron stores, as reflected in a decline in serum iron and serum ferritin. Other patients who have normal or low iron stores before rhEpo therapy need concomitant administration of iron to achieve an optimal erythropoietic response.
Figure 5.

Response of an anephric patient to recombinant human erythropoietin (rhEpo) therapy. Note that, before therapy, the patient was severely anemic and transfusion dependent. Treatment with rhEpo resulted in a reticulocytosis followed by a progressive increase in hemoglobin. The dose of rhEpo had to be lowered to prevent the hemoglobin from rising too high. Before rhEpo therapy, the patient was severely iron overloaded. The marked increase in red cell mass following therapy was accompanied by a significant reduction in iron stores. RBC, red blood cells; Fe, iron; TIBC, total iron binding capacity; Sat., saturation (Data from Eschbach et al. 1987.)
The other common use of rhEpo has been in treating anemia in cancer patients with chemotherapy-associated anemia. In addition, rhEpo is effective in AIDs patients who have developed severe anemia as a result of antiviral therapy. A minority of patients with myelodysplastic anemia also responds to rhEpo. In some clinics, rhEpo is used in the management of the anemia of prematurity as well in surgical patients, in both the preoperative and postoperative periods.
The impressive efficacy and safety of rhEpo therapy notwithstanding, a number of problems warrant careful consideration. The most evident are the high cost of treatment and the need for parenteral administration. Of equal concern are outcome studies on large numbers of renal dialysis patients showing that doses of rhEpo and darbepoietin alfa that raise the hemoglobin level >12 g/dl are associated with thrombosis of arteriovenous access, impaired control of hypertension, and a small but significant increase in mortality (Besarab et al. 1998; Singh et al. 2006; Phrommintikul et al. 2007). Cancer patients are inherently at higher risk to develop thrombosis, but this risk is significantly enhanced in those receiving rhEpo therapy (Bohlius et al. 2006; Bennett et al. 2008).
It is likely that the prothrombotic effects of rhEpo dose are due to the high doses given rather than to the increase in hemoglobin level. As mentioned earlier in this article, nonerythropoietic effects are widespread and complex. Early reports suggest that rhEpo may also promote enhanced tumor growth in vivo (Henke et al. 2003; Leyland-Jones 2003) and in vitro (Mohyeldin et al. 2005). However, EpoR expression is low in the great majority of tumors and though expressed at the mRNA level, it cannot be detected on the cell surface (Sinclair et al. 2008). Moreover, more recent meta-analyses mentioned above do not support the notion that rhEpo therapy promotes tumor progression in vivo (Bohlius et al. 2006; Bennett et al. 2008).
RECENT DEVELOPMENTS
In view of the above-mentioned concerns regarding long-term rhEpo therapy, there has been increasing interest in developing erythroid-stimulating agents that offer advantages over rhEpo and darbepoietin alfa. One approach entailed the production of a large phage library expressing short (∼15-residue) random peptides that are then screened for sequences that bind with relatively high affinity and specificity to the Epo receptor. This strategy led to the identification of peptides that lack any homology with Epo but are active both in vitro and in vivo in stimulating erythropoiesis through the same signaling pathway that is triggered by Epo (Livnah et al. 1996; Wrighton et al. 1996). However, in contrast to recent progress with thrombopoietin mimetic agents, little progress has been made to date in developing these Epo mimetic peptides into safe and effective drugs.
Another approach involves structural modification of Epo in the interest of prolonging its longevity in the circulation. Continuous erythropoietin receptor activator (CERA) is a derivative in which a 30 kDa methoxy-polyethylene glycol polymer has been covalently linked to Epo. It has a half-life in the circulation of ∼135 h, about sixfold greater than that of darbepoietin alfa (Macdougall 2005; Macdougall et al. 2006; Provenzano et al. 2007). This attribute greatly reduces the frequency of parenteral administration. CERA has been through extensive clinical trials and has passed muster in terms of both safety and efficacy. As a result, it received approval from the European Commission and the American Food and Drug Administration in 2008. However, it has not yet gained wide use in clinical practice.
The most novel strategy for pharmacologic stimulation of erythropoiesis involves the activation of HIF by inhibition of HIF-α prolylhydroxylases. As shown in Figure 4, small molecule analogs of α-ketoglutarate could prevent HIF-α hydroxylation, even in well-oxygenated cells and thereby induce expression not only of endogenous Epo but also of other HIF-dependent genes that are required for robust erythropoiesis. Several companies have developed inhibitors that induce Epo and erythropoiesis in mice (Safran et al. 2006; Yan et al. 2010) and in humans (Yan et al. 2010). Because these agents suppress expression of hepcidin (Volke et al. 2009), they may be particularly effective in the treatment of anemia of chronic inflammation. HIF prolyl hydroxylase inhibitors offer for the first time the possibility of inexpensive oral therapy for stimulation of erythropoiesis. Of course, a major concern is specificity because such a large number of genes are regulated by HIF. The lead compounds that are currently undergoing clinical trials appear to inhibit HIF-2α specifically. If so, they may also enhance duodenal iron absorption because the divalent metal transporter 1 (DMT1), like Epo, is up-regulated by HIF-2α.
For more information on all aspects of erythropoietin, see the recent detailed and comprehensive review by Wenger and Kurtz (2011).
For more detailed information on Epo structure, interaction with receptor, and signal transduction, see review by Jelkmann and Wagner (2004).
For more information on this topic, see review by Noguchi et al. (2008).
For more information on Epo and the brain, see reviews by Jelkmann (2005) and Noguchi et al. (2007).
For more information on this topic, see review by Fandrey (2004).
For more information on this subject, see reviews by Bunn (2007), Elliott (2008), and Jelkmann (2008).
Editors: David Weatherall, Alan N. Schechter, and David G. Nathan
Additional Perspectives on Hemoglobin and Its Diseases available at www.perspectivesinmedicine.org
REFERENCES
- Ang SO, Chen H, Hirota K, Gordeuk VR, Jelinek J, Guan Y, Liu E, Sergueeva AI, Miasnikova GY, Mole D, et al. 2002. Disruption of oxygen homeostasis underlies congenital Chuvash polycythemia. Nat Genet 32: 614–621 [DOI] [PubMed] [Google Scholar]
- Arany Z, Huang LE, Eckner R, Bhattacharya S, Jiang C, Goldberg MA, Bunn HF, Livingston DM 1996. Participation by the p300/CBP family of proteins in the cellular response to hypoxia. Proc Natl Acad Sci 93: 12969–12973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bachmann S, Le Hir M, Eckardt K 1993. Co-localization of erythropoietin mRNA and ecto-5′-nucleotidase immunoreactivity in peritubular cells of rat renal cortex indicates that fibroblasts produce erythropoietin. J Histochem Cytochem 41: 335–341 [DOI] [PubMed] [Google Scholar]
- Bahlmann FH, DeGroot K, Duckert T, Niemczyk E, Bahlmann E, Boehm SM, Haller H, Fliser D 2003. Endothelial progenitor cell proliferation and differentiation is regulated by erythropoietin. Kidney Int 64: 1648–1652 [DOI] [PubMed] [Google Scholar]
- Bahlmann FH, Song R, Boehm SM, Mengel M, von Wasielewski R, Lindschau C, Kirsch T, de Groot K, Laudeley R, Niemczyk E, et al. 2004. Low-dose therapy with the long-acting erythropoietin analogue darbepoetin α persistently activates endothelial Akt and attenuates progressive organ failure. Circulation 110: 1006–1012 [DOI] [PubMed] [Google Scholar]
- Bazan JF 1990. Haemopoietic receptors and helical cytokines. Immunol Today 11: 350–354 [DOI] [PubMed] [Google Scholar]
- Beall CM, Cavalleri GL, Deng L, Elston RC, Gao Y, Knight J, Li C, Li JC, Liang Y, McCormack M, et al. 2010. Natural selection on EPAS1 (HIF2α) associated with low hemoglobin concentration in Tibetan highlanders. Proc Natl Acad Sci 107: 11459–11464 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck I, Ramirez S, Weinmann R, Caro J 1991. Enhancer element at the 3′-flanking region controls transcriptional response to hypoxia in the human erythropoietin gene. J Biol Chem 266: 15563–15566 [PubMed] [Google Scholar]
- Bennett CL, Silver SM, Djulbegovic B, Samaras AT, Blau CA, Gleason KJ, Barnato SE, Elverman KM, Courtney DM, McKoy JM, et al. 2008. Venous thromboembolism and mortality associated with recombinant erythropoietin and darbepoetin administration for the treatment of cancer-associated anemia. JAMA 299: 914–924 [DOI] [PubMed] [Google Scholar]
- Berk L, Burchenal J, Wood TM, Castle W 1948. Oxygen saturation of sternal marrow blood with special reference to pathogenesis of polycythemia vera. Proc Soc Exp Biol Med 69: 316–320 [DOI] [PubMed] [Google Scholar]
- Berra E, Benizri E, Ginouves A, Volmat V, Roux D, Pouyssegur J 2003. HIF prolyl-hydroxylase 2 is the key oxygen sensor setting low steady-state levels of HIF-1α in normoxia. EMBO J 22: 4082–4090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berridge MV, Fraser JK, Carter JM, Lin FK 1988. Effects of recombinant human erythropoietin on megakaryocytes and on platelet production in the rat. Blood 72: 970–977 [PubMed] [Google Scholar]
- Besarab A, Bolton WK, Browne JK, Egrie JC, Nissenson AR, Okamoto DM, Schwab SJ, Goodkin DA 1998. The effects of normal as compared with low hematocrit values in patients with cardiac disease who are receiving hemodialysis and epoetin. N Engl J Med 339: 584–590 [DOI] [PubMed] [Google Scholar]
- Blanchard KL, Acquaviva AM, Galson DL, Bunn HF 1992. Hypoxic induction of the human erythropoietin gene: Cooperation between the promoter and enhancer, each of which contains steroid receptor response elements. Mol Cell Biol 12: 5373–5385 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bohlius J, Wilson J, Seidenfeld J, Piper M, Schwarzer G, Sandercock J, Trelle S, Weingart O, Bayliss S, Djulbegovic B, et al. 2006. Recombinant human erythropoietins and cancer patients: Updated meta-analysis of 57 studies including 9353 patients. J Natl Cancer Inst 98: 708–714 [DOI] [PubMed] [Google Scholar]
- Boissel J-P, Lee W-R, Presnell SR, Cohen FE, Bunn HF 1993. Erythropoietin structure-function relationships: Mutant proteins that test a model of tertiary structure. J Biol Chem 268: 15983–15993 [PubMed] [Google Scholar]
- Brines M 2010. The therapeutic potential of erythropoiesis-stimulating agents for tissue protection: A tale of two receptors. Blood Purificat 29: 86–92 [DOI] [PubMed] [Google Scholar]
- Brines ML, Ghezzi P, Keenan S, Agnello D, de Lanerolle NC, Cerami C, Itri LM, Cerami A 2000. Erythropoietin crosses the blood-brain barrier to protect against experimental brain injury. Proc Natl Acad Sci 97: 10526–10531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brines M, Patel NS, Villa P, Brines C, Mennini T, De Paola M, Erbayraktar Z, Erbayraktar S, Sepodes B, Thiemermann C, et al. 2008. Nonerythropoietic, tissue-protective peptides derived from the tertiary structure of erythropoietin. Proc Natl Acad Sci 105: 10925–10930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bunn HF 2007. New agents that stimulate erythropoiesis. Blood 109: 868–873 [DOI] [PubMed] [Google Scholar]
- Cario H, Schwarz K, Jorch N, Kyank U, Petrides PE, Schneider DT, Uhle R, Debatin KM, Kohne E 2005. Mutations in the von Hippel-Lindau (VHL) tumor suppressor gene and VHL-haplotype analysis in patients with presumable congenital erythrocytosis. Haematologica 90: 19–24 [PubMed] [Google Scholar]
- Carnot P, Deflandre C 1906. Sur l’activite hemopoietique des differents organeau au cours de la regeneration du sang. CR Searces Acad Sci 143: 432–435 [Google Scholar]
- Caro J, Brown S, Miller O, Murray T, Erslev AJ 1979. Erythropoietin levels in uremic nephric and anephric patients. J Lab Clin Med 93: 449–458 [PubMed] [Google Scholar]
- Cheetham JC, Smith DM, Aoki KH, Stevenson JL, Hoeffel TJ, Syed RS, Egrie J, Harvey TS 1998. NMR structure of human erythropoietin and a comparison with its receptor bound conformation. Nat Struct Biol 5: 861–866 [DOI] [PubMed] [Google Scholar]
- Chen ZY, Asavaritikrai P, Prchal JT, Noguchi CT 2007. Endogenous erythropoietin signaling is required for normal neural progenitor cell proliferation. J Biol Chem 282: 25875–25883 [DOI] [PubMed] [Google Scholar]
- Chen J, Connor KM, Aderman CM, Smith LE 2008. Erythropoietin deficiency decreases vascular stability in mice. J Clin Invest 118: 526–533 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark DA, Dessypris EN 1986. Effects of recombinant erythropoietin on murine megakaryocytic colony formation in vitro. J Lab Clin Med 108: 423–429 [PubMed] [Google Scholar]
- Cockman ME, Masson N, Mole DR, Jaakkola P, Chang GW, Clifford SC, Maher ER, Pugh CW, Ratcliffe PJ, Maxwell PH 2000. Hypoxia inducible factor-α binding and ubiquitylation by the von Hippel-Lindau tumor suppressor protein. J Biol Chem 275: 25733–25741 [DOI] [PubMed] [Google Scholar]
- Cotes PM 1989. Physiological studies of erythropoietin in plasma. In Erythropoietin (ed. Jelkmann W, Gross AJ), pp. 57–79 Springer, Berlin [Google Scholar]
- Dame C, Bartmann P, Wolber E, Fahnenstich H, Hofmann D, Fandrey J 2000. Erythropoietin gene expression in different areas of the developing human central nervous system. Brain Res Dev Brain Res 125: 69–74 [DOI] [PubMed] [Google Scholar]
- Dame C, Sola MC, Lim KC, Leach KM, Fandrey J, Ma Y, Knopfle G, Engel JD, Bungert J 2004. Hepatic erythropoietin gene regulation by GATA-4. J Biol Chem 279: 2955–2961 [DOI] [PubMed] [Google Scholar]
- Digicaylioglu M, Bichet S, Marti HH, Wenger RH, Rivas LA, Bauer C, Gassmann M 1995. Localization of specific erythropoietin binding sites in defined areas of the mouse brain. Proc Natl Acad Sci 92: 3717–3720 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebert BL, Bunn HF 1998. Regulation of transcription by hypoxia requires a multiprotein complex that includes hypoxia-inducible factor 1, an adjacent transcription factor, and pp300/CREB binding protein. Mol Cell Biol 18: 4089–4096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrenreich H, Hasselblatt M, Dembowski C, Cepek L, Lewczuk P, Stiefel M, Rustenbeck HH, Breiter N, Jacob S, Knerlich F, et al. 2002. Erythropoietin therapy for acute stroke is both safe and beneficial. Mol Med 8: 495–505 [PMC free article] [PubMed] [Google Scholar]
- Ehrenreich H, Weissenborn K, Prange H, Schneider D, Weimar C, Wartenberg K, Schellinger PD, Bohn M, Becker H, Wegrzyn M, et al. 2009. Recombinant human erythropoietin in the treatment of acute ischemic stroke. Stroke 40: e647–e656 [DOI] [PubMed] [Google Scholar]
- Elliott S 2008. Erythropoiesis-stimulating agents and other methods to enhance oxygen transport. Brit J Pharmacol 154: 529–541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott S, Lorenzini T, Chang D, Barzilay J, Delorme E, Giffin J, Hesterberg L 1996. Fine-structure epitope mapping of anti-erythropoietin monoclonal antibodies reveals a model of recombinant human erythropoietin protein structure. Blood 87: 2702–2713 [PubMed] [Google Scholar]
- Elliott S, Lorenzini T, Chang D, Barzilay J, Delorme E 1997. Mapping of the active site of recombinant human erythropoietin. Blood 89: 493–502 [PubMed] [Google Scholar]
- Epstein AC, Gleadle JM, McNeill LA, Hewitson KS, O’Rourke J, Mole DR, Mukherji M, Metzen E, Wilson MI, Dhanda A, et al. 2001. C. elegans EGL-9 and mammalian homologs define a family of dioxygenases that regulate HIF by prolyl hydroxylation. Cell 107: 43–54 [DOI] [PubMed] [Google Scholar]
- Erslev A 1953. Humoral regulation of red cell production. Blood 8: 349–357 [PubMed] [Google Scholar]
- Erslev A 1980. Blood and mountains. In Blood, pure and eloquent (ed. Wintrobe M), pp. 256–280 McGraw Hill, New York [Google Scholar]
- Erslev AJ 1993. The discovery of erythropoietin. ASAIO J 39: 89–92 [PubMed] [Google Scholar]
- Erslev AJ, Caro J 1984. Pure erythrocytosis classified according to erythropoietin titers. Am J Med 76: 57–61 [DOI] [PubMed] [Google Scholar]
- Erslev AJ, Caro J, Besarab A 1985. Why the kidney? Nephron 41: 213–216 [DOI] [PubMed] [Google Scholar]
- Erslev AJ, Wilson J, Caro J 1987. Erythropoietin titers in anemic, nonuremic patients. J Lab Clin Med 109: 429–433 [PubMed] [Google Scholar]
- Eschbach JW, Ergie JC, Downing MR, Browne JK, Adamson JW 1987. Correction of the anemia of endstage renal disease with recombinant human erythropoietin. N Engl J Med 316: 73–78 [DOI] [PubMed] [Google Scholar]
- Fandrey J 2004. Oxygen-dependent and tissue-specific regulation of erythropoietin gene expression. Am J Physiol Regul Integr Comp Physiol 286: R977–R988 [DOI] [PubMed] [Google Scholar]
- Fiordaliso F, Chimenti S, Staszewsky L, Bai A, Carlo E, Cuccovillo I, Doni M, Mengozzi M, Tonelli R, Ghezzi P, et al. 2005. A nonerythropoietic derivative of erythropoietin protects the myocardium from ischemia-reperfusion injury. Proc Natl Acad Sci 102: 2046–2051 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischl M, Galpin JE, Levine JD, Groopman JE, Henry DH, Kennedy P, Miles S, Robbins W, Starrett B, Zalusky R, et al. 1990. Recombinant human erythropoietin for patients with AIDS treated with zidovudine. N Engl J Med 322: 1488–1493 [DOI] [PubMed] [Google Scholar]
- Fox HM 1947. Daphnia haemoglobin. Nature 160: 431. [DOI] [PubMed] [Google Scholar]
- Fox HM 1949. Haemoglobin in Crustacea. Nature 164: 59. [DOI] [PubMed] [Google Scholar]
- Frede S, Freitag P, Geuting L, Konietzny R, Fandrey J 2011. Oxygen-regulated expression of the erythropoietin gene in the human renal cell line REPC. Blood 117: 4905–4914 [DOI] [PubMed] [Google Scholar]
- Fried W, Plzak L, Jacobson L, Goldwasser E 1956. Studies on erythropoiesis II. Assay of erythropoietin in hypophysectomized rats. Proc Soc Exp Biol Med 92: 203–207 [DOI] [PubMed] [Google Scholar]
- Furlow PW, Percy MJ, Sutherland S, Bierl C, McMullin MF, Master SR, Lappin TR, Lee FS 2009. Erythrocytosis-associated HIF-2α mutations demonstrate a critical role for residues C-terminal to the hydroxylacceptor proline. J Biol Chem 284: 9050–9058 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gale DP, Harten SK, Reid CD, Tuddenham EG, Maxwell PH 2008. Autosomal dominant erythrocytosis and pulmonary arterial hypertension associated with an activating HIF2 α mutation. Blood 112: 919–921 [DOI] [PubMed] [Google Scholar]
- Galson DL, Tsuchiya T, Tendler DS, Huang LE, Ren Y, Ogura T, Bunn HF 1995. The orphan receptor hepatic nuclear factor 4 functions as a transcriptional activator for tissue-specific and hypoxia-specific erythropoetin gene expression and is antagonized by EAR3/COUP-TF1. Mol Cell Biol 15: 2135–2144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gnarra JR, Zhou S, Merrill MJ, Wagner JR, Krumm A, Papavassiliou E, Oldfield EH, Klausner RD, Linehan WM 1996. Post-transcriptional regulation of vascular endothelial growth factor mRNA by the product of the VHL tumor suppressor gene. Proc Natl Acad Sci 93: 10589–10594 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg MA, Glass GA, Cunningham JM, Bunn HF 1987. The regulated expression of erythropoietin by two human hepatoma cell lines. Proc Natl Acad Sci 84: 7972–7976 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldwasser E 1996. Erythropoietin, a somewhat personal history. Perspect Biol Med 40: 18–32 [DOI] [PubMed] [Google Scholar]
- Goldwasser E, Kung CK-H, Eliason J 1974. On the mechanism of erythropoietin-induced differentiation. J Biol Chem 249: 4202–4206 [PubMed] [Google Scholar]
- Gordeuk VR, Prchal JT 2006. Vascular complications in Chuvash polycythemia. Semin Thromb Hemost 32: 289–294 [DOI] [PubMed] [Google Scholar]
- Gorr TA, Cahn JD, Yamagata H, Bunn HF 2004. Hypoxia-induced synthesis of hemoglobin in the crustacean Daphnia magna is hypoxia-inducible factor-dependent. J Biol Chem 279: 36038–36047 [DOI] [PubMed] [Google Scholar]
- Gradin K, McGuire J, Wenger RH, Kvietikova I, Whitelaw M, Toftgard R, Tora L, Gassman M, Poellinger L 1996. Functional interference between hypoxia and dioxin signal transduction pathways: Competition for recruitment of the Arnt transcription factor. Mol Cell Biol 16: 5221–5231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant WC, Root WS 1952. Fundamental stimulus for erythropoiesis. Physiol Rev 32: 449–498 [DOI] [PubMed] [Google Scholar]
- Gruber M, Hu CJ, Johnson RS, Brown EJ, Keith B, Simon MC 2007. Acute postnatal ablation of Hif-2α results in anemia. Proc Natl Acad Sci 104: 2301–2306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu YZ, Moran SM, Hogenesch JB, Wartman L, Bradfield CA 1998. Molecular characterization and chromosomal localization of a third α-class hypoxia inducible factor subunit, HIF3α. Gene Expr 7: 205–213 [PMC free article] [PubMed] [Google Scholar]
- Heeschen C, Aicher A, Lehmann R, Fichtlscherer S, Vasa M, Urbich C, Mildner-Rihm C, Martin H, Zeiher AM, Dimmeler S 2003. Erythropoietin is a potent physiologic stimulus for endothelial progenitor cell mobilization. Blood 102: 1340–1346 [DOI] [PubMed] [Google Scholar]
- Henke M, Laszig R, Rube C, Schafer U, Haase KD, Schilcher B, Mose S, Beer KT, Burger U, Dougherty C, et al. 2003. Erythropoietin to treat head and neck cancer patients with anaemia undergoing radiotherapy: Randomised, double-blind, placebo-controlled trial. Lancet 362: 1255–1260 [DOI] [PubMed] [Google Scholar]
- Hermine O, Beru N, Pech N, Goldwasser E 1991. An autocrine role for erythropoietin in mouse hematopoietic cell differentiation. Blood 78: 2253. [PubMed] [Google Scholar]
- Hickey MM, Lam JC, Bezman NA, Rathmell WK, Simon MC 2007. von Hippel-Lindau mutation in mice recapitulates Chuvash polycythemia via hypoxia-inducible factor-2α signaling and splenic erythropoiesis. J Clin Invest 117: 3879–3889 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman EC, Reyes H, Chu F-F, Sander F, Conley LH, Brooks BA, Hankinson O 1991. Cloning of a factor required for activity of the Ah (dioxin) receptor. Science 252: 954–958 [DOI] [PubMed] [Google Scholar]
- Horiguchi H, Kayama F, Oguma E, Willmore WG, Hradecky P, Bunn HF 2000. Cadmium and platinum suppression of erythropoietin production in cell culture. Blood 96: 3743–3747 [PubMed] [Google Scholar]
- Hu B, Wright E, Campbell L, Blanchard KL 1997. In vivo analysis of DNA-protein interactions on the human erythropoietin enhancer. Mol Cell Biol 17: 851–856 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang LE, Arany Z, Livingston DM, Bunn HF 1996. Activation of hypoxia-inducible transcription factor depends primarily upon redox-sensitive stabilization of its a subunit. J Biol Chem 271: 32253–32259 [DOI] [PubMed] [Google Scholar]
- Huang LE, Gu J, Schau M, Bunn HF 1998. Regulation of hypoxia-inducible factor 1α is mediated by an oxygen-dependent degradation domain via the ubiquitin-proteasome pathway. Proc Natl Acad Sci 95: 7987–7992 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iliopoulos O, Levy AP, Jiang CWG, Kaelin J, Goldberg MA 1996. Negative regulation of hypoxia-inducible genes by the von Hippel-Lindau protein. Proc Natl Acad Sci 93: 10595–10599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imagawa S, Izumi T, Miura Y 1994. Positive and negative regulation of the erythropoietin gene. J Biol Chem 269: 9038–9044 [PubMed] [Google Scholar]
- Imagawa S, Yamamoto M, Miura Y 1997. Negative regulation of the erythropoietin gene expression by the GATA transcription factors. Blood 89: 1430–1439 [PubMed] [Google Scholar]
- Ivan M, Kondo K, Yang H, Kim W, Valiando J, Ohh M, Salic A, Asara JM, Lane WS, Kaelin WG Jr 2001. HIFα targeted for VHL-mediated destruction by proline hydroxylation: Implications for O2 sensing. Science 292: 464–468 [DOI] [PubMed] [Google Scholar]
- Ivan M, Haberberger T, Gervasi DC, Michelson KS, Gunzler V, Kondo K, Yang H, Sorokina I, Conaway RC, Conaway JW, et al. 2002. Biochemical purification and pharmacological inhibition of a mammalian prolyl hydroxylase acting on hypoxia-inducible factor. Proc Natl Acad Sci 99: 13459–13464 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iyer NV, Kotch LE, Agani F, Leung SW, Laughner E, Wenger RH, Gassmann M, Gearhart JD, Lawler AM, Yu AY, et al. 1998. Cellular and developmental control of O2 homeostasis by hypoxia-inducible factor 1α. Genes Dev 12: 149–162 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaakkola P, Mole DR, Tian YM, Wilson MI, Gielbert J, Gaskell SJ, Kriegsheim A, Hebestreit HF, Mukherji M, Schofield CJ, et al. 2001. Targeting of HIF-α to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science 292: 468–472 [DOI] [PubMed] [Google Scholar]
- Jacobs K, Shoemaker C, Rudersdorf R, Neill SD, Kaufman RJ, Mufson A, Seehra J, Jones SS, Hewick R, Fritsch EF, et al. 1985. Isolation and characterization of genomic and cDNA clones of human erythropoietin. Nature 313: 806–810 [DOI] [PubMed] [Google Scholar]
- Jacobson LO, Marks E, Gaston E, Goldwasser E 1957. Role of the kidney in erythropoiesis. Nature 179: 633–634 [DOI] [PubMed] [Google Scholar]
- Jelkmann W 1992. Erythropietin: Structure, control of production, and function. Physiol Rev 72: 449–489 [DOI] [PubMed] [Google Scholar]
- Jelkmann W 2005. Effects of erythropoietin on brain function. Curr Pharm Biotechnol 6: 65–79 [DOI] [PubMed] [Google Scholar]
- Jelkmann W 2008. Developments in the therapeutic use of erythropoiesis stimulating agents. Brit J Haematol 141: 287–297 [DOI] [PubMed] [Google Scholar]
- Jelkmann W, Wagner K 2004. Beneficial and ominous aspects of the pleiotropic action of erythropoietin. Ann Hematol 83: 673–686 [DOI] [PubMed] [Google Scholar]
- Kapitsinou PP, Liu Q, Unger TL, Rha J, Davidoff O, Keith B, Epstein JA, Moores SL, Erickson-Miller CL, Haase VH 2010. Hepatic HIF-2 regulates erythropoietic responses to hypoxia in renal anemia. Blood 116: 3039–3048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koury MJ, Bondurant MC 1990. Erythropoietin retards DNA breakdown and prevents programmed death in erythroid progenitor cells. Science 248: 378–381 [DOI] [PubMed] [Google Scholar]
- Koury ST, Bondurant MC, Koury MJ 1988. Localization of erythropoietin synthesizing cells in murine kidneys by in situ hybridization. Blood 71: 524–527 [PubMed] [Google Scholar]
- Koury ST, Koury MJ, Bondurant MC, Caro J, Graber SE 1989. Quantitation of erythropoietin-producing cells in kidneys of mice by in situ hybridization: Correlation with hematocrit, renal erythopoietin mRNA, and serum erythropoietin concentration. Blood 74: 645–651 [PubMed] [Google Scholar]
- Koury ST, Bondurant MC, Koury MJ, Semenza GL 1991. Localization of cells producing erythropoietin in murine liver by in situ hybridization. Blood 77: 2497–2503 [PubMed] [Google Scholar]
- Krumdieck N 1943. Erythropoietic substance in the serum of anemic animals. Proc Soc Exp Biol Med 54: 14–17 [Google Scholar]
- Lacombe C, Silva J-LD, Bruneval P, Fournier J-G, Wendling F, Casadevall N, Camilleri J-P, Bariety J, Varet B, Tambourin P 1988. Pertubular cells are the site of erythropoietin synthesis in the murine hypoxic kidney. J Clin Invest 81: 620–623 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ladroue C, Carcenac R, Leporrier M, Gad S, Le Hello C, Galateau-Salle F, Feunteun J, Pouyssegur J, Richard S, Gardie B 2008. PHD2 mutation and congenital erythrocytosis with paraganglioma. N Engl J Med 359: 2685–2692 [DOI] [PubMed] [Google Scholar]
- La Ferla K, Reimann C, Jelkmann W, Hellwig-Burgel T 2002. Inhibition of erythropoietin gene expression signaling involves the transcription factors GATA-2 and NF-κB. FASEB J 16: 1811–1813 [DOI] [PubMed] [Google Scholar]
- Lando D, Peet DJ, Gorman JJ, Whelan DA, Whitelaw ML, Bruick RK 2002a. FIH-1 is an asparaginyl hydroxylase enzyme that regulates the transcriptional activity of hypoxia-inducible factor. Genes Dev 16: 1466–1471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lando D, Peet DJ, Whelan DA, Gorman JJ, Whitelaw ML 2002b. Asparagine hydroxylation of the HIF transactivation domain a hypoxic switch. Science 295: 858–861 [DOI] [PubMed] [Google Scholar]
- Leist M, Ghezzi P, Grasso G, Bianchi R, Villa P, Fratelli M, Savino C, Bianchi M, Nielsen J, Gerwien J, et al. 2004. Derivatives of erythropoietin that are tissue protective but not erythropoietic. Science 305: 239–242 [DOI] [PubMed] [Google Scholar]
- Le Minh K, Klemm K, Abshagen K, Eipel C, Menger MD, Vollmar B 2007. Attenuation of inflammation and apoptosis by pre- and posttreatment of darbepoetin-α in acute liver failure of mice. Am J Pathol 170: 1954–1963 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leyland-Jones B 2003. Breast cancer trial with erythropoietin terminated unexpectedly. Lancet Oncol 4: 459–460 [DOI] [PubMed] [Google Scholar]
- Lin F-K, Suggs S, Lin C-H, Browne JK, Smalling R, Egrie JC, Chen KK, Fox GM, Martin F, Stabinsky Z, et al. 1985. Cloning and expression of the human erythropoietin gene. Proc Natl Acad Sci 82: 7580–7584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livnah O, Stura EA, Johnson DL, Middleton SA, Mulcahy LS, Wrighton NC, Dower WJ, Jolliffe LK, Wilson IA 1996. Functional mimicry of a protein hormone by a peptide agonist: The EPO receptor complex at 2.8 A. Science 273: 464–471 [DOI] [PubMed] [Google Scholar]
- Livnah O, Stura EA, Middleton SA, Johnson DL, Jolliffe LK, Wilson IA 1999. Crystallographic evidence for preformed dimers of erythropoietin receptor before ligand activation. Science 283: 987–990 [DOI] [PubMed] [Google Scholar]
- Macdougall IC 2005. CERA (Continuous erythropoietin receptor activator): A new erythropoiesis-stimulating agent for the treatment of anemia. Curr Hematol Rep 4: 436–440 [PubMed] [Google Scholar]
- Macdougall IC, Robson R, Opatrna S, Liogier X, Pannier A, Jordan P, Dougherty FC, Reigner B 2006. Pharmacokinetics and pharmacodynamics of intravenous and subcutaneous continuous erythropoietin receptor activator (C.E.R.A.) in patients with chronic kidney disease. Clin J Am Soc Nephrol 1: 1211–1215 [DOI] [PubMed] [Google Scholar]
- Madan A, Lin C, Hatch SL, Curtin PT 1995. Regulated basal, inducible, and tissue-specific human erythropoietin gene expression in transgenic mice requires multiple cis DNA sequences. Blood 85: 2735–2741 [PubMed] [Google Scholar]
- Mahon PC, Hirota K, Semenza GL 2001. FIH-1: A novel protein that interacts with HIF-1α and VHL to mediate repression of HIF-1 transcriptional activity. Genes Dev 15: 2675–2686 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makita T, Hernandez-Hoyos G, Chen TH, Wu H, Rothenberg EV, Sucov HM 2001. A developmental transition in definitive erythropoiesis: Erythropoietin expression is sequentially regulated by retinoic acid receptors and HNF4. Genes Dev 15: 889–901 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marti HH, Wenger RH, Rivas LA, Straumann U, Digicaylioglu M, Henn V, Yonekawa Y, Bauer C, Gassmann M 1996. Erythropoietin gene expression in human, monkey and murine brain. Eur J Neurosci 8: 666–676 [DOI] [PubMed] [Google Scholar]
- Martini M, Teofili L, Cenci T, Giona F, Torti L, Rea M, Foa R, Leone G, Larocca LM 2008. A novel heterozygous HIF2AM535I mutation reinforces the role of oxygen sensing pathway disturbances in the pathogenesis of familial erythrocytosis. Haematologica 93: 1068–1071 [DOI] [PubMed] [Google Scholar]
- Masson N, Willam C, Maxwell PH, Pugh CW, Ratcliffe PJ 2001. Independent function of two destruction domains in hypoxia-inducible factor-α chains activated by prolyl hydroxylation. EMBO J 20: 5197–5206 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masuda S, Okano M, Yamagishi K, Nagao M, Ueda M, Sasaki R 1994. A novel site of erythropoietin production: Oxygen-dependent production in cultured rat astrocytes. J Biol Chem 269: 19488–19493 [PubMed] [Google Scholar]
- Maxwell PH, Osmond MK, Pugh CW, Heryet A, Nicholls LG, Tan CC, Doe BG, Ferguson DJP, Johnson MH, Ratcliffe PJ 1993a. Identification of the renal erythropoietin-producing cells using transgenic mice. Kidney Int 44: 1149–1162 [DOI] [PubMed] [Google Scholar]
- Maxwell PH, Pugh CW, Ratcliffe PJ 1993b. Inducible operation of the erythropoietin 3′ enhancer in multiple cell lines: Evidence for a widespread oxygen-sensing mechanism. Proc Natl Acad Sci 90: 2423–2427 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maxwell PH, Ferguson DJP, Osmond MK, Pugh CW, Heryet A, Doe BG, Johnson MH, Ratcliffe PJ 1994. Expression of a homologously recombined erythopoietin-SV40 T antigen fusion gene in mouse liver: Evidence for erythropoietin production by Ito cells. Blood 84: 1823–1830 [PubMed] [Google Scholar]
- Maxwell PH, Wiesener MS, Chang G-W, Clifford SC, Vaux EC, Cockman ME, Wykoff CC, Pugh CW, Maher ER, Ratcliffe PJ 1999. The tumor suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature 399: 271–275 [DOI] [PubMed] [Google Scholar]
- McDonald TP, Cottrell MB, Clift RE, Cullen WC, Lin FK 1987. High doses of recombinant erythropoietin stimulate platelet production in mice. Exp Hematol 15: 719–721 [PubMed] [Google Scholar]
- McLeod DL, Shreeve MM, Axelrad AA 1974. Improved plasma culture system for production of erythrocytic colonies in vitro: Quantitative assay method for CFU-E. Blood 44: 517–534 [PubMed] [Google Scholar]
- Means RT Jr 1994. Clinical application of recombinant erythropoietin in the anemia of chronic disease. Hematol Oncol Clin North Am 8: 933–944 [PubMed] [Google Scholar]
- Messinezy M, Westwood NB, El-Hemaidi I, Marsden JT, Sherwood RS, Pearson TC 2002. Serum erythropoietin values in erythrocytoses and in primary thrombocythaemia. Brit J Haematol 117: 47–53 [DOI] [PubMed] [Google Scholar]
- Miescher F 1893. Über die Beziehungen zwischen Meereshöhe und Beschaffenheit des Blutes. Corresp Bl Schweiz Aerzte 23: 809–830 [Google Scholar]
- Miller CB, Jones RJ, Piantadosi S, Abeloff MD, Spivak JL 1990. Decreased erythropoietin response in patients with the anemia of cancer. N Engl J Med 322: 1689–1692 [DOI] [PubMed] [Google Scholar]
- Minamishima YA, Moslehi J, Bardeesy N, Cullen D, Bronson RT, Kaelin WG Jr 2008. Somatic inactivation of the PHD2 prolyl hydroxylase causes polycythemia and congestive heart failure. Blood 111: 3236–3244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyake T, Hung CKH, Goldwasser E 1977. Purification of human erythropoietin. J Biol Chem 252: 5558–5564 [PubMed] [Google Scholar]
- Mohyeldin A, Lu H, Dalgard C, Lai SY, Cohen N, Acs G, Verma A 2005. Erythropoietin signaling promotes invasiveness of human head and neck squamous cell carcinoma. Neoplasia 7: 537–543 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nathan DG, Schupak E, Stohlman F Jr, Merrill JP 1964. Erythropoiesis in anephric man. J Clin Invest 43: 2158–2165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishiya D, Omura T, Shimada K, Matsumoto R, Kusuyama T, Enomoto S, Iwao H, Takeuchi K, Yoshikawa J, Yoshiyama M 2006. Effects of erythropoietin on cardiac remodeling after myocardial infarction. J Pharm Sci 101: 31–39 [DOI] [PubMed] [Google Scholar]
- Noguchi CT, Asavaritikrai P, Teng R, Jia Y 2007. Role of erythropoietin in the brain. Crit Rev Oncol Hemat 64: 159–171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noguchi CT, Wang L, Rogers HM, Teng R, Jia Y 2008. Survival and proliferative roles of erythropoietin beyond the erythroid lineage. Expert Rev Mol Med 10: e36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obara N, Suzuki N, Kim K, Nagasawa T, Imagawa S, Yamamoto M 2008. Repression via the GATA box is essential for tissue-specific erythropoietin gene expression. Blood 111: 5223–5232 [DOI] [PubMed] [Google Scholar]
- Ohh M, Park CW, Ivan M, Hoffman MA, Kim TY, Huang LE, Pavletich N, Chau V, Kaelin WG 2000. Ubiquitination of hypoxia-inducible factor requires direct binding to the β-domain of the von Hippel-Lindau protein. Nat Cell Biol 2: 423–427 [DOI] [PubMed] [Google Scholar]
- Paliege A, Rosenberger C, Bondke A, Sciesielski L, Shina A, Heyman SN, Flippin LA, Arend M, Klaus SJ, Bachmann S 2010. Hypoxia-inducible factor-2α-expressing interstitial fibroblasts are the only renal cells that express erythropoietin under hypoxia-inducible factor stabilization. Kidney Int 77: 312–318 [DOI] [PubMed] [Google Scholar]
- Parsa CJ, Kim J, Riel RU, Pascal LS, Thompson RB, Petrofski JA, Matsumoto A, Stamler JS, Koch WJ 2004. Cardioprotective effects of erythropoietin in the reperfused ischemic heart: A potential role for cardiac fibroblasts. J Biol Chem 279: 20655–20662 [DOI] [PubMed] [Google Scholar]
- Pastore YD, Jelinek J, Ang S, Guan Y, Liu E, Jedlickova K, Krishnamurti L, Prchal JT 2003. Mutations in the VHL gene in sporadic apparently congenital polycythemia. Blood 101: 1591–1595 [DOI] [PubMed] [Google Scholar]
- Patel NS, Sharples EJ, Cuzzocrea S, Chatterjee PK, Britti D, Yaqoob MM, Thiemermann C 2004. Pretreatment with EPO reduces the injury and dysfunction caused by ischemia/reperfusion in the mouse kidney in vivo. Kidney Int 66: 983–989 [DOI] [PubMed] [Google Scholar]
- Percy MJ, Zhao Q, Flores A, Harrison C, Lappin TR, Maxwell PH, McMullin MF, Lee FS 2006. A family with erythrocytosis establishes a role for prolyl hydroxylase domain protein 2 in oxygen homeostasis. Proc Natl Acad Sci 103: 654–659 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Percy MJ, Furlow PW, Beer PA, Lappin TR, McMullin MF, Lee FS 2007. A novel erythrocytosis-associated PHD2 mutation suggests the location of a HIF binding groove. Blood 110: 2193–2196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Percy MJ, Beer PA, Campbell G, Dekker AW, Green AR, Oscier D, Rainey MG, van Wijk R, Wood M, Lappin TR, et al. 2008a. Novel exon 12 mutations in the HIF2A gene associated with erythrocytosis. Blood 111: 5400–5402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Percy MJ, Furlow PW, Lucas GS, Li X, Lappin TR, McMullin MF, Lee FS 2008b. A gain-of-function mutation in the HIF2A gene in familial erythrocytosis. N Engl J Med 358: 162–168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Percy MJ, Chung YJ, Harrison C, Mercieca J, Hoffbrand VJ, Dinardo CL, Dantos PCJL, Fonseca GHH, Gualalndro SFM, Pereira AC, et al. 2012. Two new mutations in the HIF2A gene associated with erythrocytosis. Am J Hematol 87: 439–442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrotta S, Nobili B, Ferraro M, Migliaccio C, Borriello A, Cucciolla V, Martinelli V, Rossi F, Punzo F, Cirillo P, et al. 2006. Von Hippel-Lindau-dependent polycythemia is endemic on the island of Ischia: Identification of a novel cluster. Blood 107: 514–519 [DOI] [PubMed] [Google Scholar]
- Peyssonnaux C, Zinkernagel AS, Schuepbach RA, Rankin E, Vaulont S, Haase VH, Nizet V, Johnson RS 2007. Regulation of iron homeostasis by the hypoxia-inducible transcription factors (HIFs). J Clin Invest 117: 1926–1932 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phrommintikul A, Haas SJ, Elsik M, Krum H 2007. Mortality and target haemoglobin concentrations in anaemic patients with chronic kidney disease treated with erythropoietin: A meta-analysis. Lancet 369: 381–388 [DOI] [PubMed] [Google Scholar]
- Prchal JF, Prchal JT 1999. Molecular basis for polycythemia. Curr Opin Hematol 6: 100–109 [DOI] [PubMed] [Google Scholar]
- Provenzano R, Besarab A, Macdougall IC, Ellison DH, Maxwell AP, Sulowicz W, Klinger M, Rutkowski B, Correa-Rotter R, Dougherty FC 2007. The continuous erythropoietin receptor activator (C.E.R.A.) corrects anemia at extended administration intervals in patients with chronic kidney disease not on dialysis: Results of a phase II study. Clin Nephrol 67: 306–317 [PubMed] [Google Scholar]
- Pugh CW, Tan CC, Jones RW, Ratcliffe PJ 1991. Functional analysis of an oxygen-regulated transcriptional enhancer lying 3′ to the mouse erythropoietin gene. Proc Natl Acad Sci 88: 10553–10557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pugh CW, Ebert BL, Ebrahim O, Ratcliffe PJ 1994. Characterisation of functional domains within the mouse erythropoietin 3′ enhancer conveying oxygen-regulated responses in different cell lines. Biochem Biophys Acta 1217: 297–306 [DOI] [PubMed] [Google Scholar]
- Rankin EB, Biju MP, Liu Q, Unger TL, Rha J, Johnson RS, Simon MC, Keith B, Haase VH 2007. Hypoxia-inducible factor-2 (HIF-2) regulates hepatic erythropoietin in vivo. J Clin Invest 117: 1068–1077 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reissmann KR 1950. Studies on the mechanism of erythropoietic stimulation in parabiotic rats during hypoxia. Blood 5: 372–380 [PubMed] [Google Scholar]
- Remy I, Wilson IA, Michnick SW 1999. Erythropoietin receptor activation by a ligand-induced conformation change. Science 283: 990–993 [DOI] [PubMed] [Google Scholar]
- Rosa R, Prehu M-O, Beuzard Y, Rosa J 1978. The first case of a complete deficiency of diphosphoglycerate mutase in human erythrocytes. J Clin Invest 62: 907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruhenstroth-Bauer G 1950. Versuche zum Nachweis eines spezifischen erythropetischen Hormons. Arch Exp Pathol Pharmakol 211: 32–56 [PubMed] [Google Scholar]
- Ryan HE, Lo J, Johnson RS 1998. HIF-1α is required for solid tumor formation and embryonic vascularization. EMBO J 17: 3005–3015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Safran M, Kim WY, O’Connell F, Flippin L, Gunzler V, Horner JW, Depinho RA, Kaelin WG Jr 2006. Mouse model for noninvasive imaging of HIF prolyl hydroxylase activity: Assessment of an oral agent that stimulates erythropoietin production. Proc Natl Acad Sci 103: 105–110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakanaka M, Wen TC, Matsuda S, Masuda S, Morishita E, Nagao M, Sasaki R 1998. In vivo evidence that erythropoietin protects neurons from ischemic damage. Proc Natl Acad Sci 95: 4635–4640 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salceda S, Caro J 1997. Hypoxia-inducible factor 1α (HIF-1α) protein is rapidly degraded by the ubiquitin-proteasome system under normoxic conditions. Its stabilization by hypoxia depends on redox-induced changes. J Biol Chem 272: 22642–22647 [DOI] [PubMed] [Google Scholar]
- Schmeding M, Boas-Knoop S, Lippert S, Ruehl M, Somasundaram R, Dagdelen T, Neuhaus P, Neumann UP 2008. Erythropoietin promotes hepatic regeneration after extended liver resection in rats. J Gastroenterol Hepatol 23: 1125–1131 [DOI] [PubMed] [Google Scholar]
- Schmeding M, Hunold G, Ariyakhagorn V, Rademacher S, Boas-Knoop S, Lippert S, Neuhaus P, Neumann UP 2009. Erythropoietin reduces ischemia-reperfusion injury after liver transplantation in rats. Transpl Int 22: 738–746 [DOI] [PubMed] [Google Scholar]
- Schmid TC, Loffing J, Le Hir M, Kaissling B 1994. Distribution of ecto-5′-nucleotidase in the rat liver: Effect of anaemia. Histochemistry 101: 439–447 [DOI] [PubMed] [Google Scholar]
- Schuster SJ, Koury ST, Bohrer M, Salceda S, Caro J 1992. Cellular sites of extrarenal and renal erythropoietin production in anemic rats. Brit J Hematol 81: 153–159 [DOI] [PubMed] [Google Scholar]
- Scortegagna M, Ding K, Zhang Q, Oktay Y, Bennett MJ, Bennett M, Shelton JM, Richardson JA, Moe O, Garcia JA 2005. HIF-2α regulates murine hematopoietic development in an erythropoietin-dependent manner. Blood 105: 3133–3140 [DOI] [PubMed] [Google Scholar]
- Semenza GL, Wang GL 1992. A nuclear factor induced by hypoxia via de novo protein synthesis binds to the human erythropoietin gene enhancer at a site required for transcriptional activation. Mol Cell Biol 12: 5447–5454 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Semenza GL, Trystman M, Gearhart JD, Antonarakis S 1989. Polycythemia in transgenic mice expressing the human erythropoietin gene. Proc Natl Acad Sci 86: 2301–2305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Semenza GL, Dureza RC, Traystman MD, Gearhart JD, Antonarakis SE 1990. Human erythropoietin gene expression in transgenic mice: Multiple transcription initiation sites and cis-acting regulatory elements. Mol Cell Biol 10: 930–938 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Semenza GL, Koury ST, Nejfelt MK, Gearhart JD, Antonarakis SE 1991a. Cell-type-specific and hypoxia-inducible expression of the human erythropoietin gene in transgenic mice. Proc Natl Acad Sci 88: 8725–8729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Semenza GL, Nejfelt MK, Chi SM, Antonarakis SE 1991b. Hypoxia-inducible nuclear factors bind to an enhancer element located 3′ to the human erythropoietin gene. Proc Natl Acad Sci 88: 5680–5684 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sepodes B, Maio R, Pinto R, Sharples E, Oliveira P, McDonald M, Yaqoob M, Thiemermann C, Mota-Filipe H 2006. Recombinant human erythropoietin protects the liver from hepatic ischemia-reperfusion injury in the rat. Transpl Int 19: 919–926 [DOI] [PubMed] [Google Scholar]
- Sergeyeva A, Gordeuk VR, Tokarev YN, Sokol L, Prchal JH, Prchal JT 1997. Congenital polycythemia in Chuvashia. Blood 89: 2148–2154 [PubMed] [Google Scholar]
- Sharples EJ, Patel N, Brown P, Stewart K, Mota-Philipe H, Sheaff M, Kieswich J, Allen D, Harwood S, Raftery M, et al. 2004. Erythropoietin protects the kidney against the injury and dysfunction caused by ischemia-reperfusion. J Am Soc Nephrol 15: 2115–2124 [DOI] [PubMed] [Google Scholar]
- Simonson TS, Yang Y, Huff CD, Yun H, Qin G, Witherspoon DJ, Bai Z, Lorenzo FR, Xing J, Jorde LB, et al. 2010. Genetic evidence for high-altitude adaptation in Tibet. Science 329: 72–75 [DOI] [PubMed] [Google Scholar]
- Sinclair AM, Rogers N, Busse L, Archibeque I, Brown W, Kassner PD, Watson JE, Arnold GE, Nguyen KC, Powers S, et al. 2008. Erythropoietin receptor transcription is neither elevated nor predictive of surface expression in human tumour cells. Brit J Cancer 98: 1059–1067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh A, Eckardt KU, Zimmermann A, Gotz KH, Hamann M, Ratcliffe PJ, Kurtz A, Reinhart WH 1993. Increased plasma viscosity as a reason for inappropriate erythropoietin formation. J Clin Invest 91: 251–256 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh AK, Szczech L, Tang KL, Barnhart H, Sapp S, Wolfson M, Reddan D 2006. Correction of anemia with epoetin alfa in chronic kidney disease. N Engl J Med 355: 2085–2098 [DOI] [PubMed] [Google Scholar]
- Spivak JL 1994. Recombinant human erythropoietin and the anemia of cancer [editorial]. Blood 84: 997–1004 [PubMed] [Google Scholar]
- Stohlman FJ, Rath C, Rose J 1954. Evidence for a humoral regulation of erythropoiesis. Blood 9: 721–733 [PubMed] [Google Scholar]
- Stopka T, Zivny JH, Stopkova P, Prchal JF, Prchal JT 1998. Human hematopoietic progenitors express erythropoietin. Blood 91: 3766–3772 [PubMed] [Google Scholar]
- Suzuki N, Ohneda O, Takahashi S, Higuchi M, Mukai HY, Nakahata T, Imagawa S, Yamamoto M 2002. Erythroid-specific expression of the erythropoietin receptor rescued its null mutant mice from lethality. Blood 100: 2279–2288 [DOI] [PubMed] [Google Scholar]
- Syed RS, Reid SW, Li C, Cheetham JC, Aoki KH, Liu B, Zhan H, Osslund TD, Chirino AJ, Zhang J, et al. 1998. Efficiency of signalling through cytokine receptors depends critically on receptor orientation. Nature 395: 511–516 [DOI] [PubMed] [Google Scholar]
- Takeda K, Aguila HL, Parikh NS, Li X, Lamothe K, Duan LJ, Takeda H, Lee FS, Fong GH 2008. Regulation of adult erythropoiesis by prolyl hydroxylase domain proteins. Blood 111: 3229–3235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan CC, Eckardt K-U, Ratcliffe PJ 1991. Organ distribution of erythopoietin messenger RNA in normal and uremic rats. Kidney Int 40: 69–76 [DOI] [PubMed] [Google Scholar]
- Tian H, McKnight SL, Russell DW 1997. Endothelial PAS domain protein 1 (EPAS1), a transcription factor selectively expressed in endothelial cells. Genes Dev 11: 72–82 [DOI] [PubMed] [Google Scholar]
- Tsai PT, Ohab JJ, Kertesz N, Groszer M, Matter C, Gao J, Liu X, Wu H, Carmichael ST 2006. A critical role of erythropoietin receptor in neurogenesis and post-stroke recovery. J Neurosci 26: 1269–1274 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Wijk R, Sutherland S, Van Wesel AC, Huizinga EG, Percy MJ, Bierings M, Lee FS 2010. Erythrocytosis associated with a novel missense mutation in the HIF2A gene. Haematologica 95: 829–832 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaziri ND, Zhou XJ, Liao SY 1994. Erythropoietin enhances recovery from cisplatin-induced acute renal failure. Am J Physiol 266: F360–F366 [DOI] [PubMed] [Google Scholar]
- Viault F 1890. Sur l’augmentation considerable du nombre des globules rouges dans le sang chez les habitants des hautes plateaux de l’Amerique du sud. CR Acad Sci Paris 111: 918–919 [Google Scholar]
- Volke M, Gale DP, Maegdefrau U, Schley G, Klanke B, Bosserhoff AK, Maxwell PH, Eckardt KU, Warnecke C 2009. Evidence for a lack of a direct transcriptional suppression of the iron regulatory peptide hepcidin by hypoxia-inducible factors. PloS ONE 4: e7875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang GL, Semenza GL 1993a. Desferrioxamine induces erythropoietin gene expression and hypoxia-inducible factor 1 DNA-binding activity: Implications for models of hypoxia signal transduction. Blood 82: 3610–3615 [PubMed] [Google Scholar]
- Wang GL, Semenza GL 1993b. General involvement of hypoxia-inducible factor 1 in transcriptional response to hypoxia. Proc Natl Acad Sci 90: 4304–4308 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang GL, Semenza GL 1995. Purification and characterization of hypoxia-inducible factor-1. J Biol Chem 270: 1230–1237 [DOI] [PubMed] [Google Scholar]
- Wang GL, Jiang B-H, Rue EA, Semenza GL 1995a. Hypoxia-inducible factor 1 is a basic helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci 92: 5510–5514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang GL, Jiang BH, Rue EA, Semenza GL 1995b. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci 92: 5510–5514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warnecke C, Zaborowska Z, Kurreck J, Erdmann VA, Frei U, Wiesener M, Eckardt KU 2004. Differentiating the functional role of hypoxia-inducible factor (HIF)-1α and HIF-2α (EPAS-1) by the use of RNA interference: Erythropoietin is a HIF-2α target gene in Hep3B and Kelly cells. FASEB J 18: 1462–1464 [DOI] [PubMed] [Google Scholar]
- Wen D, Boissel J-P, Showers M, Ruch BC, Bunn HF 1994. Erythropoietin structure-function relationships: Identification of functionally important domains. J Biol Chem 269: 22839–22846 [PubMed] [Google Scholar]
- Wenger RH, Kurtz A 2011. Erythropoietin. Compr Physiol 1: 1759–1794 [DOI] [PubMed] [Google Scholar]
- Wenger RH, Stiehl DP, Camenisch G 2005. Integration of oxygen signaling at the consensus HRE. Sci STKE 2005: re12. [DOI] [PubMed] [Google Scholar]
- Wood SM, Gleadle JM, Pugh CW, Hankinson O, Ratcliffe PJ 1996. The role of the aryl hydrocarbon receptor nuclear translocator (ARNT) in hypoxic induction of gene expression. J Biol Chem 269: 15117–15123 [DOI] [PubMed] [Google Scholar]
- Wrighton NC, Farrell FX, Chang R, Kashyap AK, Barbone FP, Mulcahy LS, Johnson DL, Barrett RW, Jolliffe LK, Dower WJ 1996. Small peptides as potent mimetics of the protein hormone erythropoietin. Science 273: 458–463 [DOI] [PubMed] [Google Scholar]
- Yamaji R, Okada T, Moriya M, Naito M, Tsuruo T, Miyatake K, Nakano Y 1996. Brain capillary endothelial cells express two forms of erythropoietin receptor mRNA. Eur J Biochem 239: 494–500 [DOI] [PubMed] [Google Scholar]
- Yan L, Colandrea VJ, Hale JJ 2010. Prolyl hydroxylase domain-containing protein inhibitors as stabilizers of hypoxia-inducible factor: Small molecule-based therapeutics for anemia. Expert Opin Ther Pat 20: 1219–1245 [DOI] [PubMed] [Google Scholar]
- Yang CW, Li C, Jung JY, Shin SJ, Choi BS, Lim SW, Sun BK, Kim YS, Kim J, Chang YS, et al. 2003. Preconditioning with erythropoietin protects against subsequent ischemia-reperfusion injury in rat kidney. FASEB J 17: 1754–1755 [DOI] [PubMed] [Google Scholar]
- Yi X, Liang Y, Huerta-Sanchez E, Jin X, Cuo ZX, Pool JE, Xu X, Jiang H, Vinckenbosch N, Korneliussen TS, et al. 2010. Sequencing of 50 human exomes reveals adaptation to high altitude. Science 329: 75–78 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu F, White SB, Zhao Q, Lee FS 2001. HIF-1α binding to VHL is regulated by stimulus-sensitive proline hydroxylation. Proc Natl Acad Sci 98: 9630–9635 [DOI] [PMC free article] [PubMed] [Google Scholar]