

Abstract
Cannabidiol (CBD) is a phytocannabinoid with therapeutic properties for numerous disorders exerted through molecular mechanisms that are yet to be completely identified. CBD acts in some experimental models as an anti-inflammatory, anticonvulsant, anti-oxidant, anti-emetic, anxiolytic and antipsychotic agent, and is therefore a potential medicine for the treatment of neuroinflammation, epilepsy, oxidative injury, vomiting and nausea, anxiety and schizophrenia, respectively. The neuroprotective potential of CBD, based on the combination of its anti-inflammatory and anti-oxidant properties, is of particular interest and is presently under intense preclinical research in numerous neurodegenerative disorders. In fact, CBD combined with Δ9-tetrahydrocannabinol is already under clinical evaluation in patients with Huntington's disease to determine its potential as a disease-modifying therapy. The neuroprotective properties of CBD do not appear to be exerted by the activation of key targets within the endocannabinoid system for plant-derived cannabinoids like Δ9-tetrahydrocannabinol, i.e. CB1 and CB2 receptors, as CBD has negligible activity at these cannabinoid receptors, although certain activity at the CB2 receptor has been documented in specific pathological conditions (i.e. damage of immature brain). Within the endocannabinoid system, CBD has been shown to have an inhibitory effect on the inactivation of endocannabinoids (i.e. inhibition of FAAH enzyme), thereby enhancing the action of these endogenous molecules on cannabinoid receptors, which is also noted in certain pathological conditions. CBD acts not only through the endocannabinoid system, but also causes direct or indirect activation of metabotropic receptors for serotonin or adenosine, and can target nuclear receptors of the PPAR family and also ion channels.
Keywords: cannabidiol, cannabinoid signalling system, Huntington's disease, neonatal ischaemia, neuroprotection, Parkinson's disease
Overview on the therapeutic properties of CBD
Cannabidiol (CBD) is one of the key cannabinoid constituents in the plant Cannabis sativa in which it may represent up to 40% of cannabis extracts [1]. However, contrarily to Δ9-tetrahydrocannabinol (Δ9-THC), the major psychoactive plant-derived cannabinoid, which combines therapeutic properties with some important adverse effects, CBD is not psychoactive (it does not activate CB1 receptors [2]), it is well-tolerated and exhibits a broad spectrum of therapeutic properties [3]. Even, combined with Δ9-THC in the cannabis-based medicine Sativex® (GW Pharmaceuticals Ltd, Kent, UK), CBD is able to enhance the beneficial properties of Δ9-THC while reducing its negative effects [4]. Based on this relatively low toxicity, CBD has been studied, even at the clinical level, alone or combined with other phytocannabinoids, to determine its therapeutic efficacy in different central nervous system (CNS) and peripheral disorders [3]. In the CNS, CBD has been reported to have anti-inflammatory properties, thus being useful for neuroinflammatory disorders [5], including multiple sclerosis for which CBD combined with Δ9-THC (Sativex®) has been recently licenced as a symptom-relieving agent for the treatment of spasticity and pain [6]. Based on its anticonvulsant properties, CBD has been proposed for the treatment of epilepsy [7–9], and also for the treatment of sleep disorders based on its capability to induce sleep [10]. CBD is also anti-emetic, as are most of the cannabinoid agonists, but its effects are independent of CB1 receptors and are possibly related to its capability to modulate serotonin transmission (see [11] and below). CBD has antitumoural properties that explain its potential against various types of cancer [12, 13]. Moreover, CBD has recently shown an interesting profile for psychiatric disorders, for example, it may serve as an antipsychotic and be a promising compound for the treatment of schizophrenia [14–17], but it also has potential as an anxiolytic [18] and antidepressant [19], thus being also effective for other psychiatric disorders. Lastly, based on the combination of its anti-inflammatory and anti-oxidant properties, CBD has been demonstrated to have an interesting neuroprotective profile as indicated by results obtained through intense preclinical research into numerous neurodegenerative disorders, in particular the three disorders addressed in this review, neonatal ischaemia (CBD alone) [20], Huntington's disease (HD) (CBD combined with Δ9-THC as in Sativex®) [21–23] or Parkinson's disease (PD) (CBD probably combined with the phytocannabinoid Δ9-tetrahydrocannabivarin, Δ9-THCV) [24, 25], work that has recently progressed to the clinical area in some specific cases [26]. The neuroprotective potential of CBD for the management of certain other neurodegenerative disorders, e.g. Alzheimer's disease, stroke and multiple sclerosis, has also been investigated in studies that have yielded some positive results [27–33]. However, these data will be considered here only very briefly.
Overview on the mechanisms of action of CBD
The therapeutic properties of CBD do not appear to be exerted by the activation of key targets within the endocannabinoid system for plant-derived cannabinoids like Δ9-THC, i.e. CB1 and CB2 receptors. CBD has in general negligible activity at these cannabinoid receptors [2], so it has been generally assumed that most of its pharmacological effects are not a priori pharmacodynamic in nature and related to the activation of specific signalling pathways, but related to its innate chemical properties, in particular with the presence of two hydroxyl groups (see below) that enables CBD to have an important anti-oxidant action [2]. However, in certain pathological conditions (i.e. damage of immature brain), CBD has shown some activity at the CB2 receptor exerted directly ([20], see also Table 1) or indirectly through an inhibitory effect on the mechanisms of inactivation (i.e. transporter, FAAH enzyme) of endocannabinoids [34, 35], enhancing the action of these endogenous molecules at the CB2 receptor but also at the CB1 and at other receptors for endocannabinoids, i.e. TRPV1 [35] and TRPV2 [36] receptors.
Table 1.
A selection of receptors, ion channels, enzymes and cellular uptake processes that CBD has been reported to activate, antagonize or inhibit in vitro
| CBD concentration | Pharmacological target and effect | Reference |
|---|---|---|
| Receptors and channels | ||
| <1 µm | CB1 receptor (−) | [40] |
| CB2 receptor (−) | [40] | |
| GPR55 (−) | [41]† | |
| 5-HT3A ligand-gated channel (−)* | [42] | |
| TRPM8 cation channel (−) | [36] | |
| TRPA1 cation channel (+) | [36] | |
| 1–10 µm | PPARγ nuclear receptor (+) | [41]† |
| CaV3 T-type Ca2+ channels (−) | [43] | |
| TRPV1 cation channel (+) | [36] | |
| TRPV2 cation channel (+) | [36] | |
| >10 µm | 5-HT1A receptor (+) | [44]† |
| µ and δ opioid receptors (−)* | [44]† | |
| α1 and α1β glycine ligand-gated channels (+)* | [45] | |
| Enzymes | ||
| <1 µm | CYP1A1 (−) | [46] |
| 1–10 µm | CYP1A2 & CYP1B1 (−) | [46] |
| CYP2B6 (−) | [47] | |
| CYP2D6 (−) | [48] | |
| CYP3A5 (−) | [49] | |
| Mg2+-ATPase (−) | [44]† | |
| Arylalkylamine N-acetyltransferase (−) | [50] | |
| Indoleamine-2,3-dioxygenase (−) | [51] | |
| 15-lipoxygenase (−) | [52] | |
| Phospholipase A2 (+) | [44]† | |
| Glutathione peroxidase (+) | [13, 53] | |
| Glutathione reductase (+) | [13, 53] | |
| >10 µm | CYP2A6 (−) | [47] |
| CYP3A4 and CYP3A7 (−) | [49] | |
| Fatty acid amide hydrolase (−) | [36] | |
| 5-lipoxygenase (−) | [52] | |
| Superoxide dismutase (−) | [53] | |
| Catalase (−) | [53] | |
| NAD(P)H-quinone reductase (−) | [53] | |
| Progesterone 17α-hydroxylase (−) | [54, 55] | |
| Testosterone 6β-hydroxylase (−) | [54] | |
| Testosterone 16α-hydroxylase (−) | [54] | |
| Transporters and cellular uptake | ||
| <1 µm | Adenosine uptake by cultured microglia and macrophages (−) | [44]† |
| Calcium uptake by synaptosomes (−) | [44]† | |
| 1–10 µm | NE, DA, 5-HT and GABA uptake by synaptosomes (−) | [44]† |
| Anandamide and palmitoylethanolamide cellular uptake (−) | [36] | |
| P-glycoprotein (drug efflux transporter) (−) | [56] | |
| >10 µm | Choline uptake by rat hippocampal homogenates (−) | [44]† |
5-HT, 5-hydroxytryptamine; DA, dopamine; GABA, γ-aminobutyric acid; NE, norepinephrine.
Apparent allosteric modulation; (+). activation; (−), inhibition or antagonism.
Review article.
However, the anti-oxidant profile of CBD, as well as the few effects it exerts through targets within the endocannabinoid system in certain pathophysiological conditions, cannot completely explain all of the many pharmacological effects of CBD, prompting a need to seek out possible targets for this phytocannabinoid outside the endocannabinoid system. There is, indeed, already evidence that CBD can affect serotonin receptors (i.e. 5HT1A) [18, 19, 28], adenosine uptake [37], nuclear receptors of the PPAR family (i.e. PPAR-γ) [38, 39] and many other pharmacological targets (see Table 1 including references [40–56]). In part, this information derives from numerous studies directed at identifying the pharmacological actions that CBD produces in vitro. This phytocannabinoid has been found to display a wide range of actions in vitro some at concentrations in the submicromolar range, and others at concentrations between 1 and 10 µm or above 10 µm. Its pharmacological targets include a number of receptors, ion channels, enzymes and cellular uptake processes (summarized in Table 1). There is evidence too that CBD can inhibit delayed rectifier K+ and L-type Ca2+ currents and evoked human neutrophil migration, activate basal microglial cell migration, and increase membrane fluidity, all at submicromolar concentrations, and that at concentrations between 1 and 10 µm it can inhibit the proliferation of human keratinocytes and of certain cancer cells (reviewed in [44]). At concentrations between 1 and 10 µm, CBD has also been reported to be neuroprotective, to reduce signs of oxidative stress, to modulate cytokine release and to increase calcium release from neuronal and glial intracellular stores (reviewed in [44]), and at 15 µM to induce mRNA expression of several phosphatases in prostate and colon cancer cells [57].
As will be discussed in the following section, the question of which of these many actions contributes most towards the beneficial effects that CBD displays in vivo in animal models of neurodegenerative disorders such as PD and HD remains to be fully investigated. Also still to be explored is the possibility that CBD may ameliorate signs and symptoms of such disorders and others (i.e. psychiatric disorders), at least in part, by potentiating activation of 5-HT1A receptors by endogenously released serotonin. Thus, although CBD only activates the 5-HT1A receptor at concentrations above 10 µm (Table 1), it can, at the much lower concentration of 100 nm enhance the ability of the 5-HT1A receptor agonist, 8-hydroxy-2-(di-n-propylamino)tetralin to stimulate [35S]-GTPγS binding to rat brainstem membranes [58]. Furthermore, there is evidence first, that activation of 5-HT1A receptors can ameliorate specific symptoms in PD [59, 60] and second, that beneficial effects displayed by CBD in vivo in animal models of ischaemic injury [27, 28], hepatic encephalopathy [61], anxiety, stress and panic [18, 62–64], depression [19], pain [65] and nausea and vomiting [66] are all mediated by increased activation of the 5-HT1A receptor. Importantly, the dose–response curve of CBD for the production of its effects in several of these models has been found to be bell-shaped [19, 28, 62, 65, 67, 68]. This is a significant observation since it strengthens the hypothesis that CBD can act in vivo to potentiate 5-HT-induced activation of 5-HT1A receptors. Thus, the concentration–response curve of CBD for its enhancement of 8-hydroxy-2-(di-n-propylamino)tetralin-induced stimulation of [35S]-GTPγS binding to rat brainstem membranes is also bell-shaped [58].
CBD as a neuroprotective agent
In contrast to the neuroprotective properties of cannabinoid receptor agonists [69, 70], those of CBD do not seem to be attributable to the control of excitotoxicity via the activation of CB1 receptors and/or to the control of microglial toxicity via the activation of CB2 receptors. Thus, except in preclinical models of neonatal ischaemia (see below and [20]), CBD has been found not to display any signs of CB1 or CB2 receptor activation, and yet is no less active than cannabinoid receptor agonists against the brain damage produced by different types of cytotoxic insults ([71–75], reviewed in [76]). What then are the cannabinoid receptor-independent mechanisms by which CBD acts as a neuroprotective agent? Finding the correct answer to this question is not easy, although data obtained in numerous investigations into different pathological conditions associated with brain damage indicate that CBD does normalize glutamate homeostasis [71, 72], reduce oxidative stress [73, 77] and attenuate glial activation and the occurrence of local inflammatory events [74, 78]. Furthermore, a recent study by Juknat et al. [79] has strongly demonstrated the existence of notable differences in the genes that were altered by CBD (not active at CB1 or CB2 receptors) and those altered by Δ9-THC (active at both these receptors) in inflammatory conditions in an in vitro model. These authors found a greater influence of CBD on genes controlled by nuclear factors known to be involved in the regulation of stress responses (including oxidative stress) and inflammation [79]. This agrees with the idea that there may be two key processes underlying the neuroprotective effects of CBD. The first and the most classic mechanism is the capability of CBD to restore the normal balance between oxidative events and anti-oxidant endogenous mechanisms [69] that is frequently disrupted in neurodegenerative disorders, thereby enhancing neuronal survival. As has been mentioned above [73, 77], this capability seems to be inherent to CBD and structurally-similar compounds, i.e. Δ9-THC, cannabinol, nabilone, levonantradol and dexanabinol, as it would depend on the innate anti-oxidant properties of these compounds and be cannabinoid receptor-independent. Alternatively, or in addition, the anti-oxidant effect of CBD may involve intracellular mechanisms that enhance the ability of endogenous anti-oxidant enzymes to control oxidative stress, in particular the signaling triggered by the transcription factor nuclear factor-erythroid 2-related Factor 2 (nrf-2), as has been found in the case of other classic anti-oxidants. According to this idea, CBD may bind to an intracellular target capable of regulating this transcription factor which plays a major role in the control of anti-oxidant-response elements located in genes encoding for different anti-oxidant enzymes of the so-called phase II-anti-oxidant response (see proposed mechanism in Figure 1). This possibility is presently under investigation (reviewed in [69]).
Figure 1.
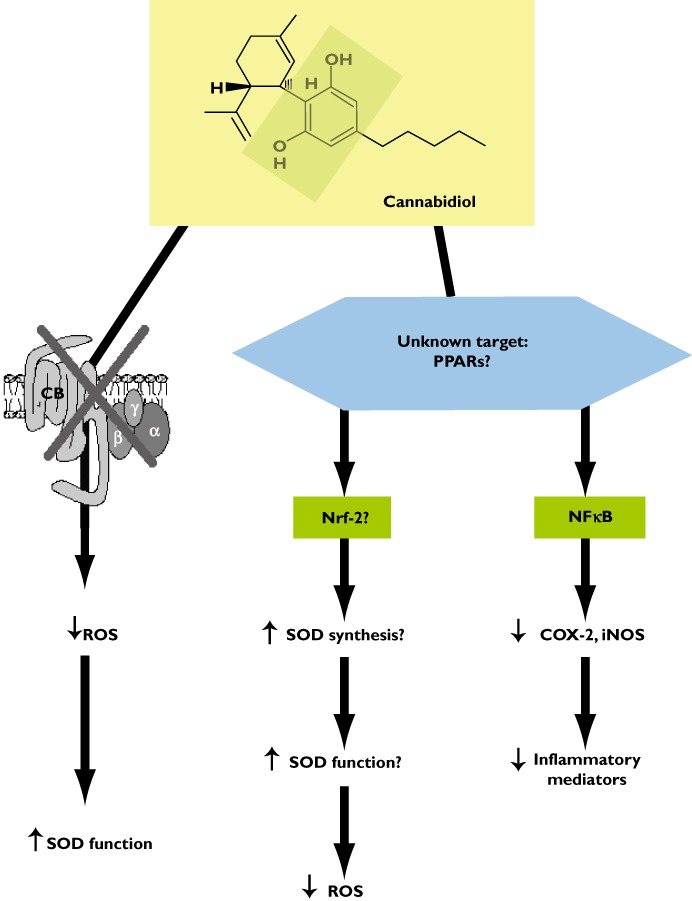
Mechanisms proposed for the neuroprotective effects exerted by CBD in neurodegenerative disorders.
The second key mechanism for CBD as a neuroprotective compound involves its anti-inflammatory activity that is exerted by mechanisms other than the activation of CB2 receptors, the canonic pathway for the anti-inflammatory effects of most of cannabinoid agonists [70]. Anti-inflammatory effects of CBD have been related to the control of microglial cell migration [80] and the toxicity exerted by these cells, i.e. production of pro-inflammatory mediators [81], similarly with the case of cannabinoid compounds targeting the CB2 receptor [70]. However, a key element in this CBD effect is the inhibitory control of NFκB signalling activity and the control of those genes regulated by this transcription factor (i.e. iNOS) [31, 81]. This inhibitory control of NFκB signalling may be exerted by reducing the phosphorylation of specific kinases (i.e. p38 MAP kinase) involved in the control of this transcription factor and by preventing its translocation to the nucleus to induce the expression of pro-inflammatory genes [31]. However, it has been recently proposed that CBD may bind the nuclear receptors of the PPAR family, in particular the PPAR-γ[38, 39] (Table 1) and it is well known that these receptors antagonize the action of NFκB, reducing the expression of pro-inflammatory enzymes (i.e. iNOS, COX-2), pro-inflammatory cytokines and metalloproteases, effects that are elicited by different cannabinoids including CBD (reviewed in [9, 39]). Therefore, it could well be that CBD may produce its anti-inflammatory effects by the activation of these nuclear receptors and the regulation of their downstream signals although various aspects of this mechanism are pending further research and confirmation (see proposed mechanism in Figure 1).
Other mechanisms proposed for the neuroprotective effects of CBD include: (i) the contribution of 5HT1A receptors, e.g. in stroke [27, 28], (ii) the inhibition of adenosine uptake [37], e.g. in neonatal ischaemia ([20], see below) and (iii) specific signalling pathways (e.g. WNT/β-catenin signaling) that play a role in β-amyloid-induced GSK-3β activation and tau hyperphosphorylation in Alzheimer's disease [82].
CBD in specific neurodegenerative disorders: from basic to clinical studies
Although the neuroprotective properties of CBD have been already examined in numerous acute or chronic neurodegenerative disorders, we will address here only three disorders, i.e. neonatal ischaemia, HD and PD, in which a clinical evaluation of CBD, as monotherapy or in combination with other phytocannabinoids, is already in progress or may be developed soon. CBD has demonstrated significant effects in preclinical models of these three disorders, but, in some cases, its combination with other phytocannabinoids (i.e. Δ9-THC for HD, Δ9-THCV for PD) revealed some interesting synergies that may be extremely useful at the clinical level.
CBD and neonatal ischaemia
Brain damage by hypoxia-ischaemia (HI) affects 0.3% subjects over 65 years old in developed countries leading to more than 150 000 deaths per year in the USA (for review see [83]). Although less prevalent, newborn hypoxic-ischaemic brain damage (NHIBD) is of great importance too. Approximately 0.1–0.2% live term births experience perinatal asphyxia with one third of them developing a severe neurological syndrome. About 25% of severe NHIBD leads to lasting sequelae and about 20% to death. Energy failure during ischaemia provokes the dysfunction of ionic pumps in neurons, leading to accumulation of ions and excitotoxic substances such as glutamate. The consequent increase in intracellular calcium content aggravates the neuron dysfunction and activates different enzymes, starting different processes of immediate and programmed cell death. During post ischaemic reperfusion, inflammation and oxidative stress aggravate and amplify such responses, increasing and spreading neuron and glial cell damage. Excitotoxicity, inflammation and oxidative stress play, therefore, a particularly relevant role in HI-induced brain cell death in newborns [83].
Unfortunately, the therapeutic outcome in NHIBD is still very limited and there is a strong need for novel strategies. We have solid evidence that CBD may be a good candidate to be tested in NHIBD at the clinical level. Using forebrain slices from newborn mice subjected to glucose-oxygen deprivation, a well-known in vitro model of NHIBD, we have already reported that CBD is able to reduce necrotic and apoptotic damage [20]. This neuroprotective effect is related to the modulation of excitotoxicity, oxidative stress and inflammation, as CBD normalizes the release of glutamate and cytokines as well as the induction of iNOS and COX-2 [20]. Surprisingly, we found that co-incubation of CBD with the CB2 receptor antagonist AM-630 abolished all these protective effects, suggesting that CB2 receptors are somehow involved in neuroprotective effects of CBD in immature brain [20]. In addition, adenosine receptors, in particular A2A receptors, seem to be also involved in these neuroprotective effects of CBD in the immature brain as revealed by the fact that the effect of CBD in this model was abolished by co-incubation with the A2A receptor antagonist SCH58261 [20]. CBD has been tested further in an in vivo model of NHIBD in newborn pigs, which closely resembles the actual human condition. In this model, the administration of CBD after the HI insult also reduces immediate brain damage by modulating cerebral haemodynamic impairment and brain metabolic derangement, and preventing the appearance of brain oedema and seizures. These neuroprotective effects are not only free from side effects but also associated with some beneficial cardiac, haemodynamic and ventilatory effects [84]. These protective effects restore neurobehavioural performance in the following 72 h post HI [85].
CBD and Huntington's disease
HD is an inherited neurodegenerative disorder caused by a mutation in the gene encoding the protein huntingtin. The mutation consists of a CAG triplet repeat expansion translated into an abnormal polyglutamine tract in the amino-terminal portion of huntingtin, which due to a gain of function becomes toxic for specific striatal and cortical neuronal subpopulations, although a loss of function in mutant huntingtin has been also related to HD pathogenesis (see [86] for review). Major symptoms include hyperkinesia (chorea) and cognitive deficits (see [87] for review). At present, there is no specific pharmacotherapy to alleviate motor and cognitive symptoms and/or to arrest/delay disease progression in HD. Thus, even though a few compounds have produced encouraging effects in preclinical studies (i.e. minocycline, coenzyme Q10, unsaturated fatty acids, inhibitors of histone deacetylases) none of the findings obtained in these studies have yet led on to the development of an effective medicine [88]. Importantly, therefore, following on from an extensive preclinical evaluation using different experimental models of HD, clinical tests are now being performed with cannabinoids, and this includes the use of CBD combined with Δ9-THC [26]. To get here, CBD was first studied in rats lesioned with 3-nitropropionic acid, a mitochondrial toxin that replicates the complex II deficiency characteristic of HD patients and that provokes striatal injury by mechanisms that mainly involve the Ca++-regulated protein calpain and generation of ROS. Neuroprotective effects in this experimental model were found with CBD alone [21] or combined with Δ9-THC as in Sativex®[22], and in both cases, these effects were not blocked by selective antagonists of either CB1 or CB2 receptors, thus supporting the idea that these effects are caused by the anti-oxidant and cannabinoid receptor-independent properties of these phytocannabinoids. It is possible, however, that this anti-oxidant/neuroprotective effect of phytocannabinoids involves the activation of signalling pathways implicated in the control of redox balance (i.e. nrf-2/ARE), as mentioned before. CBD has also been studied in rats lesioned with malonate, a model of striatal atrophy that involves mainly glial activation, inflammatory events and activation of apoptotic machinery. CBD alone did not provide protection in this model as only CB2 receptor agonists were effective [89], but the combination of CBD with Δ9-THC used in Sativex® was highly effective in this model, by preserving striatal neurons, and this protective effect involved both CB1 and CB2 receptors [23]. It is interesting to note that Δ9-THC alone produced biphasic effects in this model whereas CB1 receptor blockade aggravated the striatal damage [90]. We are presently studying the efficacy of this phytocannabinoid combination in a transgenic murine model of HD, i.e. R6/2 mice, in which the activation of both CB1 and CB2 receptors has already been found to induce beneficial effects [91, 92]. This solid preclinical evidence has provided substantial support for the evaluation of Sativex®, or equivalent cannabinoid-based medicines, as a new disease-modifying therapy in HD patients. Previous clinical studies had already used CBD, but they concentrated on symptom relief (i.e. chorea) rather than on disease progression and they did not show any significant improvement [93, 94]. We are presently engaged in a novel phase II-clinical trial with Sativex® as a disease-modifying agent in presymptomatic and early symptomatic patients [26], the outcome of which will be known soon.
CBD and Parkinson's disease
PD is also a progressive neurodegenerative disorder whose aetiology has been, however, associated with environmental insults, genetic susceptibility or interactions between both causes [95]. The major clinical symptoms in PD are tremor, bradykinesia, postural instability and rigidity, symptoms that result from the severe dopaminergic denervation of the striatum caused by the progressive death of dopaminergic neurons of the substantia nigra pars compacta[96]. CBD has also been found to be highly effective as a neuroprotective compound in experimental models of parkinsonism, i.e. 6-hydroxydopamine-lesioned rats, by acting through anti-oxidant mechanisms that seem to be independent of CB1 or CB2 receptors [24, 25, 97]. This observation is particularly important in the case of PD due to the relevance of oxidative injury to this disease, and because the hypokinetic profile of cannabinoids that activate CB1 receptors represents a disadvantage for this disease because such compounds can acutely enhance rather than reduce motor disability, as a few clinical data have already revealed (reviewed in [98]). Therefore, major efforts are being directed at finding cannabinoid molecules that may provide neuroprotection through their anti-oxidant properties and that may also activate CB2 receptors, but not CB1 receptors, or that may even block CB1 receptors, actions which may provide additional benefits, for example by relieving symptoms such as bradykinesia. One interesting example of a compound with this profile is the phytocannabinoid Δ9-THCV, which is presently under investigation in preclinical models of PD [25]. Thus, there could well be clinical advantages to administering Δ9-THCV together with CBD as this might induce symptomatic relief (due to the blockade of CB1 by Δ9-THCV) and neuroprotection (due to the anti-oxidant and anti-inflammatory properties of both CBD and Δ9-THCV). The combination of CBD with Δ9-THCV (rather than with Δ9-THC) would merit investigation in parkinsonian patients (reviewed in [9, 99]), as previous data obtained in clinical studies have indicated that CBD was effective in the relief of some PD-related symptoms such as dystonia, although not in others like tremor [100], but its combination with Δ9-THC, which can activate CB1 receptors, failed to improve parkinsoniam symptoms or to attenuate levodopa-induced dyskinesias [101].
Concluding remarks and futures perspectives
The experimental evidence presented in this review supports the idea that, from a pharmaceutical point of view, CBD is an unusually interesting molecule. As presented above, its actions are channeled through several biochemical mechanisms and yet it causes essentially no undesirable side effects and its toxicity is negligible [2]. It has shown valuable activities in numerous pharmaceutically important areas: (i) it is a potent anti-oxidant [73], which may partly explain its neuroprotective effects in PD [24, 25], and possibly in cerebral ischaemia-reperfusion (reviewed in [83]), (ii) it has been evaluated in human epileptic patients with very positive results [7–9], (iii) it has shown activity in mice with several autoimmune diseases, i.e. type-1 diabetes [102] and rheumatoid arthritis [103], (iv) it lowers the effects of myocardial ischaemic-reperfusion injury in mice [104], (v) it reduces microglial activation in mice and hence may slow the progression of Alzheimer's disease [78], (vi) it protects against hepatic ischaemia/reperfusion injury in animals [105] and has shown considerable activity in an animal model of hepatic encephalopathy [106], (vii) it even lowers anxiety (in humans) [107] and (viii) it is already in use, together with Δ9-THC, in a buccal spray (Sativex®) to lower symptoms of multiple sclerosis [6]. The presence of CBD in Sativex® enhances the positive effects of Δ9-THC whilst reducing its adverse effects, in concordance with previous data that indicated that CBD alters some of the effects of Δ9-THC, i.e. it lowers the acute memory-impairing effects and anxiety produced by Δ9-THC [108]. In addition, cannabis with high CBD content presumably leads to fewer psychotic experiences than cannabis with a highest proportion of Δ9-THC [17].
It is possible that CBD has not become a licensed medicine (except in Sativex®) because of patenting problems. However, commercial issues apart, CBD has tremendous potential as a new medicine. Thus, because the mechanisms that underlie its anti-inflammatory effects are different from those of prescribed drugs, it could well prove to be of considerable benefit to a large number of patients, who for various reasons are not sufficiently helped by existing drugs. In type 1-diabetes, we have shown that in mice CBD very significantly lowers the number of insulin-producing cells that are affected even after the disease has advanced [102]. Its neuroprotective effects are extremely valuable as no drugs exist that have similar properties. Surprisingly very few CBD derivatives have been evaluated and compared with CBD. At least one of them, CBD-dimethylheptyl-7-oic acid, is more potent than CBD as an anti-inflammatory agent [109]. Aren't we missing a valuable new pathway to a family of very promising new therapeutic agents?
Acknowledgments
The experimental work carried out by our group and that has been mentioned in this review article, has been supported during the last years by grants from CIBERNED (CB06/05/0089), MICINN (SAF2009-11847), CAM (S2011/BMD-2308) and GW Pharmaceuticals Ltd. The authors are indebted to all colleagues who contributed in this experimental work and to Yolanda García-Movellán for administrative support.
Competing Interests
JFR, OS and CG are supported by GW Pharma for research on phytocannabinoids and motor disorders. JMO and MRP have received funds for research from GW Pharma, Ltd. RP's research is supported in part by funding from GW Pharmaceuticals. RM is a consultant of GW Pharma.
REFERENCES
- 1.Grlic L. A comparative study on some chemical and biological characteristics of various samples of cannabis resin. Bull Narc. 1976;14:37–46. [Google Scholar]
- 2.Mechoulam R, Parker LA, Gallily R. Cannabidiol: an overview of some pharmacological aspects. J Clin Pharmacol. 2002;42:11S–9S. doi: 10.1002/j.1552-4604.2002.tb05998.x. [DOI] [PubMed] [Google Scholar]
- 3.Mechoulam R, Peters M, Murillo-Rodriguez E, Hanus LO. Cannabidiol-recent advances. Chem Biodivers. 2007;4:1678–92. doi: 10.1002/cbdv.200790147. [DOI] [PubMed] [Google Scholar]
- 4.Russo E, Guy GW. A tale of two cannabinoids: the therapeutic rationale for combining tetrahydrocannabinol and cannabidiol. Med Hypotheses. 2006;66:234–46. doi: 10.1016/j.mehy.2005.08.026. [DOI] [PubMed] [Google Scholar]
- 5.Costa B, Colleoni M, Conti S, Parolaro D, Franke C, Trovato AE, Giagnoni G. Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn Schmiedebergs Arch Pharmacol. 2004;369:294–9. doi: 10.1007/s00210-004-0871-3. [DOI] [PubMed] [Google Scholar]
- 6.Sastre-Garriga J, Vila C, Clissold S, Montalban X. THC and CBD oromucosal spray (Sativex®) in the management of spasticity associated with multiple sclerosis. Expert Rev Neurother. 2011;11:627–37. doi: 10.1586/ern.11.47. [DOI] [PubMed] [Google Scholar]
- 7.Cunha JM, Carlini EA, Pereira AE, Ramos OL, Pimentel C, Gagliardi R, Sanvito WL, Lander N, Mechoulam R. Chronic administration of cannabidiol to healthy volunteers and epileptic patients. Pharmacology. 1980;21:175–85. doi: 10.1159/000137430. [DOI] [PubMed] [Google Scholar]
- 8.Cortesi M, Fusar-Poli P. Potential therapeutical effects of cannabidiol in children with pharmacoresistant epilepsy. Med Hypotheses. 2007;68:920–1. doi: 10.1016/j.mehy.2006.09.030. [DOI] [PubMed] [Google Scholar]
- 9.Hill AJ, Williams CM, Whalley BJ, Stephens GJ. Phytocannabinoids as novel therapeutic agents in CNS disorders. Pharmacol Ther. 2012;133:79–97. doi: 10.1016/j.pharmthera.2011.09.002. [DOI] [PubMed] [Google Scholar]
- 10.Murillo-Rodríguez E, Millán-Aldaco D, Palomero-Rivero M, Mechoulam R, Drucker-Colín R. Cannabidiol, a constituent of Cannabis sativa, modulates sleep in rats. FEBS Lett. 2006;580:4337–45. doi: 10.1016/j.febslet.2006.04.102. [DOI] [PubMed] [Google Scholar]
- 11.Parker LA, Rock EM, Limebeer CL. Regulation of nausea and vomiting by cannabinoids. Br J Pharmacol. 2011;163:1411–22. doi: 10.1111/j.1476-5381.2010.01176.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ligresti A, Moriello AS, Starowicz K, Matias I, Pisanti S, De Petrocellis L, Laezza C, Portella G, Bifulco M, Di Marzo V. Antitumor activity of plant cannabinoids with emphasis on the effect of cannabidiol on human breast carcinoma. J Pharmacol Exp Ther. 2006;318:1375–87. doi: 10.1124/jpet.106.105247. [DOI] [PubMed] [Google Scholar]
- 13.Massi P, Vaccani A, Bianchessi S, Costa B, Macchi P, Parolaro D. The non-psychoactive cannabidiol triggers caspase activation and oxidative stress in human glioma cells. Cell Mol Life Sci. 2006;63:2057–66. doi: 10.1007/s00018-006-6156-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Leweke FM, Schneider U, Radwan M, Schmidt E, Emrich HM. Different effects of nabilone and cannabidiol on binocular depth inversion in man. Pharmacol Biochem Behav. 2000;66:175–81. doi: 10.1016/s0091-3057(00)00201-x. [DOI] [PubMed] [Google Scholar]
- 15.Zuardi AW, Crippa JA, Hallak JE, Moreira FA, Guimarães FS. Cannabidiol, a Cannabis sativa constituent, as an antipsychotic drug. Braz J Med Biol Res. 2006;39:421–9. doi: 10.1590/s0100-879x2006000400001. [DOI] [PubMed] [Google Scholar]
- 16.Leweke FM, Piomelli D, Pahlisch F, Muhi D, Gerth CW, Hoyer C, Klosterkötter J, Hellmich M, Koethe D. Cannabidiol enhances anandamide signaling and alleviates psychotic symptoms of schizophrenia. Transl Psychiatry. 2012;2:e94. doi: 10.1038/tp.2012.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Schubart CD, Sommer IE, van Gastel WA, Goetgebuer RL, Kahn RS, Boks MP. Cannabis with high cannabidiol content is associated with fewer psychotic experiences. Schizophr Res. 2011;130:216–21. doi: 10.1016/j.schres.2011.04.017. [DOI] [PubMed] [Google Scholar]
- 18.Gomes FV, Resstel LBM, Guimarães FS. The anxiolytic-like effects of cannabidiol injected into the bed nucleus of the stria terminalis are mediated by 5-HT1A receptors. Psychopharmacology. 2011;213:465–73. doi: 10.1007/s00213-010-2036-z. [DOI] [PubMed] [Google Scholar]
- 19.Zanelati TV, Biojone C, Moreira FA, Guimarães FS, Joca SRL. Antidepressant-like effects of cannabidiol in mice: possible involvement of 5-HT1A receptors. Br J Pharmacol. 2010;159:122–8. doi: 10.1111/j.1476-5381.2009.00521.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Castillo A, Tolón MR, Fernández-Ruiz J, Romero J, Martinez-Orgado J. The neuroprotective effect of cannabidiol in an in vitro model of newborn hypoxic-ischemic brain damage in mice is mediated by CB2 and adenosine receptors. Neurobiol Dis. 2010;37:434–40. doi: 10.1016/j.nbd.2009.10.023. [DOI] [PubMed] [Google Scholar]
- 21.Sagredo O, Ramos JA, Decio A, Mechoulam R, Fernández-Ruiz J. Cannabidiol reduced the striatal atrophy caused 3-nitropropionic acid in vivo by mechanisms independent of the activation of cannabinoid, vanilloid TRPV1 and adenosine A2A receptors. Eur J Neurosci. 2007;26:843–51. doi: 10.1111/j.1460-9568.2007.05717.x. [DOI] [PubMed] [Google Scholar]
- 22.Sagredo O, Pazos MR, Satta V, Ramos JA, Pertwee RG, Fernández-Ruiz J. Neuroprotective effects of phytocannabinoid-based medicines in experimental models of Huntington's disease. J Neurosci Res. 2011;89:1509–18. doi: 10.1002/jnr.22682. [DOI] [PubMed] [Google Scholar]
- 23.Valdeolivas S, Satta V, Pertwee RG, Fernández-Ruiz J, Sagredo O. Sativex-like combination of phytocannabinoids is neuroprotective in malonate-lesioned rats, an inflammatory model of Huntington's disease: role of CB1 and CB2 receptors. ACS Chem Neurosci. 2012;3:400–6. doi: 10.1021/cn200114w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lastres-Becker I, Molina-Holgado F, Ramos JA, Mechoulam R, Fernández-Ruiz J. Cannabinoids provide neuroprotection against 6-hydroxydopamine toxicity in vivo and in vitro: relevance to Parkinson's disease. Neurobiol Dis. 2005;19:96–107. doi: 10.1016/j.nbd.2004.11.009. [DOI] [PubMed] [Google Scholar]
- 25.García C, Palomo-Garo C, García-Arencibia M, Ramos J, Pertwee RG, Fernández-Ruiz J. Symptom-relieving and neuroprotective effects of the phytocannabinoid Δ9-THCV in animal models of Parkinson's disease. Br J Pharmacol. 2011;163:1495–506. doi: 10.1111/j.1476-5381.2011.01278.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.García de Yébenes J. Phase II-clinical trial on neuroprotection with cannabinoids in Huntington's disease (SAT-HD) EudraCT 2010-024227-24.
- 27.Hayakawa K, Mishima K, Nozako M, Ogata A, Hazekawa M, Liu A-X, Fujioka M, Abe K, Hasebe N, Egashira N, Iwasaki K, Fujiwara M. Repeated treatment with cannabidiol but not Δ9-tetrahydrocannabinol has a neuroprotective effect without the development of tolerance. Neuropharmacology. 2007;52:1079–87. doi: 10.1016/j.neuropharm.2006.11.005. [DOI] [PubMed] [Google Scholar]
- 28.Mishima K, Hayakawa K, Abe K, Ikeda T, Egashira N, Iwasaki K, Fujiwara M. Cannabidiol prevents cerebral infarction via a serotonergic 5-hydroxytryptamine1A receptor-dependent mechanism. Stroke. 2005;36:1071–6. doi: 10.1161/01.STR.0000163083.59201.34. [DOI] [PubMed] [Google Scholar]
- 29.Braida D, Pegorini S, Arcidiacono MV, Consalez GG, Croci L, Sala M. Post-ischemic treatment with cannabidiol prevents electroencephalographic flattening, hyperlocomotion and neuronal injury in gerbils. Neurosci Lett. 2003;346:61–4. doi: 10.1016/s0304-3940(03)00569-x. [DOI] [PubMed] [Google Scholar]
- 30.Hayakawa K, Mishima K, Fujiwara M. Therapeutic potential of non-psychotropic cannabidiol in ischemic stroke. Pharmaceuticals. 2010;3:2197–212. doi: 10.3390/ph3072197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Esposito G, De Filippis D, Maiuri MC, De Stefano D, Carnuccio R, Iuvone T. Cannabidiol inhibits inducible nitric oxide synthase protein expression and nitric oxide production in beta-amyloid stimulated PC12 neurons through p38 MAP kinase and NF-kappaB involvement. Neurosci Lett. 2006;99:91–5. doi: 10.1016/j.neulet.2006.01.047. [DOI] [PubMed] [Google Scholar]
- 32.Iuvone T, Esposito G, Esposito R, Santamaria R, Di Rosa M, Izzo AA. Neuroprotective effect of cannabidiol, a non-psychoactive component from Cannabis sativa, on beta-amyloid-induced toxicity in PC12 cells. J Neurochem. 2004;89:134–41. doi: 10.1111/j.1471-4159.2003.02327.x. [DOI] [PubMed] [Google Scholar]
- 33.Kozela E, Lev N, Kaushansky N, Eilam R, Rimmerman N, Levy R, Ben-Nun A, Juknat A, Vogel Z. Cannabidiol inhibits pathogenic T cells, decreases spinal microglial activation and ameliorates multiple sclerosis-like disease in C57BL/6 mice. Br J Pharmacol. 2011;163:1507–19. doi: 10.1111/j.1476-5381.2011.01379.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.de Filippis D, Iuvone T, d'amico A, Esposito G, Steardo L, Herman AG, Pelckmans PA, de Winter BY, de Man JG. Effect of cannabidiol on sepsis-induced motility disturbances in mice: involvement of CB receptors and fatty acid amide hydrolase. Neurogastroenterol Motil. 2008;20:919–27. doi: 10.1111/j.1365-2982.2008.01114.x. [DOI] [PubMed] [Google Scholar]
- 35.Bisogno T, Hanus L, De Petrocellis L, Tchilibon S, Ponde DE, Brandi I, Moriello AS, Davis JB, Mechoulam R, Di Marzo V. Molecular targets for cannabidiol and its synthetic analogues: effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br J Pharmacol. 2001;134:845–52. doi: 10.1038/sj.bjp.0704327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.De Petrocellis L, Ligresti A, Moriello AS, Allarà M, Bisogno T, Petrosino S, Stott CG, Di Marzo V. Effects of cannabinoids and cannabinoid-enriched cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br J Pharmacol. 2011;163:1479–94. doi: 10.1111/j.1476-5381.2010.01166.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Carrier EJ, Auchampach JA, Hillard CJ. Inhibition of an equilibrative nucleoside transporter by cannabidiol: a mechanism of cannabinoid immunosuppression. Proc Natl Acad Sci USA. 2006;103:7895–900. doi: 10.1073/pnas.0511232103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.O'Sullivan SE, Kendall DA. Cannabinoid activation of peroxisome proliferator-activated receptors: potential for modulation of inflammatory disease. Immunobiology. 2010;215:611–6. doi: 10.1016/j.imbio.2009.09.007. [DOI] [PubMed] [Google Scholar]
- 39.Esposito G, Scuderi C, Valenza M, Togna GI, Latina V, De Filippis D, Cipriano M, Carratù MR, Iuvone T, Steardo L. Cannabidiol reduces Aβ-induced neuroinflammation and promotes hippocampal neurogenesis through PPARγ involvement. PLoS ONE. 2011;6:e28668. doi: 10.1371/journal.pone.0028668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Thomas A, Baillie GL, Phillips AM, Razdan RK, Ross RA, Pertwee RG. Cannabidiol displays unexpectedly high potency as an antagonist of CB1 and CB2 receptor agonists in vitro. Br J Pharmacol. 2007;150:613–23. doi: 10.1038/sj.bjp.0707133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Pertwee RG, Howlett AC, Abood ME, Alexander SPH, Di Marzo V, Elphick MR, Greasley PJ, Hansen HS, Kunos G, Mackie K, Mechoulam R, Ross RA. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: beyond CB1 and CB2. Pharmacol Rev. 2010;62:588–631. doi: 10.1124/pr.110.003004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yang K-H, Galadari S, Isaev D, Petroianu G, Shippenberg TS, Oz M. The nonpsychoactive cannabinoid cannabidiol inhibits 5-hydroxytryptamine3A receptor-mediated currents in Xenopus laevis oocytes. J Pharmacol Exp Ther. 2010;333:547–54. doi: 10.1124/jpet.109.162594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ross HR, Napier I, Connor M. Inhibition of recombinant human T-type calcium channels by Δ9-tetrahydrocannabinol and cannabidiol. J Biol Chem. 2008;283:16124–34. doi: 10.1074/jbc.M707104200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Pertwee RG. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Δ9-tetrahydrocannabinol, cannabidiol and Δ9-tetrahydrocannabivarin. Br J Pharmacol. 2008;153:199–215. doi: 10.1038/sj.bjp.0707442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ahrens J, Demir R, Leuwer M, de la Roche J, Krampfl K, Foadi N, Karst M, Haeseler G. The nonpsychotropic cannabinoid cannabidiol modulates and directly activates alpha-1 and alpha-1-beta glycine receptor function. Pharmacology. 2009;83:217–22. doi: 10.1159/000201556. [DOI] [PubMed] [Google Scholar]
- 46.Yamaori S, Kushihara M, Yamamoto I, Watanabe K. Characterization of major phytocannabinoids, cannabidiol and cannabinol, as isoform-selective and potent inhibitors of human CYP1 enzymes. Biochem Pharmacol. 2010;79:1691–8. doi: 10.1016/j.bcp.2010.01.028. [DOI] [PubMed] [Google Scholar]
- 47.Yamaori S, Maeda C, Yamamoto I, Watanabe K. Differential inhibition of human cytochrome P450 2A6 and 2B6 by major phytocannabinoids. Forensic Toxicol. 2011;29:117–24. [Google Scholar]
- 48.Yamaori S, Okamoto Y, Yamamoto I, Watanabe K. Cannabidiol, a major phytocannabinoid, as a potent atypical inhibitor for CYP2D6. Drug Metab Dispos. 2011;39:2049–56. doi: 10.1124/dmd.111.041384. [DOI] [PubMed] [Google Scholar]
- 49.Yamaori S, Ebisawa J, Okushima Y, Yamamoto I, Watanabe K. Potent inhibition of human cytochrome P450 3A isoforms by cannabidiol: role of phenolic hydroxyl groups in the resorcinol moiety. Life Sci. 2011;88:730–6. doi: 10.1016/j.lfs.2011.02.017. [DOI] [PubMed] [Google Scholar]
- 50.Koch M, Dehghani F, Habazettl I, Schomerus C, Korf H-W. Cannabinoids attenuate norepinephrine-induced melatonin biosynthesis in the rat pineal gland by reducing arylalkylamine N-acetyltransferase activity without involvement of cannabinoid receptors. J Neurochem. 2006;98:267–78. doi: 10.1111/j.1471-4159.2006.03873.x. [DOI] [PubMed] [Google Scholar]
- 51.Jenny M, Santer E, Pirich E, Schennach H, Fuchs D. Δ9-tetrahydrocannabinol and cannabidiol modulate mitogen-induced tryptophan degradation and neopterin formation in peripheral blood mononuclear cells in vitro. J Neuroimmunol. 2009;207:75–82. doi: 10.1016/j.jneuroim.2008.12.004. [DOI] [PubMed] [Google Scholar]
- 52.Takeda S, Usami N, Yamamoto I, Watanabe K. Cannabidiol-2′,6′-dimethyl ether, a cannabidiol derivative, is a highly potent and selective 15-lipoxygenase inhibitor. Drug Metab Dispos. 2009;37:1733–7. doi: 10.1124/dmd.109.026930. [DOI] [PubMed] [Google Scholar]
- 53.Usami N, Yamamoto I, Watanabe K. Generation of reactive oxygen species during mouse hepatic microsomal metabolism of cannabidiol and cannabidiol hydroxy-quinone. Life Sci. 2008;83:717–24. doi: 10.1016/j.lfs.2008.09.011. [DOI] [PubMed] [Google Scholar]
- 54.Watanabe K, Motoya E, Matsuzawa N, Funahashi T, Kimura T, Matsunaga T, Arizono K, Yamamoto I. Marijuana extracts possess the effects like the endocrine disrupting chemicals. Toxicology. 2005;206:471–8. doi: 10.1016/j.tox.2004.08.005. [DOI] [PubMed] [Google Scholar]
- 55.Funahashi T, Ikeuchi H, Yamaori S, Kimura T, Yamamoto I, Watanabe K. In vitro inhibitory effects of cannabinoids on progesterone 17α-hydroxylase activity in rat testis microsomes. J Health Sci. 2005;51:369–75. [Google Scholar]
- 56.Zhu HJ, Wang J-S, Markowitz JS, Donovan JL, Gibson BB, Gefroh HA, DeVane CL. Characterization of P-glycoprotein inhibition by major cannabinoids from marijuana. J Pharmacol Exp Ther. 2006;317:850–7. doi: 10.1124/jpet.105.098541. [DOI] [PubMed] [Google Scholar]
- 57.Sreevalsan S, Joseph S, Jutooru I, Chadalapaka G, Safe SH. Induction of apoptosis by cannabinoids in prostate and colon cancer cells is phosphatase dependent. Anticancer Res. 2011;31:3799–807. [PMC free article] [PubMed] [Google Scholar]
- 58.Rock EM, Bolognini D, Limebeer CL, Cascio MG, Anavi-Goffer S, Fletcher PJ, Mechoulam R, Pertwee RG, Parker LA. Cannabidiol, a non-psychotropic component of cannabis, attenuates vomiting and nausea-like behaviour via indirect agonism of 5-HT1A somatodendritic autoreceptors in the dorsal raphe nucleus. Br J Pharmacol. 2012;165:2620–34. doi: 10.1111/j.1476-5381.2011.01621.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Dupre KB, Eskow KL, Barnum CJ, Bishop C. Striatal 5-HT1A receptor stimulation reduces D1 receptor-induced dyskinesia and improves movement in the hemiparkinsonian rat. Neuropharmacology. 2008;55:1321–8. doi: 10.1016/j.neuropharm.2008.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Ohno Y. Therapeutic role of 5-HT1A receptors in the treatment of schizophrenia and Parkinson's disease. CNS Neurosci Ther. 2011;17:58–65. doi: 10.1111/j.1755-5949.2010.00211.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Magen I, Avraham Y, Ackerman Z, Vorobiev L, Mechoulam R, Berry EM. Cannabidiol ameliorates cognitive and motor impairments in bile-duct ligated mice via 5-HT1A receptor activation. Br J Pharmacol. 2010;159:950–7. doi: 10.1111/j.1476-5381.2009.00589.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Campos AC, Guimarães FS. Involvement of 5HT1A receptors in the anxiolytic-like effects of cannabidiol injected into the dorsolateral periaqueductal gray of rats. Psychopharmacology. 2008;199:223–30. doi: 10.1007/s00213-008-1168-x. [DOI] [PubMed] [Google Scholar]
- 63.Resstel LBM, Tavares RF, Lisboa SFS, Joca SRL, Corrêa FMA, Guimarães FS. 5-HT1A receptors are involved in the cannabidiol-induced attenuation of behavioural and cardiovascular responses to acute restraint stress in rats. Br J Pharmacol. 2009;156:181–8. doi: 10.1111/j.1476-5381.2008.00046.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Soares VdeP, Campos AC, de Bortoli VC, Zangrossi H, Guimarães FS, Zuardi AW. Intra-dorsal periaqueductal gray administration of cannabidiol blocks panic-like response by activating 5-HT1A receptors. Behav Brain Res. 2010;213:225–9. doi: 10.1016/j.bbr.2010.05.004. [DOI] [PubMed] [Google Scholar]
- 65.Maione S, Piscitelli F, Gatta L, Vita D, De Petrocellis L, Palazzo E, de Novellis V, Di Marzo V. Non-psychoactive cannabinoids modulate the descending pathway of antinociception in anaesthetized rats through several mechanisms of action. Br J Pharmacol. 2011;162:584–96. doi: 10.1111/j.1476-5381.2010.01063.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Rock EM, Goodwin JM, Limebeer CL, Breuer A, Pertwee RG, Mechoulam R, Parker LA. Interaction between non-psychotropic cannabinoids in marihuana: effect of cannabigerol (CBG) on the anti-nausea or anti-emetic effects of cannabidiol (CBD) in rats and shrews. Psychopharmacology. 2011;215:505–12. doi: 10.1007/s00213-010-2157-4. [DOI] [PubMed] [Google Scholar]
- 67.Kwiatkowska M, Parker LA, Burton P, Mechoulam R. A comparative analysis of the potential of cannabinoids and ondansetron to suppress cisplatin-induced emesis in the Suncus murinus (house musk shrew) Psychopharmacology. 2004;174:254–9. doi: 10.1007/s00213-003-1739-9. [DOI] [PubMed] [Google Scholar]
- 68.Parker LA, Kwiatkowska M, Burton P, Mechoulam R. Effect of cannabinoids on lithium-induced vomiting in the Suncus murinus (house musk shrew) Psychopharmacology. 2004;171:156–61. doi: 10.1007/s00213-003-1571-2. [DOI] [PubMed] [Google Scholar]
- 69.Fernández-Ruiz J, García C, Sagredo O, Gómez-Ruiz M, de Lago E. The endocannabinoid system as a target for the treatment of neuronal damage. Expert Opin Ther Targets. 2010;14:387–404. doi: 10.1517/14728221003709792. [DOI] [PubMed] [Google Scholar]
- 70.Fernández-Ruiz J, Romero J, Velasco G, Tolón RM, Ramos JA, Guzmán M. Cannabinoid CB2 receptor: a new target for controlling neural cell survival? Trends Pharmacol Sci. 2007;28:39–45. doi: 10.1016/j.tips.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 71.Hampson AJ, Grimaldi M, Lolic M, Wink D, Rosenthal R, Axelrod J. Neuroprotective antioxidants from marijuana. Ann N Y Acad Sci. 2000;899:274–82. [PubMed] [Google Scholar]
- 72.El-Remessy AB, Khalil IE, Matragoon S, Abou-Mohamed G, Tsai NJ, Roon P, Caldwell RB, Caldwell RW, Green K, Liou GI. Neuroprotective effect of (-)Δ9-tetrahydrocannabinol and cannabidiol in N-methyl-D-aspartate-induced retinal neurotoxicity: involvement of peroxynitrite. Am J Pathol. 2003;163:1997–2008. doi: 10.1016/s0002-9440(10)63558-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hampson AJ, Grimaldi M, Axelrod J, Wink D. Cannabidiol and (-)Δ9-tetrahydrocannabinol are neuroprotective antioxidants. Proc Natl Acad Sci USA. 1998;95:8268–73. doi: 10.1073/pnas.95.14.8268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ruiz-Valdepeñas L, Martínez-Orgado JA, Benito C, Millán A, Tolón RM, Romero J. Cannabidiol reduces lipopolysaccharide-induced vascular changes and inflammation in the mouse brain: an intravital microscopy study. J Neuroinflammation. 2011;8:5. doi: 10.1186/1742-2094-8-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kwiatkoski M, Guimarães FS, Del-Bel E. Cannabidiol-treated rats exhibited higher motor score after cryogenic spinal cord injury. Neurotox Res. 2012;21:271–80. doi: 10.1007/s12640-011-9273-8. [DOI] [PubMed] [Google Scholar]
- 76.Iuvone T, Esposito G, De Filippis D, Scuderi C, Steardo L. Cannabidiol: a promising drug for neurodegenerative disorders? CNS Neurosci Ther. 2009;15:65–75. doi: 10.1111/j.1755-5949.2008.00065.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Marsicano G, Moosmann B, Hermann H, Lutz B, Behl C. Neuroprotective properties of cannabinoids against oxidative stress: role of the cannabinoid receptor CB1. J Neurochem. 2002;80:448–56. doi: 10.1046/j.0022-3042.2001.00716.x. [DOI] [PubMed] [Google Scholar]
- 78.Martín-Moreno AM, Reigada D, Ramírez BG, Mechoulam R, Innamorato N, Cuadrado A, de Ceballos ML. Cannabidiol and other cannabinoids reduce microglial activation in vitro and in vivo: relevance to Alzheimer's disease. Mol Pharmacol. 2011;79:964–73. doi: 10.1124/mol.111.071290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Juknat A, Pietr M, Kozela E, Rimmerman N, Levy R, Coppola G, Geschwind D, Vogel Z. Differential transcriptional profiles mediated by exposure to the cannabinoids cannabidiol and Δ(9)-tetrahydrocannabinol in BV-2 microglial cells. Br J Pharmacol. 2012 doi: 10.1111/j.1476-5381.2011.01461.x. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Walter L, Franklin A, Witting A, Wade C, Xie Y, Kunos G, Mackie K, Stella N. Nonpsychotropic cannabinoid receptors regulate microglial cell migration. J Neurosci. 2003;23:1398–405. doi: 10.1523/JNEUROSCI.23-04-01398.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Esposito G, Scuderi C, Savani C, Steardo L, Jr, De Filippis D, Cottone P, Iuvone T, Cuomo V, Steardo L. Cannabidiol in vivo blunts beta-amyloid induced neuroinflammation by suppressing IL-1beta and iNOS expression. Br J Pharmacol. 2007;151:1272–9. doi: 10.1038/sj.bjp.0707337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Esposito G, De Filippis D, Carnuccio R, Izzo AA, Iuvone T. The marijuana component cannabidiol inhibits beta-amyloid-induced tau protein hyperphosphorylation through Wnt/beta-catenin pathway rescue in PC12 cells. J Mol Med (Berl) 2006;84:253–8. doi: 10.1007/s00109-005-0025-1. [DOI] [PubMed] [Google Scholar]
- 83.Martinez-Orgado J, Fernandez-Lopez D, Lizasoain I, Romero J. The seek of neuroprotection: introducing cannabinoids. Recent Patents. CNS Drug Discov. 2007;2:131–9. doi: 10.2174/157488907780832724. [DOI] [PubMed] [Google Scholar]
- 84.Alvarez FJ, Lafuente H, Rey-Santano MC, Mielgo VE, Gastiasoro E, Rueda M, Pertwee RG, Castillo AI, Romero J, Martínez-Orgado J. Neuroprotective effects of the non-psychoactive cannabinoid cannabidiol in hypoxic-ischemic newborn piglets. Pediatr Res. 2008;64:653–8. doi: 10.1203/PDR.0b013e318186e5dd. [DOI] [PubMed] [Google Scholar]
- 85.Lafuente H, Alvarez FJ, Pazos MR, Alvarez A, Rey-Santano MC, Mielgo V, Murgia-Esteve X, Hilario E, Martinez-Orgado J. Cannabidiol reduces brain damage and improves functional recovery after acute hypoxia-ischemia in newborn pigs. Pediatr Res. 2011;70:272–7. doi: 10.1203/PDR.0b013e3182276b11. [DOI] [PubMed] [Google Scholar]
- 86.Zuccato C, Valenza M, Cattaneo E. Molecular mechanisms and potential therapeutical targets in Huntington's disease. Physiol Rev. 2010;90:905–81. doi: 10.1152/physrev.00041.2009. [DOI] [PubMed] [Google Scholar]
- 87.Roze E, Bonnet C, Betuing S, Caboche J. Huntington's disease. Adv Exp Med Biol. 2010;685:45–63. [PubMed] [Google Scholar]
- 88.Johnson CD, Davidson BL. Huntington's disease: progress toward effective disease-modifying treatments and a cure. Hum Mol Genet. 2010;19:R98–R102. doi: 10.1093/hmg/ddq148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Sagredo O, González S, Aroyo I, Pazos MR, Benito C, Lastres-Becker I, Romero JP, Tolón RM, Mechoulam R, Brouillet E, Romero J, Fernández-Ruiz J. Cannabinoid CB2 receptor agonists protect the striatum against malonate toxicity: relevance for Huntington's disease. Glia. 2009;57:1154–67. doi: 10.1002/glia.20838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Lastres-Becker I, Bizat N, Boyer F, Hantraye P, Brouillet E, Fernández-Ruiz J. Effects of cannabinoids in the rat model of Huntington's disease generated by an intrastriatal injection of malonate. Neuroreport. 2003;14:813–6. doi: 10.1097/00001756-200305060-00007. [DOI] [PubMed] [Google Scholar]
- 91.Palazuelos J, Aguado T, Pazos MR, Julien B, Carrasco C, Resel E, Sagredo O, Benito C, Romero J, Azcoitia I, Fernández-Ruiz J, Guzmán M, Galve-Roperh I. Microglial CB2 cannabinoid receptors are neuroprotective in Huntington's disease excitotoxicity. Brain. 2009;132:3152–64. doi: 10.1093/brain/awp239. [DOI] [PubMed] [Google Scholar]
- 92.Blázquez C, Chiarlone A, Sagredo O, Aguado T, Pazos MR, Resel E, Palazuelos J, Julien B, Salazar M, Börner C, Benito C, Carrasco C, Diez-Zaera M, Paoletti P, Díaz-Hernández M, Ruiz C, Sendtner M, Lucas JJ, de Yébenes JG, Marsicano G, Monory K, Lutz B, Romero J, Alberch J, Ginés S, Kraus J, Fernández-Ruiz J, Galve-Roperh I, Guzmán M. Loss of striatal type 1 cannabinoid receptors is a key pathogenic factor in Huntington's disease. Brain. 2011;134:119–36. doi: 10.1093/brain/awq278. [DOI] [PubMed] [Google Scholar]
- 93.Sandyk R, Snider SR, Consroe P, Elias SM. Cannabidiol in dystonic movement disorders. Psychiatry Res. 1986;18:291. doi: 10.1016/0165-1781(86)90117-4. [DOI] [PubMed] [Google Scholar]
- 94.Consroe P, Laguna J, Allender J, Snider S, Stern L, Sandyk R, Kennedy K, Schram K. Controlled clinical trial of cannabidiol in Huntington's disease. Pharmacol Biochem Behav. 1991;40:701–8. doi: 10.1016/0091-3057(91)90386-g. [DOI] [PubMed] [Google Scholar]
- 95.Thomas B, Beal MF. Parkinson's disease. Hum Mol Genet. 2007;16:R183–R194. doi: 10.1093/hmg/ddm159. [DOI] [PubMed] [Google Scholar]
- 96.Nagatsu T, Sawada M. Biochemistry of postmortem brains in Parkinson's disease: historical overview and future prospects. J Neural Transm Suppl. 2007;72:113–20. doi: 10.1007/978-3-211-73574-9_14. [DOI] [PubMed] [Google Scholar]
- 97.García-Arencibia M, González S, de Lago E, Ramos JA, Mechoulam R, Fernández-Ruiz J. Evaluation of the neuroprotective effect of cannabinoids in a rat model of Parkinson's disease: importance of antioxidant and cannabinoid receptor-independent properties. Brain Res. 2007;1134:162–70. doi: 10.1016/j.brainres.2006.11.063. [DOI] [PubMed] [Google Scholar]
- 98.Fernández-Ruiz J. The endocannabinoid system as a target for the treatment of motor dysfunction. Br J Pharmacol. 2009;156:1029–40. doi: 10.1111/j.1476-5381.2008.00088.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Fernández-Ruiz J, Moreno-Martet M, Rodríguez-Cueto C, Palomo-Garo C, Gómez-Cañas M, Valdeolivas S, Guaza C, Romero J, Guzmán M, Mechoulam R, Ramos JA. Prospects for cannabinoid therapies in basal ganglia disorders. Br J Pharmacol. 2011;163:1365–78. doi: 10.1111/j.1476-5381.2011.01365.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Consroe P, Sandyk R, Snider SR. Open label evaluation of cannabidiol in dystonic movement disorders. Int J Neurosci. 1986;30:277–82. doi: 10.3109/00207458608985678. [DOI] [PubMed] [Google Scholar]
- 101.Carroll CB, Bain PG, Teare L, Liu X, Joint C, Wroath C, Parkin SG, Fox P, Wright D, Hobart J, Zajicek JP. Cannabis for dyskinesia in Parkinson disease: a randomized double-blind crossover study. Neurology. 2004;63:1245–50. doi: 10.1212/01.wnl.0000140288.48796.8e. [DOI] [PubMed] [Google Scholar]
- 102.Weiss L, Zeira M, Reich S, Slavin S, Raz I, Mechoulam R, Gallily R. Cannabidiol arrests onset of autoimmune diabetes in NOD mice. Neuropharmacology. 2008;54:244–9. doi: 10.1016/j.neuropharm.2007.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Malfait AM, Gallily R, Sumariwalla PF, Malik AS, Andreakos E, Mechoulam R, Feldmann M. The nonpsychoactive cannabis constituent cannabidiol is an oral anti-arthritic therapeutic in murine collagen-induced arthritis. Proc Natl Acad Sci USA. 2000;97:9561–6. doi: 10.1073/pnas.160105897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Durst R, Danenberg H, Gallily R, Mechoulam R, Meir K, Grad E, Beeri R, Pugatsch T, Tarsish E, Lotan C. Cannabidiol, a nonpsychoactive cannabis constituent, protects against myocardial ischemic reperfusion injury. Am J Physiol Heart Circ Physiol. 2007;293:H3602–H3607. doi: 10.1152/ajpheart.00098.2007. [DOI] [PubMed] [Google Scholar]
- 105.Mukhopadhyay P, Rajesh M, Horváth B, Bátkai S, Park O, Tanchian G, Gao RY, Patel V, Wink DA, Liaudet L, Haskó G, Mechoulam R, Pacher P. Cannabidiol protects against hepatic ischemia/reperfusion injury by attenuating inflammatory signaling and response, oxidative/nitrative stress, and cell death. Free Radic Biol Med. 2011;50:1368–81. doi: 10.1016/j.freeradbiomed.2011.02.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Avraham Y, Grigoriadis N, Poutahidis T, Vorobiev L, Magen I, Ilan Y, Mechoulam R, Berry E. Cannabidiol improves brain and liver function in a fulminant hepatic failure-induced model of hepatic encephalopathy in mice. Br J Pharmacol. 2011;162:1650–8. doi: 10.1111/j.1476-5381.2010.01179.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Bergamaschi MM, Queiroz RH, Chagas MH, de Oliveira DC, De Martinis BS, Kapczinski F, Quevedo J, Roesler R, Schröder N, Nardi AE, Martín-Santos R, Hallak JE, Zuardi AW, Crippa JA. Cannabidiol reduces the anxiety induced by simulated public speaking in treatment-naïve social phobia patients. Neuropsychopharmacology. 2011;36:1219–26. doi: 10.1038/npp.2011.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Morgan CJ, Schafer G, Freeman TP, Curran HV. Impact of cannabidiol on the acute memory and psychotomimetic effects of smoked cannabis: naturalistic study. Br J Psychiatry. 2010;197:285–90. doi: 10.1192/bjp.bp.110.077503. [DOI] [PubMed] [Google Scholar]
- 109.Sumariwalla PF, Gallily R, Tchilibon S, Fride E, Mechoulam R, Feldmann M. A novel synthetic, nonpsychoactive cannabinoid acid (HU-320) with antiinflammatory properties in murine collagen-induced arthritis. Arthritis Rheum. 2004;50:985–98. doi: 10.1002/art.20050. [DOI] [PubMed] [Google Scholar]