

Abstract
OBJECTIVE
Ad36, a human adenovirus, increases adiposity but improves glycemic control in animal models. Similarly, natural Ad36 infection is cross-sectionally associated with greater adiposity and better glycemic control in humans. This study compared longitudinal observations in indices of adiposity (BMI and body fat percentage) and glycemic control (fasting glucose and insulin) in Ad36-infected versus uninfected adults.
RESEARCH DESIGN AND METHODS
Baseline sera from Hispanic men and women (n = 1,400) were screened post hoc for the presence of Ad36-specific antibodies. Indices of adiposity and glycemic control at baseline and at ∼10 years past the baseline were compared between seropositive and seronegative subjects, with adjustment for age and sex. In addition to age and sex, indices of glycemic control were adjusted for baseline BMI and were analyzed only for nondiabetic subjects.
RESULTS
Seropositive subjects (14.5%) had greater adiposity at baseline, compared with seronegative subjects. Longitudinally, seropositive subjects showed greater adiposity indices but lower fasting insulin levels. Subgroup analyses revealed that Ad36-seropositivity was associated with better baseline glycemic control and lower fasting insulin levels over time in the normal-weight group (BMI ≤25 kg/m2) and longitudinally, with greater adiposity in the overweight (BMI 25–30 kg/m2) and obese (BMI >30 kg/m2) men. Statistically, the differences between seropositive and seronegative individuals were modest in light of the multiple tests performed.
CONCLUSIONS
This study strengthens the plausibility that in humans, Ad36 increases adiposity and attenuates deterioration of glycemic control. Panoptically, the study raises the possibility that certain infections may modulate obesity or diabetes risk. A comprehensive understanding of these under-recognized factors is needed to effectively combat such metabolic disorders.
A deeper understanding of the factors that modulate adiposity or glycemic control may aid in the development of more effective treatment and prevention strategies for combating obesity or diabetes. Although the contribution of various behavioral and physiological risk factors for obesity or diabetes is extensively investigated, that of several putative factors remains under-investigated (1). Ad36, a human adenovirus, is one such putative factor originally described for its adipogenic property (2). Subsequent data indicated that Ad36 is likely to modulate both adiposity and glycemic control. In various animal models, experimental Ad36 infection significantly increases adiposity (3–7) and yet, seemingly paradoxically, improves glycemic control (6,7). In vitro and ex vivo mechanistic studies indicate that Ad36 expands adipose tissue by recruiting adipocyte progenitors and increasing their commitment, differentiation, and lipid accumulation (8–12). In agreement with these findings, fat cells of humans harboring Ad36 DNA in adipose tissue have greater adipogenic ability compared with fat cells of those without Ad36 DNA (10). In addition, Ad36 increases cellular glucose uptake in adipose tissue and skeletal (13,14), reduces hepatic lipid accumulation and glucose release from hepatocytes (7,15), and upregulates adiponectin, an insulin-sensitizing adipocytokine (7,16), which may collectively contribute to systemic glycemic control improved by Ad36 (6,7). An improvement in glycemic control via the expansion of adipose tissue has been recently documented in several animal models (17–19) and has been suggested in humans receiving some antidiabetic agents (20). Similarly, Ad36 may mediate its effect on glycemic control by increasing adiposity.
In humans, Ad36 infection is prevalent globally (21), with a prevalence of ∼15% in United States adults (7,22). Because of the substantial prevalence, if Ad36 influences adiposity or glycemic control in humans, then the potential impact could be considerable. However, ethical considerations preclude experimental infection of humans to determine a causal role of Ad36 in modulating adiposity or glycemic control. Several cross-sectional studies show that natural Ad36 infection in humans is associated with greater adiposity but better glycemic control. Association of Ad36 infection with greater BMI or greater prevalence of obesity is reported in adults from the United States (22), Italy (23), and Korea (24), and in children from the United States (25) and Korea (26,27). Ad36 infected human twins are significantly heavier compared with their uninfected counterparts (22). However, two studies did not find an association of Ad36 with obesity in military recruits or Scandinavian subjects (2). In four cohorts totaling >1,500 white, black, and Hispanic men, women, and children from the United States, Ad36 infection significantly predicted better indices of glycemic control that were independent of race, sex, age, and adiposity (7). Other smaller studies also showed significantly lower HbA1c levels and higher adiponectin in Ad36-infected adults (13,16). In addition, Ad36 infection is associated with lower hepatic lipid levels in United States and Italian adults and children (7,28).
Similar to the phenotype induced by Ad36 in experimentally infected animals, overall human cross-sectional studies show an association of Ad36 infection with greater adiposity, better glycemic control, and lower hepatic lipid levels. This congruence of data from animal and human studies supports a similar role for Ad36 in humans. However, causality cannot be established unequivocally because of the observational nature of these studies. A prospective approach could help elucidate the long-term association of Ad36 infection with adiposity and glycemic control within a person over time, and could markedly strengthen causal inferences by assessing temporality. Such determination may advance our understanding of multifactorial nature of obesity or diabetes, and may lead to novel prevention or treatment strategies.
This study screened baseline serum samples of Hispanic adults enrolled in San Antonio Family Heart Study for a history of natural Ad36 infection, as indicated by neutralizing antibodies. Data on obesity and glycemic control were collected for ∼10 years past baseline, which allowed the comparison of changes in indices of adiposity (BMI and body fat percentage) and glycemic control (fasting glucose and insulin) in Ad36-seropositive vs. Ad36-seronegative individuals.
RESEARCH DESIGN AND METHODS
Experimental design
Baseline sera samples of 1,400 Mexican American persons enrolled in the San Antonio Family Heart Study (29) were screened for the presence of neutralizing antibodies to Ad36 by serum neutralization assay as described. Subsequently, seropositivity was evaluated for association with measures of adiposity available at baseline and ∼10 years later.
San Antonio Family Heart Study
The study was initiated in 1991 with the goal of investigating the genetic factors influencing cardiovascular disease and related quantitative risk factors in Mexican American individuals from San Antonio, Texas. Multigenerational families were ascertained from largely Hispanic neighborhoods of San Antonio via a single adult Mexican American proband with a sufficient number of first-degree relatives to be eligible. Families were randomly ascertained without enrichment for specific diseases or other traits. All study participants gave informed consent. Body weight, body fat, fasting glucose, and insulin determined at baseline and at 10-year follow-up visit were used for analyses. Baseline body composition was determined by the bioimpedance method using a Valhalla Bio-impedance Analyzer (Valhalla Scientific, San Diego, CA). The 10-year follow-up body composition also was determined by a bioimpedance analyzer from Xitron Technologies (San Diego, CA). Plasma glucose was measured using an Abbott V/P Analyzer, and serum concentrations of insulin were measured using commercial radioimmunoassay kits (Diagnostic Products).
Serum neutralization assay
Presence of neutralizing antibodies in serum was determined by the “constant virus-decreasing serum” method as described (22). This is a labor-intensive assay but a highly specific and sensitive gold standard to determine neutralizing antibodies. Ad36 required for the assay was originally obtained from American Type Culture collection (Cat # VR913) and was plaque-purified and propagated in A549 cells (human lung cancer cell line) (3,4).
Statistical analyses
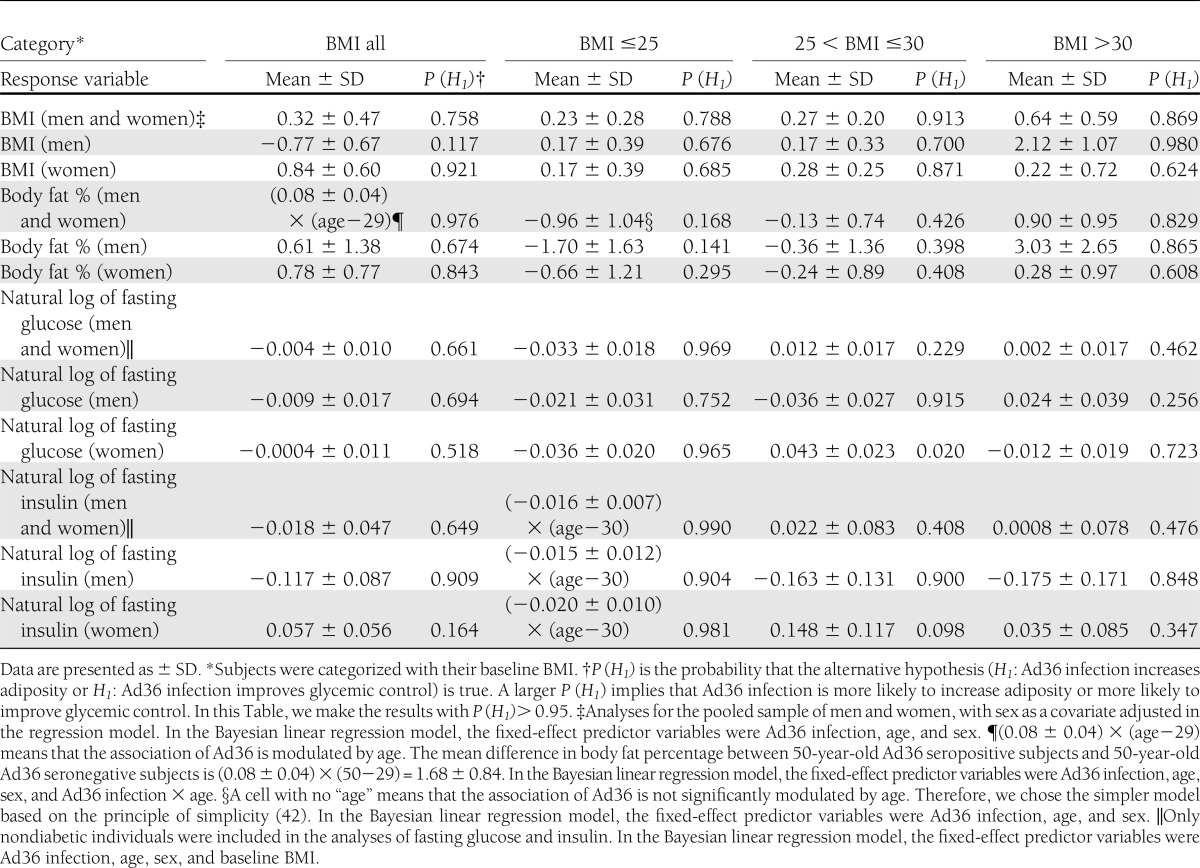
The statistical analyses for baseline measures were performed with the R package BLR (Bayesian linear regression) (30,31) to account for the degrees of genetic similarity between subjects within a pedigree and can adjust for covariates in the regression models (see Supplementary Materials for details). The data set comprises 153 pedigrees (with most participants belonging to large multigenerational pedigrees and a few participants being members of small pedigrees or singletons). BMI, body fat percentage, fasting glucose, and insulin were adjusted for age and sex. In addition, fasting glucose and insulin were also adjusted for BMI. Table 2 lists means and SDs of the differences in baseline indices of adiposity and glycemic control between Ad36 seropositive and seronegative subjects. In the table we also list the probability that the alternative hypothesis (H1: Ad36 infection increases adiposity or H1: Ad36 infection improves glycemic control) is true.
Table 2.
Differences in baseline indices of adiposity and glycemic control between Ad36 seropositive and seronegative subjects
The statistical analyses for longitudinal data were performed with the R package Pedigreemm (pedigree-based mixed-effect models) (32) (see Supplementary Materials for details). The measurements on adiposity and glycemic control at three time points (baseline, ∼5 years, and ∼10 years past the baseline) were jointly modeled in the mixed-effect model. The genetic similarity between subjects within a pedigree was considered by the Pedigreemm. Consistent with the previous analyses for baseline measures, BMI, body fat percentage, fasting glucose, and insulin were adjusted for age and sex. In addition, fasting glucose and insulin also were adjusted for BMI. P values were based on one-tailed tests according to our hypotheses about the association of Ad36 with obesity and glycemic control. Significance was considered at P < 0.05.
Furthermore, to gain better insight, we also examined the associations of Ad36 infection with BMI, body fat percentage, fasting glucose, and insulin for different subgroups of sex and BMI. Despite a continuous measure, BMI is commonly categorized as “normal weight,” “overweight,” or “obese” (BMI kg/m2: <25, 25–30, and >30, respectively). Glycemic control as indicated by the levels of fasting glucose or insulin is expected to deteriorate with age in the elderly or even the middle-aged (33,34). A lower increase in fasting glucose or insulin values based on the longitudinal observations over the 10-year period was used to indicate less deterioration of glycemic control.
RESULTS
Baseline
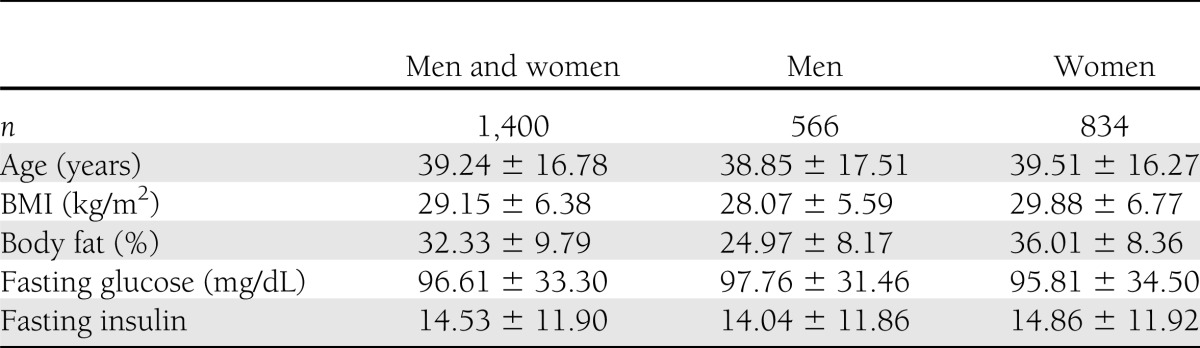
At baseline, men and women were almost 40 years of age on average and had a mean BMI in the overweight category (29.15 ± 6.38 kg/m2) (Table 1). Their mean fasting glucose and insulin levels were toward the upper limit of the normal range. Of the 834 women, and of the 566 men, 132 (15.8%) and 72 (12.7%), respectively, were seropositive for Ad36. To avoid any potential confounding effect of antidiabetic drugs on the variation in glucose and insulin levels, diabetic individuals (174 seronegative [14.5%] and 31 seropositive [15.2%]) were not included in the analyses of glycemic indices. A separate analysis for glycemic indices of diabetic and nondiabetic individuals was conducted (the diabetes status was regarded as a covariate adjusted in the model), which was presented in the Supplementary Materials.
Table 1.
Baseline characteristics (mean ± SD)
Compared with their seronegative counterparts, seropositive men and women were more likely to have greater body fat percentage (Table 2). BMI subgroup analyses indicated that Ad36 seropositivity was associated with better baseline glycemic control in the normal-weight group (BMI ≤25 kg/m2). When the BMI subgroups were separated by sex, seropositive normal-weight women, but not men, had lower baseline fasting glucose and insulin levels (Table 2).
Longitudinal data analyses for indices of adiposity and glycemic control
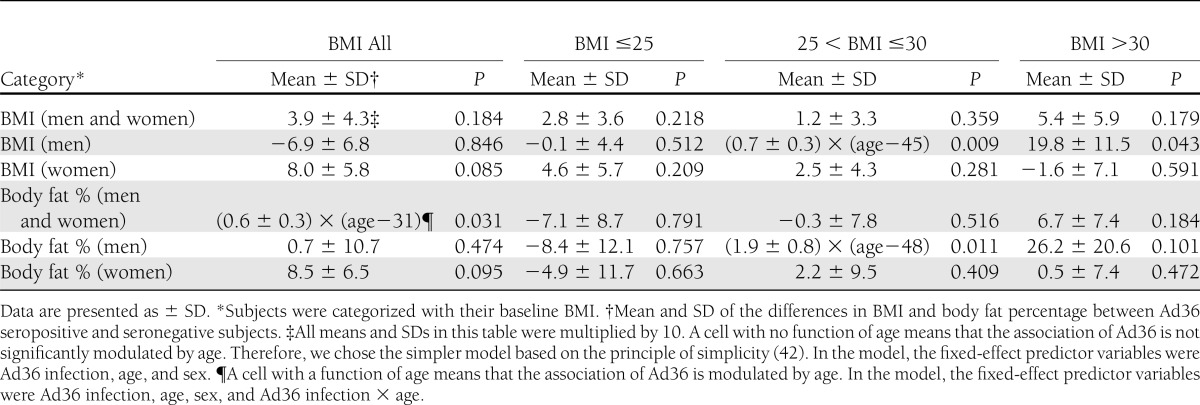
Seropositive men and women of all BMI, when analyzed together, showed a greater body fat percentage compared with seronegative counterparts (Table 3). From the BMI subgroup analyses, the overweight and obese seropositive men had a significant greater BMI compared with the overweight and obese seronegative men.
Table 3.
Longitudinal data analyses for BMI and body fat percentage
Men and women of all BMI, when analyzed together, showed lower fasting insulin levels for the seropositive group compared with the seronegative counterpart (Table 4). The BMI/sex subgroup analyses indicated that mainly the seropositive normal-weight men and women and the overweight men contributed to this association. Obese seropositive subjects did not show significantly different fasting glucose or insulin levels compared with their seronegative counterparts. Supplementary Table 1 summarizes the key findings presented in Tables 2–4.
Table 4.
Longitudinal data analyses for fasting glucose and insulin in nondiabetic subjects
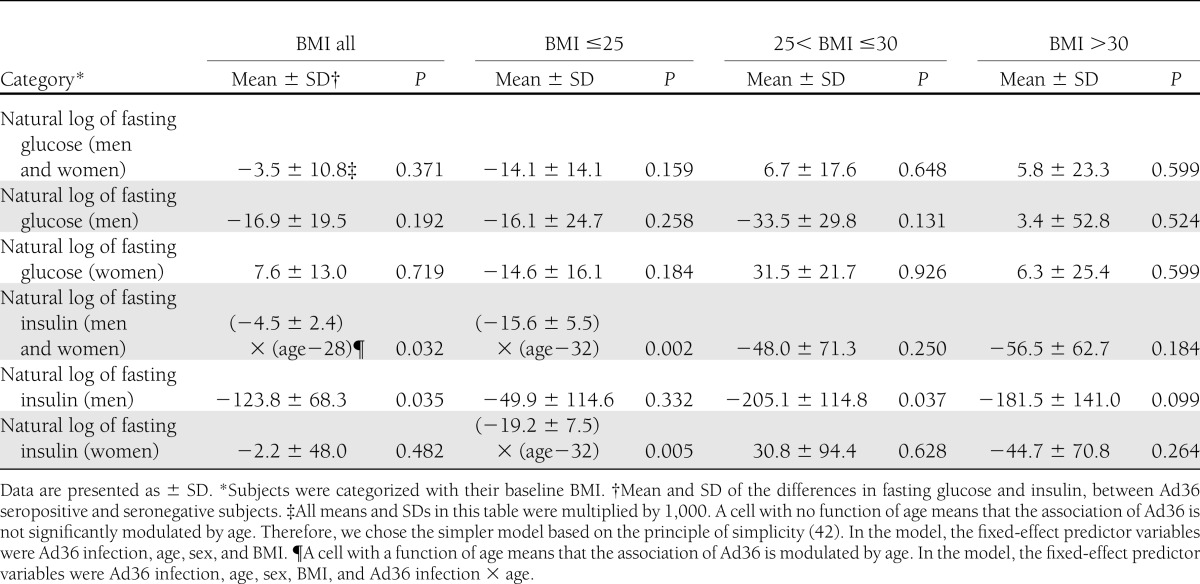
Note that only nondiabetic individuals were included in the analyses of fasting glucose and insulin. However, when the analyses were conducted by including diabetic and nondiabetic individuals, but adjusting for the diabetes status, the findings were similar (Supplementary Table 2). Ad36 seropositivity was associated with lower fasting glucose and insulin levels in only normal-weight men and women (Supplementary Table 2). Thus, despite diabetes status, Ad36 is associated with better glycemic profile, as we previously reported (13).
CONCLUSIONS
The results confirm a cross-sectional association of Ad36 infection with greater adiposity, as reported by several earlier studies (22–27). More importantly, this is the first report to show the longitudinal association of Ad36 infection with adiposity and glycemic control. The key finding of this study is that, as hypothesized, natural Ad36 infection is associated with greater adiposity and less deterioration in glycemic control in Hispanic men and women over time. Clearly, even a longitudinal association does not establish causality, and interpretation of these findings within the context of this study and in relation to other studies with Ad36 is required.
Overall, Ad36 seropositivity was associated with greater adiposity and attenuated deterioration in better glycemic control, both cross-sectionally and longitudinally (Supplementary Table 1). Interestingly, further analyses revealed that the association of Ad36 seropositivity with glycemic control and adiposity may be mainly contributed by two different subgroups, women with BMI ≤25 (for glycemic control) and men with BMI >25 (for adiposity), respectively.
In animal models, the dual effect of Ad36 on adiposity and glycemic control (6,7) appears similar to that of the thiazolidinedione class of antidiabetic drugs (20), which increase adiposity but improve diabetes. However, very recent vitro data indicate that unlike the action of the thiazolidinediones, the two effects of Ad36 may be separable (7,16). The dichotomy of association of Ad36 seropositivity with adiposity and glycemic control with two separate subgroups also suggests that the adipogenic and glycemic associations of Ad36 infection may not be interdependent.
When men and women were analyzed together, Ad36 seropositivity was associated with lower fasting insulin levels and greater body fat percentage over time. However, when analyzed separately, women appear to mainly contribute to the association with lower fasting insulin, whereas men appear to mainly contribute to the association with greater adiposity. Although the explanation for this observation is unknown, a hypothesis can be offered. Animal data indicate that adiponectin, an important insulin-sensitizing adipokine, may be a key player in mediating the insulin-sensitizing effect of Ad36 (7). Also, Ad36 infection is associated with greater adiponectin levels in humans (16). Adiponectin levels are higher in women compared with men (35), which may facilitate a greater effect of Ad36 on glycemic control in that sex. Conversely, adiponectin levels decrease in obesity or with a high-fat diet (36), which may explain a subdued association of Ad36 with better glycemic control in obese subjects.
Although it is unclear why some sex and BMI subgroups did not show significant associations with Ad36 infection, it could potentially indicate the contribution of unknown factors specific to these subgroups. Importantly, there is no information about other potential factors such as diet or exercise, which may influence adiposity or glycemic control in this population. How genetic and environmental factors can modulate the phenotypic effect of a virus is aptly illustrated by norovirus infection. This virus increases the susceptibility of an individual to Crohn disease only in the presence of a mutation in host gene Atg16L1 and in the presence of commensal bacteria (37).
Similarly, factors related to Ad36 infection, such as the virulence, the time lapsed after exposure to infection, and host factors, including race-/ethnicity-specific response to infection, severity of infection, and other lifestyle factors pertinent to this population may vary in subgroups and may collectively shape the phenotypic association to Ad36 infection. The first step toward understanding such genetic and nongenetic contributors of the metabolic associations of Ad36 is to identify the subgroup differences.
Study limitations
Although the results are consistent with our expectations based on previous reports by us and others (2,7,13,38,39), the seropositivity-based differences in adiposity and glycemic control are modest in magnitude. Moreover, we investigated the association of Ad36 seropositivity with adiposity and glycemic control in three BMI subgroups and both sexes. This required the adjustment for multiple comparisons. Following the commonly used guidelines on statistical reporting (40), we reported the exact P values. However, if a conservative Bonferroni correction is applied to the multiple testing (2 sexes × 3 BMI categories × 4 traits × 2 types of analysis [baseline and longitudinal] = 48 tests), the P value for significance should be considered to be approximately ≤0.001. None of our results reach this level of statistical significance, but such an adjustment is likely overly conservative given the correlated nature of the phenotypic variables. In any case, caution when interpreting these results is warranted.
BMI is a continuous variable and an argument could be made against stratifying by obesity level for analyses. However, to reflect clinical relevance of our findings, we selected the three categories of BMI that are used extensively. Also, a recent study suggested that weight gain over time may depend on baseline adiposity (41). Therefore, we used BMI subgroup analyses, which revealed some interesting insight about the contribution of individual BMI categories to the results.
Neutralizing antibodies to viruses generally persist for a long time. So, identifying the seropositive group in these longitudinal analyses is straightforward. However, time elapsed since Ad36 infection is unknown in this study. Also, it is unknown if the phenotypic association with seropositivity fades with time. Ad36 antibody status was determined only at baseline. Therefore, it is possible that subjects deemed seronegative at baseline may have experienced seroconversion later during the 10-year follow-up period. This would presumably bias the comparison against the current findings (by including such seroconverted subjects in the seronegative category). The significance was noted in the Ad36 seropositive groups, despite this possibility and despite the long follow-up duration. Although screening for neutralizing antibodies is highly resource-intensive, determining antibody status at the end of the follow-up period may strengthen the association further.
Despite these limitations, this study is an important initial step in unraveling our understanding of how some pathogens may chronically interact with metabolic pathways. Although this study indicates an association, and not causation, taken together with previous in vitro and in vivo data, and with human cross-sectional data, these associations render more plausible the idea that in humans Ad36 increases adiposity and improves glycemic control. Importantly, in specific subgroups, Ad36 seropositivity may be a protective factor for hyperglycemia or insulin resistance. If so, then this is potentially a property that could be harnessed to improve diabetes, as demonstrated by the experimental Ad36 infection of mice with diet-induced hyperglycemia (7). Clearly, it is impractical to infect humans with Ad36 to improve diabetes. However, E4orf1, a viral protein that is necessary and sufficient for the antihyeprglyemic effect of Ad36 (15), may be used to develop an antidiabetic agent. Improvement in glycemic control predicted by natural Ad36 infection in humans conceptually supports the potential of the Ad36 E4orf1 protein to improve glycemic control in humans. A recent study reported another potential implication of determining Ad36 infection status. In response to a reduced energy diet, Ad36 seropositive adults lose more weight and have improved insulin sensitivity and reduced hepatic steatosis compared with seronegative individuals (39). Ad36 infection appears to be a predictor of response to weight loss treatment.
Importantly, the results advance the possibility that in addition to genetic, physiological, and behavioral factors, certain asymptomatic infections may lastingly influence human metabolism. A deeper understanding of these under-recognized factors may help in designing treatment or prevention strategies for such metabolic disorders.
Supplementary Material
Acknowledgments
The pedigree study is supported by a grant from the U.S. National Heart, Lung, and Blood Institute (HL-045522 to J.B.). Funds for serological assays were provided by the National Institutes of Health (HL-080149 to H.G.). The funding agencies had no role in the analysis, and interpretation of data, in the writing of the report, or in the decision to submit the manuscript for publication.
For N.V.D., the following patents are granted or have been applied for: Patent number 6,127,113: Viral obesity methods and compositions; Patent number 6,664,050: Viral obesity methods and compositions; Patent number US 8,008,436B2, dated August 30, 2011: Adenovirus 36 E4orf1 gene and protein and their uses; Provisional patent filed: Adenovirus Ad36 E4orf1 protein for prevention and treatment of nonalcoholic fatty liver disease, July 2010; Provisional patent filed: Enhanced glycemic control using Ad36 E4orf1 and AKT1 inhibitor, January 2012. N.V.D. has ongoing grant support from Vital Health Interventions for determining antidiabetic properties of E4orf1 protein. D.B.A. has, anticipates, or has had financial interests with the Frontiers Foundation; Federal Trade Commission; Vivus, Inc.; Kraft Foods; University of Wisconsin; University of Arizona; Paul, Weiss, Wharton & Garrison LLP; and Sage Publications. No other potential conflicts of interest relevant to this article were reported.
W.-Y.L., R.R., N.L., and D.B.A. analyzed data. W.-Y.L., N.L., and D.B.A. prepared tables and wrote the statistical analyses section/methods. O.D. conducted entire serum neutralization analyses and organized data. W.-Y.L., O.D., R.R., N.L., D.B.A., J.E.C., A.G.C., J.B., C.T.L., and H.G. reviewed the manuscript. N.V.D. generated hypotheses, coordinated the project, provided input for laboratory and statistical analyses and interpretation, and prepared the manuscript draft. J.E.C., A.G.C., and J.B. provided phenotypic and genotypic data. J.B. supervised the parent study. C.T.L. planned and coordinated the study. H.G. analyzed data, edited the manuscript, and planned and coordinated the study. All authors, external and internal, had full access to all of the data (including statistical reports and tables) in the study and can take responsibility for the integrity of the data and the accuracy of the data analysis. H.G. and N.V.D. are the guarantors of this work and, as such, had full access to all the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis.
Parts of this study were presented in abstract form at the Annual Meeting of The Obesity Society, San Antonio, Texas, 20–24 September 2012.
The authors thank the participants of the San Antonio Family Heart Study, and Dr. Ana Inés Vázquez and Dr. Gustavo de los Campos (both of the University of Alabama at Birmingham) for technical support.
Footnotes
This article contains Supplementary Data online at http://care.diabetesjournals.org/lookup/suppl/doi:10.2337/dc12-1089/-/DC1.
References
- 1.McAllister EJ, Dhurandhar NV, Keith SW, et al. Ten putative contributors to the obesity epidemic. Crit Rev Food Sci Nutr 2009;49:868–913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dhurandhar NV. A framework for identification of infections that contribute to human obesity. Lancet Infect Dis 2011;11:963–969 [DOI] [PubMed] [Google Scholar]
- 3.Dhurandhar NV, Israel BA, Kolesar JM, Mayhew GF, Cook ME, Atkinson RL. Increased adiposity in animals due to a human virus. Int J Obes Relat Metab Disord 2000;24:989–996 [DOI] [PubMed] [Google Scholar]
- 4.Dhurandhar NV, Israel BA, Kolesar JM, Mayhew G, Cook ME, Atkinson RL. Transmissibility of adenovirus-induced adiposity in a chicken model. Int J Obes Relat Metab Disord 2001;25:990–996 [DOI] [PubMed] [Google Scholar]
- 5.Dhurandhar NV, Whigham LD, Abbott DH, et al. Human adenovirus Ad-36 promotes weight gain in male rhesus and marmoset monkeys. J Nutr 2002;132:3155–3160 [DOI] [PubMed] [Google Scholar]
- 6.Pasarica M, Shin AC, Yu M, et al. Human adenovirus 36 induces adiposity, increases insulin sensitivity, and alters hypothalamic monoamines in rats. Obesity (Silver Spring) 2006;14:1905–1913 [DOI] [PubMed] [Google Scholar]
- 7.Krishnapuram R, Dhurandhar EJ, Dubuisson O, et al. Template to improve glycemic control without reducing adiposity or dietary fat. Am J Physiol Endocrinol Metab 2011;300:E779–E789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vangipuram SD, Sheele J, Atkinson RL, Holland TC, Dhurandhar NV. A human adenovirus enhances preadipocyte differentiation. Obes Res 2004;12:770–777 [DOI] [PubMed] [Google Scholar]
- 9.Rathod M, Vangipuram SD, Krishnan B, Heydari AR, Holland TC, Dhurandhar NV. Viral mRNA expression but not DNA replication is required for lipogenic effect of human adenovirus Ad-36 in preadipocytes. Int J Obes (Lond) 2007;31:78–86 [DOI] [PubMed] [Google Scholar]
- 10.Pasarica M, Mashtalir N, McAllister EJ, et al. Adipogenic human adenovirus Ad-36 induces commitment, differentiation, and lipid accumulation in human adipose-derived stem cells. Stem Cells 2008;26:969–978 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pasarica M, Loiler S, Dhurandhar NV. Acute effect of infection by adipogenic human adenovirus Ad36. Arch Virol 2008;153:2097–2102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rathod MA, Rogers PM, Vangipuram SD, McAllister EJ, Dhurandhar NV. Adipogenic cascade can be induced without adipogenic media by a human adenovirus. Obesity (Silver Spring) 2009;17:657–664 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rogers PM, Mashtalir N, Rathod MA, et al. Metabolically favorable remodeling of human adipose tissue by human adenovirus type 36. Diabetes 2008;57:2321–2331 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wang ZQ, Cefalu WT, Zhang XH, et al. Human adenovirus type 36 enhances glucose uptake in diabetic and nondiabetic human skeletal muscle cells independent of insulin signaling. Diabetes 2008;57:1805–1813 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dhurandhar EJ, Dubuisson O, Mashtalir N, Krishnapuram R, Hegde V, Dhurandhar NV. E4orf1: a novel ligand that improves glucose disposal in cell culture. PLoS ONE 2011;6:e23394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dubuisson O, Dhurandhar EJ, Krishnapuram R, et al. PPARgamma-independent increase in glucose uptake and adiponectin abundance in fat cells. Endocrinology 2011;152:3648–3660 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Khan T, Muise ES, Iyengar P, et al. Metabolic dysregulation and adipose tissue fibrosis: The role of collagen VI. Mol Cell Biol 2009;6:1575–1591 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kim JY, van de Wall E, Laplante M, et al. Obesity-associated improvements in metabolic profile through expansion of adipose tissue. J Clin Invest 2007;117:2621–2637 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Gavrilova O, Marcus-Samuels B, Graham D, et al. Surgical implantation of adipose tissue reverses diabetes in lipoatrophic mice. J Clin Invest 2000;105:271–278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Larsen TM, Toubro S, Astrup A. PPARgamma agonists in the treatment of type II diabetes: is increased fatness commensurate with long-term efficacy? Int J Obes Relat Metab Disord 2003;27:147–161 [DOI] [PubMed] [Google Scholar]
- 21.Mast TC, Kierstead L, Gupta SB, et al. International epidemiology of human pre-existing adenovirus (Ad) type-5, type-6, type-26 and type-36 neutralizing antibodies: correlates of high Ad5 titers and implications for potential HIV vaccine trials. Vaccine 2010;28:950–957 [DOI] [PubMed] [Google Scholar]
- 22.Atkinson RL, Dhurandhar NV, Allison DB, et al. Human adenovirus-36 is associated with increased body weight and paradoxical reduction of serum lipids. Int J Obes (Lond) 2005;29:281–286 [DOI] [PubMed] [Google Scholar]
- 23.Trovato GM, Castro A, Tonzuso A, et al. Human obesity relationship with Ad36 adenovirus and insulin resistance. Int J Obes (Lond) 2009;33:1402–1409 [DOI] [PubMed] [Google Scholar]
- 24.Na HN, Kim J, Lee HS, et al. Association of human adenovirus-36 in overweight Korean adults. Int J Obes (Lond) 2012;36:281–285 [DOI] [PubMed] [Google Scholar]
- 25.Gabbert C, Donohue M, Arnold J, Schwimmer JB. Adenovirus 36 and obesity in children and adolescents. Pediatrics 2010;126:721–726 [DOI] [PubMed] [Google Scholar]
- 26.Atkinson RL, Lee I, Shin HJ, He J. Human adenovirus-36 antibody status is associated with obesity in children. Int J Pediatr Obes 2010;5:157–160 [DOI] [PubMed] [Google Scholar]
- 27.Na HN, Hong YM, Kim J, Kim HK, Jo I, Nam JH. Association between human adenovirus-36 and lipid disorders in Korean schoolchildren. Int J Obes (Lond) 2010;34:89–93 [DOI] [PubMed] [Google Scholar]
- 28.Trovato GM, Martines GF, Garozzo A, et al. Ad36 adipogenic adenovirus in human non-alcoholic fatty liver disease. Liver Int 2010;30:184–190 [DOI] [PubMed] [Google Scholar]
- 29.Mitchell BD, Kammerer CM, Blangero J, et al. Genetic and environmental contributions to cardiovascular risk factors in Mexican Americans. The San Antonio Family Heart Study. Circulation 1996;94:2159–2170 [DOI] [PubMed] [Google Scholar]
- 30.de los Campos G, Naya H, Gianola D, et al. Predicting quantitative traits with regression models for dense molecular markers and pedigree. Genetics 2009;182:375–385 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pérez P, de Los Campos G, Crossa J, Gianola D. Genomic-enabled prediction based on molecular markers and pedigree using the Bayesian linear regression package in R. Plant Genome 2010;3:106–116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Vazquez AI, Bates DM, Rosa GJ, Gianola D, Weigel KA. Technical note: an R package for fitting generalized linear mixed models in animal breeding. J Anim Sci 2010;88:497–504 [DOI] [PubMed] [Google Scholar]
- 33.Pascot A, Lemieux S, Lemieux I, et al. Age-related increase in visceral adipose tissue and body fat and the metabolic risk profile of premenopausal women. Diabetes Care 1999;22:1471–1478 [DOI] [PubMed] [Google Scholar]
- 34.Petersen KF, Befroy D, Dufour S, et al. Mitochondrial dysfunction in the elderly: possible role in insulin resistance. Science 2003;300:1140–1142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ai M, Otokozawa S, Asztalos BF, et al. Adiponectin: an independent risk factor for coronary heart disease in men in the Framingham offspring Study. Atherosclerosis 2011;217:543–548 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Pajvani UB, Scherer PE. Adiponectin: systemic contributor to insulin sensitivity. Curr Diab Rep 2003;3:207–213 [DOI] [PubMed] [Google Scholar]
- 37.Cadwell K, Patel KK, Maloney NS, et al. Virus-plus-susceptibility gene interaction determines Crohn’s disease gene Atg16L1 phenotypes in intestine. Cell 2010;141:1135–1145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Krishnapuram R, Kirk-Ballard H, Dhurandhar EJ, et al. NV D. Insulin receptor-independent upregulation of cellular glucose uptake. Int J Obes (Lond) 2013;37:146–153 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Trovato GM, Martines GF, Trovato FM, et al. Adenovirus-36 seropositivity enhances effects of nutritional intervention on obesity, bright liver, and insulin resistance. Dig Dis Sci 2012;57:535–544 [DOI] [PubMed] [Google Scholar]
- 40.Bailar JC, 3rd, Mosteller F. Guidelines for statistical reporting in articles for medical journals. Amplifications and explanations. Ann Intern Med 1988;108:266–273 [DOI] [PubMed] [Google Scholar]
- 41.Williams PT. Evidence that obesity risk factor potencies are weight dependent, a phenomenon that may explain accelerated weight gain in western societies. PLoS ONE 2011;6:e27657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zellner A, Keuzenkamp H, McAleer M. Simplicity, inference and modelling: keeping it sophisticatedly simple. Cambridge, Cambridge University Press, 2004 [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.