

Abstract
Several Gram-negative pathogenic bacteria employ N-acyl-L-homoserine lactone (HSL) quorum sensing (QS) system to control their virulence traits. Degradation of acyl-HSL signal molecules by quorum quenching enzyme (QQE) results in a loss of pathogenicity in QS-dependent organisms. The QQE activity of actinomycetes in rhizospheric soil and inside plant tissue was explored in order to obtain novel strains with high HSL-degrading activity. Among 344 rhizospheric and 132 endophytic isolates, 127 (36.9%) and 68 (51.5%) of them, respectively, possessed the QQE activity. The highest HSL-degrading activity was at 151.30 ± 3.1 nmole/h/mL from an endophytic actinomycetes isolate, LPC029. The isolate was identified as Streptomyces based on 16S rRNA gene sequence similarity. The QQE from LPC029 revealed HSL-acylase activity that was able to cleave an amide bond of acyl-side chain in HSL substrate as determined by HPLC. LPC029 HSL-acylase showed broad substrate specificity from C6- to C12-HSL in which C10HSL is the most favorable substrate for this enzyme. In an in vitro pathogenicity assay, the partially purified HSL-acylase efficiently suppressed soft rot of potato caused by Pectobacterium carotovorum ssp. carotovorum as demonstrated. To our knowledge, this is the first report of HSL-acylase activity derived from an endophytic Streptomyces.
1. Introduction
Bacterial pathogens infecting plants, animals, and humans cause a tremendous economic loss worldwide. Conventional treatments, such as application of antibacterial agents, significantly contribute to selection of resistant microorganisms as well as environmental contamination [1]. A number of concerns on using antibiotics have brought investigators to search for alternative strategies to combat with these pathogens. Attenuation of virulence phenotypes rather than killing of causative agents has gained an interest since this strategy does not introduce selective pressure which potentially leads to development of antibiotic-resistant bacteria [2]. Certain bacterial pathogens regulate their virulence by monitoring population density which is known as quorum sensing (QS) mechanism [3–5]. In Gram-negative bacteria, QS is often mediated by N-acyl-L-homoserine lactone (HSL) signal molecules [4]. Interference of HSL-dependent QS, commonly known as quorum quenching (QQ), has been demonstrated to be an effective antimicrobial strategy for controlling virulent pathogens [6–9]. QQ can be achieved by degradation of QS signal molecules by enzymatic digestion, significantly decreasing functions of signaling molecules. The QQ enzymes (QQEs) have been reported from several Gram-positive bacteria including Arthrobacter sp. IBN110, Bacillus sp. 240B1, Geobacillus kaustophilus strain HTA426, Mycobacterium avium subsp. paratuberculosis K-10, Solibacillus silvestris StLB046 [10], and Streptomyces M664 [8]. However, there are differences in the catalytic spectrum among QQEs toward HSLs. In the search for QQE-producing bacteria, actinomycetes are of great interest since they possess an ability to produce and secrete various extracellular hydrolytic enzymes [11, 12]. A number of actinomycetes have been isolated from several natural sources, including rhizospheric soil and plant tissues. Biological functions of actinomycetes predominantly depend on sources from which the bacteria are isolated. Natural resources in megabiodiversity with high selective pressure and microbial competition in tropical regions are well recognized as an important resource of new anti-microbial agents as well as QQE [13].
To date, the isolation of actinomycetes with high QQE activity from endophytes has never been reported. Screening for QQE-producing actinomycetes in this study showed the diversity and abundance of HSL-degrading actinomycetes from soil and plant tissues for the first time. The QQE from the high activity strain was shown to hydrolyze HSL with a broad range of chain length from N-hexanoyl-L-homoserine lactone (C6HSL) to N-dodecanoyl-L-homoserine lactone (C12HSL); hence, this enzyme could be potentially used to attenuate virulence of a broad range of bacterial pathogens with different QS signal molecules.
2. Materials and Methods
2.1. Isolation of Rhizospheric and Endophytic Actinomycetes
Rhizospheric soil samples were collected from various provinces in Thailand, including Bangkok, Chanthaburi, Nongbualumpoo, Prachinburi, and Rayong provinces. To isolate rhizospheric actinomycetes, 1.0 g of soil was serially diluted in 4 mL of 0.85% normal saline solution (NSS). The appropriate dilution was spread on selective agars, which are Pridham's and water proline supplemented with 25 μg/mL nalidixic acid and 50 μg/mL cyclohexamide [14]. Isolates with typical actinomycetes colony morphology were selected for screening of HSL-degrading activity. In order to isolate endophytic actinomycetes, leaves, fruits, seeds, and stems of plant samples were collected from various regions in Thailand including Bangkok, Salaya botanical garden in Nakhon pathom, and Nam Nao national park in Phetchabun. All dirt was removed by running tap water. Samples were air-dried at room temperature for one week. Plant samples were cut into small pieces. Surface sterilization was performed by treating with 10% sodium hypochlorite (MERCK), 70% ethanol and followed by washing twice with sterile water. The samples were then air-dried and placed on water agar medium (containing only 1.5% agar) supplemented with 25 μg/mL nalidixic acid and 50 μg/mL cyclohexamide. Actinomycetes colonies were selected for further study.
2.2. HSL Inhibition Assay
Actinomycetes isolates were grown on Waskman's agar (1% glucose, 0.5% peptone, 0.5% meat extract, 0.3% NaCl, and 1.2% agar) for 3 days, subcultured in 301 broth (2.4% starch, 0.1% glucose, 0.3% peptone, 0.5% meat extract, and 0.3% CaCO3), and further incubated at 28°C with shaking for 6 days. Forty microliters of cell-free supernatant was mixed with an equal volume of 40 µM N-decanoyl-L-homoserine lactone (C10HSL) in 0.1 M Tris-HCl pH 6.8. The mixture was incubated at 28°C for 1 h with gentle agitation and the reaction was stopped by heating at 95°C for 5 min. Then 10 µL of reaction mixture was used to determine the remaining of C10HSL on a bioassay plate. For determination of HSL-degrading activity, Agrobacterium tumefaciens NTL4 (pZLR4) (kindly provided by Professor Stephen K. Farrand, Department of Crop Sciences and Microbiology, University of Illinois, USA) was used as a biosensor strain in bioassay to long-chain HSL (C8 to C12HSL) [15]. Chromobacterium violaceum CV026 (kindly provided by Professor Paul Williams, Department of Molecular Microbiology, University of Nottingham, UK) was used to detect N-butanoyl-L-homoserine lactone (C4HSL), N-hexanoyl-L-homoserine lactone (C6HSL), and 3-oxohexanoyl-L-homoserine lactone (3-oxo-C6HSL) [16]. Each reporter strain was cultivated overnight in nutrient broth (NB) and Luria-Bertani broth (LB), respectively. To determine HSL-degrading activity qualitatively, an overnight culture of the biosensor strain (2.5 mL) was mixed with 5 mL of agar. The mixture was overlaid on an AB minimal medium bioassay plate [17] supplemented with 40 µg/mL of 5-bromo-4-chloro-3-indolyl-D-galactopyranoside (X-gal) for A. tumefaciens NTL4 (pZLR4) and Luria-Bertani agar (LA) for C. violaceum (CV026). A well on an agar plate was made by punching with a cork borer (∅ = 0.4 cm). Ten microliters of the heat inactivated reaction mixture was dropped in each well. All plates were incubated at 30°C overnight to allow color zone developing. Blue and purple color zones were developed around colonies of A. tumefaciens NTL4 (pZLR4) and C. violaceum (CV026), respectively, by the induction of the residual HSL in the reaction mixture. The residual amounts of HSL were calculated using relationship equations based on the color zone size and known amounts of HSL [6]. Relative activity of HSL-degrading enzyme was calculated by using the following formula: relative HSL-degrading activity = (HSL-degrading activity/initial amount of substrate) × 100. For quantitative HSL-degrading activity determination, bioassay agar medium in the plate was cut into separated slices across the plate (1 cm in width). Five microliters of the reaction mixture was added to one end of an agar slice and then the culture of biosensor strain at an OD600 of 1 was progressively spotted (0.6 µL per spot) at further distances from the loaded reaction mixture. The last induced color colony to the origin of the reaction mixture sample in each agar slice was measured. Amounts of HSL in the reaction mixture were determined based on relationship equation of adding known amounts of HSL to the bioassay slices and determining the distance of the color colony from the origin. HSL-degrading activity reported as nmole/h was calculated from subtraction of the initial amount of HSL substrate with leftover amount from the enzymatic digestion.
2.3. Strain Identification
The 16S rRNA gene of isolate LPC029 was amplified using conserved primers [18]. PCR was performed for 30 cycles at 95°C for 45 seconds, 45°C for 45 seconds, and 72°C from 1.5 minutes. The PCR product was sequenced by the dideoxy chain-termination method [19].
2.4. Preparation of Partially Purified HSL-Degrading Enzyme
The endophytic isolate LPC029 was harvested after 6 days of culture in 1,000 mL 301 medium by centrifugation at 4°C (10,000 g) for 10 min. The supernatant was filtered through a 0.45 μm membrane and stored at 4°C. Ammonium sulfate was added to the culture supernatant to achieve 60% saturation and the solution was left at 4°C for 16 h. The precipitate was collected by centrifugation, dissolved, in 6 mL of 20 mM sodium phosphate buffer pH 7.0, and dialyzed (dialysis tubing, MW cut-off 10 kDa; Sigma, USA) overnight against the same buffer. The dialyzed sample was then collected and stored at 4°C.
2.5. HSL Degradation Assay
C10HSL was used as a substrate in the determination of hydrolytic products of the HSL-degrading enzyme. Substrates were prepared as a stock solution at 8 mM in 80% ethanol. The HSL-degrading product was analyzed by mixing 13 µg of the partially purified HSL-degrading enzyme and 350 mM of C10HSL in 700 mL of 20 mM sodium phosphate buffer pH 7.0. After incubation at 30°C for 16 h, the reaction mixture was extracted three times with equal volumes of ethyl acetate. The organic phase was evaporated to dryness at 40°C by a rotary evaporator. The dried sample was dissolved in 50 µL of HPLC grade methanol. Five microliters of HSL-degraded products was introduced onto Shiseido Capcell Pak C18 (4.6 × 250 mm I.D.; particle size 5 µm) previously equilibrated with 10% acetonitrile. The elution was performed with linear gradient of 10–90% acetonitrile, at a flow rate of 1 mL/min under the following condition: 10% acetonitrile from 0 to 8 min; linear gradient to 90% acetonitrile from 8 to 35 min. Homoserine lactone (3.5 mM) and 35 mM of C10HSL were used as a standard. HPLC chromatogram of the products was analyzed by diode array detector with UV ranging from 200 to 280 nm (Agilent 1200 HPLC). To check substrate specificity, HSL-acylase was reacted with different HSL-substrates including C4HSL, C6HSL, 3-oxo-C6HSL, 3-oxo-C8HSL, C10HSL, and C12HSL. Reaction mixture was composed of 0.4 mM each of HSL substances in 20 mM sodium phosphate at pH 7.0 and 2 µg of the partially purified HSL-degrading enzyme in 50 µL. The mixture was incubated at 30°C for 1 h. The HSL-substrate that remained in the reaction mixture was determined by the bioassay method as described above.
2.6. In Vitro Pathogenicity Assay
The assay was performed as described by Burr et al. [20]. The potatoes' tubers of about the same size were washed with tab water and pretreated in 5% sodium hypochlorite (Merck) for 10 min then soaked with sterile water. The tubers were dried in a laminar flow cabinet. P. carotovorum ssp. carotovorum (Pcc) was cultured in 50 ml of LB broth at 30°C overnight. The culture at OD600 of 1 was further serially diluted with 0.85% NSS to 10−5. Ten microliters of 100,000-folded dilution of Pcc (ca. 1000 cfu/10 µL) was mixed with 2 µg of partially purified HSL-degrading enzyme. The reaction was then further incubated at 30°C for 1 h and was inoculated into a potato tuber. A two-hundred-microliter pipette tip was used to punch a hole of 22 mm in depth, 2 holes per tuber. The two holes in each potato tuber were each filled with 0.85% NSS (negative control), 1000 cells of Pcc (positive control), the mixture of Pcc and HSL-acylase, and HSL-acylase alone. The inoculated potato tubers were further sealed with a sterilized sticker. To make a moisture condition, the tubers were wrapped with aluminum foil and sterile moist towels. The wrapped-potato tubers were incubated in a closed box at 30°C for 3 days.
3. Results and Discussion
3.1. Screening of HSL-Degrading Actinomycetes and HSL-Degrading Activity
The abundance and diversity of HSL-degrading actinomycetes isolated from soil and plant tissues were assessed. They were isolated on the basis of their typical morphologies (filamentous growth, spore chain, and several types of convex and margin colonies on selective medium) according to Bergey's Manual of Systematic Bacteriology [21]. While 344 actinomycetes could be isolated from 43 rhizospheric soil samples, only 132 isolates were obtained from 64 plant samples. Among these actinomycetes isolates, the number of isolates with HSL degrading activity was found in higher frequency in endophytic isolates (51.5%) than in rhizospheric isolates (36.9%) (Table 1). While several previous studies have indicated the HSL-degrading activity in soil bacteria [8, 22, 23], this study is the first report of such activity in endophytic actinomycetes. The evolution race for survival in tropical ecosystem reinforces strong competition among organisms, which may result in a plethora of chemical molecules [24], and enzymes [11, 12] with biological functions. Consequently, there is a high probability that microorganisms associated with tropical plants might be a source of bioactive compounds and enzymes. Among our isolates with HSL-degrading activity, 4 rhizospheric actinomycetes (PS032, SWP036, SWP042, and SWP043) and 5 endophytic actinomycetes (LPC026, LPC029, PC005, PC052, and PC053) were found to be the potential strains in HSL degrading as they were able to degrade HSL of greater than 96.9% relative activity (Table 1). Quantitative measurement of enzymatic activity among these 9 isolates showed that isolate LPC029 from Gmelina arborea Roxb. possessed the highest HSL-degrading activity at 151.30 ± 3.1 nmole/h/mL (Table 2). Strain LPC029 was later identified with 16S rRNA gene sequence (GenBank accession number KC153060) to be closely related to S. globisporus at 99% homology. The presence of HSL-degrading activity in both soil and plant isolates may be due to their common need of HSL-degraded product as carbon and energy sources [25] and for competition with other microbes to protect their ecological niche [26]. In addition, endophytic bacteria have previously been reported to be involved in several biological functions such as promoting plant growth, biocontrol agents, and phytoremediation [24, 27–29]. These functions stem from the production of their natural compounds and antipathogen metabolites inside the host plant [29].
Table 1.
Relative HSL-degrading activity of actinomycetes isolated from rhizospheric and endophytic samples.
| Location | No. of isolates |
Relative HSL-degrading activity* (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 | >0–50 | >50–75 | >75–87.5 | >87.5–93.8 | >93.8–96.9 | >96.9–100 | ||
| Rhizospheric samples | ||||||||
| Bangkok | 278 | 182 | 54 | 16 | 11 | 8 | 4 | 3 |
| Chanthaburi | 13 | 6 | 5 | 0 | 0 | 1 | 1 | 0 |
| Nongbualumpoo | 3 | 2 | 0 | 0 | 0 | 0 | 0 | 1 |
| Prachinburi | 27 | 12 | 9 | 3 | 3 | 0 | 0 | 0 |
| Rayong | 23 | 15 | 8 | 0 | 0 | 0 | 0 | 0 |
|
| ||||||||
| Total | 344 | 217 | 76 | 19 | 14 | 9 | 5 | 4 |
|
| ||||||||
| Total (percentage) | 217 (63.1%) | 127 (36.9%) | ||||||
|
| ||||||||
| Endophytic samples | ||||||||
| Bangkok | 43 | 29 | 12 | 0 | 1 | 1 | 0 | 0 |
| Nakhonpathom | 29 | 20 | 8 | 1 | 0 | 0 | 0 | 0 |
| Phetchabun | 60 | 15 | 25 | 0 | 10 | 1 | 4 | 5 |
|
| ||||||||
| Total | 132 | 64 | 45 | 1 | 11 | 2 | 4 | 5 |
|
| ||||||||
| Total (percentage) | 64 (48.5%) | 68 (51.5%) | ||||||
*The activity of each isolate was qualitatively determined by comparing with known concentrations of C10HSL (200, 100, 50, 25, 12.5, 0.625, and 0 pmole).
Table 2.
Quantitative HSL-degrading activity of isolates with high efficiency of HSL degradation.
| Source | Name of isolate | HSL-degrading activity (nmole/h/mL) |
|---|---|---|
| Rhizosphere soil, Nongbualumpoo | PS032 | 143.42 ± 1.5 |
| Rhizosphere soil, Bangkok | SWP036 | 144.67 ± 2.1 |
| Rhizosphere soil, Bangkok | SWP042 | 144.37 ± 1.5 |
| Rhizosphere soil, Bangkok | SWP043 | 144.26 ± 1.3 |
| Leaf of Ta khram (Garuga pinnata Roxb.) | LPC026 | 146.45 ± 2.8 |
| Leaf of So (Gmelina arborea Roxb.) | LPC029 | 151.30 ± 3.1 |
| Leaf of Kling klang dong (Stephania venosa Spreng.) | PC005 | 146.93 ± 1.0 |
| Leaf of Malabar melastome (Melastoma malabathricum L.) | PC052 | 143.45 ± 4.3 |
| Leaf of Grape-leaf Wood Rose (Merremia vitifolia Hallier f.) | PC053 | 147.68 ± 1.8 |
3.2. HPLC Analysis of C10HSL Hydrolysis by LPC029 HSL-Degrading Enzyme
The products of HSL degradation by enzyme from LPC029 were analyzed by HPLC after ethyl acetate extraction. Hydrolysis of C10HSL by LPC029 enzyme resulted in releasing HSL as shown by HPLC (Figure 1(c)) compared to standard HSL (Figure 1(a)) and C10HSL substrate alone (Figure 1(b)). Therefore the HSL-degrading enzyme from LPC029 was proven to be HSL-acylase according to the deacylation activity to break the amide bond of C10HSL, out of which HSL was released as an end product. The substrate (C10HSL) was incompletely hydrolyzed (Figure 1(c)) due to an insufficient amount of enzyme used in the assay. HSL-acylase has been explored in (i) Gram-negative bacteria including Comamonas strain D1 [9], Pseudomonas aeruginosa PAO1 [30], P. aeruginosa PAI-A [31], P. syringae [32], Ralstonia solanacearum GMI1000 [33], Shewanella sp. strain MIB 015 [7], Tenacibaculum maritimum [34], and Variovorax paradoxus VAI-C [25]; (ii) Gram-positive bacteria including Rhodococcus erythropolis strain W2 [35] and Streptomyces sp. M664 [8]; and (iii) cyanobacteria such as Anabaena sp. PCC 7120 [36]. Comparison of HSL-acylase activity among these HSL-degrading microbes is difficult due to the difference of both quantitative and qualitative measurement methods. Nevertheless, substrate specificity of the enzymes could be compared qualitatively. Although the enzymes were derived from a variety of microorganisms, most of HSL-acylases were active against HSL with the carbon chain less than 8 atoms [10]. Testing of the partial purified enzyme from LPC029 with various HSL substrates showed the ability to degrade medium-to-long chains of HSL (C6–C12) in which C10HSL was the most preferred substrate (Figure 2). The activity of this enzyme was not dependent on the substitution of oxo-group on the third carbon atom (Figure 2) which showed no significant difference between two substrates, C6HSL and 3-oxo-C6HSL (P ≥ 0.05). The results suggested the high possibility of LPC029 HSL-acylase in inhibiting QS of medium-chain HSLs dependent on Pcc (3-oxo-C6HSL, 3-oxo-C8HSL) and Burkholderia cepacia (C6HSL, C8HSL) and might provide the most effective quenching agent against long-chain HSLs produced by Vibrio anguillarum (C10HSL) and P. aeruginosa (C12HSL).
Figure 1.
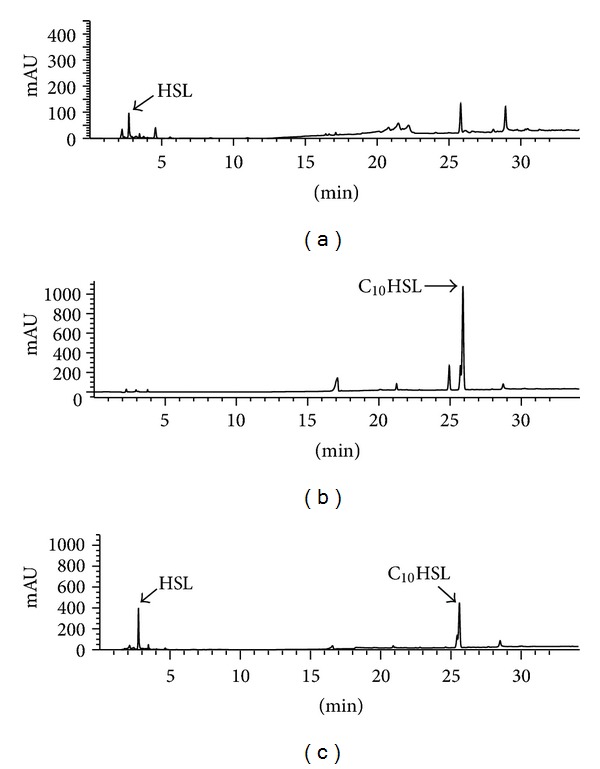
HPLC analysis of enzymatic hydrolysis product of C10HSL with the HSL-degrading enzyme from LPC029. The initial C10HSL (35 mM) was reacted with partially purified enzyme from LPC029 for 16 h and separated with HPLC. HPLC profiles of (a) HSL standard (3.5 mM); (b) unreacted C10HSL (35 mM); and (c) reaction product of C10HSL with LPC029 HSL-degrading enzyme. The product peak was eluted at 2.8 min. mAU is the abbreviation for milli-absorbance unit.
Figure 2.
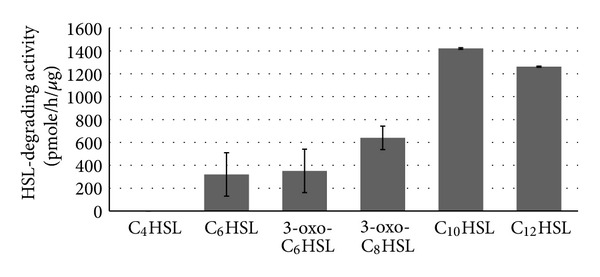
Substrate specificity of LPC029 HSL-degrading enzyme. Partially purified enzyme was mixed with each of HSLs. The activity was determined based on HSL leftover on C. violaceum CV026 and A. tumefaciens NTL4 (pZLR4) bioassay plates. Relative activity is given in parenthesis. Bars indicate SD values of four replicates.
3.3. Effect of LPC029 HSL-Degrading Enzyme on Pathogenicity
In vitro efficacy of HSL-degrading enzyme from LPC029 to quench QS-regulated functions was determined by Pectobacterium carotovorum ssp. carotovorum (Pcc) mediated soft-rotting potato tuber assay. In Pcc, the virulence and pathogenicity are QS dependent, but different strains of this subspecies prefer different signal molecules and they are divided into two groups. One group used 3-oxo-C6HSL as a predominant signal molecules, whereas another group exploited mainly 3-oxo-C8HSL as HSL-dependent QS [37]. Inoculation of Pcc (103 cells per tuber) evidently resulted in tissue maceration. The addition of 2 µg of partially purified HSL-acylase resulted in a statistically significant reduction in soft-rot weight (P < 0.05) as shown in Figure 3(e). In the control experiment, neither NSS nor HSL-acylase inoculation induced soft rot in plant tissue (Figures 3(a) and 3(d), resp.). Inoculation of Pcc alone (103 cells) caused the highest plant tissue necrosis (Figure 3(b)) with the highest soft-rot weight (Figure 3(e)). When the same number of Pcc (103 cells) was mixed with 2 µg of HSL-acylase before inoculation, the enzyme attenuated the bacterial pathogenicity by reduction of plant tissue necrosis (Figure 3(c)) and caused reduction of soft-rot weight (Figure 3(e)). Several studies have shown the effectiveness of QS targeting strategy in reducing virulence of P. aeruginosa [38], Enterobacteriaceae [39], B. cepacia [40], P. carotovorum [23, 41, 42], Serratia liquefaciens [43], Agrobacterium tumefaciens [23], and some aquatic consortia [44]. These results offer an alternative strategy to alternate virulence of plant pathogen using QS-degrading enzyme from endophytic Streptomyces. Due to its ability to degrade long-chain HSL with moderate activity toward short-chain HSL, the QQE from Streptomyces LPC029 might be of interest to control certain plant and human pathogens. However, more in-depth in vitro and also in vivo investigations are warranted.
Figure 3.

Effects of LPC029 HSL-degrading enzyme on pathogenicity of Pcc on potato tubers. Pathogenicity of Pcc was determined by inspection of lesion zones induced upon inoculation of potato tuber at two different sites. (a) Negative control consisting of tuber treated with 0.85% NSS, (b) inoculation of Pcc alone at 103 cells per tuber, (c) inoculation of Pcc at 103 cells mixed with 2 µg of partial purified HSL-acylase per tuber, (d) 2 µg of partially purified HSL-acylase, and (e) pathogenicity of Pcc determined by soft-rot weight. Soft-rot tissue from treatment (a)–(d) was measured after inoculation of Pcc for 3 days; bars indicate SD values of four replicates.
4. Conclusion
The HSL-degrading activity among rhizospheric and endophytic actinomycetes was assessed and the latter was found to possess this enzyme activity at higher frequency than the soil isolates. It is interesting to note that this study was the first report on studying the distribution of HSL-degrading enzyme in actinomycetes as well as the first report on finding such an activity in endophytic actinomycetes. Endophytic Streptomyces LPC029 showed the highest C10HSL-degrading activity at 151.30 ± 3.1 nmole/h/mL and its enzyme showed broad substrate specificity against medium-to-long-chain HSLs. The enzyme was classified as HSL-acylase which hydrolyzes an amide bond between an acyl-side chain and a homoserine lactone and releases a free HSL. The partially purified enzyme from LPC029 attenuated soft-rot diseases caused by Pcc, which made the strain as well as many other QQ actinomycetes in this study potential candidates for biocontrol against QS-dependent phytopathogens. Further study on growth conditions including nutrients, temperature, pH, and aerations is required to maximize enzyme productivity in the selected strain(s).
Acknowledgments
This work was supported by the MRG5180031 Grant of the Thailand Research Fund (TRF) and the Commission on Higher Education (CHE) and supported in part by the Center of Excellence on Agricultural Biotechnology, Science and Technology Postgraduate Education and Research Development Office, Office of Higher Education Commission, Ministry of Education (AG-BIO/PERDO-CHE). The authors thank Dr. Tavan Janvilisri and Dr. Pahol Kosiyachinda for critical reading of the paper.
References
- 1.Chadwick D, Goode J. Antibiotic Resistance: Origins, Evolution, Selection, and Spread. New York, NY, USA: John Wiley and Sons; 1997. [Google Scholar]
- 2.Clatworthy AE, Pierson E, Hung DT. Targeting virulence: a new paradigm for antimicrobial therapy. Nature Chemical Biology. 2007;3(9):541–548. doi: 10.1038/nchembio.2007.24. [DOI] [PubMed] [Google Scholar]
- 3.Miller MB, Bassler BL. Quorum sensing in bacteria. Annual Review of Microbiology. 2001;55:165–199. doi: 10.1146/annurev.micro.55.1.165. [DOI] [PubMed] [Google Scholar]
- 4.Whitehead NA, Barnard AML, Slater H, Simpson NJL, Salmond GPC. Quorum-sensing in Gram-negative bacteria. FEMS Microbiology Reviews. 2006;25(4):365–404. doi: 10.1111/j.1574-6976.2001.tb00583.x. [DOI] [PubMed] [Google Scholar]
- 5.Winans SC. Command, control and communication in bacterial pathogenesis. Trends in Microbiology. 1998;6(10):382–383. doi: 10.1016/s0966-842x(98)01338-9. [DOI] [PubMed] [Google Scholar]
- 6.Dong YH, Xu JL, Li XZ, Zhang LH. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora . Proceedings of the National Academy of Sciences of the United States of America. 2000;97(7):3526–3531. doi: 10.1073/pnas.060023897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Morohoshi T, Nakazawa S, Ebata A, Kato N, Ikeda T. Identification and characterization of N-acylhomoserine lactone-acylase from the fish intestinal Shewanella sp. strain MIB015. Bioscience, Biotechnology and Biochemistry. 2008;72(7):1887–1893. doi: 10.1271/bbb.80139. [DOI] [PubMed] [Google Scholar]
- 8.Park SY, Kang HO, Jang HS, Lee JK, Koo BT, Yum DY. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Applied and Environmental Microbiology. 2005;71(5):2632–2641. doi: 10.1128/AEM.71.5.2632-2641.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Uroz S, Oger P, Chhabra SR, Cámara M, Williams P, Dessaux Y. N-acyl homoserine lactones are degraded via an amidolytic activity in Comamonas sp. strain D1. Archives of Microbiology. 2007;187(3):249–256. doi: 10.1007/s00203-006-0186-5. [DOI] [PubMed] [Google Scholar]
- 10.Hong KW, Koh CL, Sam CK, Yin WF, Chan KG. Quorum quenching revisited-from signal decays to signalling confusion. Sensors. 2012;12(4):4661–4696. doi: 10.3390/s120404661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Saadoun I, Rawashdeh R, Dayeh T, Ababneh Q, Mahasneh A. Isolation, characterization and screening for fiber hydrolytic enzymes-producing streptomycetes of Jordanian forest soils. Biotechnology. 2007;6(1):120–128. [Google Scholar]
- 12.Tan H, Deng Z, Cao L. Isolation and characterization of actinomycetes from healthy goat faeces. Letters in Applied Microbiology. 2009;49(2):248–253. doi: 10.1111/j.1472-765X.2009.02649.x. [DOI] [PubMed] [Google Scholar]
- 13.Bull AT, Goodfellow M, Slater JH. Biodiversity as a source of innovation in biotechnology. Annual Review of Microbiology. 1992;46:219–252. doi: 10.1146/annurev.mi.46.100192.001251. [DOI] [PubMed] [Google Scholar]
- 14.Anupama M, Narayana KJP, Vijayalakshmi M. Screening of Streptomyces purpeofuscus for antimicrobial metabolites. Research Journal of Microbiology. 2007;2(12):992–994. [Google Scholar]
- 15.Cha C, Gao P, Chen YC, Shaw PD, Farrand SK. Production of acyl-homoserine lactone quorum-sensing signals by gram-negative plant-associated bacteria. Molecular Plant-Microbe Interactions. 1998;11(11):1119–1129. doi: 10.1094/MPMI.1998.11.11.1119. [DOI] [PubMed] [Google Scholar]
- 16.McClean KH, Winson MK, Fish L, et al. Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology. 1997;143(12):3703–3711. doi: 10.1099/00221287-143-12-3703. [DOI] [PubMed] [Google Scholar]
- 17.Chilton MD, Currier TC, Farrand SK. Agrobacterium tumefaciens DNA and PS8 bacteriophage DNA not detected in crown gall tumors. Proceedings of the National Academy of Sciences of the United States of America. 1974;71(9):3672–3676. doi: 10.1073/pnas.71.9.3672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lane DJ. 16S/23S rRNA sequencing. In: Stackerbrand E, Goodfellow M, editors. Nucleic Acid Techniques in Bacterial Systematics. John Wiley and Sons; 1991. [Google Scholar]
- 19.Sanger F, Nicklen S, Coulson AR. DNA sequencing with chain-terminating inhibitors. Proceedings of the National Academy of Sciences of the United States of America. 1977;74(12):5463–5467. doi: 10.1073/pnas.74.12.5463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Burr T, Barnard AML, Corbett MJ, Pemberton CL, Simpson NJL, Salmond GPC. Identification of the central quorum sensing regulator of virulence in the enteric phytopathogen, Erwinia carotovora: The VirR repressor. Molecular Microbiology. 2006;59(1):113–125. doi: 10.1111/j.1365-2958.2005.04939.x. [DOI] [PubMed] [Google Scholar]
- 21.Holt JG, Krieg NR, Sneath PHA, Staley JT, Williams ST. Bergey's Manual of Determinative Bacteriology. Hagerstown, Md, USA: Lippincott Williams & Wilkins; 1994. [Google Scholar]
- 22.Lee SJ, Park SY, Lee JJ, Yum DY, Koo BT, Lee JK. Genes encoding the N-acyl homoserine lactone-degrading enzyme are widespread in many subspecies of Bacillus thuringiensis . Applied and Environmental Microbiology. 2002;68(8):3919–3924. doi: 10.1128/AEM.68.8.3919-3924.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Molina L, Constantinescu F, Michel L, Reimmann C, Duffy B, Défago G. Degradation of pathogen quorum-sensing molecules by soil bacteria: a preventive and curative biological control mechanism. FEMS Microbiology Ecology. 2003;45(1):71–81. doi: 10.1016/S0168-6496(03)00125-9. [DOI] [PubMed] [Google Scholar]
- 24.Strobel G. Harnessing endophytes for industrial microbiology. Current Opinion in Microbiology. 2006;9(3):240–244. doi: 10.1016/j.mib.2006.04.001. [DOI] [PubMed] [Google Scholar]
- 25.Leadbetter JR, Greenberg EP. Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus . Journal of Bacteriology. 2000;182(24):6921–6926. doi: 10.1128/jb.182.24.6921-6926.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dong YH, Zhang LH. Quorum sensing and quorum-quenching enzymes. Journal of Microbiology. 2005;43:101–109. [PubMed] [Google Scholar]
- 27.Rosenblueth M, Martínez-Romero E. Bacterial endophytes and their interactions with hosts. Molecular Plant-Microbe Interactions. 2006;19(8):827–837. doi: 10.1094/MPMI-19-0827. [DOI] [PubMed] [Google Scholar]
- 28.Ryan RP, Germaine K, Franks A, Ryan DJ, Dowling DN. Bacterial endophytes: recent developments and applications. FEMS Microbiology Letters. 2008;278(1):1–9. doi: 10.1111/j.1574-6968.2007.00918.x. [DOI] [PubMed] [Google Scholar]
- 29.Strobel G, Daisy B. Bioprospecting for microbial endophytes and their natural products. Microbiology and Molecular Biology Reviews. 2003;67(4):491–502. doi: 10.1128/MMBR.67.4.491-502.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Huang JJ, Petersen A, Whiteley M, Leadbetter JR. Identification of QuiP, the product of gene PA1032, as the second acyl-homoserine lactone acylase of Pseudomonas aeruginosa PAO1. Applied and Environmental Microbiology. 2006;72(2):1190–1197. doi: 10.1128/AEM.72.2.1190-1197.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Huang JJ, Han JI, Zhang LH, Leadbetter JR. Utilization of acyl-homoserine lactone quorum signals for growth by a soil Pseudomonad and Pseudomonas aeruginosa PAO1. Applied and Environmental Microbiology. 2003;69(10):5941–5949. doi: 10.1128/AEM.69.10.5941-5949.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shepherd RW, Lindow SE. Two dissimilar N-aeyl-homoserine lactone acylases of Pseudomonas syringae influence colony and biofilm Morphology. Applied and Environmental Microbiology. 2009;75(1):45–53. doi: 10.1128/AEM.01723-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chen CN, Chen CJ, Liao CT, Lee CY. A probable aculeacin A acylase from the Ralstonia solanacearum GMI1000 is N-acyl-homoserine lactone acylase with quorum-quenching activity. BMC Microbiology. 2009;9, article 89 doi: 10.1186/1471-2180-9-89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Romero M, Avendaño-Herrera R, Magariños B, Cámara M, Otero A. Acylhomoserine lactone production and degradation by the fish pathogen Tenacibaculum maritimum, a member of the Cytophaga-Flavobacterium-Bacteroides (CFB) group. FEMS Microbiology Letters. 2010;304(2):131–139. doi: 10.1111/j.1574-6968.2009.01889.x. [DOI] [PubMed] [Google Scholar]
- 35.Uroz S, Chhabra SR, Cámara M, Williams P, Oger P, Dessaux Y. N-acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology. 2005;151(10):3313–3322. doi: 10.1099/mic.0.27961-0. [DOI] [PubMed] [Google Scholar]
- 36.Romero M, Diggle SP, Heeb S, Cámara M, Otero A. Quorum quenching activity in Anabaena sp. PCC 7120: identification of AiiC, a novel AHL-acylase. FEMS Microbiology Letters. 2008;280(1):73–80. doi: 10.1111/j.1574-6968.2007.01046.x. [DOI] [PubMed] [Google Scholar]
- 37.Jafra S, Jalink H, Van Der Schoor R, Van Der Wolf JM. Pectobacterium carotovorum subsp. carotovorum strains show diversity in production of and response to N-acyl homoserine lactones. Journal of Phytopathology. 2006;154(11-12):729–739. [Google Scholar]
- 38.Sio CF, Otten LG, Cool RH, et al. Quorum quenching by an N-acyl-homoserine lactone acylase from Pseudomonas aeruginosa PAO1. Infection and Immunity. 2006;74(3):1673–1682. doi: 10.1128/IAI.74.3.1673-1682.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ravn Flodgaard L, Christensen AB, Molin S, Givskov M, Gram L. Influence of food preservation parameters and associated microbiota on production rate, profile and stability of acylated homoserine lactones from food-derived Enterobacteriaceae. International Journal of Food Microbiology. 2003;84(2):145–156. doi: 10.1016/s0168-1605(02)00405-1. [DOI] [PubMed] [Google Scholar]
- 40.Aguilar C, Bertani I, Venturi V. Quorum-sensing system and stationary-phase sigma factor (rpoS) of the onion pathogen Burkholderia cepacia genomovar I type strain, ATCC 25416. Applied and Environmental Microbiology. 2003;69(3):1739–1747. doi: 10.1128/AEM.69.3.1739-1747.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dong YH, Wang LH, Xu JL, Zhang HB, Zhang XF, Zhang LH. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature. 2001;411(6839):813–817. doi: 10.1038/35081101. [DOI] [PubMed] [Google Scholar]
- 42.Dong YH, Zhang XF, Xu JL, Zhang LH. Insecticidal Bacillus thuringiensis Silences Erwinia carotovora virulence by a new form of microbial antagonism, signal interference. Applied and Environmental Microbiology. 2004;70(2):954–960. doi: 10.1128/AEM.70.2.954-960.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hjelmgaard T, Persson T, Rasmussen TB, Givskov M, Nielsen J. Synthesis of furanone-based natural product analogues with quorum sensing antagonist activity. Bioorganic and Medicinal Chemistry. 2003;11(15):3261–3271. doi: 10.1016/s0968-0896(03)00295-5. [DOI] [PubMed] [Google Scholar]
- 44.Xu F, Byun T, Deussen HJ, Duke KR. Degradation of N-acylhomoserine lactones, the bacterial quorum-sensing molecules, by acylase. Journal of Biotechnology. 2003;101(1):89–96. doi: 10.1016/s0168-1656(02)00305-x. [DOI] [PubMed] [Google Scholar]