

Background: Collagen macromolecules are biologically relevant substrates in tissue remodeling and bone-related diseases.
Results: We investigated the action of cysteine proteases on the structural integrity and mechanical functionality of collagen fibers.
Conclusion: Using ultrastructural and biochemical techniques, we present a model of collagen fiber degradation via cathepsin K.
Significance: Our data provide new insights in matrix degradation and may allow new strategies to inhibit it.
Keywords: Atomic Force Microscopy, Collagen, Cysteine Protease, Electron Microscopy (EM), Glycosaminoglycan, Mechanical Properties
Abstract
Excessive cathepsin K (catK)-mediated turnover of fibrillar type I and II collagens in bone and cartilage leads to osteoporosis and osteoarthritis. However, little is known about how catK degrades compact collagen macromolecules. The present study is aimed to explore the structural and mechanical consequences of collagen fiber degradation by catK. Mouse tail type I collagen fibers were incubated with either catK or non-collagenase cathepsins. Methods used include scanning electron microscopy, protein electrophoresis, atomic force microscopy, and tensile strength testing. Our study revealed evidence of proteoglycan network degradation, followed by the progressive disassembly of macroscopic collagen fibers into primary structural elements by catK. Proteolytically released GAGs are involved in the generation of collagenolytically active catK-GAG complexes as shown by AFM. In addition to their structural disintegration, a decrease in the tensile properties of fibers was observed due to the action of catK. The Young's moduli of untreated collagen fibers versus catK-treated fibers in dehydrated conditions were 3.2 ± 0.68 GPa and 1.9 ± 0.65 GPa, respectively. In contrast, cathepsin L, V, B, and S revealed no collagenase activity, except the disruption of proteoglycan-GAG interfibrillar bridges, which slightly decreased the tensile strength of fibers.
Introduction
Fibrillar collagens are the crucial architectural and mechanical support elements of mammalian connective tissues such as bones, cartilage, and tendons (1, 2). Significant efforts have been made to elucidate the fine structure of collagen fibers. Its smallest unit is the triple helical collagen consisting of three intertwined α-chains (two α1(I) and one α2(I) in type I collagen and three α1(II) in type II collagen (3, 4). Recently, attention has been given to the molecular packing of collagen molecules into microfibrils (5) and their self-organization into larger fibrils. The elucidation of these substructures is based on a variety of methods including x-ray diffraction, electron tomography, electron density mapping, electron microscopy, and atomic force microscopy (6–10). Furthermore, some studies have suggested that the mutual interactions of these collagen fibrils within a fiber depends on proteoglycan-GAG2 interfibrillar bridges (11), and these proteoglycans are also essential in collagen fibrillogenesis. Proteoglycans consist of leucine-rich repeat core proteins such as biglycans, decorins, and fibromodulins with covalently attached glycosaminoglycan chains of dermatan sulfate (DS) (12, 13), keratan sulfate (11), or chondroitin sulfate (14). These GAG chains are associated with nearby GAG chains through electrostatic interactions and build bridges between proteoglycan core proteins present on the collagen fibril surface (15). Several studies have shown that polysaccharide chains of these interfibrillar bridges are responsible for the mechanical integrity between collagen fibrils in connective tissues (16, 17). The excellent mechanical properties of collagen fibers at different hierarchical levels have been studied for many years using advance techniques such as direct tensile tests, bending tests, AFM force spectrometry, nanoscale indentation methods, etc. (18–23). However, the action of cysteine proteases such as catK on the structural and mechanical functionality of these macromolecules is less clear.
Significant progress has been made to understand the degradation of triple helical collagen molecules by collagenases (24–28), but little is known about the mechanism of their action on collagen fibers. One of the most effective collagenases is catK, a papain-like cysteine protease highly expressed in osteoclasts. catK is responsible for the bulk collagen degradation in physiological and pathological bone remodeling and thus a key pharmaceutical target for the development of anti-osteoporotic (29) and anti-arthritic drugs (30). In this study, we report the in vitro collagen fiber degradation by catK. Using SEM and AFM, we present a model of collagen fiber degradation by this protease. Time course studies of the degradation of collagen fibers and the proteolytic release of α-chains from tropocollagen molecules reveal the simultaneous progress of both these processes at the same time. Moreover, the effect of catK activity on collagen fibers is compared with the action of non-collagenolytic cathepsins and the consequences of cathepsin exposure on the mechanical strength and physical properties of fibers are evaluated.
EXPERIMENTAL PROCEDURES
Materials
All chemicals and solvents used in the present study were of analytical grade. C4-S, DS, dithiothreitol (DTT), and EDTA were purchased from Sigma. For the in vitro collagen fiber degradation assay, 100 mm sodium acetate buffer (pH 5.5) containing 2.5 mm DTT and 2.5 mm EDTA was used. Z-Phe-Arg-MCA was purchased from Bachem (Weil am Rhein, Germany). Glutaraldehyde was procured from Sigma and Milli-Q water was used for imaging experiments. Dimethylmethylene blue was purchased from Sigma.
Isolation of Collagen Fibers
Type I collagen fibers were isolated from tail tendons of 3-month-old C57BL/6 mice as described in Ref. 31. Briefly, bundles of white fibers were pulled out from the distal end of mouse tail using surgical clamps and collected in PBS. These fibers were sterilized with 70% ethanol, air-dried, and transferred to a sterile bottle for further use. Freshly isolated collagen fibers were used for the present experiments.
Proteases
Human cathepsins K, V, S, and L were expressed in Pichia pastoris using the pPIC9K vector (32, 33). Cathepsin proteins were purified by chromatography using N-butyl-Sepharose and SP-Sepharose (Amersham Biosciences) (34), and their active site concentrations were determined by E-64 titration as described previously (35). Recombinant human cathepsin B was kindly provided by Dr. J. Mort from the Shriner's Hospital for Sick Children (Montreal, QC, Canada).
In Vitro Collagen Fiber Degradation
Insoluble type I collagen fibers (1 mg) were incubated with wild type catK and non-collagenase cathepsins (L, V, B, or S) with each at 3 μm concentration in 100 mm sodium acetate buffer, pH 5.5, containing 2.5 mm DTT and EDTA for different time intervals (up to 20 h) at 28 °C. Digest experiments were performed in the absence and presence of 1.5 μm C4-S or DS to analyze the effect of external GAGs on the collagenolytic activity of catK. The reaction was stopped by the addition of 10 μm E-64 at respective time intervals. Subsequently, the reaction mixture was centrifuged for 20 min, and the supernatant was taken and subjected to SDS-PAGE analysis using 9% Tris/glycine gels. Bands were visualized by Coomassie Brilliant Blue R-250 staining and analyzed by the bioimaging system, SYNGENE. Prestained protein ladders (PAGE, Invitrogen) was used for size determination. The collagenase activity of these proteases was evaluated on the basis of the generation and loss of αI and α2 bands after SDS-PAGE.
Electron Microscopy Imaging and Measurements
Scanning electron microscopy was used to characterize collagen fibers before and after enzymatic treatment. Collagen fibers were incubated with wild type catK or non collagenase cathepsins under the conditions as described above. At different time points of the enzymatic digestion, reactions were stopped with E-64, and collagen fibers were separated, rinsed with water and fixed with 2.5% glutaraldehyde (pH 7.4) at room temperature and then rinsed several times with distilled water. Samples were dehydrated by transferring through increasing concentrations of ethanol. After passing through anhydrous ethanol, samples were transferred into a critical point dryer. Following the drying procedure, samples were mounted on a metal stub with double-sided carbon adhesive tape and coated with Au/Pd in Hummer VI Sputtering System (AnaTech, Union City, CA). Samples were imaged by Helios NanoLabTM 650 (FEI, Hillsboro, OR) scanning electron microscope, operated at 2–10 kV. Experiments were repeated several times to confirm the results, and samples were observed carefully without any beam damage. Micrographs were taken from different spots of the same sample at similar magnification, and width measurements were done using software provided with the Helios microscope.
Atomic Force Microscopy Observations
Further imaging studies were carried out to observe catK-GAG complexes and released products during the proteolytic degradation of collagen fibers using AFM (Cypher scanning probe microscope, Asylum Research, Santa Barbara, CA). Collagen fibers were incubated with catK to perform the collagenase assay as described above, and reaction supernatants were collected at different time points for AFM analysis. Reaction supernatants were deposited on freshly cleaved mica for 10 min, rinsed with distilled water, and dried using a stream of nitrogen. Imaging was done in air using tapping mode and images (512 × 512 pixel scans) were acquired at a scanning rate of 3 Hz. Silicon tips (Model- AC160TS, Asylum Research) with a radius of 7 nm were used to record the images at resonance frequency of 300 kHz and spring constant of 42 n/m. The background that corresponds to the mica surface in the AFM images was corrected using the first-order flattening function of the Asylum Research 101010 + 1901 macro working with Igor Pro (version 6.22A; Wavemetrics, Inc., Portland, OR) below a threshold set at ∼100–300 pm. Section analyses revealing the size of the proteins or protein complexes were done using the same software.
Weight Determination
Degradation effect of cathepsins (K, L, V, B, and S) on collagen fibers was also interpreted in the form of weight loss. Collagen fibers were treated with different cathepsins, and their mass loss due to enzymatic digestion was measured over sequential time points (0–20 h) using Mettler Toledo AG285 analytical balance. Fibers were isolated from the reaction mixtures and washed with milli-Q water and dried in vacuum. Analyses of numerical data were performed using statistical software and presented as mean ± S.D.
Dimethylmethylene Blue Assay for Quantitative GAG Determination
Collagen fibers were incubated with catK and non-collagenase cathepsins as per given protocol, and reaction supernatant was collected at different time points to quantify the released GAGs. Dimethylmethylene blue solution was prepared by dissolving 16 mg of dye in 1 liter of water with 2.37 g of NaCl, 3.04 g of glycine, and 95 ml of 0.1 m HCl, and assay was performed according to the manufacturer's instructions (36). Absorbance was measured at 525 nm using a microtiter plate reader (Spectra Max 190 software; Soft Max Pro; version 5.2). The concentration of sulfated GAGs was determined by a C4-S standard curve.
Tensile Strength Testing of Cathepsin-treated Collagen Fibers
Collagen fibers (n = 8 to 14) of similar diameter (45.5 ± 6.5 μm) were treated with different cathepsins and evaluated for their effects on the mechanical properties of fibers using tensile testing method in dehydrated condition (37). Both SEM and optical microscopy were used to determine the diameters of fibers before and after enzymatic treatments for micromechanical testing. The mechanical properties (Young's modulus, tensile strength, and ultimate strain) of single fibers were obtained from uniaxial tensile tests. A gauge length of 10 mm was selected in these experiments for 30-mm-long collagen fibers. Before testing, the fibers were fixed on a frame, and their diameters were determined from the average of microscopic measurements from different spots along the length of fiber. Then, the frame was clamped on a universal MTS Synergie RT100 type tensile machine equipped with a 50 n capacity load cell, and the edges of the frame are cut away. The fibers were stretched to failure at a constant crosshead displacement of 5 mm/min. The mechanical properties were determined in accordance with the Norme Française (NF) 25-704 standard, which takes into account the compliance of the loading frame. The Young's moduli were calculated on the linear part of the stress-strain curve.
RESULTS
Control Collagen Fibers
SEM analysis of untreated fibers demonstrated average diameters of 45.5 ± 6.5 μm (Fig. 1A). At higher magnification, collagen fibers showed a parallel arrangement of fibrils (∼200 nm) with a typical d-banding pattern, fused together with proteoglycan-GAG interfibrillar bridges (Fig. 2A). Random areas of control fibers were imaged over different time intervals of incubation in cathepsin activity buffer at pH 5.5 for up to 20 h, which revealed no changes in the fiber structures and minor increases in diameters. Our results are similar to those of previously reported structural SEM-based collagen fibril studies (38, 39). SDS-PAGE analysis of supernatants of fiber incubation mixtures did not show any Coomassie-positive fragments, indicating the intactness of the collagen fibers (see also Fig. 7A, control lane).
FIGURE 1.

Progressive dissociation of type I collagen fibers by catK. SEM micrographs of: untreated collagen fiber (A), fibers incubated with catK for 2 h (B), 10 h (C), and 12 to 14 h (D), showing the degradation of mouse tail collagen fibers (45.5 ± 6.5 μm) by 3 μm catK at pH 5.5 and 28 °C. The intact fiber is degraded in a stepwise process into fibril bundles (3.5 ± 1.5 μm) and fibrils (∼70–200 nm) within 14 h. Bars represent 30 μm.
FIGURE 2.

Degradation of different subhierarchical structures of type I collagen fiber by catK. A, scanning electron micrographs show the parallel arrangement of fibrils in an untreated collagen fiber, connected through interfibrillar proteoglycan-GAG cross-links. B, removal of these surface proteoglycan-GAG bridges after a 1-h incubation by catK. C, dissociation of collagen fiber into fibril bundles (3.5 ± 1.5 μm) can be seen at 4 h of catK treatment (bar, 5 μm). D and E, further dissociation of fibril bundles into fibrils having diameters between ∼70–200 nm but still displaying the d-periodicity at 7 h (D) and 12 h (E) of incubation. After 14 h, fibrils further decreased their diameters and lost their d-periodicity. F, unfolding of collagen fibrils. Bars represent 2 μm.
FIGURE 7.

Cathepsin-mediated degradation of collagen fibers. A, comparative SDS-PAGE of collagen fiber degradation products after incubation with activity buffer (C, control), catK, cathepsin L, cathepsin V, cathepsin B, cathepsin S, and catK+NaCl (enzyme concentration, 3 μm) at pH 5.5 and 28 °C for 4 h, respectively. Only catK is able to release significant amounts of tropocollagen fragments from insoluble collagen fibers. B, diameter analysis of collagen fibers after incubation with activity buffer (n = 14), catK, non-collagenase cathepsins (cathepsins L, V, B, and S), and catK+NaCl (3 μm enzyme concentration), respectively, at different time points (0 h, 2 h, and 20 h) (n = 8). C, mass analysis of collagen fibers at different points (0–20 h) of incubation with activity buffer, catK, or non-collagenase cathepsins. As shown, incubation of collagen fibers with activity buffer in the absence of cathepsins does not affect fiber diameters and their appearance, and no α-bands are observed in SDS-PAGE. Note that K+NaCl represents the catK+NaCl.
Degradation of Insoluble Type I Collagen Fibers by catK
In this study, we investigated the time-dependent progressive disintegration of collagen fibers in the presence of catK by high-resolution SEM incubation of collagen fibers with recombinant human catK revealed a dramatic degradation resulting in the disruption of the arrangement of fibrils within a fiber as shown in Fig. 1, A–D. At higher magnification, significant structural changes were observed at 1-h postincubation with catK as the fiber surface became irregular when compared with control specimens. Fibril bundles were not tightly packed, and the fiber surface displayed the loss of proteoglycan-GAG bridges between fibrils (Fig. 2B). Examination of 4-h catK-treated samples showed the splitting of collagen fibers (diameter, 45.5 ± 6.5 μm) into small fibril bundles (diameter, 3.5 ± 1.5 μm) (Fig. 2C), which with increasing incubation time further dissociated into fibrils (diameter ∼ 70–200 nm) (Fig. 2, D and E). Intact collagen fibers disappeared after incubation with catK within 14 h, but different subhierarchical structures of the fibers were still present. SEM analysis of 14–17 h specimens revealed the further degradation of fibrils, and subsequently, these fibrils lost their d-periodicity due to the excessive unfolding of microfibrils (Fig. 2F). After 17 h, any remaining fibrillar structure elements lost their structural integrity and were completely dissolved or disappeared within 20 h.
Fig. 3A demonstrates the degradation of collagen fibers assessed by SDS-PAGE where the visibility of α-bands represents the collagenase activity of wild type catK. αI and α2 band-sized degradation products reached their maximum between 4 and 7 h of incubation time and then gradually decreased. This implies that catK degrades solubilized tropocollagen fragments into low molecular weight fragments as we have previously demonstrated for soluble collagen preparations (24, 25). After 20 h of incubation with catK, no distinguishable collagen fragments were visible. Thus, throughout the degradation of collagen fibers, both processes, the unfolding of fibrils and microfibrils and the degradation of triple helical collagen molecules, occurred simultaneously.
FIGURE 3.

A, representative SDS-PAGE analysis of collagen fiber degradation products (α1 and α2-chains) after incubation with catK at different time points up to 20 h. α-Chains reach a maximum between 4–7 h and are subsequently degraded. B, quantitative analysis of released GAGs in digest mixture by catK, non-collagenase cathepsins (catL, -V, -B, -S) and catK + NaCl at different time points (0 h, 4 h, 10 h, and 20 h), compared with control fibers incubated with activity buffer in the absence of cathepsins.
Quantitative Analysis of Released Sulfated GAGs
Spectrophotometric GAG determination in collagen fiber digest mixtures revealed a gradual increase in soluble GAGs by catK activity reaching 12.2 ± 2.3 μg after 0.5 h and up to 22.0 ± 1.4 μg (∼1.1 μm) after 20 h in the supernatants. This amount of GAGs is sufficient to allow collagen degradation by catK-GAG complexes as demonstrated previously (40). In contrast, soluble GAGs released from undigested collagen fibers reached a maximum of less than 2.0 ± 0.08 μg after 20 h of incubation in the acidic reaction buffer. However, the activity of non-collagenolytic cathepsins also caused the release of soluble GAGs, which ranged from 3.1 ± 1.4 μg for catV to 5.9 ± 0.96 μg, 6.5 ± 4.6 μg, 7.9 ± 1.8 μg for catS, catB, and catL after 20 h of incubation at pH 5.5. Interestingly, fibers treated with catK in the presence of NaCl only revealed 7.5 ± 1.2 μg of released GAGs after 20 h, which was in the range of non-collagenolytic cathepsins. This indicates that the degradation of proteoglycans and GAG release by catK is not the result of its collagenolytic activity. Other non-collagenolytic cathepsins can do the same as shown in Fig. 3B but with an overall lesser efficacy. Non-collagenolytic cathepsins can only cleave proteoglycan/GAG interactions located on the surface of fibers or otherwise accessible, whereas catK will be able to reach cryptic proteoglycans due to its collagenolytic activity. The collagenase-dependent dissociation of collagen fibers into fibrils will likely make more proteoglycans available for degradation and thus would explain the increased GAG release by catK. The concentrations of released GAGs by catK and other cathepsins at different time points are shown in Fig. 3B.
Atomic Force Microscopy Analysis of catK-mediated Fiber Degradation Products
The release of GAGs from collagen fibers by cathepsins and in particular by catK suggests that the GAGs required for catK-GAG complex formation are provided by cathepsin activity. AFM scanning analysis revealed that these polysaccharide chains form complexes with catK. Supernatants of collagen fiber incubation mixtures were taken prior to the addition of catK, immediately after the addition of catK, after 30 min, 4 h, and 7 h, and subjected to AFM analysis. The supernatant of collagen fibers alone did not show any discernable structural entities in the micrographs. After the addition of catK, numerous small particles were observed, which may represent the presence of monomeric catK molecules (Fig. 4A). Considering the sample dehydration effect, standard curves were constructed using reference proteins of different sizes.3 According to these standard curves, it was determined that the apparent height of monomeric catK particles was 4.3 ± 1.7 nm, which is very similar to the observed catK size in the crystal structure. Microscopic observation of catK-treated collagen fibers exhibited the loss of surface interactions within 1 h and thus the probable release of GAGs. This was further corroborated by the finding of increased levels of GAGs in catK-treated collagen fibers (Fig. 3B). After 30 min of incubation with catK, aggregates of potentially several catK molecules and GAGs were observed with apparent heights of 5.5 ± 1.1 nm (Fig. 4B). This minor height difference is expected due to interaction between catK and GAG strands forming larger size globular complexes. Similar structures (Fig. 4B, arrow) were observed when catK and C4-S were incubated alone (Fig. 4C). In contrast, catK in the absence of C4-S provided single dot images as seen in Fig. 4A (data not shown). After 4 h, fibrillar materials associated with globular aggregates appeared, and after 7 h, large amount of fibrillar materials of different heights were visible (Fig. 4, D and E). These fibrils likely correspond to tropocollagen aggregates and released fibrillar substructures accumulated in the digest mixtures, which are subsequently degraded by catK-GAG complexes. SDS-PAGE analysis of the solubilized cleavage products revealed tropocollagen-sized α-chain-related bands of 100–130 kDa. Thus, AFM images at 7 h represent released microfibrillar fragments but may also be interpreted as partially refibrilized soluble tropocollagen fragments during sample preparation.
FIGURE 4.
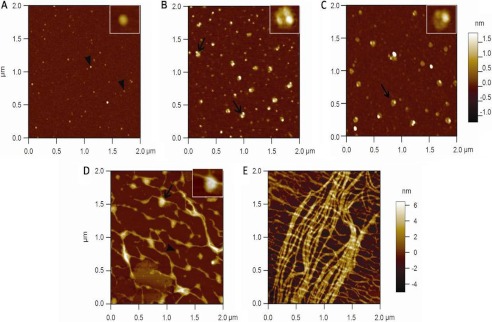
AFM analysis of soluble fractions of type I collagen fiber degradation by catK. AFM images of supernatants of catK/type I collagen digests at 0-h incubation ((A) only catK molecules of 4.3 ± 1.7 nm height are visible; catK in buffer alone gives identical images (not shown)), 0.5-h incubation ((B) CatK-GAG complexes of 5.5 ± 1.1 nm height are present), and catK/collagen digests at 4-h incubation (D), and 7 h ((E) between 4–7 h of increasing amounts of fibrillar material of different heights appears). C, similar catK-GAG complexes as B (here, catK and C4-S were incubated in the absence of collagen at a catK:GAG ratio similar to those present in the digestion supernatant after 30 min; 3 μm catK:30 μg/ml C4-S). The upper right insets are electronic magnifications of catK and CatK-GAG complexes. The colored bars on the right in the upper and lower panels represent the height scale of A–E, respectively. Diameter measurements were done by taking the average sample heights from different cross-sections. The black arrowheads point to catK molecules, and black arrows point to the catK-GAG complexes.
Degradation of Type I Collagen Fibers by catK in the Presence of Additional GAGs and NaCl
Type I collagen fibers were separately incubated with catK-C4-S and catK-DS mixtures (3 μm catK + 30 μg/ml) at 28 °C to determine the effects of externally applied GAGs on fibers degradation. Scanning microscopy analysis and in vitro digest data revealed no significant differences in the degradation level of collagen fibers in the presence or absence of external GAGs (Fig. 5, A–D). As the degradation of collagen-associated proteoglycans by catK releases sufficient amounts of GAGs, no externally added GAGs are required for the degradation of native collagen fibers.
FIGURE 5.

Degradation of type I collagen fibers by catK in the presence of C4-S. Shown are SEM micrographs of untreated collagen fiber (A), fibers incubated with catK/C4-S for 4 h (B), and 10 h (C), showing the degradation of collagen fibers (45.5 ± 6.5 μm). The catK concentration is 3 μm in presence of 30 μg/ml C4-S at pH 5.5 and 28 °C. Bars represent 30 μm. The structural degradation of collagen fiber shows a dramatic increase in diameter after 2 h, and their dissociation into fibril bundles after 4 h is followed by their dissociation into subfibrillar structures. Between 7–14 h, the degradation of fibrils was clearly visible. After 14 h, compact fibers disappeared as shown in catK degradation in the absence of external GAGs. In conclusion, the degradation of collagen fiber is similar in both cases, in the absence or presence of endogenous C4-S. The released GAGs from collagen fibers by catK are sufficient for the formation of catK-GAG complexes to degrade the compact collagen fiber and fibrils. D, SDS-PAGE analysis of collagen fiber degradation products (α1- and α2-chains) after incubation with 3 μm catK in the presence of 30 μg/ml C4-S at different time points up to 20 h. The time-dependent degradation pattern of α-bands is similar to that observed in the absence of additional GAGs. Experiments performed with catK/DS mixtures showed identical results (not shown).
Adding 300 mm NaCl to the digest completely inhibited the dissociation of the collagen fiber into smaller fibrils but did not block the removal of GAGs. After 20 h of incubation, the collagen fibers stayed intact but showed an ∼53% increase in fiber diameter (Fig. 6E) and the release of GAGs (7.5 ± 1.2 μg) into the digest mixture. This indicates that electrostatic interactions are needed for the degradation of collagen but not for the degradation of collagen-associated proteoglycans. This is further supported by the finding that in the presence of 300 mm NaCl no α-band-sized fragments were observed in SDS-PAGE analysis (Fig. 7A, catK + NaCl lane). As NaCl at this concentration does not have a significant effect on the kinetic parameters of the hydrolysis of a fluorogenic peptide substrate or on the degradation of gelatin (denatured collagen) but prevents the complex formation with GAGs, it can be concluded that monomeric catK cleaves proteoglycans but does not cleave fibrillar collagens as previously shown for the degradation of soluble triple helical collagen (40).
FIGURE 6.

Collagen fiber degradation by non-collagenolytic cathepsins. SEM micrographs of collagen fibers after 20 h Incubation with cathepsin L (A), cathepsin V (B), cathepsin B (C), cathepsin S (D), and catK+NaCl ((E) 3 μm enzyme concentrations) at pH 5.5 and room temperature. Left panel demonstrates the morphology of intact collagen fibers after 20-h cathepsin treatment; bars represent 30 μm. Right panel shows the magnified surface view of fibers after 20-h incubation with cathepsins L, V, B, S, and catK+NaCl. The images clearly demonstrate the arrangement of fibrils after enzymatic action and removal of collagen fiber-associated proteoglycans. Bars represent 2 μm. CatK in the presence of 300 mm NaCl and cathepsin L remove proteoglycans and partially degrade the collagen fiber at its surface. Note the collagen fibers incubated with activity buffer in the absence of cathepsins reveal no structural changes after 20 h.
Degradation of Insoluble Type I Collagen Fibers by Non-collagenolytic Cathepsins
SEM-based analysis of the collagen fibers treated with different cathepsins revealed that cathepsins B, S, and V only removed proteoglycan-GAG bridges between fibrils (Fig. 6, A–D). SDS-PAGE data confirmed their lack of collagenolytic activity. No collagen α-bands were observed. In contrast to cathepsins B, S, and V, cathepsin L activity displayed the removal of the proteoglycans and a partial liberation of α-band-sized fragments (Fig. 7A). SDS-PAGE analysis further revealed the presence of very low amounts of lower molecular weight sized α-chain-sized degradation products, indicating a very weak collagenase activity. However, this should be carefully interpreted as this could be a consequence of the degradation of partially denatured tropocollagen molecules within the fiber accessible for degradation by cathepsin L.
Effects of Cathepsins on Collagen Fiber Diameter and Weight
There were minor changes in the diameters and weights of collagen fibers when incubated with activity buffer in the absence of enzymes for up to 20 h. In contrast, incubation of fibers with catK showed initial increases of fiber diameters from 70–100% after 2 h due to subsequent loss of fiber integrity. The complete disappearance of fiber structure occurred after 14 h. The initial increase in fiber diameter is likely attributed to the loss of interfibrillar GAG bridges, which may lead to a loosening of the fiber structure. Less dramatic increases in fiber diameter were observed by the action of non-collagenase cathepsins. Scanning microscopy measurements showed ∼62% enhancement in diameter after 20 h of incubation with cathepsin L, ∼45% with cathepsin V or cathepsin B, ∼59% with cathepsin S, and ∼53% with catK+NaCl (Fig. 7B). Weight determinations of digested collagen fibers showed a 100% loss in mass after 14 h by catK, demonstrating the complete digest of the fiber material. Cathepsin L and catK + NaCl treated samples showed a ∼35% weight losses and cathepsins B-, S-, and V-treated samples about a 20–27% weight loss after 20 h of incubation (Fig. 7C). Here, the weight loss is primarily related to the degradation of proteoglycans and in the case of cathepsin L, a partial loss of collagen is also likely as shown by the release of small amounts of α-chain-related degradation products into the supernatants of the degradation mixture.
Effects of Cysteine Proteases on Mechanical Properties of Collagen Fiber
The mechanical properties of collagen fibers vary with different enzymatic treatments. The stress-strain curves obtained from the tensile test of control fibers, catK, non-collagenase cathepsins (L, V, B, and S) and catK+NaCl-treated fibers are shown in Fig. 8A. The differences in the diameter of fibers due to the specific activity of these proteases (Fig. 7B) are the likely cause for the variation of the tensile strength properties of the fibers. The average Young's moduli for control fibers were 3.2 ± 0.68 GPa, catK-treated fibers were 1.9 ± 0.65 GPa and for non-collagenase cathepsin (L, V, B, and S)-treated fibers between 2.0 ± 0.42 GPa to 2.7 ± 0.76 GPa (Fig. 8B). However, control fibers had significantly higher ultimate tensile strength (2.13 ± 0.71 n) than catK-treated fibers (0.95 ± 0.55 N) and non-collagenase cathepsin-treated fibers. The tensile results are displayed in Table 1 and show that for control fibers, the stress at the break is slightly higher (650 ± 110 MPa) compared with catK (241 ± 121 MPa) and non-collagenase cathepsin treated fibers (381 ± 190 MPa to 556 ± 86 MPa). Both, stress at break and strain at failure were highest for control fibers and lowest for catK-treated fibers (Fig. 8C). The maximum stretching was observed in control fibers (39 ± 7%). On the other hand, catK-treated collagen fibers had a strain at failure of 19 ± 8%. Approximately 27 ± 4%, 25 ± 3%, 29 ± 6%, 31 ± 7%, and 31 ± 4% stretching of collagen fibers were observed after incubation with cathepsins L, V, B, S, and catK+NaCl, respectively. The strain rate of collagen fibers decreased as a result of proteolytic activity of non-collagenase cathepsins because these proteases rupture the proteoglycan-GAG bridges between fibrils, which affects the mechanical integrity between the fibrils of collagen fibers (16, 17).
FIGURE 8.

Mechanical strength measurements on collagen fibers after cathepsin treatment. A, typical stress-strain curves of control collagen fibers incubated with activity buffer in the absence of proteases for 20 h (n = 14). Fibers were incubated with non-collagenase cathepsins (L, V, B, S, or catK+NaCl) for 20 h (n = 8), and collagen fibers were incubated with catK (n = 8) for 2 h. Stress-strain curves were obtained from the displacement of 5 mm/min in dry conditions. B, graph showing the Young's moduli of control collagen fibers and cathepsins K, L, V, B, S, and catK+ NaCl-treated fibers. Young's moduli were calculated on the linear part of the stress-strain curve. The relative error in the tensile modulus of these fibers is due to the function of different cathepsins and changes in fiber diameter. C, graph showing the stress at break (■) and strain at break (▵) of control and cathepsin (K, L, V, B, S, and catK+NaCl)-treated collagen fibers. Note that incubation time for catK-treated fibers is 2 h because after 2 h, fibers lost their structural integrity, which interferes in diameter measurements. 2-h catK-treated fibers show weak mechanical properties compared with other cathepsin-treated and control fibers after 20 h.
TABLE 1.
Mechanical properties of control and cathepsin-treated collagen fibers
| Fiber treatment | Diameter ± S.D.a | Strength ± S.D.b | Young's modulus ± S.D. | Stress at break ± S.D. | Strain at failure ± S.D.c |
|---|---|---|---|---|---|
| μm | n | GPa | MPa | % | |
| Control fibers (n = 14) | 60.03 ± 15.56 | 2.13 ± 0.71 | 3.21 ± 0.68 | 650 ± 110 | 39 ± 7 |
| CatK-treated fibers (n = 8) | 81.31 ± 22.71 | 0.95 ± 0.55 | 1.93 ± 0.65 | 241 ± 121 | 19 ± 8 |
| CatL-treated fibers (n = 8) | 76.02 ± 17.22 | 1.79 ± 0.77 | 1.77 ± 0.62 | 381 ± 190 | 27 ± 4 |
| CatV-treated fibers (n = 8) | 70.09 ± 18.25 | 1.89 ± 0.63 | 2.29 ± 0.59 | 478 ± 189 | 25 ± 3 |
| CatB-treated fibers (n = 7) | 65.95 ± 7. 59 | 1.54 ± 0.44 | 2.12 ± 0.26 | 479 ± 142 | 29 ± 6 |
| CatS-treated fibers (n = 8) | 71.16 ± 16.50 | 2.01 ± 0.95 | 2.78 ± 0.76 | 556 ± 86 | 31 ± 7 |
| CatK+NaCl-treated fibers (n = 8) | 67.17 ± 14.12 | 1.99 ± 0.50 | 2.07 ± 0.42 | 512 ± 157 | 31 ± 4 |
a Average diameters of collagen fibers calculated from multiple spots on the same fiber in dry conditions.
b Ultimate force in Newton (N).
c Ultimate strain in percentage (%) representing the maximum stretching value till failure.
DISCUSSION
Model of Collagen Fiber Degradation by catK
Most studies related to collagenases use soluble collagens as substrate, although the biologically relevant substrates in tissue remodeling and diseases such as osteoporosis, arthritis, and fibroses are insoluble collagen fibers or fibrils. Only recently, the fine structure of collagen fibrils has been better understood (41). The morphology of collagen fibers is characterized by a regular arrangement of fibrils tightly bundled together through GAG-mediated proteoglycan interactions as shown in previous reports (15, 39, 42, 43). We and others (24) have previously demonstrated the unique collagenase activity of catK, which is able to cleave at multiple sites within the triple helical region of tropocollagen. But how is catK able to degrade collagen fibers so efficiently? Our microscopic analysis and in situ proteolytic digestion results of collagen fiber degradation by catK demonstrated that the dissociation of proteoglycans from the fibers occurred prior to the collagen degradation and that their removal will expose additional areas on the fiber surface for the collagenolytic attack (44). However, the removal of proteoglycans alone does not appear to be sufficient to disintegrate a macro fiber into smaller subfibrils, as cathepsin B and cathepsin V are unable to do so and cathepsin L is only able to do so in a limited manner (45, 46).
We have previously demonstrated that the collagenase activity of catK requires GAGs as cofactors (25). These GAGs are likely provided by the degradation of collagen-associated proteoglycans to form collagenolytically active catK-GAG complexes (47). In this report, we demonstrated that catK releases GAGs from collagen fibers, and this reaction correlates with the release of collagen α-chains from the fiber. NaCl, which has been previously shown to block soluble collagen degradation and complex formation of catK with GAGs (40), completely inhibits collagen fiber disassembly and degradation. Although our data are indicative that NaCl prevents collagen degradation by inhibiting the formation of collagenolytically active catK-GAG complexes, it should be noted that we cannot rule out an effect on the binding of catK to collagen by itself.
Our SEM data of catK-treated collagen fibers in the presence of NaCl also provide direct evidence that in the absence of catK-GAG complexes, monomeric catK still exerts its proteoglycan degradation capability but lacks it collagen-degrading activity. This suggests that catK-GAG complexes are only required for the degradation of collagen fibrils as previously demonstrated for soluble collagen fragments (25, 40). The stepwise degradation of collagen molecules from the surface of fibrils will allow access to cryptic proteoglycans within the macro fiber core, which will lead to the observed splitting of the fiber into smaller and smaller subfibrils and simultaneously to an increased GAG release by catK. As expected the progressive unfolding of the fibers made them less stable and thus more accessible for further degradation. Scheme 1 summarizes the proposed mechanism of collagen fiber degradation by catK.
SCHEME 1.

Schematic presentation of stepwise collagen fiber degradation by catK. White and blue thin strands are proteoglycan-GAG chains, single red dots are catK molecules, and aggregates of red dots are catK/GAG complexes.
Mechanical Properties of Collagen Fibers Influenced by Cathepsin Activity
Tendons are organized in a hierarchical order from tropocollagen, microfibrils, and fibrils to fibers (4, 5, 41). Previous work has demonstrated the mechanical characterizations of different hierarchical structures of collagen fibers (20, 21, 23, 48–50). The comparison of undigested control collagen fibers with cathepsin-treated fibers in dehydrated conditions clearly indicated that the orientation, tensile strength, and strain rate of collagen fibers changed due to the activity catK and non-collagenase cathepsins. As a consequence of cathepsin activity, fiber diameter increased, and Young's moduli and ultimately the tensile strength of collagen fibers decreased (Figs. 7B and 8B). Our micromechanical results suggest that stress-strain curves of control and cathepsin-treated collagen fibers exhibit both linear and non-linear regions, which are in agreement with previous studies (19, 21). We find that control collagen fibers incubated in activity buffer up to 20 h and analyzed in dehydrated conditions had smaller diameters (∼60 μm) and displayed a higher Young's modulus, ultimate strain, and strength. However, an ∼70% increase in diameter was observed after 2 h of incubation time, leading ultimately to the complete degradation of collagen fibers by catK. The stress at the break, strain at failure, Young's modulus, and ultimate tensile strength of catK-treated collagen fibers was comparatively lower than control and non-collagenase-treated fibers in dehydrated conditions (Table 1).
The above results suggest that the control collagen fiber appears to be more stable than catK and other non-collagenase cathepsin-treated fibers. We believe that the load-bearing mechanism between the collagen fibrils may disrupt due to these enzymes. Fibrillar collagens in combination with proteoglycans form a network to provide mechanical integrity to tissues (15–17). Proteoglycan/GAG interactions are mainly responsible to hold fibrils together within a collagen fiber and intermolecular cross-links in the telopeptide regions of triple helical collagen maintain the hierarchy of collagen molecules within fibrils (5). From our analysis, it is clear that non-collagenase cathepsins (L, V, S, and B) are only capable of destabilizing the fiber structure in a limited manner. Neither the proteoglycan degradation nor their previously described telopeptidase activity (45, 46) allows for the disintegration of collagen fibers as observed with catK. Therefore, the structures of collagen fibers are still intact after incubation with non-collagenase cathepsins up to 20 h but clearly displayed a decrease in the tensile strength of fibers (19). However, catK possesses both proteoglycan and triple helical collagen degradation abilities, which severely interfere with the structural integrity as well as the mechanical strength of collagen fibers.
In summary, we demonstrated the stepwise degradation of collagen fibers by catK. SEM experiments revealed disintegration of large collagen fibers into smaller fibril bundles and microfibrils and their final dissolution. This process is accompanied by the simultaneous degradation of collagen-associated proteoglycans, which provide soluble GAGs to form collagenolytically active catK-GAG complexes. AFM studies suggested the formation of defined catK-GAG complexes during the degradation process. Tensile strength studies showed the destabilizing effects of catK activity on the mechanical properties of collagen fibers. However, we also investigated that destruction of proteoglycan-collagen interactions by non-collagenase cathepsins (catL, V, B, and S). These proteases are capable of partially destabilizing collagen fibers by removing collagen-associated proteoglycans, which are reflected by the altered diameter of collagen fibers and decreased mechanical stability.
Acknowledgment
We are thankful to Antoine Kervoelen for kind help in the mechanical experiments and for the technology support of the Centre for High Throughput Phenogenomics.
This work was supported by Canadian Institutes of Health Research Grant MOP89974 and the Canada Research Chair award.
X. Du and D. Brömme, unpublished data.
- GAG
- glycosaminoglycan
- SEM
- scanning electron microscopy
- AFM
- atomic force microscopy
- GPa
- gigapascal
- Z
- benzyloxycarbonyl
- MCA
- 4-methyl-7-coumarylamide
- E-64
- carboxy-trans-2,3-epoxypropionyl-leucylamido-(4-guanidino)butane
- C4-S
- chondroitin 4-sulfate
- catK
- cathepsin K
- DS
- dermatan sulfate
- MPa
- megapascal.
REFERENCES
- 1. Prockop D. J., Kivirikko K. I. (1995) Collagens: molecular biology, diseases, and potentials for therapy. Annu. Rev. Biochem. 64, 403–434 [DOI] [PubMed] [Google Scholar]
- 2. Kadler K. E., Baldock C., Bella J., Boot-Handford R. P. (2007) Collagens at a glance. J. Cell Sci. 120, 1955–1958 [DOI] [PubMed] [Google Scholar]
- 3. Rich A., Crick F. H. (1961) The molecular structure of collagen. J. Mol. Biol. 3, 483–506 [DOI] [PubMed] [Google Scholar]
- 4. Okuyama K., Takayanagi M., Ashida T., Kakudo M. (1977) A New Structural Model for Collagen. Polymer J. 9, 341–343 [Google Scholar]
- 5. Orgel J. P., Irving T. C., Miller A., Wess T. J. (2006) Microfibrillar structure of type I collagen in situ. Proc. Natl. Acad. Sci. U.S.A. 103, 9001–9005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Orgel J. P., Miller A., Irving T. C., Fischetti R. F., Hammersley A. P., Wess T. J. (2001) The in situ supermolecular structure of type I collagen. Structure 9, 1061–1069 [DOI] [PubMed] [Google Scholar]
- 7. Zhang G., Ezura Y., Chervoneva I., Robinson P. S., Beason D. P., Carine E. T., Soslowsky L. J., Iozzo R. V., Birk D. E. (2006) Decorin regulates assembly of collagen fibrils and acquisition of biomechanical properties during tendon development. J. Cell Biochem. 98, 1436–1449 [DOI] [PubMed] [Google Scholar]
- 8. Raspanti M., Ottani V., Ruggeri A. (1989) Different architectures of the collagen fibril: morphological aspects and functional implications. Int. J. Biol. Macromol. 11, 367–371 [DOI] [PubMed] [Google Scholar]
- 9. Starborg T., Lu Y., Meadows R. S., Kadler K. E., Holmes D. F. (2008) Electron microscopy in cell-matrix research. Methods 45, 53–64 [DOI] [PubMed] [Google Scholar]
- 10. Baselt D. R., Revel J. P., Baldeschwieler J. D. (1993) Subfibrillar structure of type I collagen observed by atomic force microscopy. Biophysical Journal 65, 2644–2655 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Scott J. E. (1992) Supramolecular organization of extracellular matrix glycosaminoglycans, in vitro and in the tissues. FASEB J. 6, 2639–2645 [PubMed] [Google Scholar]
- 12. Redaelli A., Vesentini S., Soncini M., Vena P., Mantero S., Montevecchi F. M. (2003) Possible role of decorin glycosaminoglycans in fibril to fibril force transfer in relative mature tendons-a computational study from molecular to microstructural level. J. Biomech. 36, 1555–1569 [DOI] [PubMed] [Google Scholar]
- 13. Vesentini S., Redaelli A., Montevecchi F. M. (2005) Estimation of the binding force of the collagen molecule-decorin core protein complex in collagen fibril. J. Biomech. 38, 433–443 [DOI] [PubMed] [Google Scholar]
- 14. Rees S. G., Flannery C. R., Little C. B., Hughes C. E., Caterson B., Dent C. M. (2000) Catabolism of aggrecan, decorin and biglycan in tendon. Biochem. J. 350, 181–188 [PMC free article] [PubMed] [Google Scholar]
- 15. Scott J. E., Orford C. R., Hughes E. W. (1981) Proteoglycan-collagen arrangements in developing rat tail tendon. An electron microscopical and biochemical investigation. Biochem. J. 195, 573–581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Puxkandl R., Zizak I., Paris O., Keckes J., Tesch W., Bernstorff S., Purslow P., Fratzl P. (2002) Viscoelastic properties of collagen: synchrotron radiation investigations and structural model. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 357, 191–197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Liu X., Yeh M. L., Lewis J. L., Luo Z. P. (2005) Direct measurement of the rupture force of single pair of decorin interactions. Biochem. Biophys. Res. Commun. 338, 1342–1345 [DOI] [PubMed] [Google Scholar]
- 18. Gentleman E., Lay A. N., Dickerson D. A., Nauman E. A., Livesay G. A., Dee K. C. (2003) Mechanical characterization of collagen fibers and scaffolds for tissue engineering. Biomaterials 24, 3805–3813 [DOI] [PubMed] [Google Scholar]
- 19. Pins G. D., Christiansen D. L., Patel R., Silver F. H. (1997) Self-assembly of collagen fibers. Influence of fibrillar alignment and decorin on mechanical properties. Biophys. J. 73, 2164–2172 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Eppell S., Smith B., Kahn H., Ballarini R. (2006) Nano measurements with micro-devices: mechanical properties of hydrated collagen fibrils. J. Roy. Soc. Interface 3, 117–121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Silver F. H., Freeman J. W., Seehra G. P. (2003) Collagen self-assembly and the development of tendon mechanical properties. J. Biomech. 36, 1529–1553 [DOI] [PubMed] [Google Scholar]
- 22. Yang L., Fitié C. F., van der Werf K. O., Bennink M. L., Dijkstra P. J., Feijen J. (2008) Mechanical properties of single electrospun collagen type I fibers. Biomaterials 29, 955–962 [DOI] [PubMed] [Google Scholar]
- 23. van der Rijt J. A., van der Werf K. O., Bennink M. L., Dijkstra P. J., Feijen J. (2006) Micromechanical Testing of Individual Collagen Fibrils. Macromol. Biosci. 6, 697–702 [DOI] [PubMed] [Google Scholar]
- 24. Garnero P. (1998) The Collagenolytic Activity of Cathepsin K Is Unique among Mammalian Proteinases. J. Biol. Chem. 273, 32347–32352 [DOI] [PubMed] [Google Scholar]
- 25. Li Z., Yasuda Y., Li W., Bogyo M., Katz N., Gordon R. E., Fields G. B., Brömme D. (2004) Regulation of collagenase activities of human cathepsins by glycosaminoglycans. J. Biol. Chem. 279, 5470–5479 [DOI] [PubMed] [Google Scholar]
- 26. Bertini I., Fragai M., Luchinat C., Melikian M., Toccafondi M., Lauer J. L., Fields G. B. (2012) Structural Basis for Matrix Metalloproteinase 1-Catalyzed Collagenolysis. J. Am. Chem. Soc. 134, 2100–2110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Sarkar S. K., Marmer B., Goldberg G., Neuman K. C. (2012) Single-Molecule Tracking of Collagenase on Native Type I Collagen Fibrils Reveals Degradation Mechanism. Curr. Biol. 22, 1047–1056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Manka S. W., Carafoli F., Visse R., Bihan D., Raynal N., Farndale R. W., Murphy G., Enghild J. J., Hohenester E., Nagase H. (2012) Structural insights into triple-helical collagen cleavage by matrix metalloproteinase 1. Proc. Natl. Acad. Sci. U.S.A. 109, 12461–12466 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Yamashita D. S., Dodds R. A. (2000) Cathepsin K and the Design of Inhibitors of Cathepsin K. Curr. Pharm. Des. 6, 1–24 [DOI] [PubMed] [Google Scholar]
- 30. Yasuda Y., Kaleta J., Brömme D. (2005) The role of cathepsins in osteoporosis and arthritis: rationale for the design of new therapeutics. Adv. Drug Deliv. Rev. 57, 973–93 [DOI] [PubMed] [Google Scholar]
- 31. Rajan N., Habermehl J., Coté M. F., Doillon C. J., Mantovani D. (2006) Preparation of ready-to-use, storable and reconstituted type I collagen from rat tail tendon for tissue engineering applications. Nat. Protoc. 1, 2753–2758 [DOI] [PubMed] [Google Scholar]
- 32. Linnevers C. J., McGrath M. E., Armstrong R., Mistry F. R., Barnes M. G., Klaus J. L., Palmer J. T., Katz B. A., Brömme D. (1997) Expression of human cathepsin K in Pichia pastoris and preliminary crystallographic studies of an inhibitor complex. Protein Sci. 6, 919–921 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Brömme D., Li Z., Barnes M., Mehler E. (1999) Human cathepsin V functional expression, tissue distribution, electrostatic surface potential, enzymatic characterization, and chromosomal localization. Biochemistry 38, 2377–2385 [DOI] [PubMed] [Google Scholar]
- 34. Brömme D., Okamoto K., Wang B. B., Biroc S. (1996) Human cathepsin O2, a matrix protein-degrading cysteine protease expressed in osteoclasts. Functional expression of human cathepsin O2 in Spodoptera frugiperda and characterization of the enzyme. J. Biol. Chem. 271, 2126–2132 [DOI] [PubMed] [Google Scholar]
- 35. Barrett A. J., Kembhavi A. A., Brown M. A., Kirschke H., Knight C. G., Tamai M., Hanada K. (1982) l-trans-Epoxysuccinyl-leucylamido(4-guanidino)butane (E-64) and its analogues as inhibitors of cysteine proteinases including cathepsins B, H and L. Biochem. J. 201, 189–198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Farndale R. W., Sayers C. A., Barrett A. J. (1982) A direct spectrophotometric microassay for sulfated glycosaminoglycans in cartilage cultures. Connect. Tissue Res. 9, 247–248 [DOI] [PubMed] [Google Scholar]
- 37. Tan E. P., Ng S. Y., Lim C. T. (2005) Tensile testing of a single ultrafine polymeric fiber. Biomaterials 26, 1453–1456 [DOI] [PubMed] [Google Scholar]
- 38. Raspanti M., Viola M., Forlino A., Tenni R., Gruppi C., Tira M. E. (2008) Glycosaminoglycans show a specific periodic interaction with type I collagen fibrils. J. Struct. Biol. 164, 134–139 [DOI] [PubMed] [Google Scholar]
- 39. Orgel J. P., Eid A., Antipova O., Bella J., Scott J. E. (2009) Decorin Core Protein (Decoron) Shape Complements Collagen Fibril Surface Structure and Mediates Its Binding. PLoS One 4, e7028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Li Z., Hou W. S., Escalante-Torres C. R., Gelb B. D., Bromme D. (2002) Collagenase activity of cathepsin K depends on complex formation with chondroitin sulfate. J. Biol. Chem. 277, 28669–28676 [DOI] [PubMed] [Google Scholar]
- 41. Ottani V., Martini D., Franchi M., Ruggeri A., Raspanti M. (2002) Hierarchical structures in fibrillar collagens. Micron 33, 587–596 [DOI] [PubMed] [Google Scholar]
- 42. Provenzano P. P., Vanderby R., Jr. (2006) Collagen fibril morphology and organization: implications for force transmission in ligament and tendon. Matrix Biol. 25, 71–84 [DOI] [PubMed] [Google Scholar]
- 43. Paige M. F., Rainey J. K., Goh M. C. (2001) A study of fibrous long spacing collagen ultrastructure and assembly by atomic force microscopy. Micron 32, 341–353 [DOI] [PubMed] [Google Scholar]
- 44. Billinghurst R. C., Wu W., Ionescu M., Reiner A., Dahlberg L., Chen J., van Wart H., Poole A. R. (2000) Comparison of the degradation of type II collagen and proteoglycan in nasal and articular cartilages induced by interleukin-1 and the selective inhibition of type II collagen cleavage by collagenase. Arthritis Rheum. 43, 664–672 [DOI] [PubMed] [Google Scholar]
- 45. Burleigh M. C., Barrett A. J., Lazarus G. S. (1974) Cathepsin B1. A lysosomal enzyme that degrades native collagen. Biochem. J. 137, 387–398 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Kirschke H., Kembhavi A. A., Bohley P., Barrett A. J. (1982) Action of rat liver cathepsin L on collagen and other substrates. Biochem. J. 201, 367–372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Li Z., Hou W. S., Brömme D. (2000) Collagenolytic activity of cathepsin K is specifically modulated by cartilage-resident chondroitin sulfates. Biochemistry 39, 529–536 [DOI] [PubMed] [Google Scholar]
- 48. Fratzl P. (2008) Collagen: Structure and Mechanics, Springer, New York [Google Scholar]
- 49. Kato Y. P., Christiansen D. L., Hahn R. A., Shieh S. J., Goldstein J. D., Silver F. H. (1989) Mechanical properties of collagen fibres: a comparison of reconstituted and rat tail tendon fibres. Biomaterials 10, 38–42 [DOI] [PubMed] [Google Scholar]
- 50. Gupta H. S., Seto J., Wagermaier W., Zaslansky P., Boesecke P., Fratzl P. (2006) Cooperative deformation of mineral and collagen in bone at the nanoscale. Proc. Natl. Acad. Sci. 103, 17741–17746 [DOI] [PMC free article] [PubMed] [Google Scholar]