

Abstract
Ossified gastralia, clavicles and sternal ribs are known in a variety of reptilians, including dinosaurs. In sauropods, however, the identity of these bones is controversial. The peculiar shapes of these bones complicate their identification, which led to various differing interpretations in the past. Here we describe different elements from the chest region of diplodocids, found near Shell, Wyoming, USA. Five morphotypes are easily distinguishable: (A) elongated, relatively stout, curved elements with a spatulate and a bifurcate end resemble much the previously reported sauropod clavicles, but might actually represent interclavicles; (B) short, L-shaped elements, mostly preserved as a symmetrical pair, probably are the real clavicles, as indicated by new findings in diplodocids; (C) slender, rod-like bones with rugose ends are highly similar to elements identified as sauropod sternal ribs; (D) curved bones with wide, probably medial ends constitute the fourth morphotype, herein interpreted as gastralia; and (E) irregularly shaped elements, often with extended rugosities, are included into the fifth morphotype, tentatively identified as sternal ribs and/or intercostal elements. To our knowledge, the bones previously interpreted as sauropod clavicles were always found as single bones, which sheds doubt on the validity of their identification. Various lines of evidence presented herein suggest they might actually be interclavicles – which are single elements. This would be the first definitive evidence of interclavicles in dinosauromorphs. Previously supposed interclavicles in the early sauropodomorph Massospondylus or the theropods Oviraptor and Velociraptor were later reinterpreted as clavicles or furculae. Independent from their identification, the existence of the reported bones has both phylogenetic and functional significance. Their presence in non-neosauropod Eusauropoda and Flagellicaudata and probable absence in rebbachisaurs and Titanosauriformes shows a clear character polarity. This implicates that the ossification of these bones can be considered plesiomorphic for Sauropoda. The proposed presence of interclavicles in sauropods may give further support to a recent study, which finds a homology of the avian furcula with the interclavicle to be equally parsimonious to the traditional theory that furcula were formed by the fusion of the clavicles. Functional implications are the stabilizing of the chest region, which coincides with the development of elongated cervical and caudal vertebral columns or the use of the tail as defensive weapon. The loss of ossified chest bones coincides with more widely spaced limbs, and the evolution of a wide-gauge locomotor style.
Keywords: diplodocidae, furcula, interclavicle, pectoral girdle, sauropod dinosaurs
Introduction
The plesiomorphic composition of the chest region in early reptiles includes various elements of the pectoral girdle (scapula, suprascapula, pro- and metacoracoids, cleithrum, clavicle, and the interclavicle), the sternal apparatus (sternal plates and ribs) and other chest bones (gastralia; Romer, 1956; Vickaryous & Hall, 2006; see Table 1 for a summary of pectoral and sternal elements in Reptilia). The term chest bones is here informally used to include gastralia, sternal plates and ribs, clavicles, and interclavicles based on the topographic position, and independent from their embryological or evolutionary origin.
Table 1.
Bones present in the chest regions of tetrapods
| General shape and distinction | Paired /single. Number | Embryological origin (dermal / endochondral) | Articulations | Previously identified in Sauropoda | |
|---|---|---|---|---|---|
| Clavicles (Morphotype B) | Curved element | Paired, can fuse into one (furcula, questioned herein). One pair present | Dermal bone | Dorsal to coracoids (covering it sometimes), between cleithrum scapula, and interclavicle in basal reptiles. Attaches to the acromion | Yes |
| Interclavicles (Morphotype A) | Variable. Diamond shaped in early tetrapods, rod-like with or without transverse processes in crocodylians and lacertilians | Single (very rarely paired). Only one present | Dermal bone | Between the clavicles. Connects to the coracoid and/or sternal plates posteriorly. In rare cases (e.g. monotremes) attaching to acromion | No |
| Sternal Ribs(Morphotypes C and E) | Irregularly shaped, often rugose | Paired. Maximum one per dorsal rib, often less | Endochondral | Connects the sternal plates with the dorsal ribs or intercostal elements | Yes |
| Intercostal elements (Morphotype E?) | Irregularly shaped, often rugose | Paired. Maximum one per dorsal rib, often less | Endochondral | Connects sternal ribs with dorsal ribs | No |
| Gastralia (Morphotype D) | Slender, slightly curved bones. Medial elements with expanded medial end for articulation with corresponding gastralia. Anteriormost sometimes fused | Four elements per row (2 lateral, 2 medial). Up to 21 rows in large theropods | Dermal bone | Articulate among themselves and to dorsal ribs, as well as maybe the sternal apparatus | Yes |
| Cleithrum | Spoon-shaped | Paired. Only one pair present | Dermal bone | Capping scapulacoracoid, attaches to clavicles | No |
| Sternal plates | Shield-like: flat oval or reniform | Paired or single. One single or two mirrored elements | Endochondral | Articulate among themselves, sternal ribs attached to them sometimes touching the interclavicle anteriorly | Yes |
| Furcula | Wide V-shape | Single, rarely two paired elements | Dermal bone | With acromia laterally | No |
| Scapulacoracoid | Large, flat, subcircular acromion with a elongate, more narrow posterodorsal projection | Paired. One pair present | Endochondral | Dorsal and external to sternal apparatus, connected medially by cleithrum, clavicles, and interclavicle | Yes |
In Diapsida, the cleithrum was lost, and pro- and metacoracoids fused to form a single element (Vickaryous & Hall, 2006; Remes, 2008). The interclavicle is generally interpreted to have been lost in Dinosauriformes (Nesbitt, 2011). A supposed absence of clavicles was often used as synapomorphy of Dinosauria or even more inclusive clades, and as reason against the ancestry of this clade to birds (see Sereno, 1991; Novas, 1996; Yates & Vasconcelos, 2005). However, numerous reports of clavicles in various dinosaur clades imply that these elements are plesiomorphically present, and that it is mostly due to diagenetic or taphonomic reasons that they are not recovered (Yates & Vasconcelos, 2005; Remes, 2008). On the other hand, a recent study on the embryology of the avian furcula and the crocodylian interclavicle revealed that an interpretation of the furcula as derived interclavicle would be equally parsimonious as the traditional hypothesis, where the furcula represents the medially fused clavicles (Vickaryous & Hall, 2010). The suggestion of Vickaryous & Hall (2010) that all previously reported dinosaurian clavicles might actually be interclavicles would bring back the earlier stated hypothesis that the absence of clavicles would be synapomorphic for Dinosauriformes, paralleled by extant crocodylians. However, the evidence remains ambiguous, and especially Sauropodomorpha appear to have both ossified clavicles and interclavicles (von Huene, 1926; Yates & Vasconcelos, 2005; this study).
In the Sauropodomorpha, pectoral girdles usually only preserve the scapula and the coracoid, which in sauropods generally fuse during ontogeny (Ikejiri et al. 2005; Schwarz et al. 2007a,b; Remes, 2008). Putative clavicles are predicted to connect the scapulae dorsal to the coracoids but, until recently, no articulated pair had been reported in sauropods (Remes, 2008; but see Galiano & Albersdörfer, 2010 for an articulated specimen). Unpaired rod-like structures interpreted as clavicles were found in a variety of sauropod taxa, sometimes associated with the pectoral girdle (Dong & Tang, 1984; He et al. 1988; Zhang, 1988; Hatcher, 1901, 1903; Sereno et al. 1999; Harris, 2007; Remes et al. 2009). However, most of these identifications rely solely on the similarity to previously reported ‘clavicles’ – which themselves are not beyond doubt.
The sauropod scapulacoracoid articulates posteroventrally with the sternal plates, which are connected with the dorsal ribs through usually cartilaginous sternal ribs (Schwarz et al. 2007a; Remes, 2008; Hohn-Schulte, 2010). As they remain mostly cartilaginous, sternal ribs are very rarely preserved in the fossil record. The only reports are from the Apatosaurus excelsus holotype YPM 1980 (Marsh, 1896), a set of ribs associated with the holotype of A. louisae (Holland, 1915), a sternal rib fused to a sternal plate in a probable early macronarian (Tschopp & Mateus, 2012), and maybe from the holotype of Eobrontosaurus yahnahpin (Filla & Redman, 1994; Claessens, 2004). Filla & Redman (1994) initially interpreted these structures as gastralia, based on superficial similarity to gastralia in theropods, non-sauropod sauropodomorphs, and other archosaurs like crocodylians or sphenodonts. Gastralia were also reported from Gongxianosaurus shibeiensis and Jobaria tiguidensis (He et al. 1998; Sereno et al. 1999), but Claessens (2004) and Fechner (pers. comm., 2011), based on a detailed comparison with theropod and non-sauropod sauropodomorph gastralia, recently questioned this identification, and suggested them to be ossified sternal ribs instead. In short, the evolutionary and developmental origin of these elements is hotly debated, and the sauropod chest bones other than the scapulacoracoid remain poorly understood.
Herein we describe five morphotypes of bones from the chest region, and state their most probable identifications. This classification helps identifying disarticulated elements. It yields important new information on the proper reconstruction of the sauropod pectoral girdle.
Locality
The new material described herein comes exclusively from the Howe Quarry in the Bighorn Basin of Wyoming, USA (Fig. 1). Two periods of extensive excavation were conducted at this site. In 1934 Barnum Brown collected 3000–4000 bones at the Howe Quarry for the American Museum of Natural History, NY, USA (AMNH; Brown, 1935; Michelis, 2004), and in 1990 Hans-Jakob Siber reopened the site with a team of the Sauriermuseum Aathal, Switzerland (SMA) and excavated another 700–800 elements (Ayer, 2000; Michelis, 2004; Tschopp & Mateus, in press). Only one of the specimens found during these two periods has since been described in detail, and was identified as subadult specimen of a new diplodocine species (Tschopp & Mateus, in press). Brown (1935) tentatively identified the majority as Diplodocinae, except for some elements belonging to Apatosaurus or Camarasaurus (see also Michelis, 2004; Tschopp & Mateus, in press).
Fig. 1.
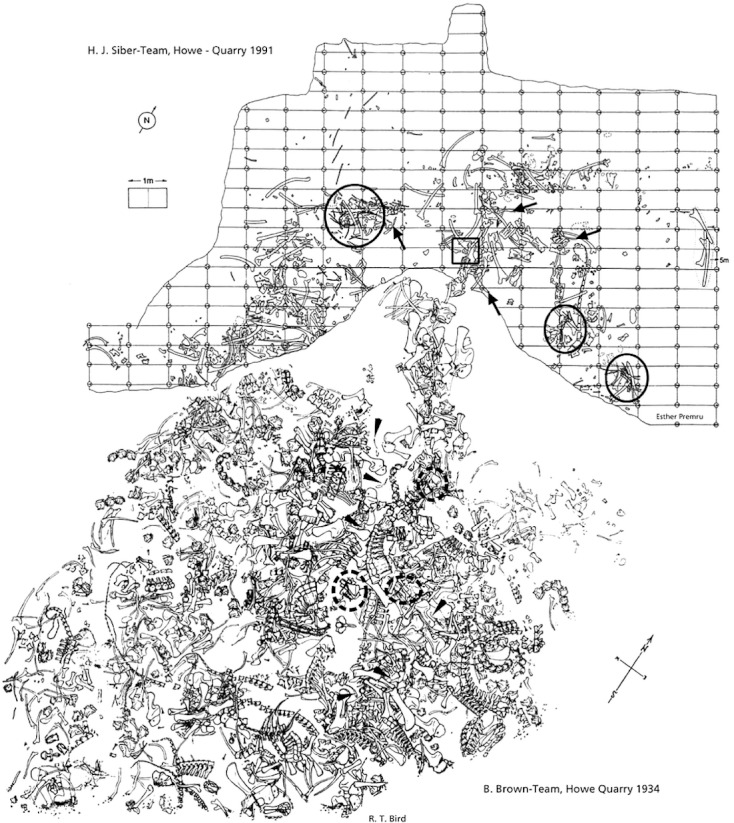
Compiled quarry map of the two excavation periods at the Howe Quarry (AMNH map below; SMA map above). Arrows indicate supposed clavicles at SMA, arrowheads possible locations of the supposed clavicle at AMNH. Circles indicate gastral or sternal baskets (full circles: SMA; dashed circles: AMNH), rectangle marks the SMA pair of symmetrical bones. AMNH map modified from Bird (1985); SMA map drawn by Esther Premru.
Both the AMNH and the SMA expeditions yielded various sets of bones resembling the gastralia or sternal ribs described from Eobrontosaurus yahnahpin (Filla & Redman, 1994). Furthermore, five single elements like the bones previously identified as sauropod clavicles, as well as two pairs of L-shaped, symmetrical elements were recovered, the identity of which is discussed below (Fig. 2; Table 2). Although such an accumulation of chest bones is unusual, none of these bones from the Howe Quarry has been reported to date. Given the predominance of Diplodocidae in the Howe Quarry, and the close association of some of the clusters of gastralia/sternal ribs and the clavicles to diplodocid cervical vertebrae, an attribution of these elements to this group can be considered highly probable.
Fig. 2.

Detail of the 1991 quarry map, with sections producing associated morphotype C–E elements enlarged (from left to right: clusters M 21, F 27 and D 28). The morphotype C–E elements are highlighted in grey in the enlarged sections.
Table 2.
New and already reported chest bones of sauropods, ordered by morphotype and first mention. Reported chest bones of unknown morphotype are listed in the end
| Specimen Number | Taxonomy | References | first identified as | Comments | |
|---|---|---|---|---|---|
| Morphotype A | CM 84 | Diplodocus carnegii | Hatcher, 1901, 1903; Nopcsa, 1905; Holland, 1906; Mcintosh 1981 | clavicle | |
| HMNS 175 (= CM 662) | Diplodocus hayi | Hatcher, 1903; Nopcsa, 1905; Holland, 1906; | clavicle | stored at CM | |
| ? | Mamenchisaurus sp. | Dong et al. 1983; | clavicle | ||
| ? | Omeisaurus junghsiensis | Dong et al. 1983; | clavicle | three elements mentioned | |
| IVPP V7262 | Datousaurus bashanensis | Dong & Tang, 1984; | clavicle | ||
| IVPP V7263 | Datousaurus bashanensis | Dong & Tang, 1984; | clavicle | ||
| T5704 | Omeisaurus tianfuensis | He et al. 1988; | clavicle | ||
| T5401 | Shunosaurus lii | Zhang, 1988; | clavicle | ||
| ANS 21122 | Suuwassea emilieae | Harris, 2007; | clavicle | ||
| NMB-1698-R | Spinophorosaurus nigerensis | Remes et al. 2009; | clavicle | ||
| AMNH 30900 | ?diplodocid | this study | interclavicle | ||
| SMA I 24-4 | ?diplodocid | this study | interclavicle | ||
| SMA L 22-3 | ?diplodocid | this study | interclavicle | ||
| SMA L 27-7 | ?diplodocid | this study | interclavicle | ||
| SMA M 25-3 | ?diplodocid | this study | interclavicle | ||
| Morphotype B | SMA 0009 | non-somphospondylian macronarian | Schwarz et al. 2007b; this study | possible furcula | pair recovered |
| GCP-CV 4229 | Spinophorosaurus nigerensis | Remes et al. 2009; | tail spikes | two elements, possibly not symmetrical | |
| DQ-SB | Diplodocidae indet. | Galiano & Albersdörfer, 2010; Galiano & Albersdörfer, 2010; A. Maltese, pers. comm. 2012 | clavicle | pair recovered | |
| DQ-TY | Diplodocidae indet. | clavicle | articulated pair (Bader et al. 2009: Fig. 4B) | ||
| KUVP 129716 | Camarasaurus sp. | – | |||
| SMA K 24-3 & 6 | ?diplodocid | this study | clavicle | pair recovered | |
| AMNH 30789 | ?diplodocid | this study | clavicle | pair recovered | |
| Morphotype C | AMNH 5760/5761 | Camarasaurus supremus | Osborn & Mook, 1921; | sternal/ventral rib | single element |
| Tate 001 | Eobrontosaurus yahnahpin | Filla & Redman, 1994; | gastralia | almost complete set including morphotype E as well | |
| AMNH 30901 | ?diplodocid | this study | ?sternal rib | ||
| SMA ? | ?diplodocid | this study | ?sternal rib | various unnumbered elements | |
| SMA C 17-5 | ?diplodocid | this study | ?sternal rib | ||
| SMA D 28-6 to 11 | ?diplodocid | this study | ?sternal rib | part of D 28-cluster | |
| SMA D 28-18 to 19 | ?diplodocid | this study | ?sternal rib | part of D 28-cluster | |
| SMA E 19-9 | ?diplodocid | this study | ?sternal rib | ||
| SMA E 21-2 to 3 | ?diplodocid | this study | ?sternal rib | ||
| SMA F 19-10 | ?diplodocid | this study | ?sternal rib | ||
| SMA F 19-21 | ?diplodocid | this study | ?sternal rib | ||
| SMA F 20-9 | ?diplodocid | this study | ?sternal rib | ||
| SMA F 27-16 to 17 | ?diplodocid | this study | ?sternal rib | part of F 27-cluster | |
| SMA F 27-33 to 35 | ?diplodocid | this study | ?sternal rib | part of F 27-cluster | |
| SMA G 21-2 | ?diplodocid | this study | ?sternal rib | ||
| SMA G 27-3 to 4 | ?diplodocid | this study | ?sternal rib | probably part of F 27-cluster | |
| SMA G 27-22 to 23 | ?diplodocid | this study | ?sternal rib | probably part of F 27-cluster | |
| SMA H 20-7 | ?diplodocid | this study | ?sternal rib | ||
| SMA H 21-2 | ?diplodocid | this study | ?sternal rib | ||
| SMA H 21-5 | ?diplodocid | this study | ?sternal rib | bears a foramen | |
| SMA H 21-9 to 10 | ?diplodocid | this study | ?sternal rib | ||
| SMA H 21-12 | ?diplodocid | this study | ?sternal rib | ||
| SMA L 21-3 to 5 | ?diplodocid | this study | ?sternal rib | probably part of M 21-cluster | |
| SMA M 21-4 | ?diplodocid | this study | ?sternal rib | part of M 21-cluster | |
| SMA M 21-6 to 7 | ?diplodocid | this study | ?sternal rib | part of M 21-cluster | |
| SMA M 21-11 | ?diplodocid | this study | ?sternal rib | part of M 21-cluster | |
| SMA M 21-13 | ?diplodocid | this study | ?sternal rib | part of M 21-cluster | |
| SMA N 22-2 | ?diplodocid | this study | ?sternal rib | probably part of M 21-cluster | |
| SMA P 19-1 | ?diplodocid | this study | ?sternal rib | ||
| SMA P 21-1 | ?diplodocid | this study | ?sternal rib | ||
| SMA S 22-3 | ?diplodocid | this study | ?sternal rib | ||
| SMA V 21-1 | ?diplodocid | this study | ?sternal rib | ||
| SMA 0011 | Diplodocidae indet. | this study | ?sternal rib | ||
| Morphotype D | SMA D 28-5 | ?diplodocid | this study | gastralia | part of D 28-cluster |
| SMA D 28-14 to 15 | ?diplodocid | this study | gastralia | part of D 28-cluster | |
| SMA F 19-11 to 12 | ?diplodocid | this study | gastralia | ||
| SMA G 21-3 | ?diplodocid | this study | gastralia | ||
| SMA M 21-2 | ?diplodocid | this study | gastralia | part of M 21-cluster | |
| SMA M 21-8 | ?diplodocid | this study | gastralia | part of M 21-cluster, fused element | |
| SMA M 21-16 | ?diplodocid | this study | gastralia | part of M 21-cluster, fused element | |
| SMA N 21-3 | ?diplodocid | this study | gastralia | probably part of M 21-cluster | |
| Morphotype E | YPM 1980 | Apatosaurus excelsus | Marsh, 1883, 1896; | sternal ribs | several elements, 2 figured |
| Tate 001 | Eobrontosaurus yahnahpin | Filla & Redman, 1994; | gastralia | almost complete set including morphotype C as well | |
| ML 684 | ?non-titanosauriform macronarian | Tschopp & Mateus, 2012; | sternal rib | fused with sternal plate | |
| SMA H 21-1 | ?diplodocid | this study | sternal rib | associated with SMA H 21-3 and morphotype C elements | |
| SMA H 21-3 | ?diplodocid | this study | sternal rib | associated with SMA H 21-1 and morphotype C elements | |
| SMA M 21-15 | ?diplodocid | this study | sternal rib | part of M 21-cluster | |
| SMA N 22-12 | ?diplodocid | this study | sternal rib/ intercostal element | probably part of M 21-cluster | |
| unknown | ?CM 3018 | ?Apatosaurus louisae | Holland, 1915; | sternal ribs | several elements, not described/figured |
| GMNH-PV 101 | Camarasaurus grandis | McIntosh et al. 1996; | gastralia | one element, figured but not described | |
| ? | Gongxianosaurus shibeiensis | He et al. 1998; | gastralia | several elements, not described, inadequately figured | |
| ? | Jobaria tiguidensis | Sereno et al. 1999; | clavicle | not described, inadequately figured | |
| ? | Jobaria tiguidensis | Sereno et al. 1999; | gastralia | several elements, not described/figured | |
| DNM ? | ?Camarasaurus | Claessens, 2004; | sternal ribs | several elements in the wall, not described/figured, possibly the same as the ones mentioned by Holland, 1915 | |
| AODF 603 | Diamantinasaurus matildae | Hocknull et al. 2009; | gastralia | fragmentary, not described/figured | |
| DQ-TY | Diplodocidae indet. | Galiano & Albersdörfer, 2010; | sternal ribs | several elements, not described/figured | |
| DQ-SB | Diplodocidae indet. | Galiano & Albersdörfer, 2010; | gastralia or sternal ribs | several elements, not described/figured | |
| DQ-EN | Diplodocidae indet. | Galiano & Albersdörfer, 2010 | sternal ribs | several elements, not described/figured | |
| DMNS 59329 | Diplodocus carnegii | Denver Museum Database | clavicle | not described/figured; probably morphotype A |
Description and discussion
Morphotype A
Previous identification: clavicles, sternal ribs, os penis (baculum; Fig. 3; Table 3; Suppl. Figs 1–5)
Fig. 3.
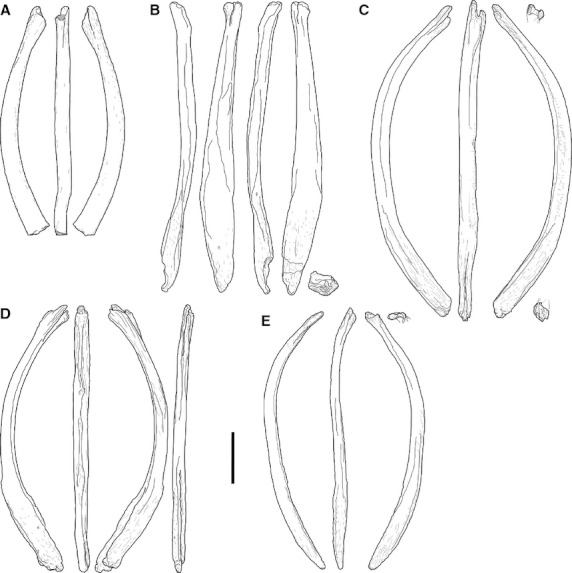
Morphotype A elements, to scale. (a) AMNH 30900; (b) SMA I 24-4; (c) SMA M 25-3; (d) SMA L 22-3; (e) SMA L 27-7. Scale bar: 10 cm. Gray areas in (a) indicate broken surfaces. Note the bifurcate end on top and the spatulate end at the bottom.
Table 3.
Measurements of new and the two first reported finds of morphotype A elements (interclavicles; in mm)
| Specimen | AMNH 30900 | SMA M 25-3 | SMA L 27-7 | SMA I 24-4 | SMA L 22-3 | CM 84 | HMNS 175 |
|---|---|---|---|---|---|---|---|
| Length along curvature | 455* | 650 | 545 | 550 | 554 | 660 | |
| Length measured straight | 437* | 585 | 484 | 540 | 510 | 613 | |
| Length spatulate portion | 185 | 173 | 200 | 206 | 225 | ||
| Width compressed end | 21* | 25 | 6 | 31 | 24 | 77 | |
| Width bifurcated end | 29 | 30 | 27 | 35 | 21 | 75 | |
| Depth of bifurcation | 17 | 30 | 17 | 19 | 41 | 50 | 40 |
Indicate incomplete measurements due to fractures, empty cells were measurements impossible to obtain.
Our interpretation: interclavicles.
General morphology
Morphotype A elements are relatively stout, elongated bones. They are usually bowed, and exhibit a spatulate and a bifurcate end. Interpreting that the concave side follows the curvature of the body, this side can be regarded as internal. The spatulate end bears more or less linear rugose striations for muscle or ligament attachment. The shafts are suboval in cross-section at mid-length, and achieve their greatest breadth at two-thirds to three-quarters of their total length, toward the spatulate end. The notch in the bifurcated end is usually only visible in internal or external view.
Morphotype A elements show some superficial similarities to dorsal ribs. They can be distinguished from ribs by the presence of the spatulate and bifid ends, the more circular cross-section at midlength, and the striated rugosities present on the spatulate end.
Howe Quarry material
Five elements were located in the collections of the AMNH and the SMA (AMNH 30900; SMA field numbers I 24-4, L 22-3, L 27-7 and M 25-3; Fig. 3). Whereas the provenance and association of the AMNH element within the Howe Quarry is unclear, the SMA specimens were found close to dorsal ribs and an associated but disarticulated series of diplodocid cervical vertebrae (I 24-4); neck and skull remains of a new diplodocine sauropod (Tschopp & Mateus, in press), and a gastral/sternal rib cage (L 22-3); anterior cervical vertebrae, a dorsal rib, some skull remains and a metatarsal (L 27-7); as well as associated with dorsal ribs, posterior diplodocid cervical vertebrae and an articulated series of midcaudal vertebrae (M 25-3). All these elements were found as single elements, which is consistent with the earlier findings of similar finds in other sauropod taxa.
Previous reports
Several bones belonging to morphotype A have been reported in the literature, and were usually identified as sauropod clavicles (Table 2). The species preserving morphotype A elements are the non-neosauropod Eusauropoda Shunosaurus lii, Spinophorosaurus nigerensis, Omeisaurus junghsiensis, O. tianfuensis, Mamenchisaurus sp. and Datousaurus bashanensis, as well as the Flagellicaudata Diplodocus carnegii, D. hayi and Suuwassea emilieae (Dong et al. 1983; Dong & Tang, 1984; He et al. 1988; Zhang, 1988; Hatcher, 1901, 1903; Harris, 2007; Remes et al. 2009). In none of these taxa, morphotype A elements were recovered in pairs, even though some of the specimens were reasonably complete and articulated.
Variation in morphology
Two different orientations of the spatulate end can be observed in the five elements from the Howe Quarry, as well as in previously reported bones belonging to morphotype A: perpendicular to the curvature of the bone, so that its thin edges face internally and externally (e.g. SMA L 22-3; Fig. 3d), or turned 90 ° (e.g. SMA I 24-4; Fig. 3b). The spatulate end can bear a deep slot-like concavity on its internal side (e.g. in SMA M 25-3 and L 22-3; Fig. 3c,d), giving the impression of a deeply bifurcated end on this side as well, in internal view. In other elements, the internal side of the spatulate end bears shallow (Diplodocus hayi HMNS 175 or Spinophorosaurus nigerensis NMB-1698-R) to distinct ridges (Suuwassea emilieae ANS 21122) with varying lengths (Hatcher, 1903; Harris, 2007; Remes et al. 2009). Differences in curvature (see the straight element SMA I 24-4 or the outwards curve in SMA L 27-7; Fig. 3b,e, respectively) are most probably of taphonomical origin.
Identification
The first reported element of morphotype A was found associated with the scapulacoracoid of Diplodocus carnegii CM 84 and was tentatively identified as clavicle (Hatcher, 1901). Although this interpretation has never been definitely confirmed, it has become generally accepted, and subsequent finds of similar bones continued to be identified as clavicles (Hatcher, 1903; Dong et al. 1983; Dong & Tang, 1984; He et al. 1988; Zhang, 1988; Harris, 2007; Remes et al. 2009). The suggestions of Nopcsa (1905) or Holland (1906) that morphotype A elements represent the os penis or sternal ribs, respectively, are improbable given the absence of bacula in extant reptiles, and because they are relatively much more massive compared with articulated sternal ribs of pterosaurs or crocodylians (Claessens et al. 2009; Crocodylus niloticus, NHM, unregistered display specimen, ET, pers. obs.).
The five elements recovered as single bones in the Howe Quarry, as well as the corresponding elements of Diplodocus carnegii (CM 84), D. hayi (HMNS 175), Spinophorosaurus nigerensis (NMB-1698-R) and Suuwassea emilieae (ANS 21122), are slightly asymmetrical (see also Hatcher, 1901, 1903; Holland, 1906), but have a longitudinal midline extending through the bifid end and dividing the expanded spatulate end in two halves. These bones could therefore also represent elements of the chest region that lie on the body midline, and their continuous findings as single elements might have been no coincidence. The two areas abutting to the right and left of the ridges subdividing the spatulate end in the elements of Suuwassea and Spinophorosaurus resemble articulation surfaces, implying that the morphotype A elements covered two symmetrical elements externally, and medially. The absence of such a ridge in the other taxa might be of taxonomic significance.
The only median pectoral element in the non-avian shoulder girdle is the interclavicle. As the morphotype A elements, also Alligator interclavicles have slightly asymmetrical outlines (Vickaryous & Hall, 2010; R. Wilhite, pers. comm. 2012). Whereas early tetrapods had diamond-shaped interclavicles (Steyer et al. 2000), crocodylomorphs and some lepidosaurs developed rod-like shapes without lateral processes (Vickaryous & Hall, 2010), similar to the elements described herein. Following this interpretation, the bifurcated end probably represents the reduced lateral processes, and the spatulate end would articulate with either the coracoids or the sternal plates – covering them externally and anteromedially.
A bone found in the pectoral girdle of the early sauropodomorph Massospondylus carinatus shows a similar spatulate expansion on one end, and in fact has first been interpreted as interclavicle (Cooper, 1981). Sereno (1991) and Yates & Vasconcelos (2005) subsequently reinterpreted this element as a clavicle, based on its similarity to paired, articulated clavicles in both Plateosaurus and other specimens of Massospondylus. The articulated specimens connect to the acromion process of the scapulae, which is typical for clavicles, but almost never the case in interclavicles (Table 1; Romer, 1956). The only reported exception is the interclavicle in some monotreme mammals, where the transverse processes reach the acromion (Klima, 1973). The articulation of these elements with the acromia of the scapulae in both Massospondylus (Yates & Vasconcelos, 2005) and Plateosaurus (B. Pabst, pers. comm., 2011; ET, pers. obs.) thus make an interpretation as clavicles more probable. However, given that sauropods appear to have both clavicles and interclavicles, Cooper's (1981) interpretation of the single Massospondylus ‘interclavicle’ might have to be reconsidered.
An alternative hypothesis would be that morphotype A represents a neomorphic element, independently developed in Sauropoda. This would be the case if interclavicles would really have gotten lost in Dinosauriformes. However, given the incompleteness of the finds of early dinosauriforms and dinosaurs, it cannot be ruled out that the absence of an ossified interclavicle is due to taphonomy. Therefore, and since the presence of an interclavicle is plesiomorphic for tetrapods, an interpretation of the sauropod morphotype A elements as interclavicle seems more appropriate.
Morphotype B
Not previously recognized (Figs 4 and 5; Suppl. Figs 6–9).
Fig. 4.
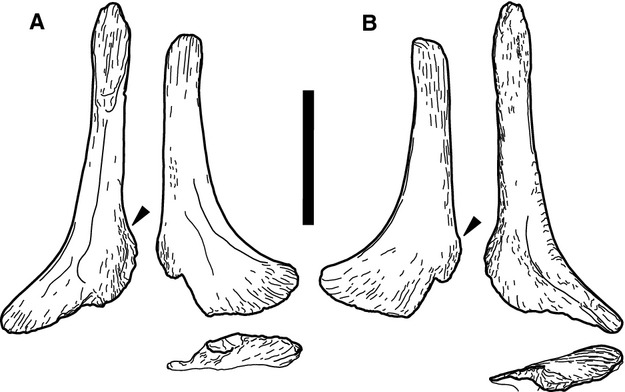
Pair of Morphotype B elements SMA K 24-3 (outer bone) and SMA K 24-6 (inner bone) in internal (a) and external (b) view. Short leg of L-shaped bones shown in perpendicular view below. Note the considerable bend of this portion in respect to the main axis of the bone. Scale bar: 10 cm.
Fig. 5.
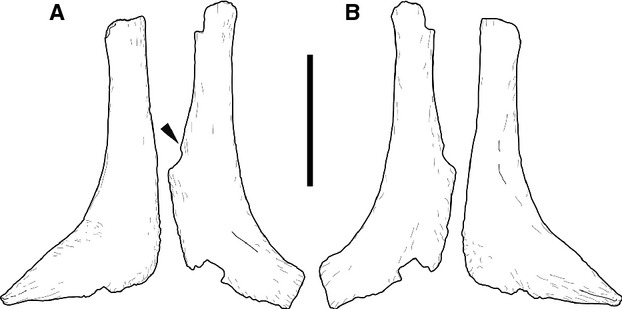
Pair of Morphotype B elements AMNH 30789 in internal (a) and external (b) view. Scale bar: 10 cm.
Our interpretation: clavicles.
General morphology
Morphotype B elements are L-shaped, and of similar thickness as morphotype A, but shorter. They are concave on one side, and convex on the opposite surface, and are usually found in pairs. Morphotype B elements have a D- to crescentic-shaped cross-section at midlength. The convex side is hereinafter interpreted as external, the flat to slightly concave surface as internal. Towards the end of the longer leg of the L, a striated rugosity develops on both sides, and the bone expands slightly. This end is broken pre-burial in one of the SMA elements (K 24-6; Fig. 4), and post mortem in both AMNH elements, so that they appear shorter and stouter (Fig. 5). The shorter leg of the L is expanded ‘backwards’ as well, especially so in the SMA specimen, forming a somewhat heel-like, rounded flange (Figs 4 and 5). Towards the tip of the short leg, the bone curves externally. This portion shows a similar but stronger striated rugosity as in the longer leg of the L.
Morphotype B elements have a peculiar morphology. The most similar bones are the anterior- or posterior-most dorsal ribs, but neither capitulum nor tuberculum are present on the morphotype B. Furthermore, the striations marking soft tissue attachment do not occur on dorsal ribs.
Howe Quarry material
Two pairs of morphotype B elements were found at this site. The first specimen was found in 1934 by the AMNH (AMNH 30789), the other pair was recovered in the SMA excavation and bear the field numbers SMA K 24-3 and K 24-6. Whereas it is clear that the two bones of AMNH 30789 were found together (this collection number was given to all bones in a plaster jacket bearing the field number 151), their placement within the Howe Quarry is impossible to locate to date. AMNH 30789 also includes chevrons and pedal material. The SMA specimens were found closely together, below several dorsal ribs, and between posterior cervical and anterior dorsal diplodocid vertebrae.
Previous reports
Not much is known about the occurrence of morphotype B elements in sauropods. The only formal reports of similar elements concerns an element recovered with an early juvenile sauropod (Schwarz et al. 2007b), as well as two bones belonging to the holotype of Spinophorosaurus nigerensis (Remes et al. 2009). Due to superficial affinities to elements in Shunosaurus lii, Remes et al. (2009) tentatively identified the Spinophorosaurus elements as tail spikes. Recently, Galiano & Albersdörfer (2010) informally reported three elements (one pair and a single bone) found associated with indeterminate diplodocid sauropods.
Variation in morphology
One of the AMNH elements develops a conspicuous ridge towards the tip of the short leg of the L. In its counterpart, this end is broken off. Contrary to the state in the AMNH elements, the same end is transversely expanded in the single morphotype B bone reported by Galiano & Albersdörfer (2010; H. Galiano, pers. comm. 2011).
The SMA specimens bear a thickened portion resembling an articulation facet at about midlength of the shorter leg of the L. As the AMNH elements show broken edges in this region, this facet might also constitute to the general morphology.
Identification
The three morphotype B elements recently reported by Galiano & Albersdörfer (2010) include a set of paired bones articulated between the acromia of the scapulae of a diplodocid specimen (DQ-SB; Fig. 6), where clavicles are supposed to attach (Romer, 1956; Vickaryous & Hall, 2006). Similarly, detailed examination of the juvenile sauropod SMA 0009 revealed that a second, L-shaped element is actually present attached to the dorsal corner of the right coracoid (Fig. 7). Schwarz et al. (2007b), in their initial description of SMA 0009, described this bone as ‘dorsally pointing tip’ of the coracoid, but a thin layer of matrix between the elements indicates that they are taphonomically pressed onto each other. A third pair of morphotype B elements, articulated between the scapulae, appears to be present in a Camarasaurus (KUVP 129716, Bader et al., 2009; A. Maltese, pers. comm. 2012).
Fig. 6.
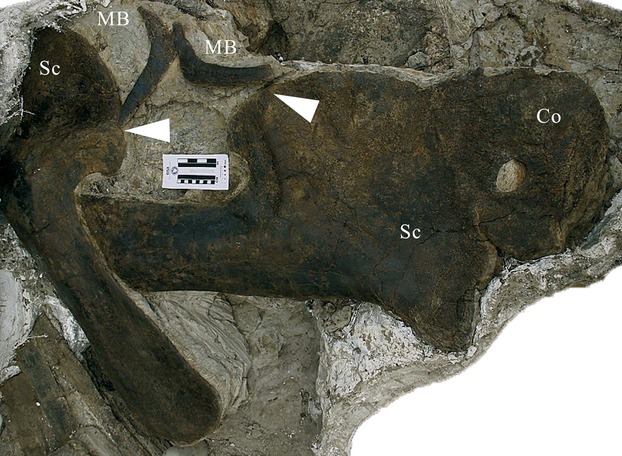
Morphotype B elements of the diplodocid DQ-SB, articulated with the acromia (arrowheads) of the scapulae, as they were found. Co, coracoid; MB, morphotype B element; Sc, scapula. Picture courtesy of H. Galiano.
Fig. 7.

Coracoid with taphonomically attached morphotype B element (MB) of the non-somphospondylian macronarian SMA 0009 in posteroventral (a) and lateral (b) view. Coracoid made semitransparent in order to visualize better the morphotype B element. Arrows indicate brightly coloured matrix present between the MB and the coracoid. CF, coracoid foramen; GL, glenoid surface. Scale bar: 2 cm.
The topology and paired appearance of these morphotype B elements strongly suggests that they fit an identification as clavicles better than morphotype A. Such an interpretation would also match previous findings of similarly shaped clavicles in ceratopsian dinosaurs (Chinnery & Weishampel, 1998: Fig. 6; Vickaryous & Hall, 2010: Fig. 5). On the other hand, paired finds of non-sauropod sauropodomorph clavicles appear to be more straight (von Huene, 1926; Yates & Vasconcelos, 2005; Martínez, 2009; B. Pabst, pers. comm., 2011), but without the bifurcated end – resembling the elements recovered from Jobaria tiguidensis (Sereno et al. 1999: Fig. 3e). However, other than the report of the presence of this bone in J. tiguidensis, no other information about which bones it was associated with, and no detailed description has been provided to date.
Another explanation would be that morphotype B comprises sternal ribs. However, a taphonomical shifting of sternal ribs into a position equal to the clavicular articulation typical for tetrapods (as is the case in the specimen DQ-SB) is highly improbable. Therefore, an interpretation of morphotype B elements as the true sauropod clavicles is the most convincing. This interpretation challenges Hatcher's (1901, 1903) identification of the morphotype A bones. As the gross morphology of the shoulder girdle remains similar in the majority of Sauropoda, a high diversity in the shape of clavicles seems improbable. Therefore, an interpretation of the morphotype A elements as interclavicles is supported as well by the presence of morphotype B bones as the true clavicles.
Morphotype C
Previous identification: gastralia, ventral ribs, sternal ribs (Fig. 8; Suppl. Figs 10–11).
Fig. 8.
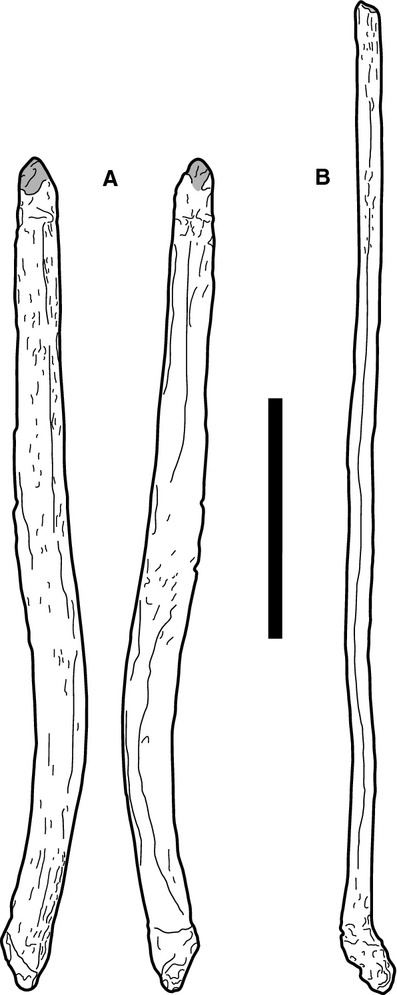
Morphotype C elements SMA H 20-7 (a) and L 21-5 (b). Both elements are incomplete, fracture surface at the top is indicated by the grey area. Scale bar: 10 cm.
Our identification: sternal ribs.
General morphology
Morphotype C elements are elongated, rod-like bones with a suboval cross-section. The smooth shafts are generally slightly curved, in some elements (e.g. SMA D 28-6, D 28-7) in two directions forming a weak S-shape. Both ends are rugose and irregular. One end is flattened and often shows differing degrees of rugosity on the two sides of the flattened portion.
The most similar bones to morphotype C elements would be cervical ribs or ossified tendons. However, cervical ribs are usually concave dorsally. Ossified tendons are often attached to other bones, or do not exhibit expanded ends, but if only a portion of the shaft of a morphotype C element is found, distinguishing between the two might be difficult.
Howe Quarry material
In the Howe Quarry sample, three clusters of gastralia/sternal ribs were found by the SMA (around field area D 28, F 27 and M 21; Fig. 2). All three clusters contain about 15 single elements. Within these clusters, morphotype C elements constitute the majority of the recovered bones. They (as well as elements belonging to morphotypes D and E) were always found in association with dorsal ribs. In the field area F 27, also two distal tail segments of different sizes as well as single posterior cervical vertebrae were recovered in the vicinity of the gastral/sternal rib cage. The M 21 cluster was associated with the skull and neck of a diplodocine sauropod, as well as single (probably diplodocid) anterior chevrons. Additional morphotype C elements come from various areas within the Howe Quarry, and were found more scattered.
Previous reports
Such elements were usually described as sternal or gastral ribs, and are often associated with bones of morphotypes D and E. Taxa reported to preserve morphotype C elements include the non-neosauropod eusauropod Jobaria tiguidensis, the diplodocids Apatosaurus louisae, Eobrontosaurus yahnahpin, as well as the macronarian Camarasaurus supremus and Diamantinasaurus matildae (Holland, 1915; Osborn & Mook, 1921; Filla & Redman, 1994; Sereno et al. 1999; Hocknull et al. 2009).
Variation in morphology
Instead of being slightly curved, some morphotype C elements remain straight during their entire length. Toward the non-flattened ends, some of the elements remain straight, whereas others show a distinct bend.
Identification
Probable sauropod gastralia and/or sternal ribs have been reported more often than clavicles, and in a wider range of taxa (Table 2). As mentioned above, their interpretation as gastralia has been challenged by Claessens (2004), pointing out their anatomical differences compared with non-sauropod sauropodomorph or theropod gastralia. Claessens (2004) proposes that Marsh (1896) was right in identifying such elements as sternal ribs.
Sternal ribs are of endochondral origin, and present in both extant birds and crocodylians, but remain often cartilaginous (Claessens, 2004; Claessens et al. 2009; R. Fechner, pers. comm., 2011). They connect the distal tips of the anterior dorsal ribs with the sternum, either directly as in birds, or articulating with the dorsal ribs through generally cartilaginous intercostal elements (Claessens et al. 2009; R. Fechner, pers. comm., 2011; pers. obs.). Within dinosaurs, only very few reports of sternal ribs exist besides the ones from Marsh (1883, 1896): they are described in hypsilophodont Ornithischia (e.g. Parks, 1926; Galton & Jensen, 1973; Weishampel & Heinrich, 1992), and Theropoda (e.g. Clark et al. 1999; Ruben et al. 2003).
Gastralia are dermal bones embedded in the abdominal musculature, and are usually thought to support the breathing apparatus and/or protect the belly (Claessens, 2004; Claessens et al. 2009). As sternal ribs, also gastralia are present in both birds and crocodylians (Claessens, 2004). Unambiguous evidence for gastralia in dinosaurs appears to exist only in Theropoda and early Sauropodomorpha (‘prosauropods’; Claessens, 2004; R. Fechner, pers. comm., 2011). None of the bones of morphotype C exhibit the typical longitudinal articulation facets that occur between the medial and lateral elements of theropod or early sauropodomorph gastralia (Claessens, 2004). On the other hand, for a bird-like sternal rib configuration, 15 elements are too many: in birds, distally expanded dorsal ribs usually connect to sternal segments through cartilage (Parks, 1926; Clark et al. 1999; Schwarz et al. 2007a), and straight or converging distal rib ends mark free ribs. Fully articulated ribcages of Apatosaurus and Diplodocus show transversely expanded ends only in the first five–seven dorsal ribs (Gilmore, 1936; Schwarz et al. 2007a) – which would allow a maximum number of 14 sternal ribs. However, the about 15 elements recovered per cluster include at least three morphotype D elements. Given that the latter most probably are not sternal ribs (see below), the maximum number of sternal ribs per individual would not be exceeded. An identification of morphotype C elements as sternal ribs is thus the most convincing.
Morphotype D
Not previously recognized (Fig. 9; Suppl. Figs 12–14).
Fig. 9.
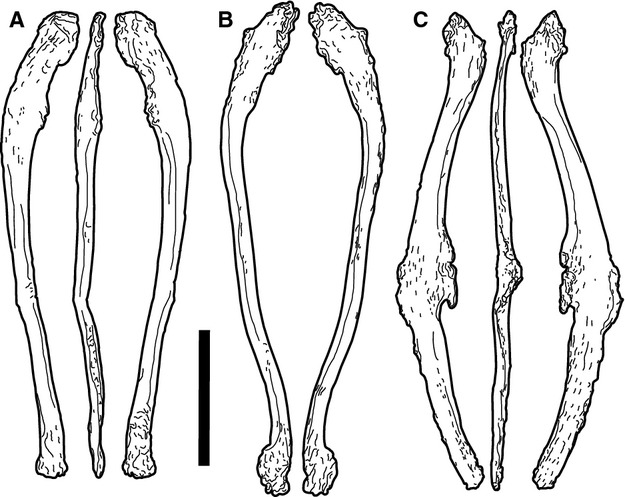
Morphotype D elements SMA D 28-5 (a), M 21-2 (b) and M 21-8 (c). The bottom end of M 21-8 is broken. Scale bar: 10 cm.
Our identification: gastralia.
General morphology
Morphotype D elements are more irregularly formed than morphotype C, shorter and thicker. They are curved bones with both ends expanded and rugose. The expansions are not equal on the two extremities, one of them being wider than the other. The wider end is flattened, very irregularly expanded, and with strong rugosities. On one side, this end is slightly convex, indicating that this side was not articulating with any other element. These bones all show some curvature at the opposite end, resulting in an outward pointing extremity.
Morphotype D elements can be very similar to bones belonging to morphotype C, and thus also to ossified tendons and cervical ribs. The wide, probably medial (see below) extremity is the best characteristic to define morphotype D. It is more irregular, wider and resembles more bony overgrowth than what is usually present in morphotype C.
Howe Quarry material
Morphotype D elements are less frequently found. However, all three gastral/sternal rib clusters in the SMA collection contain morphotype D elements. Two bones of the cluster D 28 are symmetrical, and can be nicely articulated at their wider ends (D 28-5 and 14; Fig. 10).
Fig. 10.

Proposed articulation between two morphotype D elements (left, SMA D 28-5; right, SMA D 28-14) in three views (internal/dorsal view in the centre, grey lines indicate the same morphological landmarks on the respective elements). Note the similarity to the central portion of the fused morphotype D element (Fig. 9c). Scale bar: 5 cm.
Previous reports
To our knowledge, elements of this morphotype are described for the first time in this paper. It is possible, however, that previously mentioned sets of ‘gastralia’ or ‘sternal ribs’ include morphotype D elements, but that these were not recognized as such and not figured (see Table 2).
Variation in morphology
Certain elements in the M 21-basket (e.g. SMA M 21-8; Fig. 9c) appear to be fused symmetric elements. They reproduce the slight upward curvature of two articulated opposing elements similar to the pair SMA D 28-5 and 14, and exhibit an outgrowth in the middle of the bone, which would come to lie on the body midline. This outgrowth resembles somewhat pathological bony overgrowth but also the shape of two unfused anterior or posterior gastralia with their enlarged medial ends. Towards the extremities the curvature of the bone becomes inverted in a way that the expanded ends are pointing somewhat downwards again (or probably straightly outwards when articulated). This results in a slightly sinuous curve, similar to tyrannosaur furculae described by Makovicky & Currie (1998).
Identification
Although associated with the probable sternal ribs belonging to morphotype C, bones like SMA M 21-8 (Fig. 9c), which seem to be composed of two fused elements like D 28-5 or M 21-2 (Fig. 9a or b, respectively), have no equivalent in previously described sternal ribs known to us. Sternal ribs sometimes connect to other, more anteriorly placed elements, instead of articulating directly with the sternal plates (Galton & Jensen, 1973; Clark et al. 1999; Claessens et al. 2009), but no specimen has been reported to date exhibiting fused left and right ribs.
Manual manipulation of the two corresponding elements SMA D 28-5 and 14 shows that the expanded ends would articulate relatively nicely in a way similar to the midline joint of two gastralia in non-sauropod sauropomorphs and theropods (Fig. 10). Median gastralia of the anterior-most row were previously shown to fuse in certain cases, thereby forming irregularly shaped and asymmetric sutures (e.g. Makovicky & Currie, 1998; Claessens, 2004). Such a development resembles much the herein described fused elements. Morphotype D is thus most convincingly interpreted as the anterior-most gastralia, close to the sternal apparatus.
Morphotype E
Previous identification: gastralia, sternal ribs (Fig. 11; Suppl. Figs 15–16).
Fig. 11.
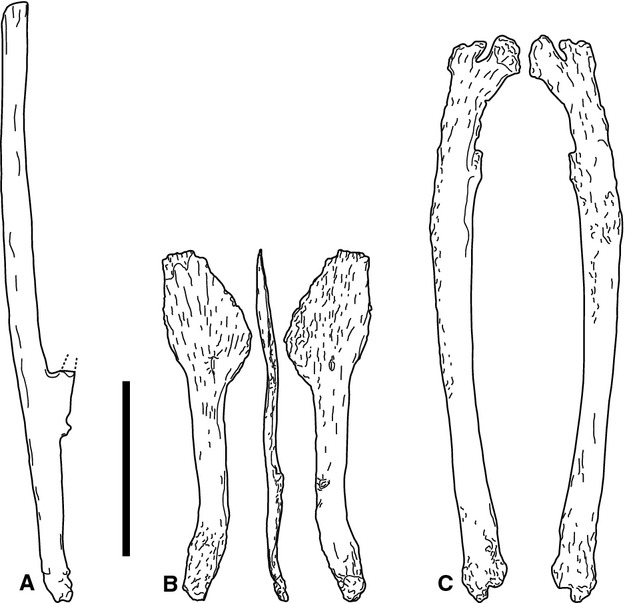
Morphotype E elements SMA H 21-3 (a), N 22-12 (b) and M 21-15 (c). Note the irregular shapes that do not allow an assignation to any other morphotype. Dotted lines in (a) indicate direction of the broken hook-like projection. Scale bar: 10 cm.
Our identification: sternal ribs and/or intercostal elements.
General morphology
Bones belonging to morphotype E have irregular shapes that cannot be included in any of the above defined morphotypes. Peculiar morphologies include projections (e.g. SMA H 21-1; Fig. 11a), irregular expansions (SMA N 22-12; Fig. 11b), and bifurcated ends (SMA M 21-15; Fig. 11c). Due to their particular shapes, morphotype E elements do not resemble any other bone in the sauropod skeleton.
Howe Quarry material
Few of these elements were recovered at the Howe Quarry, always in association with bones of the morphotypes C and D. SMA H 21-1 and 3 form a symmetrical pair.
Previous reports
Similar elements include bones of Apatosaurus excelsus, identified as sternal ribs by Marsh (1883, 1896), and some of the elements of the gastral basket of Eobrontosaurus yahnahpin described by Filla & Redman (1994).
Variation in morphology
SMA H 21-1 and H 21-3 develop a projection approximately at one-third to two-fifths of their entire length, which appears to proceed at an acute angle to the longer portion of the bone (Fig. 11a). How long this projection is remains unclear, as their ends are broken in both elements. M 21-15 is a rather thick bone of medium length, compared with the usual gastralia/sternal ribs. Both ends are flattened, one of them is markedly and slightly asymmetrically bifurcated (Fig. 11c). On the edge running from the longer portion of the bifurcation, somewhat inwards, a tubercle can be seen with fractured bone surface so that the original expansion of this feature can not be determined. The opposing end is irregular as well, exhibiting a very shallow notch. N 22-12 is a short and very thin bone, with one end greatly expanded in two dimensions, forming a spatulate shape with irregular margins, and a weak, radiating striation extending from the centre of the bone towards the outer margins on both sides (Fig. 11b). At the base of this expansion, both sides are marked by a well visible foramen that lie on the same level in regard to the long axis of the bone, and only very slightly displaced perpendicular to the long axis. Towards the other end, at about two-thirds of the entire length, there is a rugose tubercle. Further towards this end, the bone curves and becomes more rugose again.
Identification
Considering a crocodylian arrangement, morphotype E elements (like the particularly shaped SMA N 22-12, and maybe also the short elements described by Filla & Redman, 1994: Fig. 11h,i,q,r) might represent intercostal elements. Furthermore, the two SMA elements with the projection (H 21-1 and H 21-3), as well as the very irregularly shaped bone figured by Marsh (1896): Figs 12 and 13) resemble somewhat the posterior sternal ribs in the pterosaur Rhamphorhynchus (Claessens et al. 2009: Fig. 2d). The projections as well as the bifurcations might have articulated with more anterior sternal ribs.
Fig. 12.
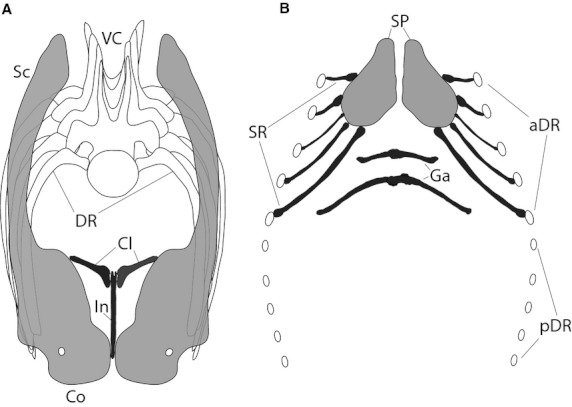
Reconstruction of the pectoral girdle and the chest region of an indeterminate diplodocid sauropod, based on the finds reported. Light grey elements represent pectoral girdle elements not discussed in the paper, dark grey elements mark the bones identified as chest bone morphotypes in this paper. Anterior (a) and ventral (b) view. Abbreviations: aDR, anterior dorsal ribs; Cl, clavicle (morphotype B); Co, coracoid; DR, dorsal rib; Ga, gastralia (morphotype D); In, interclavicle (morphotype A); pDR, posterior dorsal ribs; Sc, scapula; SP, sternal plates; SR, sternal ribs (morphotypes C and E); VC, vertebral column. Modified from Schwarz et al. (2007a; a) and Filla & Redman (1994; b).
Fig. 13.
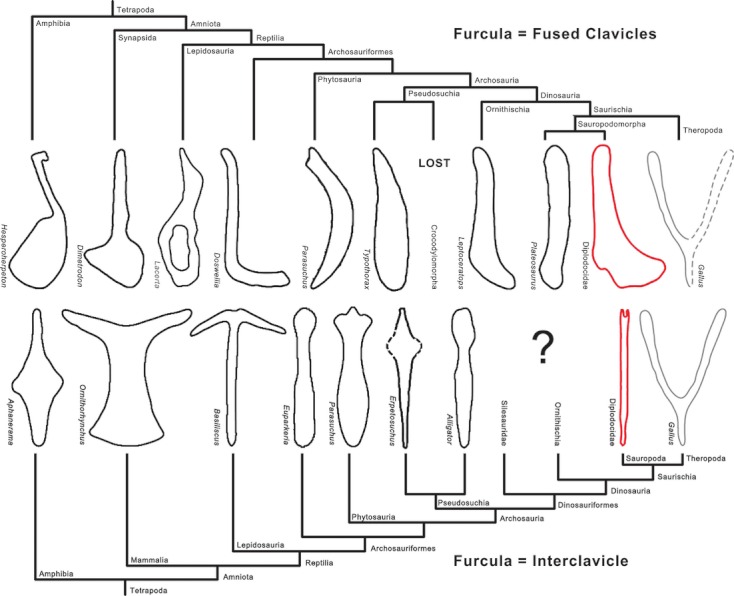
Evolution of the furcula, comparison between the two hypotheses. Note the gap within Dinosauriformes in the furcula-interclavicle hypothesis. Line drawings scaled to same size. Eaton & Stewart (1960: Hesperoherpeton); Chatterjee (1978: Parasuchus); Klima (1987: Ornithorhynchus); Rieppel (1992: Lacerta); Steyer et al. (2000: Aphanerama); Benton & Walker (2002: Erpetosuchus); Martz (2002: Typothorax); Vickaryous & Hall (2010: Dimetrodon; 2010: Alligator, Basilicus, Gallus, Leptoceratops); Remes (2008: Euparkeria); Dilkes & Sues (2009: Doswellia).
As gastral and sternal ribs have differing developmental origins (Table 1), a histological analysis might yield more definitive results concerning the identity of morphotypes C–E, but should include elements of all of them. However, histological sections of dermal and endochondral bones are difficult to distinguish in a fully ossified state (T. Scheyer, pers. comm., 2009). Such a study has thus to await further analyses and comparisons of known gastralia and sternal ribs in extant animals, and lies outside the scope of this paper.
Morphological implications
The rarity of finds of ossified chest elements other than the scapulacoracoid or the sternal plates render proper identifications difficult, especially due to the fact that they are often recovered disarticulated from the corresponding pectoral girdle. This might imply that the soft tissue connection between them and the pectoral girdle or the sternal apparatus was not very strong during lifetime – and as a consequence, chest bones were possibly easily disarticulated if not lost entirely before burial. When preserved and found, the indistinct shape and consequential difficulties identifying these elements make them more likely to not be reported or not even collected, increasing such a taphonomic bias even more. Nonetheless, the herein reported bones indicate that additional elements like the clavicles, interclavicle, sternal ribs, gastralia, and possibly intercostal elements do ossify in some sauropod taxa. However, it must be noted that no articulated sauropod specimen has yet been found preserving both morphotype A (interclavicles) and B elements (clavicles). The finds, where clavicles were found articulated (DQ-SB, KUVP 129716) or associated (SMA 0009) with the scapulacoracoids, do not appear to preserve an interclavicle. In the Howe Quarry sample described herein, all the elements were found disarticulated in a bonebed. Nonetheless, the tight association with diplodocid material suggests that if not from the same individuals, they were at least from the same taxon. A novel reconstruction of the diplodocid pectoral girdle and sternal apparatus, taking these interpretations into account, is shown in Fig. 12.
The only species from which both types of bones are reported is Spinophorosaurus nigerensis, but they are from two different individuals (Remes et al. 2009). The somewhat L-shaped elements of the holotype of Spinophorosaurus nigerensis appear to be of considerably different sizes, which was one of the reasons leading to their identification as tail spikes (Remes et al. 2009; R. Kosma and A. Ritter, pers. comm., 2011). However, the Spinophorosaurus elements resemble much more the L-shaped bones found at the Howe Quarry than the supposed Shunosaurus tail spikes (Zhang, 1988). They do not bear the typical osteoderm surface rugosity as seen in thyreophorans and sauropods, neither shows the specimen any club-like distal extension as present in Shunosaurus. Compared with the Howe Quarry material, the base of the Spinophorosaurus elements (which would correspond to the shorter leg of the L) is slightly broader, and the two legs curve gently into each other, giving the entire bone a rather triangular outline. Also, the obviously broken edges of the preserved elements shed some doubt on them being of considerably different size as described by Remes et al. (2009). Given that these elements were found below the scapula (Remes et al. 2009), an interpretation as clavicles seems possible for the Spinophorosaurus elements, and the original material should be reassessed under the light of the new findings.
Functional implications
An ossification of such a variety of chest elements creates anchor attachments for musculature and stabilizes the entire pectoral girdle, the sternal apparatus and in case of ossified gastralia also the rest of the trunk. A substitution of soft tissues (probably cartilage or ligaments in the case of the sauropod pectoral apparatus) by bone helps to cope with higher loads (Romer, 1956; Haines, 1969). The sauropod taxa exhibiting ossification of the various chest elements show some correlation with taxa usually interpreted to use their tail as laterally swinging defensive weapon, indicated by the presence of tail clubs (Shunosaurus, Omeisaurus, Mamenchisaurus; Zhang, 1988; Dong et al. 1989; Xing et al. 2009), possible tail spikes (Shunosaurus, Spinophorosaurus; Zhang, 1988; Remes et al. 2009; but see above) or whip-lash tails (Suuwassea, Diplodocus; Hatcher, 1901; Harris, 2006). Lateral movements of the tail might request a firm trunk in order to not disequilibrate the entire animal. Since the pelvic girdle – in contrast to the shoulder girdle – is co-ossified with the vertebral column, reinforcements would be particularly essential in the pectoral girdle. Furthermore, a posteriorly located centre of mass, as present especially in diplodocids, appears to induce important lateral stresses to the pectoral girdle during locomotion (Sander et al. 2011).
The loss of ossified chest elements coincides with the evolution of the particular wide-gauge locomotor style of titanosauriform sauropods (Wilson & Carrano, 1999; Carrano, 2005). The question remains, if the loss of ossified chest bones allowed the wider spacing of the legs, or if the latter enhanced stability enough to render the ossification useless. The presence of clavicles and/or interclavicles in Datousaurus, Jobaria, and Camarasaurus might just represent an example of retained plesiomorphies without strong functional significance.
Phylogenetical implications
The proposed presence of ossified elements in the chest region of some sauropods has also phylogenetical implications – even in case some of the above-stated interpretations would remain controversial. Although a taphonomical bias leading to the absence of chest bones in titanosauriforms and rebbachisaurs cannot be excluded to date, their distinctly taxonomically restricted appearance appears striking. As an ossification of additional chest elements in diplodocids and early eusauropods does also make sense in a functional point of view (see above), their presence is herein interpreted as plesiomorphic for Sauropoda, whereas the loss of ossified clavicles, interclavicles, sternal and/or gastral ribs might result a synapomorphy for Titanosauriformes, and maybe Rebbachisauridae as well. The single findings of sternal ribs in the lithostrotian Diamantinasaurus Hocknull et al. 2009) remain doubtful, or might represent exceptions to the rule.
The presence of interclavicles in dinosaurs is herein stated for the first time with direct morphological evidence. The other reported possible dinosaurian interclavicles were reinterpreted as clavicles or furculae, and thus their identification remains ambiguous (Cooper, 1981; Yates & Vasconcelos, 2005). This supports Vickaryous & Hall's (2010) statement that the theropod and avian furcula could also be homologous to the interclavicle instead of representing the fused clavicles, as generally proposed (Yates & Vasconcelos, 2005; Nesbitt et al. 2009). The evolutionary gap between non-dinosauriform interclavicles and theropod furculae, for which we did not have conclusive data before, is shortened by the presence of interclavicles in sauropods. If the interclavicle-furcula homology would get confirmed by future studies, theropods would have reduced the stem-like central body of the interclavicle as seen in sauropods to the hypocleidium, and the transverse processes would have been enlarged, and would have substituted the clavicles, which would have gotten lost early in theropod evolution. However, because both clavicles and interclavicles are present in sauropods, conclusions have to await finds of articulated specimens of early dinosaurs, or dinosauriforms, which might shed more light on the evolution of the theropod pectoral girdle. The strongest evidence against the furcula-interclavicle homology are the topology of the furcula (it articulates with the acromion as clavicles do; see Nesbitt et al. 2009), and the pairs of bones found in Massospondylus and early theropods, which articulate with the scapular acromion laterally, and among themselves medially (Yates & Vasconcelos, 2005; Nesbitt et al., 2009). Since in articulation, they resemble much the theropod furcula, and Nesbitt et al. (2009) interpreted them as an intermediate evolutionary state between unfused clavicles and the furcula. The competing hypotheses are summarized in Fig. 13.
Conclusions
Several elements recovered at the Howe Quarry (Bighorn County, Wyoming, USA) resemble bones previously identified as sauropod clavicles, as well as gastralia and/or sternal ribs. The finding of pairs of symmetric bones associated with pectoral girdle elements sheds new light on these old interpretations. In fact, detailed investigations lead to the conclusion that the bones previously supposed to represent clavicles, most probably are interclavicles, with the symmetrical, L-shaped pairs being the true claviculae. This supports the result of developmental studies of Vickaryous & Hall (2010), which questions the loss of the interclavicle in Dinosauria – and proposes a homology between the avian furcula and the reptilian interclavicle. This would change the usual interpretation that the furcula represents the fused clavicles.
A review of the occurrence of such bones within Sauropoda implies that the tendency to ossify interclavicles, clavicles, and sternal and/or gastral ribs has a distinct taxonomic distribution, with non-neosauropod Eusauropoda and Flagellicaudata representing the plesiomorphic state, and Titanosauriformes as well as possibly Rebbachisauridae exhibiting the derived condition. Functional implications of retaining the ossified chest bones include the stabilization of the trunk in order to have a firm base for lateral movements of elongated necks and tails. On the other hand, the loss of these osseous elements could have allowed the evolution of the wide-gauge locomotion in Macronaria.
Institutional abbreviations
AMNH, American Museum of Natural History, New York; ANS, Academy of Natural Sciences, Philadelphia, Pennsylvania; AODF, Australian Age of Dinosaurs Fossil; CM, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania; DMNS, Denver Museum of Nature and Science, Denver, Colorado; DNM, Dinosaur National Monument, Vernal, Utah; DQ, Dinosauria International (Dana Quarry collection), New York; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China; KUVP, University of Kansas Natural History Museum, Kansas; NMB: Naturhistorisches Museum Braunschweig, Germany; SMA, Sauriermuseum Aathal, Switzerland; Tate, Tate Geological Museum, Casper, Wyoming; YPM, Yale Peabody Museum, New Haven, Connecticut.
Acknowledgments
We thank Hans-Jakob Siber, Thomas Bolliger (SMA), Mark Norell, Carl Mehling and Alana Gishlick (AMNH) for the possibility to study and describe specimens under their care. Carl Mehling and Henry Galiano (New York, USA) shared pictures of previously undescribed material. Matthew Lamanna, Amy Henrici (CM), Ted Daeschler, Ned Gilmore (ANS), Ralf Kosma and Ulrich Joger (NMB) provided access to their collections. We greatly appreciate the fruitful discussions with Matthew Vickaryous (Univ. of Guelph, Canada) on interclavicular development, and with Regina Fechner (Univ. Bochum, Germany) on sauropodomorph gastralia and sternal ribs, and her comments on a first draft of this manuscript. We want to thank the editorial staff of the Journal of Anatomy, Julia Clarke and Edward Fenton for their help, as well as two anonymous reviewers for thoughtful and detailed reviews. Ricardo Araújo (South. Method. Univ., Dallas, TX, USA) and Jasmina Hugi (Winterthur, Switzerland) were of great help with finding literature on the embryological development of the various chest bones in reptiles, birds and mammals. Emanuele Minari (Turin, Italy) translated the description of the clavicle of Datousaurus in Dong & Tang (1984).
Emanuel Tschopp is supported by the Fundação para a Ciência e a Tecnologia doctoral fellowship SFRH/BD/66209/2009 (Ministério da Ciência, Tecnologia e Ensino superior, Portugal).
Supporting Information
Additional Supporting Information may be found in the online version of this article:
Fig. S1. Morphotype A element (interclavicle) AMNH 30900 in left lateral, internal and right lateral (from left to right). The left spur of the bifid anterior end as well as the distal posterior tip are lacking. Scale bar: 10 cm.
Fig. S2. Morphotype A element (interclavicle) SMA I 24-4 in right lateral, external, left lateral, internal (from left to right) and posterior view (bottom). Note the bifurcate anterior end towards the top of the picture. Scale bar: 10 cm.
Fig. S3. Morphotype A element (interclavicle) SMA L 22-3 in right lateral, external, left lateral and internal view (from left to right). Note the different orientation of the spatulate end compared with SMA I 24-4. Scale bar: 10 cm.
Fig. S4. Morphotype A element (interclavicle) SMA L 27-7 in left lateral, internal, right lateral (from left to right) and anterior view (top). Note the outwards curvature in the anterior half, probably due to taphonomy. Scale bar: 10 cm.
Fig. S5. Morphotype A element (interclavicle) SMA M 25-3 in left lateral, internal, right lateral (from left to right), anterior (top) and posterior view (bottom). Note the bifurcate anterior end. Scale bar: 10 cm.
Fig. S6. Morphotype B element (clavicle) SMA K 24-6 in internal, anterior, external, posterior (from left to right), lateral (top) and medial view (bottom). It is interpreted as left element of the SMA pair. Scale bar: 5 cm.
Fig. S7. Morphotype B element (clavicle) SMA K 24-3 in external (left), internal (right), lateral (top) and medial view (bottom). It is interpreted as right element of the SMA pair. Scale bar: 10 cm.
Fig. S8. Left morphotype B element (clavicle) of the pair included in AMNH 30789 in internal (left) and external view (right). The lateral-most end is lacking (top). Note the conspicuous ridge on the tip of the short leg of the L. Scale bar: 10 cm.
Fig. S9. Right morphotype B element (clavicle) of the pair included in AMNH 30789 in external (left) and internal view (right). Both the lateral-most end and the tip of the short leg of the L are lacking. Scale bar: 10 cm.
Fig. S10. Morphotype C element (sternal rib) SMA H 20-7. The end towards the top is broken. Scale bar: 10 cm.
Fig. S11. Morphotype C element (sternal rib) SMA L 21-5. The end towards the top is broken. Note the very slender and almost straight shaft. Scale bar: 10 cm.
Fig. S12. Morphotype D element (gastralia) SMA D 28-5. Note the irregular medial expansion (top) and the curved shaft. Scale bar: 10 cm.
Fig. S13. Morphotype D element (gastralia) SMA M 21-2. Note the irregular medial expansion (top) and the curved shaft. Scale bar: 10 cm.
Fig. S14. Two fused morphotype D elements (gastralia, SMA M 21-8). The lateral-most portion towards the bottom of the picture is broken, and would probably mirror the opposite end. Scale bar: 10 cm.
Fig. S15. Morphotype E element (sternal rib or intercostal element) SMA N 22-12. Scale bar: 10 cm.
Fig. S16. Morphotype E element (sternal rib or intercostal element) SMA M 21-15. Scale bar: 10 cm.
References
- Ayer J. The Howe Ranch Dinosaurs. Aathal, Switzerland: Sauriermuseum Aathal; 2000. [Google Scholar]
- Bader KS, Hasiotis ST, Martin LD. Application of forensic science techniques to trace fossils on dinosaur bones from a quarry in the Upper Jurassic Morrison Formation, Northeastern Wyoming. PALAIOS. 2009;24:140–158. [Google Scholar]
- Benton MJ, Walker AD. Erpetosuchus, a crocodile-like basal archosaur from the Late Triassic of Elgin, Scotland. Zool J Linn Soc. 2002;136:25–47. [Google Scholar]
- Brown B. Sinclair dinosaur expedition, 1934. Nat Hist. 1935;36:2–15. [Google Scholar]
- Carrano MT. The evolution of sauropod locomotion: morphological diversity of a secondarily quadrupedal radiation. In: Curry Rogers KA, Wilson JA, editors. The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press; 2005. pp. 229–251. [Google Scholar]
- Chatterjee S. A primitive parasuchid (phytosaur) reptile from the Upper Triassic Maleri Formation of India. Palaeontology. 1978;21:83–127. [Google Scholar]
- Chinnery BJ, Weishampel DB. Montanoceratops cerorhynchus (Dinosauria: Ceratopsia) and relationships among basal neoceratopsians. J Vertebr Paleontol. 1998;18:569–585. [Google Scholar]
- Claessens LPAM. Dinosaur gastralia; origin, morphology, and function. J Vertebr Paleontol. 2004;24:89–106. [Google Scholar]
- Claessens LPAM, O'Connor PM, Unwin DM. Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism. PLoS ONE. 2009;4:e4497. doi: 10.1371/journal.pone.0004497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark JM, Norell M, Chiappe LM. An oviraptorid skeleton from the late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest. Am Mus Novit. 1999;3265:1–36. [Google Scholar]
- Cooper MR. The prosauropod dinosaur Massospondylus carinatus Owen from Zimbabwe: its biology, mode of life and phylogenetic significance. Occas Pap Natl Monuments Rhod. 1981;6:690–840. [Google Scholar]
- Dilkes D, Sues H-D. Redescription and phylogenetic relationships of Doswellia kaltenbachi (Diapsida: Archosauriformes) from the Upper Triassic of Virginia. J Vertebr Paleontol. 2009;29:58–79. [Google Scholar]
- Dong Z, Tang Z. Note on a new mid-Jurassic sauropod (Datousaurus bashanensis gen. et sp. nov.) from Sichuan Basin, China. Vertebrata Pal Asiatica. 1984;22:69–75. [Google Scholar]
- Dong Z, Zhou S, Zhang Y. The dinosaurian remains from Sichuan basin, China. Palaeontologia Sinica NSC. 1983;23:139–145. [Google Scholar]
- Dong Z, Peng G, Huang D. The discovery of the bony tail club of sauropods. Vertebrata Pal Asiatica. 1989;27:219–224. [Google Scholar]
- Eaton TH, Stewart PL. A new order of fishlike Amphibia from the Pennsylvanian of Kansas. Univ Kans Mus Nat Hist. 1960;12:217–240. [Google Scholar]
- Filla BJ, Redman PD. Apatosaurus yahnahpin: a preliminary description of a new species of diplodocid dinosaur from the Late Jurassic Morrison Formation of Southern Wyoming, the first sauropod dinosaur found with a complete set of “belly ribs”. 1994;44:159–178. The Dinosaurs of Wyoming. Wyoming Geological Association 44th Annual Field Conference Guidebook. [Google Scholar]
- Galiano H, Albersdörfer R. Amphicoelias ‘brontodiplodocus’, a new sauropod, from the Morrison Formation, Big Horn Basin, Wyoming, with taxonomic reevaluation of Diplodocus, Apatosaurus, Barosaurus and other genera. Tensleep: Dinosauria International, LLC; 2010. [Google Scholar]
- Galton P, Jensen J. Skeleton of a hypsilophodontid dinosaur (Nanosaurusrex) from the Upper Jurassic of Utah. Brigham Young Univ Geol Studs. 1973;20:137–157. [Google Scholar]
- Gilmore CW. Osteology of Apatosaurus: with special reference to specimens in the Carnegie Museum. Mem Carnegie Mus. 1936;11:175–300. [Google Scholar]
- Haines RW. Epiphyses and sesamoids. In: Gans C, Bellairs Ad'A, Parsons TS, editors. Biology of the Reptilia, Vol. 1: Morphology A. London: Academic Press; 1969. pp. 81–115. [Google Scholar]
- Harris JD. The axial skeleton of Suuwassea emilieae (Sauropoda: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana, USA. Palaeontology. 2006;49:1091–1121. [Google Scholar]
- Harris JD. The appendicular skeleton of Suuwassea emilieae (Sauropoda: Flagellicaudata) from the Upper Jurassic Morrison Formation of Montana (USA) Geobios. 2007;40:501–522. [Google Scholar]
- Hatcher JB. Diplodocus (Marsh): its osteology, taxonomy, and probable habits, with a restoration of the skeleton. Mem Carnegie Mus. 1901;1:1–61. [Google Scholar]
- Hatcher JB. Additional remarks on Diplodocus. Mem Carnegie Mus. 1903;2:72–76. [Google Scholar]
- He X, Li K, Cai K. The Middle Jurassic dinosaur fauna from Dashanpu, Zigong, Sichuan. Vol. IV. Sauropod Dinosaurs (2) Omeisaurus tianfuensis. Chengdu, China: Sichuan Publishing House of Science and Technology; 1988. [Google Scholar]
- He X, Wang C, Liu S, et al. A new species of sauropod from the Early Jurassic of Gongxian County, Sichuan. Acta Geologica Sichuan. 1998;18:1–6. [Google Scholar]
- Hocknull SA, White MA, Tischler TR, et al. New mid-Cretaceous (latest Albian) dinosaurs from Winton, Queensland, Australia. PLoS ONE. 2009;4:e6190. doi: 10.1371/journal.pone.0006190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hohn-Schulte B. Form and Function of the Shoulder Girdle in Sauropod Dinosaurs: a Biomechanical Investigation with the Aid of Finite Elements (Dissertation) Bochum, Germany: Ruhr-Universität Bochum, Universitätsbibliothek; 2010. [Google Scholar]
- Holland WJ. The osteology of Diplodocus Marsh. Mem Carnegie Mus. 1906;2:225–264. [Google Scholar]
- Holland WJ. A new species of Apatosaurus. Ann Carnegie Mus. 1915;10:143–145. [Google Scholar]
- von Huene F. Vollständige Osteologie eines Plateosauriden aus dem schwäbischen Keuper. Geol Pal Abhandl. 1926;15:139–179. [Google Scholar]
- Ikejiri T, Tidwell V, Trexler DL. New adult specimens of Camarasaurus lentus highlight ontogenetic variation within the species. In: Carpenter K, Tidwell V, editors. Thunder-lizards: the Sauropodomorph dinosaurs. Bloomington: Indiana University Press; 2005. pp. 154–179. [Google Scholar]
- Klima M. Die Frühentwicklung des Schultergürtels und des Brustbeins bei den Monotremen (Mammalia: Prototheria) Adv Anat Embryol Cell Biol. 1973;47:1–78. [Google Scholar]
- Klima M. Early development of the shoulder girdle and sternum in marsupials (Mammalia: Metatheria) Adv Anat Embryol Cell Biol. 1987;109:1–91. doi: 10.1007/978-3-642-72994-2. [DOI] [PubMed] [Google Scholar]
- Makovicky PJ, Currie PJ. The presence of a furcula in tyrannosaurid theropods, and its phylogenetic and functional implications. J Vertebr Paleontol. 1998;18:143–149. [Google Scholar]
- Marsh OC. Principal characters of American Jurassic Dinosaurs. Restoration of Brontosaurus. Geol Mag. 1883;10:385–388. [Google Scholar]
- Marsh OC. The dinosaurs of North America. Washington: United States Geological Survey; 1896. [Google Scholar]
- Martínez RN. Adeopapposaurus mognai, gen. et sp. nov. (Dinosauria: Sauropodomorpha), with comments on adaptations of basal Sauropodomorpha. J Vertebr Paleontol. 2009;29:142–164. [Google Scholar]
- Martz JW. The Morphology and Ontogeny of Typothorax Coccinarum (Archosauria, Stagonolepididae) from the Upper Triassic of the American Southwest (Master thesis) Lubbock, TX, USA: Graduate Faculty, Texas Tech University; 2002. [Google Scholar]
- McIntosh JL, Miles CA, Cloward KC, et al. A new nearly complete skeleton of Camarasaurus. Bull. Gunma Mus. Natu. Hist. 1996;1:1–87. [Google Scholar]
- Michelis I. Taphonomie des Howe Quarry's (Morrison-Formation, Oberer Jura), Bighorn County, Wyoming, USA (Dissertation) Bonn, Germany: Institute of Palaeontology, University of Bonn; 2004. [Google Scholar]
- Nesbitt SJ. The early evolution of archosaurs: relationships and the origin of major clades. Bull Am Mus Nat Hist. 2011;352:1–292. [Google Scholar]
- Nesbitt SJ, Turner AH, Spaulding M, et al. The theropod furcula. J Morphol. 2009;270:856–879. doi: 10.1002/jmor.10724. [DOI] [PubMed] [Google Scholar]
- Nopcsa FB. Remarks on the supposed clavicle of the sauropodous dinosaur Diplodocus. J Zool. 1905;2:289–294. [Google Scholar]
- Novas FE. Dinosaur monophyly. J Vertebr Paleontol. 1996;16:723–741. [Google Scholar]
- Osborn HF, Mook CC. Camarasaurus, Amphicoelias, and other sauropods of Cope. Bull Am Mus Nat Hist. 1921;3:249–387. [Google Scholar]
- Parks WA. Thescelosaurus warreni, a new species of orthopodous dinosaur from the Edmonton Formation of Alberta. Can Mineral. 1926;21:1–42. [Google Scholar]
- Remes K. Evolution of the Pectoral Girdle and Forelimb in Sauropodomorpha (Dinosauria, Saurischia) (Dissertation) München, Germany: Fakultät für Geowissenschaften, LMU München; 2008. [Google Scholar]
- Remes K, Ortega F, Fierro I, et al. A new basal sauropod dinosaur from the Middle Jurassic of Niger and the early evolution of Sauropoda. PLoS ONE. 2009;4:e6924. doi: 10.1371/journal.pone.0006924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rieppel O. Studies on skeleton formation in reptiles. III. Patterns of ossification in the skeleton of Lacerta vivipara Jacquin (Reptilia, Squamata) Fieldiana Life Earth Sci. 1992;68:1–25. [Google Scholar]
- Rieppel O. Studies on skeleton formation in reptiles. V. Patterns of ossification in the skeleton of Alligator mississippiensis Daudin (Reptilia, Crocodylia) Zool J Linn Soc. 1993;109:301–325. [Google Scholar]
- Romer A. Osteology of the Reptiles. Chicago: University of Chicago Press; 1956. [Google Scholar]
- Sander PM, Christian A, Clauss M, et al. Biology of the sauropod dinosaurs: the evolution of gigantism. Biol Rev. 2011;86:117–155. doi: 10.1111/j.1469-185X.2010.00137.x. doi: 10.1111/j.1469-185X.2010.00137.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwarz D, Frey E, Meyer CA. Novel reconstruction of the orientation of the pectoral girdle in sauropods. Anat Rec (Hoboken) 2007a;290:32–47. doi: 10.1002/ar.20405. [DOI] [PubMed] [Google Scholar]
- Schwarz D, Ikejiri T, Breithaupt BH, et al. A nearly complete skeleton of an early juvenile diplodocid (Dinosauria: Sauropoda) from the Lower Morrison Formation (Late Jurassic) of north central Wyoming and its implications for early ontogeny and pneumaticity in sauropods. Hist Biol. 2007b;19:225–253. [Google Scholar]
- Sereno PC. Basal archosaurs: phylogenetic relationships and functional implications. J Vertebr Paleontol. 1991;11:1–53. [Google Scholar]
- Sereno PC, Beck AL, Dutheil DB, et al. Cretaceous sauropods from the Sahara and the uneven rate of skeletal evolution among dinosaurs. Science. 1999;286:1342–1347. doi: 10.1126/science.286.5443.1342. [DOI] [PubMed] [Google Scholar]
- Steyer J-S, Heyler D, Guillot L. Nouvelles données sur les stégocéphales du bassin de Souvigny. Revue Scientifique du Bourbonnais et du Centre de la France. 2000;1998:80–88. [Google Scholar]
- Tschopp E, Mateus O. A sternal plate of a large-sized sauropod dinosaur from the Late Jurassic of Portugal. Fundamental. 2012;20:263–266. [Google Scholar]
- Tschopp E, Mateus O. The skull and neck of a new flagellicaudatan sauropod from the Morrison Formation and its implication for the evolution and ontogeny of diplodocid dinosaurs. J Syst Paleontol. (in press), in press. [Google Scholar]
- Vickaryous MK, Hall BK. Homology of the reptilian coracoid and a reappraisal of the evolution and development of the amniote pectoral apparatus. J Anat. 2006;208:263–285. doi: 10.1111/j.1469-7580.2006.00542.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vickaryous MK, Hall BK. Comparative development of the crocodylian interclavicle and avian furcula, with comments on the homology of dermal elements in the pectoral apparatus. J Exp Zool A Ecol Genet Physiol. 2010;314:196–207. doi: 10.1002/jez.b.21326. [DOI] [PubMed] [Google Scholar]
- Weishampel DB, Heinrich R. Systematics of Hypsilophodontidae and basal Iguanodontia (Dinosauria: Ornithopoda) Hist Biol. 1992;6:159–184. [Google Scholar]
- Wilson JA, Carrano MT. Titanosaurs and the origin of “wide-gauge” trackways: a biomechanical and systematic perspective on sauropod locomotion. Paleobiology. 1999;25:252–267. [Google Scholar]
- Xing L, Ye Y, Shu C, et al. Structure, orientation, and finite element analysis of the tail club of Mamenchisaurus hochuanensis. Acta Geol Sinica. 2009;83:1031–1040. [Google Scholar]
- Yates AM, Vasconcelos CC. Furcula-like clavicles in the prosauropod dinosaur Massospondylus. J Vertebr Paleontol. 2005;25:466–468. [Google Scholar]
- Zhang Y. The Middle Jurassic dinosaur fauna from Dashanpu, Zigong, Sichuam, vol. 1: sauropod dinosaur (I): Shunosaurus. Chengdu, China: Sichuan Publishing House of Science and Technology; 1988. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.