

Abstract
Bilinguals have been shown to outperform monolinguals at suppressing task-irrelevant information. The present study aimed to identify how processing linguistic ambiguity during auditory comprehension may be associated with inhibitory control. Monolinguals and bilinguals listened to words in their native language (English) and identified them among four pictures while their eye-movements were tracked. Each target picture (e.g., hamper) appeared together with a similar-sounding within-language competitor picture (e.g., hammer) and two neutral pictures. Following each eye-tracking trial, priming probe trials indexed residual activation of target words, and residual inhibition of competitor words. Eye-tracking showed similar within-language competition across groups; priming showed stronger competitor inhibition in monolinguals than in bilinguals, suggesting differences in how inhibitory control was used to resolve within-language competition. Notably, correlation analyses revealed that inhibition performance on a nonlinguistic Stroop task was related to linguistic competition resolution in bilinguals but not in monolinguals. Together, monolingual-bilingual comparisons suggest that cognitive control mechanisms can be shaped by linguistic experience.
Keywords: Bilingualism, Inhibition, Language comprehension, Eye-tracking, Negative priming
1. Introduction
Detection and resolution of ambiguity is a core element of language processing. For example, during monolingual language comprehension, competition between linguistic alternatives arises in the presence of multiple similar-sounding words (e.g., Bradlow & Pisoni, 1999; Desroches, Newman, & Joanisse, 2008; Luce & Pisoni, 1998; Marslen-Wilson, 1987; McClelland & Elman, 1986) and multiple related word meanings (e.g., Degani & Tokowicz, 2010; Gernsbacher, 1990; Gernsbacher & Faust, 1991; Seidenberg, Tanenhaus, Leiman, & Bienkowski, 1982; Swinney, 1979). Resolution of linguistic competition may require cognitive control to focus on relevant information in the face of copeting alternatives. In general, a link between linguistic performance and cognitive control abilities has been established across a range of language processing contexts. Cognitive control has been tied to language performance both in children, where the cognitive system is developing (Nakamichi, 2007; Smith, Jones, Landau, Gershkoff-Stowe, & Samuelson, 2002) and in older adults, where it is in decline (Kemper & Sumner, 2001; Kwong See & Ryan, 1995; Taylor, O'Hara, Mumenthaler, Rosen & Yesavage, 2005). In addition, when language processing demands are high, more executive control is engaged (Kerns, 2007; Thompson-Schill, D'Esposito, Aguirre, & Farah, 1997; Thompson-Schill, D'Esposito, & Kan, 1999), and better cognitive control is associated with better linguistic performance (Christoffels, De Groot, & Kroll, 2006; Hernandez & Meschyan, 2006).
One way to examine the link between linguistic experience and domain-general cognitive function is to compare groups whose different experiences in the linguistic domain may have influenced their performance in other cognitive realms. The present research focuses on how linguistic experience may change cognitive function in bilinguals compared to monolinguals. The study has two objectives: (1) to investigate whether an inhibition mechanism is involved in auditory word comprehension in monolinguals and bilinguals, and (2) to examine the nature of such an inhibition mechanism. It was hypothesized that, if extended immersion in bilingual (high-ambiguity) language comprehension situations1 honed inhibition mechanisms that resolve competition during auditory comprehension, then bilinguals would exert cognitive control more efficiently than monolinguals. Further, if domain-general cognitive control processes were differentially involved in monolingual and bilingual language comprehension, then the relationship between word recognition and a nonlinguistic inhibitory task would differ across the two groups. More specifically, if bilingual experience resulted in recruitment and modulation of domain-general cognitive control processes, then bilinguals would show stronger relationships between inhibition during word recognition and inhibition on the nonlinguistic task, as compared to monolinguals.
Bilingual language processing involves simultaneous activation of two languages (e.g., Blumenfeld & Marian, 2007; Marian & Spivey, 2003a, 2003b) and requires an ability to efficiently control these languages (e.g., Dijkstra & van Heuven, 1998; Green, 1998) and switch from one language to the other in a context-appropriate manner (e.g., Costa & Santesteban, 2004; Rodriguez-Fornells, Balaguer, & Munte, 2006). During auditory comprehension, similar-sounding words become active across bilinguals' two languages, so that when bilinguals hear a word in one language while presented with pictures, they also look at pictures of between-language competitors that overlap phonologically across languages (e.g., when hearing marker in English, Russian-English bilinguals make eye-movements to a stamp, the Russian word for which is marka, Marian & Spivey, 2003a, 2003b). This finding has been replicated across different groups of bilinguals and language pairs (Blumenfeld & Marian, 2007; Canseco-Gonzales, Brick, Fischer, & Wagner, 2005; Cutler, Weber, & Otake, 2006; Ju & Luce, 2004; Shook & Marian, under review; Weber & Cutler, 2004; Weber & Paris, 2004), suggesting that bilinguals' experience with parallel language activation and cross-linguistic competition is universal and occurs in addition to the within-language co-activation and competition experienced by monolinguals (e.g., Luce & Pisoni, 1998; Marslen-Wilson, 1987; McClelland & Elman, 1986).
As a consequence of bilinguals' parallel language activation, bilinguals may face consistently higher degrees of linguistic competition compared to monolinguals and, as a result, their performance on executive control tasks may be altered (e.g., Bialystok, 2005, chap. 20; Kroll, 2008), with bilinguals showing cognitive advantages over monolinguals (e.g., Bialystok, 2005, chap. 20; Colzato, Bajo, van den Wildenberg, & Paolieri, 2008; Cook, 1997; Costa, Hernandez, & Sebastian-Galles, 2008; Prior & MacWhinney, 2010). For example, Bialystok and Codd (1997) compared monolingual and bilingual children on two selective-attention tasks where participants had to ignore irrelevant and conflicting information (e.g., to identify a tower containing more blocks, participants had to ignore a higher tower that actually contained fewer blocks), compared to tasks that contained no conflicting information. They found that bilingual children performed better than monolingual children on the task containing irrelevant conflicting information, but performed the same as the monolingual children on the task containing no conflicting information (for similar findings comparing bilingual and monolingual children, see Bialystok, 1999, 2010; Bialystok & Martin, 2004; Bialystok & Viswanathan, 2009). Similar bilingual advantages in inhibitory control, conflict monitoring, and task switching have been found across the lifespan (Bialystok, 2006, 2010; Bialystok, Craik, Klein, & Viswanathan, 2004; Colzato et al., 2008; Costa, Hernandez, Costa, & Sebastian-Galles, 2009; Costa et al., 2008; Kovacs & Mehler, 2009; Prior & MacWhinney, 2010).
The demands of bilingual language processing are the likely source of cognitive advantages in bilinguals (e.g., Bialystok, 2005, chap. 20; Kroll, 2008). Language processes that require greater cognitive demands in bilinguals include language switching (Prior & MacWhinney, 2010) and continued suppression of a second language during production (Linck, Hoshino, & Kroll, 2008; Linck, Kroll, & Sunderman, 2009). While parallel language activation during comprehension has been cited as a potential source of bilingual cognitive advantages (e.g., Kroll, 2008), a direct link between comprehension processes and cognitive control in bilinguals has not yet been established. Findings that identify a link between linguistic and nonlinguistic control processes would provide empirical support for the hypothesis that linguistic competition is at the root of bilingual cognitive advantages previously identified in the literature (e.g., Bialystok, 2005, chap. 20; Costa et al., 2008).
The aim of the present study was to identify a link between monolinguals' and bilinguals' ambiguity resolution during auditory comprehension and their performance on a nonlinguistic task of cognitive control. Monolingual and bilingual listeners completed a combined Eye-Tracking/Negative Priming task (Blumenfeld, 2008) that indexed both activation of multiple word candidates during auditory comprehension and subsequent suppression of irrelevant competing words (see Fig. 1). Participants heard object names, and were asked to identify these target objects from a set of items in a visual display while their eye-movements were tracked (e.g., Tanenhaus, Spivey-Knowlton, Eberhard, & Sedivy, 1995). During target identification, participants' eye-movements to target and competitor pictures reflected parallel activation of both items. In order to examine whether inhibitory control processes were differentially involved in monolingual and bilingual language comprehension, monolinguals' and bilinguals' ability to resolve competition was compared during auditory word comprehension in their native language (English).
Fig. 1.
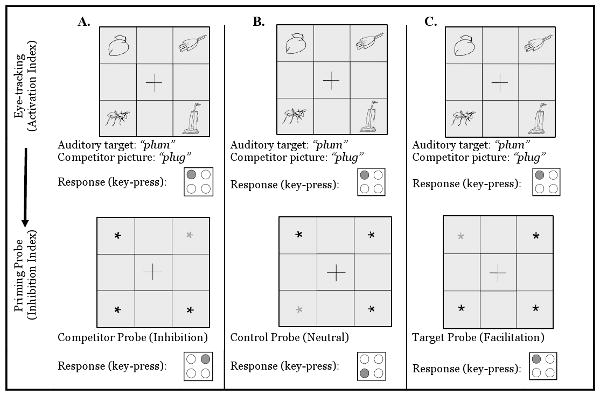
The Eye-Tracking/Negative Priming paradigm. To probe activation of similar-sounding words, participants heard words and identified corresponding pictures in the presence of phonological competitor pictures while their eye-movements were tracked (top panel). To probe inhibition of competitor words, Word Recognition trials were followed by priming probe trials (bottom panel) that probed inhibition of competitor items (grey asterisks in locations previously occupied by competitor pictures, column A), relative to control items (grey asterisks in locations previously occupied by control pictures, column B), and target items (grey asterisks in locations previously occupied by target pictures, column C). Participants responded by pressing one of four keys, arranged in a square corresponding to the location of items on the stimulus display (response row).
To index inhibition of similar-sounding competitor words that became co-activated during comprehension, we added a negative priming component to follow each eye-tracking trial. Negative priming is a unique way to covertly capture inhibition of conflicting information by indexing its influence on subsequent processing (e.g., Connelly & Hasher, 1993; Milliken, Lupianez, Debner, & Abello, 1999; Simone, Ahrens, Foerde, & Spinetta, 2006; Tipper, 1985). Participants respond to target stimuli that are presented together with conflicting competitors (such as on the eye-tracking trials). On the next trial, inhibition of competitors from the previous trial is probed by making the previous competitor the target of response. If the response on this priming probe trial is delayed, then a negative priming effect is identified, suggesting previous inhibition of the competitor (for a review, see May, Kane, & Hasher, 1995). Negative priming paradigms may rely on location-priming (Connelly & Hasher, 1993; Simone et al., 2006). Research on spatial attention reveals lengthened response times for targets that appear in locations previously occupied by competitors, and this longer response time is believed to be due to a location-specific inhibitory mechanism (e.g., May et al., 1995; Simone et al., 2006). A location-based negative priming paradigm was employed in the present study, probing residual activation or inhibition of locations that had been occupied previously by pictures of target words or similar-sounding competitor words. The negative priming paradigm has been widely used in nonlinguistic settings (e.g., Connelly & Hasher, 1993; Simone et al., 2006), and provides a unique tool since it can be used to index inhibition in trials that precede it.
Participants also completed a nonverbal variant of the Stroop task that indexed their ability to inhibit perceptual conflict in nonlinguistic input (e.g., a right-pointing arrow on the left side of the display, Liu, Banich, Jacobson, & Tanabe, 2004). This task was selected because bilinguals' advantages have previously been identified for Stroop-type inhibition (e.g., Bialystok & Codd, 1997; Costa et al., 2008), and because cognitive processes involved in performance on the Stroop task – goal maintenance and attention allocation (Kane & Engle, 2003) – closely resemble the processes involved in language comprehension. On the Stroop task, multiple salient stimulus dimensions (arrow direction and arrow location) are activated at the same time and map onto either a single or multiple responses (Kornblum, 1992, 1994). Similarly, during the auditory comprehension task, multiple perceptual dimensions become available and can map onto different responses. For example, the acoustic speech-signal of the word mouse could initially also map onto mouth. Inclusion of the non-linguistic Stroop task that resembles the word comprehension task in attentional demands made it possible to conduct correlation analyses between nonlinguistic and language-related inhibition. Such correlations would speak to whether a nonlinguistic inhibition mechanism may be recruited for language comprehension. In addition, mapping efficient performance (i.e., small Stroop effects) in the nonlinguistic domain to performance in the linguistic domain informs the nature of cognitive control in language comprehension. Specifically, a link between linguistic and nonlinguistic inhibition mechanisms would suggest that nonlinguistic cognitive processes can be influenced by linguistic experience. Finally, if correlations between nonlinguistic and linguistic inhibition differ across monolinguals and bilinguals, then it can be concluded that inhibition mechanisms may be modulated by bilingual experience.
2. Methods
2.1. Participants
Thirty English-native monolinguals (Mage = 21.4, SD = 3.9; six males) and 30 English-native bilinguals, who also spoke Spanish (Mage = 22.0, SD = 5.1; nine males), were recruited. Monolingual and bilingual groups were matched on age, t(58) = 0.5, p > .5, digit span [monolinguals: M = 17.5, SD = 2.1; bilinguals: M = 17.6, SD = 2.5; t(58) = 0.2, p > .5], English receptive vocabulary [monolinguals: M = 116.7, SD = 11.7; bilinguals: M = 116.2, SD = 12.2; t(58) = 0.1, p > .5], and nonverbal IQ [monolinguals: M = 110.5, SD = 11.8; bilinguals: M = 110.0, SD = 11.7; t(58) = 0.2, p > .5]. Bilinguals were selected to have extensive Spanish experience (as determined by acquisition of Spanish before age 8, M = 2.4, SD = 2.9) as well as substantial current exposure to Spanish (M = 21.9% of the time, SD = 14.7%). All participants were administered the Language Experience and Proficiency Questionnaire (LEAP-Q, Marian, Blumenfeld, & Kaushanskaya, 2007). Monolinguals reported speaking only English and having only minimal exposure to foreign languages (M = 1.8% of the time, SD = 2.7%). Bilingual English–Spanish participants who spoke English as a native language rated their Spanish proficiency at an average of 7.7 on a scale from 0 to 10 (SE = 1.1) across comprehension, speaking, and reading modalities. Monolinguals (M = 9.6, SD = 0.5) and bilinguals (M = 9.4, SD = 0.7) did not differ on their self-reported English proficiency across comprehension, speaking, and reading modalities, t(58) = 1.4, p >.1.
2.2. Materials
The experimental paradigm consisted of two types of trials (see Fig. 1): Word Recognition/Eye-tracking trials that indexed activation of competitor words and control words during word recognition, and Priming Probe trials that indexed inhibition of preceding competitor words, relative to control words. Priming Probe trials immediately followed Word Recognition/Eye-tracking trials, and contained grey asterisks that occurred in the locations previously occupied by pictures of competitor, control, or target words. On Word Recognition/Eye-tracking trials, more looks made to pictures of competitor vs. control words would signify that the competitor word was activated and competed during target selection. On Priming Probe trials, longer identification latencies of priming probes in locations previously occupied by competitor pictures vs. locations previously occupied by control pictures would signify that the competitor word had been inhibited.
For Word Recognition trials, stimulus displays included four pictures and a central fixation cross (see Fig. 1, top row). The four pictures in each display included: (1) a target word, (2) a competitor or control word, and (3 and 4) two control words. In total, 234 Word Recognition trials were presented. In one third of these trials (n = 78), target words were paired with competitor words that overlapped phonologically. For example, when the target word was cab, the competitor word was cat. To minimize awareness of phonological overlap, the ratio of competitor trials to control trials was set at 1:2 (Botvinick, Braver, Barch, Carter, & Cohen, 2001; Green, 1998; Henik, Bibi, Yanai, & Tzelgov, 1997). In the remaining two-thirds of Word Recognition trials (i.e., control Word Recognition trials, n = 156), target words were paired with three neutral control words. To avoid looks due to semantic co-activation (Huettig & Altmann, 2005; Yee & Sedivy, 2006), the four stimuli in each trial were not semantically related to each other.
To maximize involvement of cognitive control processes during participation, two measures were taken to ensure strong co-activation of competitor words. First, target-competitor pairs had high phonological overlap. This overlap consisted of at least two phonemes shared at the onset of the word, and duration of acoustic overlap (as measured by a native speaker of English) averaged 279.4 ms (SE = 20.8) across target-competitor pairs. Second, based on findings by Dahan, Magnusson, and Tanenhaus (2001), competitors were selected to be of higher word frequency (M = 91.9, SE = 16.9) than targets (M = 15.7, SE = 2.3), t(154) = 4.5, p < .001, as calculated using the CELEX word frequency database (Baayen, Piepen-brock, & van Rijn, 1995). Targets and competitors were balanced on neighborhood2 density (targets: M = 13.2, SD = 8.9; competitors: M = 11.7, SD = 8.7) and neighborhood frequency (targets: M = 1.9, SD = 0.5; competitors: M = 1.9, SD = 0.4), all ps > 0.5.
Concurrent with picture displays, single words were aurally presented. Auditory stimuli were pre-recorded in a sound-proof booth (44,100 Hz, 16 bits) by a native female speaker of American English using a Marantz Solid State recorder. Normalization, segmentation and insertion of equal between-word breaks were performed using Praat and Sound Studio software. During the experiment, the name of the target picture was aurally presented 500 ms after onset of the picture display (e.g., Ju & Luce, 2004; Weber & Cutler, 2004). Picture stimuli were selected from a previous study (Blumenfeld & Marian, 2007), the International Picture Naming Database (Székely et al., 2004) and the IMSI Master Clips database. Pictures were black linedrawings, and were manipulated in Photoshop to have similar line thickness. Positioning of target and competitor pictures in the four display quadrants was counterbalanced across conditions.
To combine Word Recognition trials with the three types of priming probe trials (target probes, competitor probes, and control probes), the 78 Word Recognition trials with competitors were divided into three lists of 26 stimuli. Across these three lists, stimuli were matched for word frequencies of targets, F(2, 75) = 0.2, p > .5, and competitors, F(2, 75) = 0.2, p > .5, neighborhood size of targets, F(2, 75) = 0.1, p > .5, and competitors, F(2, 75) = 0.5, p > .5, neighborhood frequency of targets, F(2, 75) = 0.2, p > .5, and competitors, F(2, 75) = 0.5, p > .5, and duration of acoustic overlap between targets and competitors, F(2, 75) = 0.02, p > .5.
On Priming Probe trials, negative location priming was used as an index of competitor word inhibition on preceding Word Recognition trials (e.g., Simone et al., 2006). A Priming Probe trial was presented 500 ms after each Word Recognition trial. As in Word Recognition trials, displays of Priming Probe trials featured four quadrants. One of these quadrants contained a grey asterisk, and the remaining three quadrants contained black asterisks. To ensure that inhibition of preceding lexical competitors was reliably indexed, three types of priming probe trials were employed: (1) control probe trials, (2) competitor probe trials, and (3) target probe trials. On control probe trials, the grey asterisk appeared in the same location as a control picture on the preceding Word Recognition trial. Since control pictures on Word Recognition trials were likely to be minimally attended to and minimally inhibited, response latencies on control probe trials acted as a baseline. On competitor probe trials, the grey asterisk appeared in the same location as the competitor picture in the previous Word Recognition trial. Since competitor pictures on Word Recognition trials were likely to be only briefly and partially attended to and then inhibited, response latencies on competitor probe trials reflected location inhibition, and were expected to be longer relative to control probe trials. Finally, on target probe trials, the grey asterisk appeared in the same location as the target on the preceding Word Recognition trial. Since target pictures on Word Recognition trials were likely to be maximally attended to and fully activated, response latencies on target probe trials reflected location facilitation, and were expected to be shorter relative to control probe and competitor probe trials. All control Word Recognition trials were followed by filler priming probe trials that had grey asterisks assigned to each quadrant an equal number of times. Of the 234 Priming Probe trials, 26 trials were control probe trials, 26 trials were competitor probe trials, 26 trials were target probe trials, and 156 trials were filler trials. For a summary of conditions and trial types in the Eye-Tracking/Negative Priming paradigm, see Fig. 2. To reduce effects due to stimulus characteristics, pairings between the 26 control probe, competitor probe, and target probe trials and the three lists of Word Recognition trials were counterbalanced across participants so that each set of Word Recognition trials was paired with each type of Priming Probe trial an equal number of times.
Fig. 2.
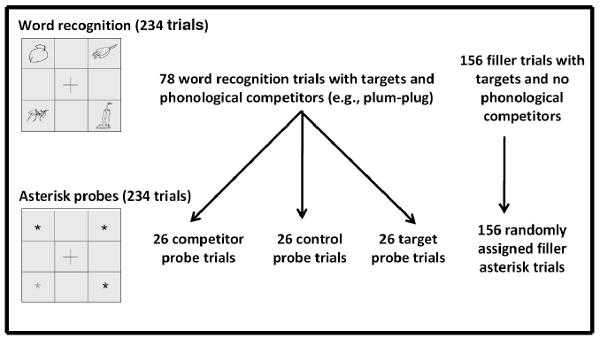
Design summary of the Eye-Tracking/Negative Priming paradigm.
2.2.1. Nonlinguistic inhibitory control
On the Stroop task (Stroop, 1935, adapted for the present study from Liu et al., 2004), cognitive conflict was created by manipulating two stimulus dimensions – arrow direction and arrow location – to be either congruent or incongruent. Participants were asked to respond to arrow direction but to ignore location. They were instructed to click a left button when they saw a leftward-facing arrow and a right button when they saw a rightward-facing arrow, while ignoring the location of the arrow. Sixty congruent trials contained a leftward-facing arrow presented to the left of the central fixation cross, and 60 contained a rightward-facing arrow presented to the right of the central fixation cross. Twenty incongruent trials contained a leftward-facing arrow presented to the right of the central fixation cross, and 20 contained a rightward-facing arrow presented to the left of the central fixation cross. The ratio of incongruent to congruent trials was maintained at 1:3, and each trial started with a 500 ms central fixation cross (to call participants' attention towards the middle of the screen), followed by a 700 ms presentation of the congruent or incongruent stimulus display, and an 800 ms presentation of a blank screen. All trials were presented in a fixed pseudo-randomized order, with an equal number of “switch trials” in the congruent and incongruent conditions.
2.3. Procedure
After completing 25 practice trials, participants listened to auditory stimuli and viewed pictures placed in four quadrants of a visual display. They identified the quadrant containing the target they heard by pressing one of four keys. Immediately following each of the Word Recognition trials, participants were presented with a Priming Probe trial. They identified the quadrant containing the grey asterisk by pressing one of the same four keys. The response keys were arranged in a square pattern, corresponding to the location of pictures and asterisks on the stimulus display. Participants used their two index fingers (upper right and left quadrants) and two thumbs (lower right and left quadrants) to respond. Participants were then presented with the nonlinguistic Stroop task. After the experimental tasks, participants were administered the nonverbal components of the Wechsler Abbreviated Scale of Intelligence (WASI, PsychCorp, 1999), the digit span component of the Comprehensive Test of Phonological Processing (CTOPP, Wagner, Torgesen, & Rashotte, 1999), the Peabody Picture Vocabulary Test (Dunn & Dunn, 1997), the Language Experience and Proficiency Questionnaire (LEAP-Q) and, if they were bilingual, the Test de Vocabolario en Imagenes Peabody (Dunn, Padilla, Lugo, & Dunn, 1986).
2.4. Data coding
Across each Word Recognition trial, eye-movements to target, competitor, and control items were coded as 1 (look) or 0 (no look) within each 33 ms window starting at the onset of the target word, and ending with the participant's response via button press. The participant had to fixate an object for at least one 33 ms frame for the look to be coded. In addition, across the duration of each trial, the total proportion of looks was calculated for targets, competitors and the averaged two controls. Since eye-movement planning takes approximately 200 ms (Hallett, 1986), timecourse analyses focused on activation beyond the initial 200 ms. Activation time curves were derived by plotting the proportion of target, competitor, and control fixations (across trials and participants) over time in milliseconds post-stimulus onset. An independent coder re-coded 15% of all data, yielding high inter-coder reliability (Pearson pair-wise correlations, r = .93, p < .001). For each Priming Probe trial, response latencies for identifying the grey asterisk were captured using Superlab software. For each participant, response latencies below 200 ms, or three standard deviations above or below the mean, were removed, resulting in omission of 3.7% of data. An additional 3.4% of data were omitted due to incorrect responses.
3. Results
Results are presented for: (1) lexical activation during word recognition, (2) inhibition during negative priming, and (3) performance on the nonlinguistic Stroop task and correlations between linguistic and nonlinguistic inhibition.
3.1. Lexical activation
Target identification accuracy and latency were examined using 2 × 2 ANOVAs with trial type (competitors, controls) as a within-subjects variable and with group (monolingual, bilingual) as a between-subjects variable. As expected, both groups identified target words less accurately in the presence of similar-sounding competitors (M = 95.3%, SE = 0.4%) than in their absence (M = 98.7%, SE = 0.2%), F(1, 58) = 61.8, p < .001, and took longer to identify the targets when competitors were present (M = 2040.0, SE = 32.8) than when they were absent (M = 1905.0, SE = 27.8), F(1, 58) = 204.0, p<.001, . No other significant effects or interactions were found, p>.1, suggesting that monolinguals and bilinguals were equally efficient at identifying target words when withinlanguage competitors were present.
The timecourse of target, competitor, and control activation for monolingual and bilingual participants was examined between 200 and 2600 ms post-stimulus onset (see Fig. 3). For both monolingual and bilingual participants, significant differences between competitor and control activation were found between 200 ms and 933 ms post-stimulus onset (all 2-tailed ps < .05, F(1, 58) = 192.3, p < .001, . Within this time-window, participants looked at competitors on average 18.7% (SE = 0.8%) of the time and at controls on average 12.5% (SE = 0.5%) of the time, consistent with within-language co-activation effects found in previous eye-tracking studies (e.g., Marian & Spivey, 2003b). No main effect of group, or interaction between condition and group were found, p > .5. Once the target activation peak had been reached, bilinguals maintained the target active longer than monolinguals, t(58) = 1.7, p < .05 (1-tailed), as illustrated by more fixations to the target in bilinguals (16.7%, SE = 1.2) than in monolinguals (14.0%, SE = 1.0) between 1099 ms and 2734 ms post-stimulus onset.
Fig. 3.

Activation timecourse of English words, and their similar-sounding English competitors, for monolinguals (A) and bilinguals (B).
3.2. Priming probe results
Priming effects were examined using a 3 × 2 ANOVA, with trial type (control probe, competitor probe, target probe) as a within-subjects variable, and with group (monolingual, bilingual) as a between-subjects variable. Accuracy rates (M = 99.7%, SE = 0.1) did not differ across groups or trial types, p > .1. For reaction time, an interaction was found between trial type and group, F(1, 58) = 10.7, p < .01, (see Fig. 4). Follow-up ANO-VAs compared response latencies on control probes to response latencies on target probes and to response latencies on competitor probes. As predicted based on previous location priming studies, for targets, results revealed that participants identified priming probes in positions previously occupied by target pictures (M = 467.8, SE = 8.4) faster than priming probes in positions previously occupied by control pictures, (M = 496.2, SE = 8.3), F(1, 58) = 30.1, p < .001, (no interaction was found, p > .5). Confirming predictions that inhibitory control differences would emerge between monolinguals and bilinguals, for competitors, a significant interaction between trial type and group, F(1, 58) = 10.7, p < .01, was found. Follow-up t-tests revealed that monolinguals identified priming probes in positions previously occupied by competitor pictures (M = 505.5, SE = 14.3) slower than priming probes in positions previously occupied by control pictures (M = 485.5, SE = 12.9), t(29) = 4.3, p < .001, consistent with expectations that an inhibition effect would be identified. Bilinguals did not differ between the two conditions, p > .5, suggesting that, inconsistent with expectations, monolinguals but not bilinguals showed inhibition of previously-activated competitor words.
Fig. 4.
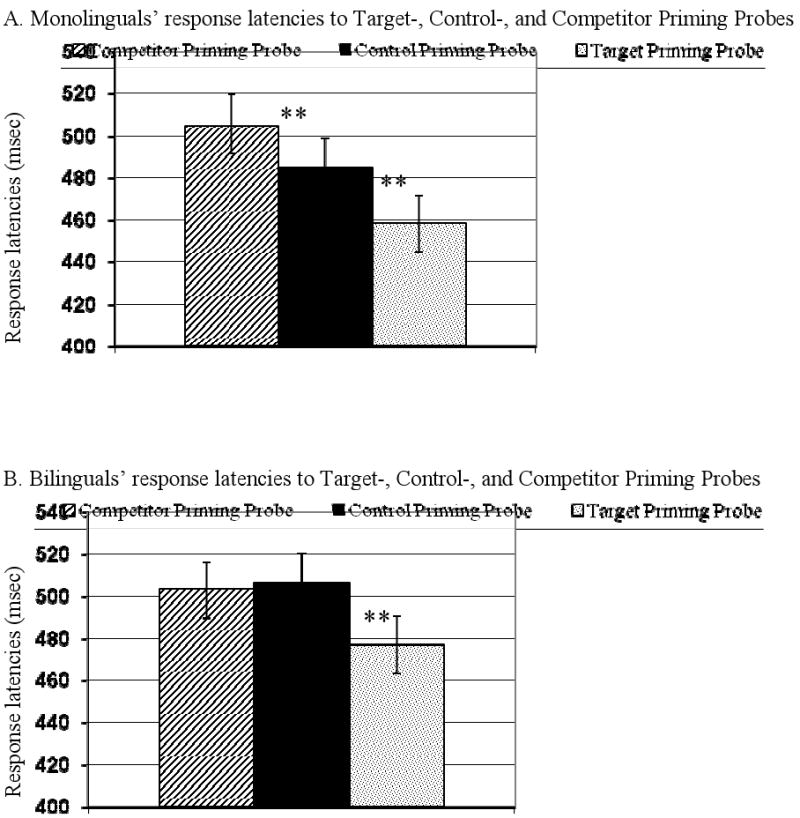
Response latencies to competitor-, control-, and target priming probes (presented as grey asterisks in positions previously occupied by competitor-, control-, and target pictures, respectively) for monolinguals (A) and bilinguals (B).
Two possible inhibition mechanisms may account for the finding that bilinguals and monolinguals differ when inhibiting previously-activated competitors. First, it is possible that inhibition effects are due to negative location priming, with attentional access to competitor locations inhibited. Second, it is possible that effects are due to inhibition at the response level, with motor responses to competitor pictures inhibited. To examine whether group differences on negative-priming trials could be localized to inhibition at the response level or to inhibition at an earlier processing stage, we examined launch latencies of eye-movements to grey asterisks. If findings of competitor inhibition were due to inhibition during early processing stages, then we would expect effects to be reflected in different launch latencies to target-, competitor-, and filler-probes. Conversely, if the effect were due to inhibition at the response level, then no differences would be observed in the launch latencies of eye-movements. For targets, results revealed that all participants were quicker to launch eye-movements to priming probes in positions previously occupied by target pictures (M = 243.5, SE = 6.1) than to priming probes in positions previously occupied by control pictures, (M = 272.1, SE = 7.0), F(1, 57) = 23.5, p<.001, . For competitors, monolinguals were slower to launch eye-movements to priming probes in positions previously occupied by competitor pictures (M = 283.6, SE = 10.1) than to priming probes in positions previously occupied by control pictures (M = 269.8, SE = 9.4), t(29) = 2.5, p < .05. Bilinguals did not differ between the two conditions, p > .5,3 suggesting that monolinguals' but not bilinguals' eye-movement launch latencies showed inhibition of previously-activated competitor words. Consistent patterns for response latencies and eye-movement launch latencies on negative-priming trials indicate that the inhibitory effect in monolinguals could at least in part be localized to a pre-response processing level.
3.3. Stroop performance and correlations with linguistic competitor inhibition
To compare monolinguals' and bilinguals' accuracy and response times on the Stroop task, 2 × 2 mixed ANOVAs were conducted, with trial type (congruent, incongruent) as a within-subjects factor, and with group (monolingual, bilingual) as a between-subjects factor. As expected, participants were less accurate, F(1, 58) = 96.8, p<.001, , and slower, F(1, 58) = 438.6, p<.001, , responding to incongruent trials (accuracy: M = 84.6%, SE = 1.5%; RT: M = 464.4 ms, SE = 7.3 ms) than to congruent trials (accuracy: M = 99.0%, SE = 0.4%; RT: M = 373.3 ms, SE = 6.1 ms). A marginal effect of group was found for accuracy (but not for reaction times, p>.5), F(1,58) = 3.5, p = .07, , with bilinguals showing overall higher accuracy (M = 93.4%, SE=1.2%) than monolinguals (M = 90.2%, SE = 1.2%). Although interactions between trial type and group did not reach significance, planned comparisons revealed a marginally larger Stroop effect for accuracy in monolinguals (Stroop size = 16.8%, ) than in bilinguals (Stroop size = 12.0%, , t(58) = 1.6, p = .05 (1-tailed), suggesting a smaller Stroop effect in bilinguals and partially confirming expectations based on previous research that a bilingual advantage in nonlinguistic inhibition would emerge.
To examine the relationship between nonlinguistic and language-related inhibition, correlation analyses were conducted between Stroop performance (i.e., the Stroop effect, as measured by reaction times on incongruent trials minus reaction times on congruent trials) and linguistic competitor inhibition (as measured by reaction times to control probes minus reaction times to competitor probes on negative-priming trials). Smaller Stroop effects indicated more efficient nonlinguistic inhibition; negative priming effects approaching zero or greater than zero indicated weaker language-related inhibition. Bilinguals showed a negative correlation between Stroop inhibition and competitor priming (r = −.5, p = .004; see Fig. 5B), suggesting that more efficient Stroop inhibition was associated with less inhibition of the linguistic competitor and providing new evidence for a functional link between linguistic and nonlinguistic inhibition in bilinguals. For linguistic targets, bilinguals showed a negative correlation between nonlinguistic Stroop inhibition and target priming (r = −.4, p < .05), suggesting that more efficient Stroop inhibition was associated with increased target activation. In contrast, in monolinguals, no significant correlation was found between nonlinguistic Stroop inhibition and competitor inhibition (r = −.1, p = .591; see Fig. 5A), or between nonlinguistic Stroop inhibition and target facilitation (r = −.2, p = .329). Together, these findings confirm our predictions of a stronger relationship between nonlinguistic and linguistic inhibition in bilinguals compared to monolinguals.
Fig. 5.
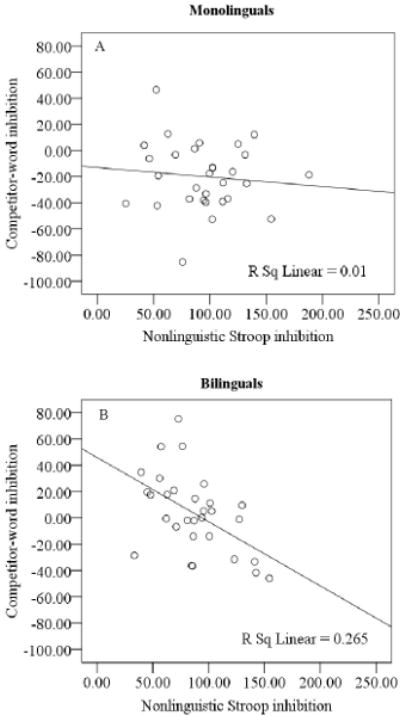
Correlations between nonlinguistic Stroop measures (incongruent–congruent RTs) and competitor word inhibition (control probecompetitor probe RTs) in monolinguals (A) and bilinguals (B).
3.4. Correlations between measures of lexical activation and inhibition
To directly relate degree of competitor activation during picture identification with measures of subsequent inhibition, correlation analyses compared competitor activation (percentage of looks to competitor minus filler pictures) to negative priming effects (response latencies during control minus competitor probe identification) and to nonlinguistic Stroop effects (response latencies during incongruent minus congruent trials).
Competitor activation during the 200–933 ms competition window was analyzed. Between 433 ms and 600 ms post-stimulus onset, monolinguals (but not bilinguals) showed a positive relationship between competitor activation and negative priming (r = .39, p < .05); in other words, the less competitor activation was present within this window the stronger the negative priming effect. Between 666.7 and 833.3 ms post-stimulus onset, bilinguals (but not monolinguals) showed a negative correlation between competitor activation and negative priming (r = −.38, p < .05); in other words, the greater the lexical activation during this time-window, the stronger the negative priming effect. The correlation between competitor activation and nonlinguistic Stroop performance was found to be significant for the entire window of competitor activation (200–933 ms post-stimulus onset, r = .39, p < .05) for bilinguals, but not monolinguals. In sum, comparisons between competitor activation during picture identification and subsequent inhibition revealed monolingual-bilingual differences, with competitor activation between 433 and 600 ms post-stimulus onset most closely associated with negative priming in monolinguals and competitor activation between 666.7 and 833.3 ms most closely associated with negative priming in bilinguals. Importantly, bilinguals also showed correlations between competitor activation and nonlinguistic Stroop performance across the duration of competitor activation, confirming that bilinguals' performance is tied to nonlinguistic inhibition.
4. Discussion
In the current study, a combined Eye-tracking/Negative Priming paradigm was employed to identify a link between auditory word comprehension and cognitive control in bilinguals. Under the hypothesis that bilingual experience acts on inhibition mechanisms used during language processing, bilingual and monolingual performance was compared. It was predicted that if bilingual experience modulated cognitive control mechanisms associated with language processing, then monolingual and bilingual groups would differ in their use of inhibition to resolve competition between similar-sounding words. Results supported this prediction. While monolinguals and bilinguals were equally efficient at identifying target words in the presence of competitors, and co-activated similar-sounding competitors to an equal extent, differences in control mechanisms were revealed in response latencies to priming probes. Importantly, results yielded a correlation between competitor word inhibition and nonlinguistic Stroop performance in bilinguals, but not monolinguals, suggesting a link between processing of linguistic ambiguity and a nonlinguistic inhibition mechanism in bilinguals.
While listening to words in their native language, English monolinguals and English-native bilinguals displayed similar activation of within-language competitor words (as reflected by eye-tracking, response accuracies and latencies), suggesting that within-language activation likely places equivalent demands on monolinguals' and bilinguals' cognitive systems. Therefore, monolingual/bilingual differences in negative priming can be interpreted as a reflection of differences in the recruitment of control mechanisms to address equivalent processing demands. Monolinguals responded slower to competitor probes than to control probes on negative-priming trials, suggesting inhibition of previously presented competitor words; bilinguals responded equally quickly to competitor and control probes. Therefore, at the time post-stimulus when inhibition of competitor words was indexed, monolinguals showed more residual inhibition than bilinguals. These patterns were reflected in launch latencies of eye-movements to priming probes, suggesting that the locus of the inhibition effect is at a pre-response processing level and confirming that location inhibition took place. Nevertheless, effects were statistically more robust when response latencies on priming probes were examined than when launch latencies of eye-movements were examined. Therefore, it is possible that response-level inhibition also played a role in the negative priming effect. Specifically, when participants experience competition between two responses, motor planning to both the target and the competitor may occur in parallel (DeSoto, Fabiani, Geary, & Gratton, 2001). Subsequently, the motor response to the competitor must be inhibited. If a motor response to the competitor picture has to be inhibited during picture identification, then subsequent re-activation of this motor response during priming probe trials may be more effortful.
The findings of competitor inhibition in monolinguals but not bilinguals are consistent with two alternative interpretations. The first interpretation is that bilinguals did not rely on inhibition to resolve competition. This, however, is an unlikely explanation since: (1) eye-tracking data suggest that bilinguals resolved competition from similar-sounding words at the same time as monolinguals, and (2) correlation findings suggest that Stroop inhibition may in fact be associated with competition resolution in bilinguals. The second interpretation is that, even though bilinguals showed no residual inhibition upon presentation of priming probes, they still relied on inhibition to resolve competition. Specifically, at the time of priming probe presentation, residual inhibition may already have dissipated in bilinguals, suggesting that bilinguals may return to a baseline activation state faster after inhibiting irrelevant information. In fact, the better bilinguals were at resolving Stroop interference, the less residual competitor inhibition they showed. The direction of this correlation suggests that, in bilinguals, more efficient nonlinguistic inhibition may be linked to faster linguistic inhibition resolution and a faster return to baseline activation.
The monolingual-bilingual differences in the current findings may point to changes in another component of cognitive control: the ability to disengage after inhibiting irrelevant information. Findings in the current study (where 500 ms had elapsed between presentation of target and priming probe trials), may reflect bilinguals' earlier return to a ‘baseline' state relative to monolinguals after successfully inhibiting the preceding phonological competitor. In other words, it is possible that bilinguals show overall less perseveration on previously-irrelevant cues than their monolingual peers. An ability to disengage more quickly from inhibiting irrelevant information may be especially important during bilingual language processing, where an irrelevant language may become relevant at any point in time. It is thus possible that a central difference between cognitive control in monolinguals and bilinguals lies in the timecourse of inhibition and the point of disengagement from inhibition.
The current findings, together with two recent negative priming studies in bilinguals (Martín, Macizo, & Bajo, 2010; Treccani, Argyri, Sorace, & Della Sala, 2009), shed light on the timecourse of inhibition in bilinguals. The present results, as well as findings from Treccani et al. (2009), point to a more efficient inhibition mechanism in bilinguals compared to monolinguals, despite of differences in findings for bilinguals across the two studies. While our findings suggest negative priming in monolinguals but not bilinguals, Treccani et al. showed stronger negative priming in bilinguals, with bilinguals making more errors on trials previously occupied by competitors during a nonlinguistic spatial negative priming task. Two possible reasons behind the differences in our findings and those by Treccani et al. may be the timing of the inhibition probe and the nature of the task. Since Treccani et al. employed a shorter time interval between presentation of target and priming probe trials (350 ms, compared to 500 ms employed in our study), stronger inhibition of irrelevant information in bilinguals suggests that bilinguals efficiently inhibit irrelevant information earlier on in the timecourse after the critical response – a finding consistent with previous work on bilingual advantages in inhibitory control. A recent study by Martín et al. (2010) shows a timecourse of inhibition in bilinguals that differs from Treccani et al.'s findings. In a cross-linguistic negative priming task, Spanish–English bilinguals made semantic relatedness judgments on English word pairs containing homographs (e.g., pie-toe, requiring a no-response in English) followed by word pairs where one item was the English translation of the previously-inhibited Spanish meaning of the homograph (e.g., foot-hand). The authors found inhibition effects when 500 ms had elapsed between the two trial types but not when 750 ms had elapsed, suggesting resolution of cross-linguistic inhibition by 750 ms after presentation of the homograph. The different timecourse of inhibition identified by Martín et al. may be due to the different locus of conflict in this task (semantic-level) as well as its cross-linguistic nature.
In general, findings of the current study, together with findings by Treccani et al. (2009) and Martín et al. (2010) suggest that bilinguals' language-related inhibitory processes may be released between 350 and 750 ms after the critical response, with variability likely to occur depending on the level of processing and locus of inhibition. It is possible that bilinguals in the current study released inhibition later than in the study by Treccani et al. (2009) but earlier than in the study by Martín et al. (2010) because of the level of processing involved. Treccani et al. presented participants with nonlinguistic stimuli while Martín et al. presented participants with semantic stimuli. The phonological-level processing required in the current study presents an intermediate level of processingwhich may be associated with inhibition of irrelevant information along an intermediate timecourse.
An additional source of information to elucidate monolingual-bilingual differences in the timecourse of inhibition resolution is to examine bilinguals with different proficiency levels. If bilingual experience is responsible for changes in the timecourse of inhibition resolution, then bilinguals with low proficiency in their second language should resemble monolinguals more than bilinguals with high proficiency in their second language. Preliminary support for this relationship comes from correlations between bilinguals' receptive vocabulary in Spanish and their linguistic inhibition performance as measured by negative priming. The more proficient bilinguals were in their non-dominant language, Spanish (as measured by the Test the Vocabulario en Imagenes Peabody, TVIP), the less residual competitor inhibition they showed (r = .4, p < .05). Therefore, it can be concluded that more proficient bilinguals were more likely to have disengaged competitor inhibition when inhibition was probed. Future research can examine whether these differences in inhibitory control across bilinguals with varying L2 proficiency levels can be tied to inhibition of cross-linguistic competitors. Such findings would confirm that cross-linguistic activation and competition are a source of changes in nonlinguistic inhibition with bilingual experience.
The finding that nonlinguistic Stroop inhibition correlates with residual competitor inhibition in bilinguals converges with findings that suggest a honed nonlinguistic Stroop effect. Bilinguals showed higher overall accuracy rates than their monolingual peers, as well as somewhat smaller Stroop effects. However, the present study yielded no robust bilingual advantages in nonlinguistic inhibitory control. Absence of such advantages may be due to the fact that participants were in their early 20s (therefore performing at ceiling) and, although they were highly proficient in both languages, were English-dominant and did not live in a fully bilingual society. Previous findings of bilingual Stroop advantages come from bilinguals who are either younger or older, and who are immersed in truly bilingual environments (e.g., Bialystok & Codd, 1997; Bia-lystok & Shapero, 2005; Costa et al., 2008; Emmorey, Borinstein, Thompson, & Gollan, 2008). Although our participants had acquired Spanish early in life (on average by 2.4 years of age) and had substantial exposure to Spanish (on average 22% of their time), recruitment limitations precluded us from finding participants who spoke an equal amount of Spanish and English on a daily basis, perhaps contributing to the lack of an absolute bilingual advantage. Despite this, current findings show a trend towards a bilingual advantage on the Stroop task and suggest that, in bilinguals, Stroop-type inhibition is closely associated with language comprehension. It is possible that, in balanced bilinguals, this link between inhibitory control and language processing may be a source of advantages in inhibitory control. Therefore, current findings point to a possible mechanism for how bilingual experience may result in cognitive changes, both within the linguistic system and in the nonlinguistic domain.
The association between linguistic processing and Stroop performance in bilinguals may stem from a similar type of inhibition required during the Stroop task and during auditory comprehension. Specifically, in both processing contexts, interference emerges due to two incongruent perceptual aspects of the same stimulus. For example, the perceptual input plu- may be automatically mapped to either plum or plug. Similarly, a left-pointing arrow on the right side of the display might be mapped to either a right-button click or a left-button click. The non-linguistic Stroop effect indexes inhibition of a competing response that is activated because an irrelevant stimulus dimension (arrow location) is perceived; similarly, the negative priming effect indexes inhibition of a word that is activated because of overlap in the perceived input. The conclusion that bilingual advantages emerge because of perceptual competition is also supported by work with bimodal bilinguals (of sign and spoken languages), who do not experience within-modality perceptual competition between languages. Emmorey, Luk, Pyers, and Bialystok (2008) found that bimodal bilinguals appear not to evince Stroop-type advantages (likely because their two languages do not compete phonologically), compared to unimodal bilinguals of two spoken languages.
In addition to the use of inhibitory control to resolve phonological competition, our results support previous findings that facilitatory mechanisms may amplify relevant information (e.g., Egner & Hirsch, 2005). To explain the relationship between inhibitory mechanisms (e.g., Green, 1998; Norman & Shallice, 1986) and facilitatory mechanisms, dual accounts that incorporate both inhibition and facilitation have been proposed to describe selective attention mechanisms (e.g., Egner & Hirsch, 2005). Findings from target and competitor activation in the present experiment provide evidence that an inhibition mechanism and an enhanced-activation/amplification mechanism may operate in tandem during monolingual and bilingual language processing. A strong relationship between response latencies on competitor priming probes (measuring residual inhibition of preceding competitors) and response latencies on target priming probes (measuring residual activation of preceding targets) was found in both bilinguals (r = .5, p = .003) and monolinguals (r = .6, p = .001). For both monolinguals and bilinguals, the less residual inhibition was indexed the more residual facilitation was found. Therefore, even though findings suggest that inhibition mechanisms may differ across monolinguals and bilinguals, the close relationship between competitor inhibition and target facilitation is consistent across groups. Interestingly, findings from eye-tracking suggest that there may be subtle differences between bilinguals and monolinguals in the timecourse of target facilitation. Bilinguals showed higher proportions of target fixation between 1099 and 2734 ms post-stimulus onset, suggesting a greater focus on the target throughout the end of the trial. It is possible that bilinguals maintain a greater focus on the target throughout the identification process in order to reduce competition from alternatives.
In conclusion, mechanisms at work during language comprehension, including parallel activation of within-language alternatives and inhibition of irrelevant information, are likely to be influenced by bilingual language experience. Bilinguals may disengage from inhibiting phonological competitors along a different timecourse than monolinguals, and this process may be closely associated with a domain-general Stroop-type inhibition mechanism. These findings have implications for models of auditory comprehension. In bilinguals, long-term demands on ambiguity resolution during comprehension may result in recruitment of a general inhibitory control mechanism. Further, via involvement in linguistic processing, aspects of cognitive control may in turn be modulated over time. Such a framework suggests that models of bilingual auditory comprehension need to be dynamic, with the nature and timecourse of inhibitory control components likely to change with linguistic experience.
Acknowledgments
We would like to thank Gerry Altmann, Ellen Bialystok, Judith Kroll, and an anonymous reviewer for valuable feedback on a previous version of the manuscript, Margarita Kaushanskaya, Guillaume Thierry, Cynthia Thompson, James Booth, Scott Schroeder, Anthony Shook, James Bartolotti, and members of the Bilingualism and Psycholinguistics Laboratory at Northwestern University for helpful discussions of this work, and Patrick Wong for assistance in recording auditory stimuli. We would also like to thank Natalie Spino, Daniela Cherbowsky, Vanessa Howes, Anna Zak, Roxanna Palma, Zahra Ali, Alyssa Greiman, Anita Goyal, and Emily Hudepohl for research assistance. This project was supported in part by a John and Lucille Clarke Scholarship and by San Diego State University Grant Program Grant #242298 to the first author, and by Grants NSF-BC-0418495 and NICHD-RO1HD059858 to the second author.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
I.e., listening contexts where bilinguals simultaneously activate their two languages either overtly (because both languages are explicitly used) or covertly (because auditory input occasionally sounds similar to words in the other language).
Phonological neighborhood size and frequency were determined using the University of Washington Speech & Hearing Lab Neighborhood Database (the Phonological Neighborhood database can be accessed at the following website: http://128.252.27.56/Neighborhood/Home.asp), and WordGen (Duyck, Desmet, Verbeke, & Brysbaert, 2004).
One bilingual participant was omitted from analyses of eye-movement launch latencies on negative-priming trials because they looked at grey asterisks on only 6 out of 78 trials, 4 of which were trials where target pictures had been identified incorrectly or with response latencies that exceeded three standard deviations.
References
- Baayen H, Piepenbrock R, van Rijn H. The CELEX lexical database (CD-ROM) Philadelphia, PA: University of Pennsylvania; Linguistic Data Consortium: 1995. [Google Scholar]
- Bialystok E. Cognitive complexity and attentional control in the bilingual mind. Child Development. 1999;70:636–644. [Google Scholar]
- Bialystok E. Consequences of bilingualism for cognitive development. In: Kroll JF, de Groot AMB, editors. Handbook of Bilingualism: Psycholinguistic Approaches. Oxford, UK: Oxford University Press; 2005. [Google Scholar]
- Bialystok E. Effect of bilingualism and computer video game experience on the Simon task. Canadian Journal of Experimental Psychology. 2006;60(1):68–79. doi: 10.1037/cjep2006008. [DOI] [PubMed] [Google Scholar]
- Bialystok E. Global-local and trail-making tasks by monolingual and bilingual children: Beyond inhibition. Developmental Psychology. 2010;46:93–105. doi: 10.1037/a0015466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bialystok E, Codd J. Cardinal limits evidence from language awareness and bilingualism for developing concepts of number. Cognitive Development. 1997;12:85–106. [Google Scholar]
- Bialystok E, Craik FIM, Klein R, Viswanathan M. Bilingualism, aging, and cognitive control: Evidence from the Simon task. Psychology and Aging. 2004;19:290–303. doi: 10.1037/0882-7974.19.2.290. [DOI] [PubMed] [Google Scholar]
- Bialystok E, Martin M. Attention and inhibition in bilingual children: Evidence from the dimensional change card sort task. Developmental Science. 2004;7:325–339. doi: 10.1111/j.1467-7687.2004.00351.x. [DOI] [PubMed] [Google Scholar]
- Bialystok E, Shapero D. Ambiguous benefits: The effect of billingualism on reversing ambiguous figures. Developmental Science. 2005;8:595–604. doi: 10.1111/j.1467-7687.2005.00451.x. [DOI] [PubMed] [Google Scholar]
- Bialystok E, Viswanathan M. Components of executive control with advantages for bilingual children in two cultures. Cognition. 2009;112:494–500. doi: 10.1016/j.cognition.2009.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blumenfeld HK. Suppression mechanisms in monolingual and bilingual language comprehension: Evidence from eye-tracking, priming, and executive tasks Ph D Dissertation. Northwestern University; Ann Arbor: UMI Dissertation Services: 2008. [Google Scholar]
- Blumenfeld HK, Marian V. Constraints on parallel activation in bilingual spoken language processing: Examining proficiency and lexical status using eye-tracking. Language and Cognitive Processes. 2007;22(5):633–660. [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD. Conflict monitoring and cognitive control. Psychological Review. 2001;108(3):624–652. doi: 10.1037/0033-295x.108.3.624. [DOI] [PubMed] [Google Scholar]
- Bradlow AR, Pisoni DB. Recognition of spoken words by native and non-native listeners: Talker-, listener-, and item-related factors. Journal of the Acoustic Society of America. 1999;106:2074–2085. doi: 10.1121/1.427952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canseco-Gonzales E, Brick C, Fischer K, Wagner K. “Carpet or Carcel” effects of speaker type, fluency, and language mode on bilingual lexical access. Proceedings of the International Symposium on Bilingualism, Spain. 2005;5:156–157. [Google Scholar]
- Christoffels IK, De Groot AMB, Kroll JF. Memory and language skills in simultaneous interpreters: The role of expertise and language proficiency. Journal of Memory and Language. 2006;54:324–345. [Google Scholar]
- Colzato LS, Bajo MT, van den Wildenberg W, Paolieri D. How does bilingualism improve executive control? A comparison of active and reactive inhibition mechanisms. Journal of Experimental Psychology: Learning, Memory and Cognition. 2008;34(2):302–312. doi: 10.1037/0278-7393.34.2.302. [DOI] [PubMed] [Google Scholar]
- Connelly SL, Hasher L. Aging and inhibition of spatial location. Journal of Experimental Psychology: Human Perception and Performance. 1993;19:1238–1250. doi: 10.1037//0096-1523.19.6.1238. [DOI] [PubMed] [Google Scholar]
- Cook VJ. The consequences of bilingualism for cognitive processing. In: de Groot A, Kroll JF, editors. Tutorials in Bilingualism. Lawrence Erlbaum; 1997. pp. 279–300. [Google Scholar]
- Costa A, Hernandez M, Costa J, Sebastian-Galles N. On the bilingual advantage in conflict processing: Now you see it now you don't. Cognition. 2009;113:135–149. doi: 10.1016/j.cognition.2009.08.001. [DOI] [PubMed] [Google Scholar]
- Costa A, Hernandez M, Sebastian-Galles N. Bilingualism aids conflict resolution: Evidence from the ANT task. Cognition. 2008;106:59–86. doi: 10.1016/j.cognition.2006.12.013. [DOI] [PubMed] [Google Scholar]
- Costa A, Santesteban M. Lexical access in bilingual speech production: Evidence from language switching in highly proficient bilinguals and L2 learners. Journal of Memory and Language. 2004;50(4):491–511. [Google Scholar]
- Cutler A, Weber A, Otake T. Asymmetric mapping from phonetic to lexical representations in second-language listening. Journal of Phonetics. 2006;34:269–284. [Google Scholar]
- Dahan D, Magnusson JS, Tanenhaus MK. Time course of frequency effects in spoken-word recognition: Evidence from eye movements. Cognitive Psychology. 2001;42:317–367. doi: 10.1006/cogp.2001.0750. [DOI] [PubMed] [Google Scholar]
- Degani T, Tokowicz N. Semantic ambiguity within and across languages: An integrative review. Journal of Experimental Psychology. 2010;63(7):1266–1303. doi: 10.1080/17470210903377372. [DOI] [PubMed] [Google Scholar]
- DeSoto CM, Fabiani M, Geary DC, Gratton G. When in doubt, do it both ways: Brain evidence of the simultaneous activation of conflicting motor responses in a spatial Stroop task. Journal of Cognitive Neuroscience. 2001;13(4):523–536. doi: 10.1162/08989290152001934. [DOI] [PubMed] [Google Scholar]
- Desroches AS, Newman RL, Joanisse M. Investigating the time course of spoken word recognition: Elecrophysiological evidence for the influence of phonological similarity. Journal of Cognitive Neuroscience. 2008;21(10):1893–1906. doi: 10.1162/jocn.2008.21142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dijkstra T, van Heuven WJB. The BIA-model and bilingual word recognition. In: Grainger J, Jacobs A, editors. Localist connectionist approaches to human cognition. Hove: LEA; 1998. [Google Scholar]
- Dunn LM, Dunn LM. Peabody picture vocabulary test (PPVT) Circle Pines, MN: American Guidance Service; 1997. [Google Scholar]
- Dunn LM, Padilla ER, Lugo DE, Dunn LM. Test de Vocabulario en Imágenes Peabody (TVIP) Circle Pines, MN: American Guidance Service; Publishing: 1986. [Google Scholar]
- Duyck W, Desmet T, Verbeke L, Brysbaert M. WordGen: A tool for word selection and non-word generation in Dutch, German, English, and French. Behavior Research Methods, Instruments & Computers. 2004;36(3):488–499. doi: 10.3758/bf03195595. [DOI] [PubMed] [Google Scholar]
- Egner T, Hirsch J. Cognitive control mechanisms resolve conflict through cortical amplification of task-relevant information. Nature Neuroscience. 2005;12:1784–1790. doi: 10.1038/nn1594. [DOI] [PubMed] [Google Scholar]
- Emmorey K, Borinstein HB, Thompson R, Gollan TH. Bimodal bilingualism. Bilingualism: Language and Cognition. 2008;11(1):1–19. doi: 10.1017/S1366728907003203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Emmorey K, Luk G, Pyers JE, Bialystok E. The source of enhanced cognitive control in bilinguals. Psychological Science. 2008;19(12):1201–1206. doi: 10.1111/j.1467-9280.2008.02224.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gernsbacher MA. Language comprehension as structure building. Hillsdale, NJ: Erlbaum; 1990. [Google Scholar]
- Gernsbacher MA, Faust M. The mechanism of suppression: A component of general comprehension skill. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1991;11:1. doi: 10.1037//0278-7393.17.2.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green DW. Mental control of the bilingual lexico-semantic system. Bilingualism: Language and Cognition. 1998;1:67–81. [Google Scholar]
- Hallett PE. In: Handbook of Perception and Human Performance. Boff K, Kaufman L, Thomas J, editors. Eye-movements; Wiley: New York: 1986. pp. 10–1.pp. 10–112. [Google Scholar]
- Henik A, Bibi U, Yanai M, Tzelgov J. The Stroop effect is largest during first trials. Abstracts of the Psychonomic Society. 1997;2:57. [Google Scholar]
- Hernandez AE, Meschyan G. Executive function is necessary to enhance lexical processing in a less proficient L2: Evidence from fMRI during picture naming. Bilingualism: Language and Cognition. 2006;9(2):177–188. [Google Scholar]
- Huettig F, Altmann GTM. Word meaning and the control of eye-fixation: Semantic competitor effects and the visual world paradigm. Cognition. 2005;96:B23–B32. doi: 10.1016/j.cognition.2004.10.003. [DOI] [PubMed] [Google Scholar]
- Ju M, Luce PA. Falling on sensitive ears. Psychological Science. 2004;15:314–318. doi: 10.1111/j.0956-7976.2004.00675.x. [DOI] [PubMed] [Google Scholar]
- Kane MJ, Engle RW. Working-memory capacity and the control of attention: The contributions of goal neglect, response competition, and task set to Stroop interference. Journal of Experimental Psychology: General. 2003;132(1):47–70. doi: 10.1037/0096-3445.132.1.47. [DOI] [PubMed] [Google Scholar]
- Kemper S, Sumner A. The structure of verbal abilities in young and older adults. Psychology and Aging. 2001;16(2):312–322. [PubMed] [Google Scholar]
- Kerns JG. Experimental manipulation of cognitive control processes causes an increase in communication disturbances in healthy volunteers. Psychological Medicine. 2007;37(7):995–1004. doi: 10.1017/S0033291706009718. [DOI] [PubMed] [Google Scholar]
- Kornblum S. Dimensional overlap and dimensional relevance in stimulus-response and stimulus–stimulus compatibility. In: Stelmach G, Requin J, editors. Tutorials in motor behavior II. Amsterdam: North-Holland; 1992. pp. 743–777. [Google Scholar]
- Kornblum S. The way irrelevant dimensions are processed depends on what they overlap with: The case of Stroop- and Simon-like stimuli. Psychological Research. 1994;56:130–135. doi: 10.1007/BF00419699. [DOI] [PubMed] [Google Scholar]
- Kovacs AM, Mehler J. Cognitive gains in 7-month-old bilingual infants. Proceedings of the National Academy of Sciences. 2009;106(16):6556–6560. doi: 10.1073/pnas.0811323106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kroll JF. Juggling two languages in one mind. Psychological Science Agenda, American Psychological Association, 22 2008 [Google Scholar]
- Kwong See ST, Ryan EB. Cognitive mediation of adult age differences in language performance. Psychology and Aging. 1995;10:458–468. doi: 10.1037//0882-7974.10.3.458. [DOI] [PubMed] [Google Scholar]
- Linck JA, Hoshino N, Kroll JF. Cross-language lexical processes and inhibitory control. Mental Lexicon. 2008;3(3):349–374. doi: 10.1075/ml.3.3.06lin. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Linck JA, Kroll JF, Sunderman G. Losing access to the native language while immersed in a second language: Evidence for the role of inhibition in second-language learning. Psychological Science. 2009;20(12):1507–1515. doi: 10.1111/j.1467-9280.2009.02480.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X, Banich MT, Jacobson BL, Tanabe JL. Common and distinct neural substrates of attentional control in an integrated Simon and spatial Stroop task as assessed by event-related fMRI. NeuroImage. 2004;22(3):1097–1106. doi: 10.1016/j.neuroimage.2004.02.033. [DOI] [PubMed] [Google Scholar]
- Luce PA, Pisoni DB. Recognizing spoken words: The neighborhood activation model. Ear and Hearing. 1998;19:1–36. doi: 10.1097/00003446-199802000-00001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marian V, Blumenfeld HK, Kaushanskaya M. The language experience and proficiency questionnaire (LEAP-Q): Assessing language profiles in bilinguals and multilinguals. Journal of Speech Language and Hearing Research. 2007;50(4):940–967. doi: 10.1044/1092-4388(2007/067). [DOI] [PubMed] [Google Scholar]
- Marian V, Spivey M. Bilingual and monolingual processing of competing lexical items. Applied Psycholinguistics. 2003a;24:173–193. [Google Scholar]
- Marian V, Spivey M. Competing activation in bilingual language processing: Within- and between-language competition. Bilingualism: Language and Cognition. 2003b;6:97–115. [Google Scholar]
- Marslen-Wilson WD. Functional parallelism in spoken word recognition. Cognition. 1987;25:71–102. doi: 10.1016/0010-0277(87)90005-9. [DOI] [PubMed] [Google Scholar]
- Martín MC, Macizo P, Bajo T. Time course of inhibitory processes in bilingual language processing. British Journal of Psychology. 2010;101(4):679–693. doi: 10.1348/000712609X480571. [DOI] [PubMed] [Google Scholar]
- May CP, Kane MJ, Hasher L. Determinants of negative priming. Psychological Bulletin. 1995;118(1):35–54. doi: 10.1037/0033-2909.118.1.35. [DOI] [PubMed] [Google Scholar]
- McClelland JL, Elman JL. The TRACE model of speech perception. Cognitive Psychology. 1986;18(1):1–86. doi: 10.1016/0010-0285(86)90015-0. [DOI] [PubMed] [Google Scholar]
- Milliken B, Lupianez J, Debner J, Abello B. Automatic and controlled processing in Stroop negative priming: The role of attentional set. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1999;25:1384–1402. [Google Scholar]
- Nakamichi K. Young children's conditional reasoning, working memory, and inhibitory control. Japanese Journal of Educational Psychology. 2007;55(3):347–358. [Google Scholar]
- Norman D, Shallice T. Attention to action: Willed and automatic control of behavior. In: Davidson R, Schwartz G, Shapiro D, editors. Consciousness and self regulation: Advances in research and theory. Vol. 4. New York, NY: Plenum; 1986. pp. 1–18. [Google Scholar]
- Prior A, MacWhinney B. A bilingual advantage in task switching. Bilingualism: Language and Cognition. 2010;13:253–362. doi: 10.1017/S1366728909990526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PsychCorp. Wechsler abbreviated scale of intelligence (WASI) San Antonio, TX: Harcourt Assessment, Inc; 1999. [Google Scholar]
- Rodriguez-Fornells A, Balaguer RD, Munte TF. Executive control in bilingual language processing. Language Learning. 2006;56:133–190. [Google Scholar]
- Seidenberg MS, Tanenhaus MK, Leiman JM, Bienkowski M. Automatic access of the meaning of ambiguous words in context: Some limitations of knowledge-based processing. Cognitive Psychology. 1982;14:489–537. [Google Scholar]
- Shook A, Marian V. Bimodal bilinguals reveal the role of top-down mechanisms in the language system [Google Scholar]
- Simone PM, Ahrens K, Foerde KEG, Spinetta M. Influence of attended repetition trials on negative priming in younger and older adults. Memory and Cognition. 2006;34(1):187–195. doi: 10.3758/bf03193397. [DOI] [PubMed] [Google Scholar]
- Smith LB, Jones SS, Landau B, Gershkoff-Stowe L, Samuelson L. Object name learning provides on-the-job training for attention. Psychological Science. 2002;13:13–19. doi: 10.1111/1467-9280.00403. [DOI] [PubMed] [Google Scholar]
- Stroop JR. Studies of interference in serial verbal reactions. Journal of Experimental Psychology. 1935;18:643–662. [Google Scholar]
- Swinney DA. Lexical access during sentence comprehension: (Re) onsideration of context effects. Journal of Verbal Learning and Verbal Behavior. 1979;18:645–659. [Google Scholar]
- Székely A, Jacobsen T, D'Amico S, Devescovi A, Andonova E, Herron D, et al. A new on-line resource for psycholinguistic studies. Journal of Memory and Language. 2004;51(2):247–250. doi: 10.1016/j.jml.2004.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanenhaus M, Spivey-Knowlton M, Eberhard K, Sedivy J. Integration of visual and linguistic information in spoken language comprehension. Science. 1995;268:1632–1634. doi: 10.1126/science.7777863. [DOI] [PubMed] [Google Scholar]
- Taylor JL, O'Hara R, Mumenthaler MS, Rosen AC, Yesavage JA. Cognitive ability, expertise, and age differences in following air-traffic control instructions. Psychology and Aging. 2005;20(1):117–133. doi: 10.1037/0882-7974.20.1.117. [DOI] [PubMed] [Google Scholar]
- Thompson-Schill SL, D'Esposito M, Aguirre GK, Farah MJ. Role of left inferior prefrontal cortex in retrieval of semantic knowledge: A reevaluation. Proceedings of the National Academy of Sciences of the United States of America. 1997;94:14792–14797. doi: 10.1073/pnas.94.26.14792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson-Schill SL, D'Esposito M, Kan IP. Effects of repetition and competition on activity in left pre-frontal cortex during word generation. Neuron. 1999;23:513–522. doi: 10.1016/s0896-6273(00)80804-1. [DOI] [PubMed] [Google Scholar]
- Tipper SP. The negative priming effect: Inhibitory priming by ignored objects. Quarterly Journal of Experimental Psychology. 1985;37A:571–590. doi: 10.1080/14640748508400920. [DOI] [PubMed] [Google Scholar]
- Treccani B, Argyri E, Sorace A, Della Sala S. Spatial negative priming in bilingualism. Psychonomic Bulletin and Review. 2009;16(2):320–327. doi: 10.3758/PBR.16.2.320. [DOI] [PubMed] [Google Scholar]
- Wagner RK, Torgesen JK, Rashotte CA. The comprehensive test of phonological processing. Austin, TX: Pro-Ed; 1999. [Google Scholar]
- Weber A, Cutler A. Lexical competition in non-native spoken-word recognition. Journal of Memory and Language. 2004;50:1–25. [Google Scholar]
- Weber A, Paris G. The origin of the linguistic gender effect in spoken-word recognition: Evidence from non-native listening. Proceedings of the Twenty-Sixth Annual Meeting of the Cognitive Science Society. 2004:1446–1451. [Google Scholar]
- Yee E, Sedivy JC. Eye movements to pictures reveal transient semantic activation during spoken word recognition. Journal of Experimental Psychology: Learning, Memory and Cognition. 2006;32(1):1–14. doi: 10.1037/0278-7393.32.1.1. [DOI] [PubMed] [Google Scholar]