

Abstract
Sequential photo-bleaching provides a non-invasive way to label individual SCs at the NMJ. The NMJ is the largest synapse of the mammalian nervous system and has served as guiding model to study synaptic structure and function. In mouse NMJs motor axon terminals form pretzel-like contact sites with muscle fibers. The motor axon and its terminal are sheathed by SCs. Over the past decades, several transgenic mice have been generated to visualize motor neurons and SCs, for example Thy1-XFP1 and Plp-GFP mice2, respectively.
Along motor axons, myelinating axonal SCs are arranged in non-overlapping internodes, separated by nodes of Ranvier, to enable saltatory action potential propagation. In contrast, terminal SCs at the synapse are specialized glial cells, which monitor and promote neurotransmission, digest debris and guide regenerating axons. NMJs are tightly covered by up to half a dozen non-myelinating terminal SCs - these, however, cannot be individually resolved by light microscopy, as they are in direct membrane contact3.
Several approaches exist to individually visualize terminal SCs. None of these are flawless, though. For instance, dye filling, where single cells are impaled with a dye-filled microelectrode, requires destroying a labelled cell before filling a second one. This is not compatible with subsequent time-lapse recordings3. Multi-spectral "Brainbow" labeling of SCs has been achieved by using combinatorial expression of fluorescent proteins4. However, this technique requires combining several transgenes and is limited by the expression pattern of the promoters used. In the future, expression of "photo-switchable" proteins in SCs might be yet another alternative5. Here we present sequential photo-bleaching, where single cells are bleached, and their image obtained by subtraction. We believe that this approach - due to its ease and versatility - represents a lasting addition to the neuroscientist's technology palette, especially as it can be used in vivo and transferred to others cell types, anatomical sites or species6.
In the following protocol, we detail the application of sequential bleaching and subsequent confocal time-lapse microscopy to terminal SCs in triangularis sterni muscle explants. This thin, superficial and easily dissected nerve-muscle preparation7,8 has proven useful for studies of NMJ development, physiology and pathology9. Finally, we explain how the triangularis sterni muscle is prepared after fixation to perform correlated high-resolution confocal imaging, immunohistochemistry or ultrastructural examinations.
Keywords: Neuroscience, Issue 71, Cellular Biology, Molecular Biology, Neurobiology, Immunology, Medicine, Anatomy, Physiology, Surgery, Triangularis sterni, explant, Schwann cells, imaging, neuromuscular junction, immunohistochemistry, bleaching, muscle, nerve, mouse, animal model
Protocol
1. Triangularis Sterni Explant (Figure 1)
Timing: 15 min.
Prepare surgical instruments: 2 pairs of forceps, 1 pair of scissors, 1 pair of spring scissors, 1 tissue culture dish (10-cm). Pre-bubbled (minimum for 15 min) cooled (4 °C) Ringer's solution with 5% CO2/ 95% O2. Fill 15-cm tissue culture dish with ice and cover it with metal plate. Prepare dissection microscope and place the 15-cm dish with ice under the dissection scope in order to cool the explant during dissection.
Lethally anesthetize the mouse by isoflurane overdose (or any other approved method of sacrifice). Spray the mouse with 70% ethanol to prevent fur contamination of the explant. For SC bleaching, we used Plp-GFP mice, which can be crossed to Thy1-OFP3 or Thy1-Membow13 for simultaneous visualization of the axon.
Using the pair of large scissors, make a midline incision of the skin over the sternum and two incisions parallel to the lower edge of the rib cage.
Using the pair of large scissors, remove the skin, open the abdominal wall, and make incisions parallel to the rib cage all the way back to the vertebral column. Then dissect off the pectoral muscles by making incisions close to the muscle insertion at the sternum.
Cut the diaphragm open just below the xiphoid cartilage and dissect off the diaphragm along its costal insertions.
Holding the rib cage by the xiphoid process with a set of forceps, start cutting the ribs off the vertebral column, as close as possible to their insertions. The left and right cuts should converge above the heart towards the manubrium sterni. Try to avoid cutting major blood vessels (especially the subclavian veins). Better visibility is achieved by gently lifting the preparation while holding onto the xiphoid process with forceps.
Make a transverse cut just below the manubrium sterni and transfer the explant into the 10-cm dish with cooled 5% CO2/ 95% O2-bubbled Ringer's solution. Place the 10-cm dish on the metal plate covering the 15-cm tissue culture dish filled with ice. All further steps should be done under the dissection microscope.
Using small spring scissors remove the remnants of thymus, pleura, diaphragm (inside) and pectoral muscles (outside; Figure 1B-D).
To fit the explant to the 3.5-cm dish, dissect off all the ribs that do not insert to the sternum. This is best done using small spring scissors and cutting the tissue in between ribs (Figure 1E).
Pin the triangularis sterni nerve-muscle explant into the Sylgard coated 3.5-cm tissue culture dish using minutien pins (shortened to < 4 mm; Figure 1G). Two pins should go through cartilaginous (white) parts of the sternum. Insert at least two pins through the ribs both on the left and right side of the explant aiming for the softer cartilaginous parts and avoiding the ribs under or close to the triangularis muscle. The more pins are used, the better (we typically use 8-10). As you pin down the preparation, make sure that the ribs are somewhat spread, fixing the muscle into a slightly stretched position.
Optional: Visualize synaptic sites containing acetylcholine receptors by adding bungarotoxin coupled to Alexa dyes (concentration of 0.8 μg/ml in 5% CO2/ 95% O2-bubbled Ringer's solution). Incubate pinned explant in Sylgard coated dish for 15-20 min at room temperature then wash several times with 5% CO2/ 95% O2-bubbled Ringer's solution.
2. Bleaching SCs and Optional Time-lapse Microscopy (Figure 2)
Timing: 30 - 45 min + optional 1 - 5 hr for time lapse.
Transfer 3.5-cm Sylgard-coated dish to a confocal microscope equipped with an argon laser (488 nm wavelength) and water immersion objectives (4x, 0.13 NA; 20x, 0.5 NA and 100x, 1.0 NA or 60x, 0.9 NA).
Optional for time-lapse microscopy: Insert 3.5-cm Sylgard-coated dish into heating ring, install superfusion system and temperature probe (Figure 2A). Make sure that none of them touch the explant, the rim of the dish or the Sylgard coating. Heat the sample to 33-35 °C. For SC bleaching without time-lapse, room temperature is better, as cells show less dynamics between bleaching steps.
With a 4x air objective observe innervation pattern of triangularis sterni muscle and find triangularis sterni endplate band. Switch to 20x dipping-cone objective and start looking for superficial regions within the endplate band (areas overlying ribs are good candidates). Change objective to 100x or 60x dipping-cone objective to check on individual NMJs. Ideal is an NMJ that is covered by several SCs and is located very superficially (Figure 2B). If you perform time-lapse imaging after bleaching, select up to 3 NMJs that are relatively close together to increase yield.
Acquire a confocal image of the NMJ before photo-bleaching individual SCs (Figure 2B). (For full resolution imaging, e.g. on an Olympus FV1000 microscope, use a 100x, 1.0 NA objective and 2.5 zoom, which results in Nyquist-limited sampling of ~0.1 μm per pixel.)
"Park" the 488 nm laser beam centrally on the nucleus of a SC using maximal power for 5 seconds (alternatively an efficient scan pattern in a small region of interest can be used, such as the "tornado scan" function on an Olympus microscope; as we use a confocal microscope, the laser is focussed into the image-conjugate planes and hence, at 488 nm with an objective of 1.0 N.A., < 0.5 μm in diameter). Re-focus on the cell and judge bleaching result. If necessary repeat bleaching step again until GFP levels are reduced to near-background levels. Acquire an image after bleaching (Figure 2C). It is critical to make sure that the bleached region is outside of any overlap between two cells - if there is overlap, bleaching in a second cell will be obvious.
Repeat step above until all but one SC are bleached (Figure 2D-E). The last "unbleached" SC is suitable for confocal time-lapse microscopy. Reconstruct single cells by subtraction of images, e.g. using Photoshop software, to delineate single SC morphology and territory (Figure 2F). Be aware that subtracting two mages adds noise (e.g. by importing them in consecutive "layers" in Photoshop and then setting the top channel to "difference" in the drop down menu at the top of the channels tab). This can be circumvented by using the "despeckle" function on the channels before subtraction and cropping away any background that obviously does not originate from the bleached cell. To optimize contrast, it can be necessary to adjust the brightness of the two channels in the "levels" window. The resulting image of the bleached cell is the superimposed on the original image of a synapse, pseudo-colored in a unique hue and made transparent to color the territory of the bleached cell in the original image (from doing this for all bleached and the last, unbleached cells the image shown in Figure 2F results).
Optional: Start confocal time-lapse microscopy after bleaching all but one SCs. Take an image at fixed intervals (for example every 5 to 10 min). Use as little laser power as possible. Up to 3 NMJs can be imaged at the same session.
Make a map of the time-lapsed NMJs by taking an image at 100x without zoom, then images of the region using 20x and 4x objectives (Figure 2G-I). This "map" is needed to find NMJs following immunohistochemistry of a fixed muscle.
For image processing, convert individual images into maximum intensity projections and save in tiff format. Combine all z-projected time-points into a stack and align in xy, e.g. using the "stackreg" plugin10 in Fiji (a package based on the open source software ImageJ; National Institutes of Health).
3. Fixation and Immunohistochemistry (Figure 3)
Timing: Overnight.
Transfer the explant into a 50-ml reaction tube containing at least 15 ml 4% PFA by carefully removing the pins. Post-fix the anterior thoracic wall containing the triangularis sterni muscle for 1 hr on ice. Rinse fixed tissue with 0.1 M glycine in PBS (to quench residual fixative and hence reduce background) for at least 10 min. Samples can be stored at this point and processed later.
Transfer fixed explant to a 10-cm Sylgard coated tissue culture dish filled with 0.01 M PBS. Cut out a trapezoid shape with one side parallel to the sternum and the other through the bone-to-cartilage transitions - this contains the triangularis muscle on its surface. Remove lower ribs with a horizontal (i.e. trans-sternal) cut so that the caudal end of the triangularis muscle can be freely accessed (Figure 3A-D).
Insert a hypodermic needle as a pin into the upper part of the trapezoid (Figure 3E). Use a hypodermic needle attached to a 1 ml syringe and forceps to remove the triangularis muscle on the surface by gently holding onto a caudal corner of the muscle and using the needle as a micro-scalpel. Make sure that cuts are made parallel to the plane of the thoracic wall. Once the caudal part of the muscle is released, insert another hypodermic needle as a pin below the muscle through the thoracic wall - this immobilizes the preparation and allows exerting a gentle pull on the muscle using forceps (Figure 3F). As you are cutting connective tissue insertions, "peel" away the muscle from the thorax. Cut last caudal attachment with spring scissors. Remove fat and blood vessel remnants using forceps. Be careful not to rip away the nerves. Note: Use separate sets of tools and dishes for working with live and fixed tissue.
Start immunohistochemistry (using a standard protocol) by transferring tissue samples into blocking solution for 1 hr on a shaker at room temperature. Smallest possible volume for staining is 200 μl using a 96-well plate.
Apply primary antibody diluted in blocking solution, 200 μl per well, overnight at 4 °C on a shaker.
Wash in 0.01 M PBS three times for 10 min.
Apply suitable secondary antibody for 1-2 hr at room temperature diluted in blocking solution.
Wash in 0.01 M PBS three times for 10 min.
Mount in anti-fading medium (e.g. Vectashield) and coverslip.
Place slide on magnetic metal plate, place magnet on top of the sample to flatten for a few hours or overnight at 4 °C. Seal with nail polish.
Representative Results
An example of a triangularis sterni explant ready for imaging dish is shown in Figure 1G. This explant is particularly suitable for imaging NMJs ex vivo as the triangularis muscle consists only of a few layers of muscle fibers. This allows obtaining high resolution images from explants derived from transgenic mouse lines that highlight either SCs (Plp-GFP2) and/or axons (Thy1-XFP1). Critical factors for high quality imaging include: (i) avoiding touching the triangularis muscle during explant preparation to prevent muscle fiber injury; (ii) selecting superficial and flat synapses to reduce depth-associated aberrations, (iii) not overheating the sample (> 37 °C) and keeping the sample well oxygenated, (iv) potentially stopping the superfusion during imaging to reduce drift or vibration; (v) carefully placing superfusion tubing and temperature probes to avoid touching the sample, as well as keeping the underside of the dish dry to prevent drift; (vi) balancing comprehensive bleaching with avoiding "collateral" signal loss in off-target SCs to maximize contrast. Muscle twitching and axon fragmentation are frequently observed phenomena, if the explant is damaged and unhealthy. Instability of the preparation results in blurry projections of confocal stacks. However, once the described sequential photo-bleaching technique works well, it reveals the morphology of single terminal SC in considerable detail (Figure 2F). The revealed morphology of single terminal SCs can be quite surprising and is not easily predicted before bleaching. Moreover, the degree of SC motility within the NMJ cannot be revealed without labeling single cells.
Following live microscopy, the triangularis sterni explant can be fixed and stained for any marker of interest (Figure 3). For this, live imaged cells can be tracked back using the map created beforehand (Figure 2G-I) and scanned at high resolution in the fixed tissue. Generally standard methods for immunohistochemistry can be used - one caveat here is the fact that antibody penetration can be quite limited in myelinated axons and requires very long antibody incubation times. Again, choosing superficial NMJs and removing fat residues from the fixed muscle well (without ripping off the nerves) helps to avoid this pitfall.
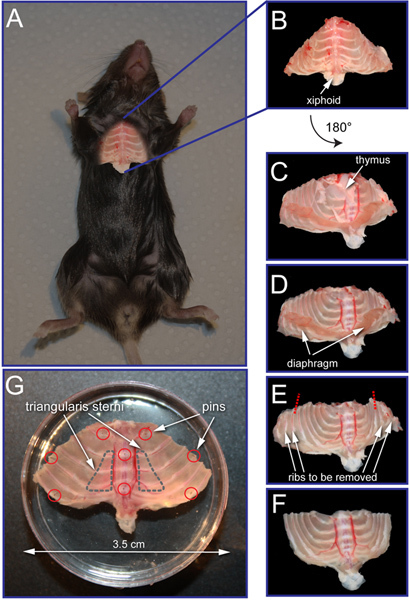 Figure 1. Triangularis sterni explant preparation. (A) To prepare the triangularis nerve-muscle explant it is necessary to dissect the complete anterior thoracic wall of the mouse (schematically superimposed on a mouse). (B) Anterior thoracic wall just after dissection. (C) Inner side of the anterior thoracic wall. Note the remnants of the thymus and diaphragm that should be removed. (D, E) Anterior thoracic wall after removal of thymus (D) and diaphragm (E). Red dashed lines indicate the entry points for incisions to remove all the ribs that do not insert to the sternum. (F) Explant ready to be pinned down into the 3.5-cm Sylgard coated dish. (G) Triangularis sterni explant pinned down and ready for imaging. Note the position of the pins (red circles) and triangularis sterni muscle localization (outlined with gray dashed line). Click here to view larger figure.
Figure 1. Triangularis sterni explant preparation. (A) To prepare the triangularis nerve-muscle explant it is necessary to dissect the complete anterior thoracic wall of the mouse (schematically superimposed on a mouse). (B) Anterior thoracic wall just after dissection. (C) Inner side of the anterior thoracic wall. Note the remnants of the thymus and diaphragm that should be removed. (D, E) Anterior thoracic wall after removal of thymus (D) and diaphragm (E). Red dashed lines indicate the entry points for incisions to remove all the ribs that do not insert to the sternum. (F) Explant ready to be pinned down into the 3.5-cm Sylgard coated dish. (G) Triangularis sterni explant pinned down and ready for imaging. Note the position of the pins (red circles) and triangularis sterni muscle localization (outlined with gray dashed line). Click here to view larger figure.
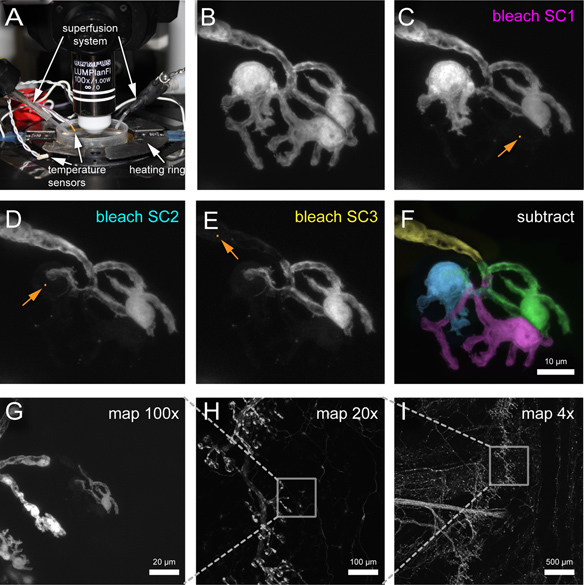 Figure 2. Sequential SC bleaching in a Plp-GFP mouse. (A) Confocal setup equipped for time-lapse microscopy with superfusion system. (B) NMJ with 3 terminal SCs which are sequentially bleached in (C-E). (F) Image subtraction using Photoshop software and pseudo-colored overlay allows visualizing single SC territories. (G-I) Map of imaged SCs to track back imaged SC after immunohistochemistry. (C-E) The orange dots highlighted by orange arrows indicate the approximate position and size of the region of interested used for bleaching (here a "tornado" scan pattern was used). Click here to view larger figure.
Figure 2. Sequential SC bleaching in a Plp-GFP mouse. (A) Confocal setup equipped for time-lapse microscopy with superfusion system. (B) NMJ with 3 terminal SCs which are sequentially bleached in (C-E). (F) Image subtraction using Photoshop software and pseudo-colored overlay allows visualizing single SC territories. (G-I) Map of imaged SCs to track back imaged SC after immunohistochemistry. (C-E) The orange dots highlighted by orange arrows indicate the approximate position and size of the region of interested used for bleaching (here a "tornado" scan pattern was used). Click here to view larger figure.
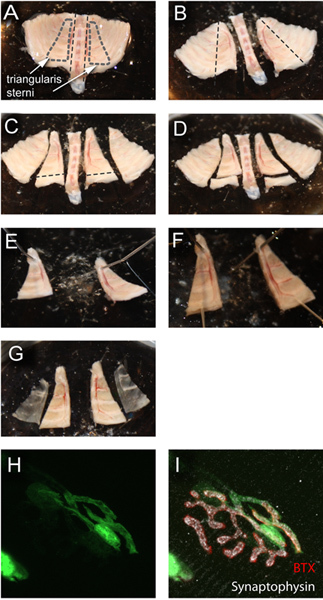 Figure 3. Dissection of the fixed triangularis sterni muscle and image of fixed NMJ. (A) Fixed triangularis sterni explant in PBS in cell culture dish coated with Sylgard. (B-D) Cuts performed with spring scissors to isolate triangularis sterni muscle. (E-G) Pinning of explant piece containing triangularis sterni muscle and removing muscle from surface. (H, I) Fixed view of the very NMJ that was imaged in Figure 2. Staining for acetylcholine receptors with bungarotoxin coupled to Alexa 647 (red) and for synaptic vesicles with an anti-synaptophysin antibody (white). Click here to view larger figure.
Figure 3. Dissection of the fixed triangularis sterni muscle and image of fixed NMJ. (A) Fixed triangularis sterni explant in PBS in cell culture dish coated with Sylgard. (B-D) Cuts performed with spring scissors to isolate triangularis sterni muscle. (E-G) Pinning of explant piece containing triangularis sterni muscle and removing muscle from surface. (H, I) Fixed view of the very NMJ that was imaged in Figure 2. Staining for acetylcholine receptors with bungarotoxin coupled to Alexa 647 (red) and for synaptic vesicles with an anti-synaptophysin antibody (white). Click here to view larger figure.
Discussion
The SC bleaching method presented here is simple, fast and versatile: (i) It enables revealing single SC morphology and dynamics by confocal microscopy based on a single transgene - which is a significant advantage when combining the approach e.g. with disease models that require additional alleles. (ii) The method is rapid, as from dissection to fixation it can be performed in half a day. (iii) The number of applications is broad, as it can be combined with other methods, such as cell ablations, imaging organelles, electrophysiology or immunohistochemistry. Additionally, the approach presented here for SCs at murine NMJs can be generalized to other cells, tissues or species - one example of an application of a related technique in zebrafish is presented in reference 6. Also other fluorescent proteins besides GFP can be used for bleaching and time lapse microscopy, for example YFP or CFP. While we have not used these fluorescent proteins in SCs, bleaching of axons shows that a less stable fluorescent protein (e.g. CFP) in this setting can be advantageous. However, a compromise between efficient bleaching and stable imaging has to be found, depending on the desired results (e.g. single image vs. time-lapse of the remaining un-bleached cells).
There are a number of limitations to the technique, however. One relates to the use of an explanted (and hence axotomized) muscle, which limits the imaging period. In previous studies we have found that - depending on the question under study - the triangularis sterni explant can be used for a maximum of 3-6 hr. Still, sequential photo-bleaching can be used in vivo, overcoming this limitation provided the sample can be sufficiently stabilized to obtain images that can be subtracted. Two further aspects need to be kept in mind, when analysing data resulting from the sequential bleaching approach: (i) Phototoxicity could alter SC dynamism, as killing a terminal SC results in rapid expansion of its neighbours11. So plausibility checks should be included, e.g. measuring the sum of expansions and retractions in a stable synapse. On average, the population of imaged SCs should not significantly grow or shrink. (ii) The morphology of all but the last SC is obtained by image subtraction. The quality of such an image depends on the obtained bleaching contrast. This is generally achieved easier in brighter mouse lines. For example, Plp-GFP works better than S100-GFP12. Image subtraction adds noise, so the last SCs is generally revealed in more detail than the others (especially also, as post-hoc imaging after fixation allows using high numerical aperture oil objectives). Hence, a morphological feature should only be accepted as real, if it is visible in "last" SCs, as well as in bleached ones.
Disclosures
No conflicts of interest declared.
Acknowledgments
We would like to thank Manuela Budak, Ljiljana Marinkovi and Kristina Wullimann for excellent technical assistance. We thank W. Macklin for providing Plp-GFP mice. T.M. is supported by the Institute of Advanced Studies (Technische Universität München), by the Deutsche Forschungsgemeinschaft (DFG; Sonderforschungsbereich SFB 596), by the Alexander-von-Humboldt-Foundation and by the national funding agency ("Bundesministerium für Bildung und Forschung") in the frame of ERA-Net NEURON "iPSoALS" and "2-photon imaging". T.M. and M.B. are supported by the Center for Integrated Protein Science (Munich). P.M. was supported by the Graduate School of Technische Universität München (TUM-GS).
References
- Feng G, et al. Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron. 2000;28:41–51. doi: 10.1016/s0896-6273(00)00084-2. [DOI] [PubMed] [Google Scholar]
- Mallon BS, Shick HE, Kidd GJ, Macklin WB. Proteolipid promoter activity distinguishes two populations of NG2-positive cells throughout neonatal cortical development. J. Neurosci. 2002;22:876–885. doi: 10.1523/JNEUROSCI.22-03-00876.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brill MS, Lichtman JW, Thompson W, Zuo Y, Misgeld T. Spatial constraints dictate glial territories at murine neuromuscular junctions. The Journal of cell biology. 2011;195:293–305. doi: 10.1083/jcb.201108005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livet J, et al. Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature. 2007;450:56–62. doi: 10.1038/nature06293. [DOI] [PubMed] [Google Scholar]
- Thompson WJ, et al. Transgenic mice with photoswitchable GFP in Schwann cells. Society for Neuroscience. 2008;Program 241 [Google Scholar]
- Williams PR, et al. In vivo development of outer retinal synapses in the absence of glial contact. J. Neurosci. 2010;30:11951–11961. doi: 10.1523/JNEUROSCI.3391-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerschensteiner M, Reuter MS, Lichtman JW, Misgeld T. Ex vivo imaging of motor axon dynamics in murine triangularis sterni explants. Nat. Protoc. 2008;3:1645–1653. doi: 10.1038/nprot.2008.160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McArdle JJ, et al. Advantages of the triangularis sterni muscle of the mouse for investigations of synaptic phenomena. J. Neurosci. Methods. 1981;4:109–115. doi: 10.1016/0165-0270(81)90044-3. [DOI] [PubMed] [Google Scholar]
- Marinkovi P, et al. Axonal transport deficits and degeneration can evolve independently in mouse models of amyotrophic lateral sclerosis. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:4296–4301. doi: 10.1073/pnas.1200658109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thevenaz P, Ruttimann UE, Unser M. A pyramid approach to subpixel registration based on intensity. IEEE Trans Image Process. 1998;7:27–41. doi: 10.1109/83.650848. [DOI] [PubMed] [Google Scholar]
- Brill MS, Lichtman JW, Thompson W, Zuo Y, Misgeld T. Spatial constraints dictate glial territories at murine neuromuscular junctions. The Journal of cell biology. 2011;195:293–305. doi: 10.1083/jcb.201108005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuo Y, et al. Fluorescent proteins expressed in mouse transgenic lines mark subsets of glia, neurons, macrophages, and dendritic cells for vital examination. J. Neurosci. 2004;24:10999–11009. doi: 10.1523/JNEUROSCI.3934-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]