

Abstract
Peroxisome proliferator-activated receptors (PPAR) are members of the superfamily of nuclear hormone receptors involved in embryonic development and differentiation of several tissues including placenta, which respond to specific ligands such as polyunsaturated fatty acids by altering gene expression. Three subtypes of this receptor have been discovered, each evolving to achieve different biological functions. The PPARs also control a variety of target genes involved in lipid homeostasis. Similar to other nuclear receptors, the transcriptional activity of PPARs is affected not only by ligand-stimulation but also by crosstalk with other molecules. For example, both PPARs and the RXRs are ligand-activated transcription factors that coordinately regulate gene expression. In addition, several mechanisms underlying negative regulation of gene expression by PPARs have been shown. It is suggested that PPARs are key messengers responsible for the translation of nutritional stimuli into changes in gene expression pathways for placental development.
1. Introduction
Peroxisome proliferator-activated receptors (PPARs) are a family of ligand activated transcription factors belonging to the nuclear hormone receptor superfamily, which mainly regulate the expression of target genes involved in lipid and energy metabolism [1–3]. Three PPAR isotypes have been identified in mammals termed PPARα, PPARβ/δ, and PPARγ [4, 5]. Each isotype is a product of a separate gene, and each one has a distinct tissue distribution relating to the distinct functions. The PPARs play key roles in the metabolic syndrome and overall health of organisms including regeneration of tissues, differentiation, lipid metabolism, and immune response [6]. From a nutritional viewpoint, the PPARs are of importance because of their ability to be activated by long chain fatty acids and their metabolites [7]. Therefore, the PPARs are recognized as candidates in order to improve metabolism and health through suitable diet. In addition, several evidences show the important role of PPARs in reproductive organs [8, 9]. PPARγ expression has been found in the granulosa, theca, and luteal cells [10]. The PPARγ may regulate the differentiation and proliferation of the ovarian cells, steroidogenesis, angiogenesis, and prostaglandin production [11], indicating that PPARs modulate the estrous cycle and pregnancy. Retinoic X receptor (RXR) is a functional partner of PPAR. RXRα and PPARγ function potently in metabolic diseases and are both important targets for antidiabetic drugs. Coactivation of RXRα and PPARγ is believed to synergize their effects on glucose and lipid metabolism [12]. The RXRα and PPARγ are essential for mouse placentogenesis [13, 14]. PPARγ is important for mouse placenta morphology [15]. In addition, PPARs have also been implicated in several aspects of early pregnancy development including implantation, placentation, and trophoblast differentiation [16–18]. Furthermore, PPARγ and RXRα are essential for cytotrophoblast cell fusion into a syncytiotrophoblast, which is obligatory for placentation, and these expressions are deregulated in pathological placenta [19]. So, the PPARs may be a link between energy metabolism and reproduction, which is frequently associated with insulin resistance. This paper will focus on the evidences of PPARs functions in placenta. We will also highlight the effects of co-modulators such as RXRs with PPARs in the experimental models.
2. Expression and Characteristics of PPAR
PPARs (α, β, and γ) are nuclear hormone receptors that are known to regulate gene transcription and protein expression levels of fatty acid transport and metabolism mediating proteins through the formation of a DNA binding heterodimer complex [2–4]. All distinct PPAR subtypes share a high degree of structural homology with other members of the superfamily, particularly in the DNA-binding domain and ligand-binding domain (Figure 1). PPARs exhibit wide-ranging and isotype-specific tissue expression pattern [3–5]. PPARα is expressed at high levels in tissues that catabolize fatty acids [20], as in the adult liver, heart, kidney, large intestine, and skeletal muscle. PPARβ/δ mRNA is ubiquitously distributed with a higher expression in the digestive tract and the placenta [21]. PPARγ is mostly expressed in the adipose tissue [22] and immune system. The three isotypes are expressed as early as week 7 of gestation in endodermal and mesodermal origin cells [23]. There are limiting data describing the PPARs expression in endometrial tissue of animal species through the estrous cycle. PPARα and PPARβ transcript levels show similar profiles during the estrous cycle [23–26]. PPARγ mRNA level is quite stable during entire estrous cycle [24–26]. However, the precise role of PPARs in the uterus is not well known, although PPARα, PPARβ, or PPARγ expressions have been known in uterus of various species. High levels during the luteal phase and low during the follicular suggest the association with steroids function.
Figure 1.
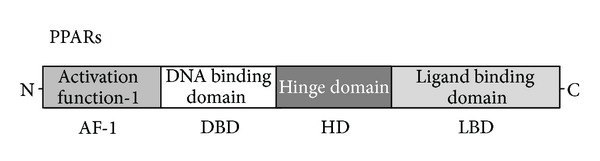
Schematic structure of PPARs protein. The predicted consensual important domain structures for each PPAR are depicted, which are common in fish species. AF-1 = activation function-1, DBD = DNA binding domain, HD = hinge domain linking DBD and LBD, LBD = C-terminal ligand binding domain.
A wide variety of compounds have been identified as PPARs ligands. Among the synthetic ligands, fibrates and thiazolidinediones are PPARα and PPARγ agonists, respectively [27]. PPARγ is also activated by prostaglandins and leukotrienes. In the presence of ligands, conformational changes of the ligand binding domain result in the recruitment of coactivator proteins, release of corepressor proteins, and subsequent assembly of a protein complex that enhances transcription of the target genes [28, 29]. A PPARα specific ligand (8S-HETE), a PPARγ ligand (15-deoxy-delta12, 14-prostaglandin J2), and a peroxisome proliferator (clofibrate) are all able to induce expression of both PPARα and PPARγ [30–32]. Subsequent work has led to the identification of various PPAR ligands that include eicosanoids, hypolipidemic agents, and antidiabetic drugs [33, 34].
Ligand activated PPARs bind as heterodimers with the RXRs on PPAR response elements. A number of PPAR target genes have been characterized to date. Most of these genes are known to have roles in lipid and glucose metabolism [35]. The endometrium is a possible place where PPARs may regulate cyclooxygenase- (COX-) 2 which catalyzes prostaglandin production [36]. They are critical to sustain the function of corpus luteum during the estrous cycle. The PPAR response element has been found upstream of the COX-2 transcriptional site. So, the activation of PPARs affects COX-2 expression in the epithelial cells. In COX-2 deficient mice, failures during the implantation and decidualization may be restored by administration of PPARβ agonists [37], suggesting common pathways of these molecules. Moreover, studies suggest that PPARs participate in uterine functions such as steroidogenesis, cytokine production, and angiogenesis during the estrous cycle and/or pregnancy [38, 39]. Interestingly, PPARs also downregulate nitric oxide synthase (NOS) in human cardiac myocytes and in human prostate cells [40]. As PPARs are expressed as cytotrophoblasts and syncytiotrophoblasts in the placenta, the activation of PPARs may stimulate the production and secretion of hormones such as gonadotropin required during pregnancy and fetal development [41, 42]. Thus, PPARs is essential for the maturation of a functional placenta.
3. Interaction with Retinoid X Receptor for Transactivation
PPARs bind to a variety of PPAR response elements (PPREs) present in the promoter regions of the responsive genes. The transcriptional regulation by PPARs requires heterodimerization with the retinoid X receptor (RXR) (Figure 2). Retinoic acid affects a broad spectrum of physiological processes, including cell growth, differentiation, morphogenesis, reproduction, and development [43], through the action of two types of receptors, the retinoic acid receptors (RARs) and the retinoid-X-receptors (RXRs). When activated by a ligand, the heterodimer modulates transcription activity. The transcriptional control by the PPAR/RXR heterodimer also requires interaction with coregulator complexes [44]. Thus, selective action of PPARs in vivo results from the interplay at a time point of each of the cofactors available. The RXRs are able to influence the transcription of a wide variety of genes, because they can activate gene transcription by binding to specific sites on DNA as homodimers and/or as the heterodimers with other related nuclear receptors including the PPARs, vitamin D receptor, and thyroid hormone receptors [45–47]. The temporal and spatial patterns of expression of PPARs and RXRs isoforms in the developing placenta have been elucidated [48]. In the human placenta, PPARα, PPARβ, and PPARγ are observed, while RXRβ is not detected. Immunocytochemistry staining results also determine the presence of PPARα, PPARβ, PPARγ, RXRα, and RXRγ to be specific to the trophoblast layer of the human chorionic villi [49]. The presence of PPAR and RXR isoforms in placenta suggests that PPAR and RXR isoforms are potential regulators of placental lipid transfer and homeostasis. The PPARs/RXRs heterodimers may play a key regulatory role in placental development. It has been suggested that the PPARs/RXRs heterodimers may also function in the modulation of trophoblast invasion. There was no significant difference in PPARs or RXRs protein expression in both amnion and choriodecidua, which have been identified in gestational tissues [50–52].
Figure 2.
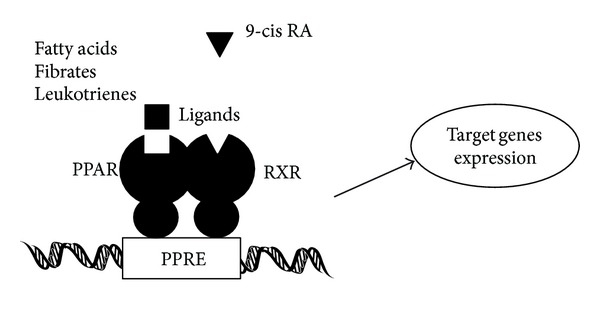
Schematic depiction of the model of mechanism of PPAR action. Similar to other nuclear hormone receptors, PPARs act as a ligand activated transcription factor. PPARs in response to the ligand binding heterodimerize with RXR and bind PPRE DNA sequences in the promoters of target genes, which are often involved in the lipid metabolism. Note that some critical molecules have been omitted for clarity.
4. Functional Interplay for the Transrepression of PPARs
The NAD(+)-dependent histone deacetylase Sir2 regulates life-span in various species [53]. Mammalian homologs of Sir2 are called Sirtuins (SIRT1–SIRT7) [54]. PPARα and SIRT1 coordinately suppress genes involved in mitochondrial function [55] (Figure 3). Calorie restriction extends lifespan in organisms ranging from yeast to mammals. Upon food withdrawal, SIRT1 protein binds to and represses genes controlled by the fat regulator PPARγ, including genes mediating fat storage. SIRT1 represses PPARγ by docking with its cofactors nuclear receptor corepressor and silencing mediator of retinoid and thyroid hormone receptors [56, 57]. The repression of PPARγ transactivation by SIRT1 inhibits lipid accumulation in adipocytes. SIRT1 also regulates angiogenesis signaling [58], which is expressed in the vasculature during blood vessel growth. Loss of SIRT1 function blocks sprouting angiogenesis and branching morphogenesis of endothelial cells with consequent downregulation of genes involved in blood vessel development and vascular remodeling. Human SIRT1 and SIRT2 are localized in the syncytiotrophoblast layer and the cytotrophoblasts of the placenta, amnion epithelium, trophoblast layer of the chorion, and decidual cells [59]. Resveratrol decreases proinflammatory TNF, IL6, and IL8 gene expression and resultant prostaglandin release from the gestational tissues [59]. SIRT1 also modulates gene expression in target tissues by regulating transcriptional coregulators or by directly interacting with transcription factors. SIRT1 overexpression prevents cytokine-mediated cytotoxicity, nitric oxide (NO) production, and inducible NO synthase expression. PPARs and SIRT1 may play a pivotal role in regulating pregnancy and parturition [59].
Figure 3.
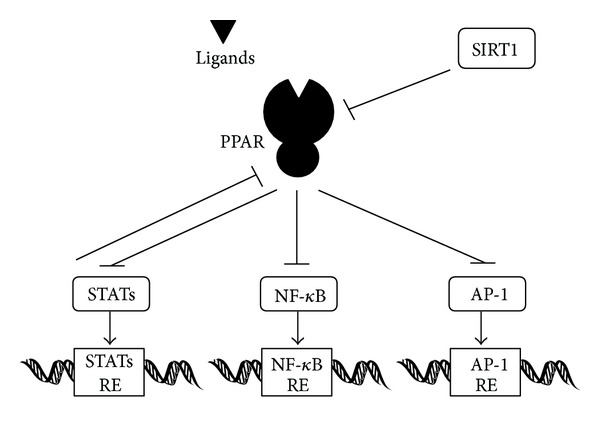
A hypothetical schematic implication of some of the PPAR regulatory network. Examples of molecules known to interact with PPARs pathway in mammals are shown. Hammerheads mean inhibition. Note that some critical pathways have been omitted for clarity.
Many of the anti-inflammatory effects of PPARγ are caused by antagonizing the activities of the transcription factors including nuclear factor-kappa B (NF-κB) (Figure 3). EPA inhibited the NF-κB pathway in myotubes in a PPARγ-dependent manner. In one way for the inhibition, PPARs and these transcription factors bind each other via protein-protein interactions and prevent binding to their response elements. The ligand-activated PPARs have been shown to interfere with DNA binding of both AP-1 and NF-κB activity [60]. Furthermore, the mitogen-activated protein kinase (MAPK) pathway is also regulated by PPARs at different levels [61]. In addition, activation of PPARγ reduces c-Jun N-terminal kinase (JNK) and p38 MAPK activation, leading to downregulation of proinflammatory gene expression [62]. The transcription factors NFκB, CCAAT/enhancer-binding protein (CEBP), and AP-1 are important transcription factor families that are involved in immune and inflammatory functions as well as in cell growth and differentiation. Human placenta is rich in diverse bioactive molecules, whose extract induces interleukin mRNA and protein expressions in a dose-dependent manner. For example, the IL8 promoter contains binding sites for the NFκB, AP-1, and CEBP. The IL-8 expression is inhibited by an inhibitor of JNK [63, 64].
Interestingly, the transcriptional expression levels of fatty acid binding proteins are upregulated in males and downregulated in females [65]. A similar trend between sexes occurs for PPARs and CEBPs, which may be the upstream regulatory elements [65]. Estrogen-related receptors have been identified as PPAR coactivators, which upregulate the expression of PPARα and PPARα-regulated genes. Estrogen has not been reported to be a PPAR ligand, but interactions between PPARs and ER proteins and their response elements have been described [66]. These interactions might be due to estrogen induced production of PPAR activating metabolites. Studies have found that 17beta-estradiol upregulates the expression of PPARα in skeletal muscle of rats [67].
A direct relationship between the PPARs activation and the inhibition of STAT5 mediated transcription has been reported [68, 69]. The PPARs do not block STAT5 tyrosine phosphorylation or do not inhibit DNA-binding activity but inhibit the transcriptional activity of STAT5. Conversely, activated STAT5 is able to inhibit PPAR-regulated gene transcription. In other words, STAT5-activating hormones and cytokines may modulate the responsiveness of PPARs to the chemical ligands. The cross-inhibition between PPAR and STAT5 proceeds in a synchronized and bidirectional manner (Figure 3). Exposure to environmental chemical activators of PPARs may thus lead to alteration of hormone induced STAT5-regulated gene expression in tissues such as placenta, where both transcription factors are expressed.
5. Perspective
PPARs are lipid-activated transcription factors that have emerged as key regulators of both lipid metabolism and inflammation, and they exert positive and negative controls over the expression of a range of genes. However, the range of transcription factors affected and the molecular mechanism involved may be different for each PPAR isoforms and cell types. Furthermore, peroxisome proliferators induce numerous alterations in lipid metabolism. A comparative approach to bring together physiological and nutritional roles of PPARs across species appears critical. It is now clear that PPARs are important in the control of placental development. PPARs may play a key role in linking lipid metabolism and reproduction systems. In addition, the PPARγ/RXRα signaling is important in human cytotrophoblast and cell fusions. A disturbed PPARγ/RXRα pathway could contribute to pathological human pregnancies. SIRT1 expression is downregulated by proinflammatory cytokines. Possessing anti-inflammatory action in human gestational tissues, the SIRT1 expression is downregulated by proinflammatory cytokines. The natural polyphenol resveratrol inhibits cytokine and prostaglandin release via the SIRT1 activation. Both mRNA and protein levels of SIRT1 are shown to decrease in placenta and fetal membranes after labor onset, which may contribute to uterine contractions associated with labor. It would be of interest to investigate the impact of different types of fatty acid, integrated into food, on ovulation capacity and fetal development. Further study of PPARs, RXRs, and SIRTs functions in placenta may indicate pathways that are common to critical processes, providing additional focus for research in important human placental diseases. In parallel, defining more specific mode of action by identifying the endogenous coactivators and modulators of these transcription factors in animal models will help to build more efficient therapeutic strategy for the diseases. Future studies using functional genomic approaches will be required to more clearly establish the complicated mechanisms by which PPARs exert their actions. Additional insight is also needed into endogenous PPAR and RXR ligands, how these molecules are formed, and how they are delivered to the nucleus in placenta. Furthermore, additional experiments are required to increase the knowledge of the way in which lipid metabolism influences reproductive functions.
Conflict of Interests
The authors declare that they have no conflict of interests.
Acknowledgments
This work was supported by grants-in-aid from the Ministry of Education, Culture, Sports, Science and Technology in Japan. In addition, this work was supported in part by the grant from SHIN-EI Pharmaceutical Co., Ltd.
Abbreviations
- AP-1:
Activator protein-1
- CEBP:
CCAAT/enhancer-binding protein
- COX:
Cyclooxygenase
- JNK:
c-Jun N-terminal kinase
- MAPK:
Mitogen activated protein kinase
- NAD(+):
Nicotinamide adenine dinucleotide
- NF-κB:
Nuclear factor-kappa B
- NO:
Nitric oxide
- PPAR:
Peroxisome proliferator-activated receptor
- PPRE:
PPAR response element
- RA:
Retinoic acid
- RE:
Response element
- RXR:
Retinoid X receptor
- SIRT1:
Silent mating type information regulation 2 homolog 1
- STAT:
Signal transducers and activators of transcription.
References
- 1.Vacca M, Degirolamo C, Mariani-Costantini R, Palasciano G, Moschetta A. Lipid-sensing nuclear receptors in the pathophysiology and treatment of the metabolic syndrome. Wiley Interdisciplinary Reviews: Systems Biology and Medicine. 2011;3(5):562–587. doi: 10.1002/wsbm.137. [DOI] [PubMed] [Google Scholar]
- 2.Schulman IG. Nuclear receptors as drug targets for metabolic disease. Advanced Drug Delivery Reviews. 2010;62(13):1307–1315. doi: 10.1016/j.addr.2010.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wahli W, Michalik L. PPARs at the crossroads of lipid signaling and inflammation. Trends in Endocrinology & Metabolism. 2012;23(7):351–363. doi: 10.1016/j.tem.2012.05.001. [DOI] [PubMed] [Google Scholar]
- 4.Becker J, Delayre-Orthez C, Frossard N, Pons F. Regulation of inflammation by PPARs: a future approach to treat lung inflammatory diseases? Fundamental and Clinical Pharmacology. 2006;20(5):429–447. doi: 10.1111/j.1472-8206.2006.00425.x. [DOI] [PubMed] [Google Scholar]
- 5.Giaginis C, Tsantili-Kakoulidou A, Theocharis S. Peroxisome proliferator-activated receptors (PPARs) in the control of bone metabolism. Fundamental and Clinical Pharmacology. 2007;21(3):231–244. doi: 10.1111/j.1472-8206.2007.00486.x. [DOI] [PubMed] [Google Scholar]
- 6.Nagy L, Szanto A, Szatmari I, Széles L. Nuclear hormone receptors enable macrophages and dendritic cells to sense their lipid environment and shape their immune response. Physiological Reviews. 2012;92(2):739–789. doi: 10.1152/physrev.00004.2011. [DOI] [PubMed] [Google Scholar]
- 7.Kruger MC, Coetzee M, Haag M, Weiler H. Long-chain polyunsaturated fatty acids: selected mechanisms of action on bone. Progress in Lipid Research. 2010;49(4):438–449. doi: 10.1016/j.plipres.2010.06.002. [DOI] [PubMed] [Google Scholar]
- 8.Martínez N, Kurtz M, Capobianco E, Higa R, White V, Jawerbaum A. PPARα agonists regulate lipid metabolism and nitric oxide production and prevent placental overgrowth in term placentas from diabetic rats. Journal of Molecular Endocrinology. 2011;47(1):1–12. doi: 10.1530/JME-10-0173. [DOI] [PubMed] [Google Scholar]
- 9.Shalom-Barak T, Nicholas JM, Wang Y, et al. Peroxisome proliferator-activated receptor γ controls Muc1 transcription in trophoblasts. Molecular and Cellular Biology. 2004;24(24):10661–10669. doi: 10.1128/MCB.24.24.10661-10669.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Puttabyatappa M, VandeVoort CA, Chaffin CL. hCG-induced down-regulation of PPARγ and liver X receptors promotes periovulatory progesterone synthesis by macaque granulosa cells. Endocrinology. 2010;151(12):5865–5872. doi: 10.1210/en.2010-0698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kowalewski MP, Dyson MT, Manna PR, Stocco DM. Involvement of peroxisome proliferator-activated receptor in gonadal steroidogenesis and steroidogenic acute regulatory protein expression. Reproduction, Fertility and Development. 2009;21(7):909–922. doi: 10.1071/RD09027. [DOI] [PubMed] [Google Scholar]
- 12.Zhang H, Xu X, Chen L, et al. Molecular determinants of magnolol targeting both RXRα and PPARγ . PLoS One. 2011;6(11, article e28253) doi: 10.1371/journal.pone.0028253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wendling O, Chambon P, Mark M. Retinoid X receptors are essential for early mouse development Land placentogenesis. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(2):547–551. doi: 10.1073/pnas.96.2.547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Barak Y, Nelson MC, Ong ES, et al. PPARγ is required for placental, cardiac, and adipose tissue development. Molecular Cell. 1999;4(4):585–595. doi: 10.1016/s1097-2765(00)80209-9. [DOI] [PubMed] [Google Scholar]
- 15.Schaiff WT, Knapp FF, Jr., Barak Y, Biron-Shental T, Nelson DM, Sadovsky Y. Ligand-activated peroxisome proliferator activated receptor γ alters placental morphology and placental fatty acid uptake in mice. Endocrinology. 2007;148(8):3625–3634. doi: 10.1210/en.2007-0211. [DOI] [PubMed] [Google Scholar]
- 16.Lambe KG, Tugwood JD. A human peroxisome-proliferator-activated receptor-γ is activated by inducers of adipogenesis, including thiazalidinedione drugs. European Journal of Biochemistry. 1996;239(1):1–7. doi: 10.1111/j.1432-1033.1996.0001u.x. [DOI] [PubMed] [Google Scholar]
- 17.Schild RL, Sonnenberg-Hirche CM, Schaiff WT, Bildirici I, Nelson DM, Sadovsky Y. The kinase p38 regulates peroxisome proliferator activated receptor-γ in human trophoblasts. Placenta. 2006;27(2-3):191–199. doi: 10.1016/j.placenta.2005.01.012. [DOI] [PubMed] [Google Scholar]
- 18.Degrelle SA, Murthi P, Evain-Brion D, Fournier T, Hue I. Expression and localization of DLX3, PPARG and SP1 in bovine trophoblast during binucleated cell differentiation. Placenta. 2011;32(11):917–920. doi: 10.1016/j.placenta.2011.08.014. [DOI] [PubMed] [Google Scholar]
- 19.Ruebner M, Langbein M, Strissel PL, et al. Regulation of the human endogenous retroviral Syncytin-1 and cell-cell fusion by the nuclear hormone receptors PPARγ/RXRα in placentogenesis. Journal of Cellular Biochemistry. 2012;113(7):2383–2396. doi: 10.1002/jcb.24110. [DOI] [PubMed] [Google Scholar]
- 20.Ringseis R, Eder K. Influence of pharmacological PPARα activators on carnitine homeostasis in proliferating and non-proliferating species. Pharmacological Research. 2009;60(3):179–184. doi: 10.1016/j.phrs.2009.01.016. [DOI] [PubMed] [Google Scholar]
- 21.Wagner KD, Wagner N. Peroxisome proliferator-activated receptor beta/delta (PPARβ/δ) acts as regulator of metabolism linked to multiple cellular functions. Pharmacology and Therapeutics. 2010;125(3):423–435. doi: 10.1016/j.pharmthera.2009.12.001. [DOI] [PubMed] [Google Scholar]
- 22.Cipolletta D, Feuerer M, Li A, et al. PPAR-γ is a major driver of the accumulation and phenotype of adipose tissue Treg cells. Nature. 2012;486(7404):549–553. doi: 10.1038/nature11132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Huin C, Corriveau L, Bianchi A, et al. Differential expression of peroxisome proliferator-activated receptors (PPARs) in the developing human fetal digestive tract. Journal of Histochemistry and Cytochemistry. 2000;48(5):603–611. doi: 10.1177/002215540004800504. [DOI] [PubMed] [Google Scholar]
- 24.Komar CM, Curry TE. Localization and expression of messenger RNAs for the peroxisome proliferator-activated receptors in ovarian tissue from naturally cycling and pseudopregnant rats. Biology of Reproduction. 2002;66(5):1531–1539. doi: 10.1095/biolreprod66.5.1531. [DOI] [PubMed] [Google Scholar]
- 25.Fedele L, Somigliana E, Frontino G, Benaglia L, Vigano P. New drugs in development for the treatment of endometriosis. Expert Opinion on Investigational Drugs. 2008;17(8):1187–1202. doi: 10.1517/13543784.17.8.1187. [DOI] [PubMed] [Google Scholar]
- 26.Gui Y, Cai Z, Silha JV, Murphy LJ. Variations in parametrial white adipose tissue mass during the mouse estrous cycle: relationship with the expression of peroxisome proliferator-activated receptor-γ and retinoic acid receptor-α . Canadian Journal of Physiology and Pharmacology. 2006;84(8-9):887–892. doi: 10.1139/y06-032. [DOI] [PubMed] [Google Scholar]
- 27.Benz V, Kintscher U, Foryst-Ludwig A. Sex-specific differences in type 2 diabetes mellitus and dyslipidemia therapy: PPAR agonists. Handbook of Experimental Pharmacology. 2012;214:387–410. doi: 10.1007/978-3-642-30726-3_18. [DOI] [PubMed] [Google Scholar]
- 28.Waku T, Shiraki T, Oyama T, Morikawa K. Atomic structure of mutant PPARγ LBD complexed with 15d-PGJ2: novel modulation mechanism of PPARγ/RXRα function by covalently bound ligands. FEBS Letters. 2009;583(2):320–324. doi: 10.1016/j.febslet.2008.12.017. [DOI] [PubMed] [Google Scholar]
- 29.Yu S, Reddy JK. Transcription coactivators for peroxisome proliferator-activated receptors. Biochimica et Biophysica Acta. 2007;1771(8):936–951. doi: 10.1016/j.bbalip.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 30.Forman BM, Tontonoz P, Chen J, Brun RP, Spiegelman BM, Evans RM. 15-deoxy-Δ12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPARγ . Cell. 1995;83(5):803–812. doi: 10.1016/0092-8674(95)90193-0. [DOI] [PubMed] [Google Scholar]
- 31.Yu Z, Schneider C, Boeglin WE, Brash AR. Epidermal lipoxygenase products of the hepoxilin pathway selectively activate the nuclear receptor PPARα . Lipids. 2007;42(6):491–497. doi: 10.1007/s11745-007-3054-4. [DOI] [PubMed] [Google Scholar]
- 32.Ibabe A, Herrero A, Cajaraville MP. Modulation of peroxisome proliferator-activated receptors (PPARs) by PPARα- and PPARγ-specific ligands and by 17β-estradiol in isolated zebrafish hepatocytes. Toxicology in Vitro. 2005;19(6):725–735. doi: 10.1016/j.tiv.2005.03.019. [DOI] [PubMed] [Google Scholar]
- 33.Kouroumichakis I, Papanas N, Zarogoulidis P, Liakopoulos V, Maltezos E, Mikhailidis DP. Fibrates: therapeutic potential for diabetic nephropathy? European Journal of Internal Medicine. 2012;23(4):309–316. doi: 10.1016/j.ejim.2011.12.007. [DOI] [PubMed] [Google Scholar]
- 34.Friedland SN, Leong A, Filion KB, et al. The cardiovascular effects of peroxisome proliferator-activated receptor agonists. American Journal of Medicine. 2012;125(2):126–133. doi: 10.1016/j.amjmed.2011.08.025. [DOI] [PubMed] [Google Scholar]
- 35.Saraf N, Sharma PK, Mondal SC, Garg VK, Singh AK. Role of PPARg2 transcription factor in thiazolidinedione-induced insulin sensitization. Journal of Pharmacy and Pharmacology. 2012;64(2):161–171. doi: 10.1111/j.2042-7158.2011.01366.x. [DOI] [PubMed] [Google Scholar]
- 36.Subbaramaiah K, Lin DT, Hart JC, Dannenberg AJ. Peroxisome proliferator-activated receptor γ ligands suppress the transcriptional activation of cyclooxygenase-2. Evidence for involvement of activator protein-1 and CREB-binding protein/p300. Journal of Biological Chemistry. 2001;276(15):12440–12448. doi: 10.1074/jbc.M007237200. [DOI] [PubMed] [Google Scholar]
- 37.Matsumoto H, Ma W, Smalley W, Trzaskos J, Breyer RM, Dey SK. Diversification of cyclooxygenase-2-derived prostaglandins in ovulation and implantation. Biology of Reproduction. 2001;64(5):1557–1565. doi: 10.1095/biolreprod64.5.1557. [DOI] [PubMed] [Google Scholar]
- 38.Ruan KH, Dogné JM. Implications of the molecular basis of prostacyclin biosynthesis and signaling in pharmaceutical designs. Current Pharmaceutical Design. 2006;12(8):925–941. doi: 10.2174/138161206776055994. [DOI] [PubMed] [Google Scholar]
- 39.Froment P, Gizard F, Defever D, Staels B, Dupont J, Monget P. Peroxisome proliferator-activated receptors in reproductive tissues: from gametogenesis to parturition. Journal of Endocrinology. 2006;189(2):199–209. doi: 10.1677/joe.1.06667. [DOI] [PubMed] [Google Scholar]
- 40.Mendez M, LaPointe MC. PPARγ inhibition of cyclooxygenase-2, PGE2 synthase, and inducible nitric oxide synthase in cardiac myocytes. Hypertension. 2003;42(4):844–850. doi: 10.1161/01.HYP.0000085332.69777.D1. [DOI] [PubMed] [Google Scholar]
- 41.Fournier T, Pavan L, Tarrade A, et al. The role of PPAR-γ/RXR-α heterodimers in the regulation of human trophoblast invasion. Annals of the New York Academy of Sciences. 2002;973:26–30. doi: 10.1111/j.1749-6632.2002.tb04601.x. [DOI] [PubMed] [Google Scholar]
- 42.Tarrade A, Schoonjans K, Guibourdenche J, et al. PPARγ/RXRα heterodimers are involved in human CGβ synthesis and human trophoblast differentiation. Endocrinology. 2001;142(10):4504–4514. doi: 10.1210/endo.142.10.8448. [DOI] [PubMed] [Google Scholar]
- 43.Tarrade A, Rochette-Egly C, Guibourdenche J, Evain-Brion D. The expression of nuclear retinoid receptors in human implantation. Placenta. 2000;21(7):703–710. doi: 10.1053/plac.2000.0568. [DOI] [PubMed] [Google Scholar]
- 44.Michalik L, Auwerx J, Berger JP, et al. International union of pharmacology. LXI. Peroxisome proliferator-activated receptors. Pharmacological Reviews. 2006;58(4):726–741. doi: 10.1124/pr.58.4.5. [DOI] [PubMed] [Google Scholar]
- 45.Wolf G. Is 9-cis-retinoic acid the endogenous ligand for the retinoic acid-X receptor? Nutrition Reviews. 2006;64(12):532–538. doi: 10.1111/j.1753-4887.2006.tb00186.x. [DOI] [PubMed] [Google Scholar]
- 46.Nezbedova P, Brtko J. 1α,25-dihydroxyvitamin D3 inducible transcription factor and its role in the vitamin D action. Endocrine Regulations. 2004;38(1):29–38. [PubMed] [Google Scholar]
- 47.Clarke SD, Thuillier P, Baillie RA, Sha X. Peroxisome proliferator-activated receptors: a family of lipid-activated transcription factors. American Journal of Clinical Nutrition. 1999;70(4):566–571. doi: 10.1093/ajcn/70.4.566. [DOI] [PubMed] [Google Scholar]
- 48.Wang Q, Fujii H, Knipp GT. Expression of PPAR and RXR isoforms in the developing rat and human term placentas. Placenta. 2002;23(8-9):661–671. doi: 10.1053/plac.2002.0855. [DOI] [PubMed] [Google Scholar]
- 49.Handschuh K, Guibourdenche J, Cocquebert M, et al. Expression and regulation by PPARγ of hCG α- and β-subunits: comparison between villous and invasive extravillous trophoblastic Cells. Placenta. 2009;30(12):1016–1022. doi: 10.1016/j.placenta.2009.09.006. [DOI] [PubMed] [Google Scholar]
- 50.Holdsworth-Carson SJ, Permezel M, Rice GE, Lappas M. Preterm and infection-driven preterm labor: the role of peroxisome proliferator-activated receptors and retinoid X receptor. Reproduction. 2009;137(6):1007–1015. doi: 10.1530/REP-08-0496. [DOI] [PubMed] [Google Scholar]
- 51.Holdsworth-Carson SJ, Permezel M, Riley C, Rice GE, Lappas M. Peroxisome Proliferator-activated receptors and retinoid X receptor-alpha in term human gestational tissues: tissue specific and labour-associated Changes. Placenta. 2009;30(2):176–186. doi: 10.1016/j.placenta.2008.11.013. [DOI] [PubMed] [Google Scholar]
- 52.Borel V, Gallot D, Marceau G, Sapin V, Blanchon L. Placental implications of peroxisome proliferator-activated receptors in gestation and parturition. PPAR Research. 2008;2008:9 pages. doi: 10.1155/2008/758562.758562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lin SJ, Ford E, Haigis M, Liszt G, Guarente L. Calorie restriction extends yeast life span by lowering the level of NADH. Genes and Development. 2004;18(1):12–16. doi: 10.1101/gad.1164804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Potente M, Ghaeni L, Baldessari D, et al. SIRT1 controls endothelial angiogenic functions during vascular growth. Genes and Development. 2007;21(20):2644–2658. doi: 10.1101/gad.435107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Oka S, Alcendor R, Zhai P, et al. PPARα-Sirt1 complex mediates cardiac hypertrophy and failure through suppression of the ERR transcriptional pathway. Cell Metabolism. 2011;14(5):598–611. doi: 10.1016/j.cmet.2011.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Legutko A, Marichal T, Fiévez L, et al. Sirtuin 1 promotes Th2 responses and airway allergy by repressing peroxisome proliferator-activated receptor-γ activity in dendritic cells. The Journal of Immunology. 2011;187(9):4517–4529. doi: 10.4049/jimmunol.1101493. [DOI] [PubMed] [Google Scholar]
- 57.Picard F, Kurtev M, Chung N, et al. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-γ . Nature. 2004;429(6993):771–776. doi: 10.1038/nature02583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Miyashita H, Watanabe T, Hayashi H, et al. Angiogenesis inhibitor vasohibin-1 enhances stress resistance of endothelial cells via induction of SOD2 and SIRT1. PLoS One. 2012;7(10, article e46459) doi: 10.1371/journal.pone.0046459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lappas M, Mitton A, Lim R, Barker G, Riley C, Permezel M. SIRT1 is a novel regulator of key pathways of human labor. Biology of Reproduction. 2011;84(1):167–178. doi: 10.1095/biolreprod.110.086983. [DOI] [PubMed] [Google Scholar]
- 60.Kim JK, Mun S, Kim MS, Kim MB, Sa BK, Hwang JK. 5, 7-Dimethoxyflavone, an activator of PPARα/γ, inhibits UVB-induced MMP expression in human skin fibroblast cells. Experimental Dermatology. 2012;21(3):211–216. doi: 10.1111/j.1600-0625.2011.01435.x. [DOI] [PubMed] [Google Scholar]
- 61.Papageorgiou E, Pitulis N, Msaouel P, Lembessis P, Koutsilieris M. The non-genomic crosstalk between PPAR-γ ligands and ERK1/2 in cancer cell lines. Expert Opinion on Therapeutic Targets. 2007;11(8):1071–1085. doi: 10.1517/14728222.11.8.1071. [DOI] [PubMed] [Google Scholar]
- 62.Pang T, Wang J, Benicky J, Sánchez-Lemus E, Saavedra JM. Telmisartan directly ameliorates the neuronal inflammatory response to IL-1β partly through the JNK/c-Jun and NADPH oxidase pathways. Journal of Neuroinflammation. 2012;9(1):p. 102. doi: 10.1186/1742-2094-9-102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Patel N, Gonsalves CS, Malik P, Kalra VK. Placenta growth factor augments endothelin-1 and endothelin-B receptor expression via hypoxia-inducible factor-1α . Blood. 2008;112(3):856–865. doi: 10.1182/blood-2007-12-130567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Maldonado-Pérez D, Brown P, Morgan K, Millar RP, Thompson EA, Jabbour HN. Prokineticin 1 modulates IL-8 expression via the calcineurin/NFAT signaling pathway. Biochimica et Biophysica Acta. 2009;1793(7):1315–1324. doi: 10.1016/j.bbamcr.2009.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhang W, Zhang Y, Zhang H, Wang J, Cui R, Dai J. Sex differences in transcriptional expression of FABPs in zebrafish liver after chronic perfluorononanoic acid exposure. Environmental Science & Technology. 2012;46(9):5175–5182. doi: 10.1021/es300147w. [DOI] [PubMed] [Google Scholar]
- 66.Keller JM, Collet P, Bianchi A, et al. Implications of peroxisome proliferator-activated receptors (PPARS) in development, cell life status and disease. International Journal of Developmental Biology. 2000;44(5):429–442. [PubMed] [Google Scholar]
- 67.Campbell SE, Mehan KA, Tunstall RJ, Febbraio MA, Cameron-Smith D. 17β-Estradiol upregulates the expression of peroxisome proliferator-activated receptor α and lipid oxidative genes in skeletal muscle. Journal of Molecular Endocrinology. 2003;31(1):37–45. doi: 10.1677/jme.0.0310037. [DOI] [PubMed] [Google Scholar]
- 68.White UA, Stephens JM. Transcriptional factors that promote formation of white adipose tissue. Molecular and Cellular Endocrinology. 2010;318(1-2):10–14. doi: 10.1016/j.mce.2009.08.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Jung HS, Lee YJ, Kim YH, Paik S, Kim JW, Lee JW. Peroxisome proliferator-activated receptor gamma/signal transducers and activators of transcription 5A pathway plays a key factor in adipogenesis of human bone marrow-derived stromal cells and 3T3-L1 preadipocytes. Stem Cells and Development. 2012;21(3):465–475. doi: 10.1089/scd.2010.0591. [DOI] [PMC free article] [PubMed] [Google Scholar]