

Abstract
The E2F family of proteins was identified based on its role in promoting the G0 to S phase transition. Research over the past several years has unveiled considerable complexity within the family, with numerous studies pointing to delegation of function for distinct family members. More recent studies highlighted in this review have expanded this picture, suggesting ways in which E2F target gene expression is refined during cell cycle progression by facilitating the acquisition of promoter-specific histone modifications. E2F associated co-activators promote activating histone marks while recruitment of co-repressors associated with E2Fs and the pRB family leads to accretion of inhibitory histone modifications that provoke chromatin compaction.
Introduction
More than 20 years after its discovery, the E2F family of transcription factors continues to intrigue us, thanks to, or perhaps in spite of, constant technological advances and the use of genetically tractable model systems. The identification of E2F1, the founding member of the family, as a protein that promotes the transition to S phase catalyzed the notion that these regulatory proteins drive cell cycle progression. E2F activity was shown to be constrained by the retinoblastoma tumor suppressor family (pRB, together with the related proteins p107 and p130, so-called pocket proteins), providing solid evidence for its important role in mediating timely cell cycle regulated gene expression patterns. However, the identification of eight E2F family members and the development of new functional genomic approaches have accelerated our progress and proven that the function of the E2F family is substantially more complex. In addition, genome-wide approaches in mammalian cells, flies, and worms have indicated that this family controls expression of genes beyond the S phase transition and indeed beyond the cell cycle by promoting differentiation and specification of distinct tissues [1-6].
Here, we review the most recent discoveries relating to controls that fine-tune E2F-mediated gene expression during cell cycle progression and suggest areas of interest for future studies.
E2F target genes and gene repression by E2F
Recent studies have identified and characterized several evolutionarily conserved complexes involved in the regulation of E2F target genes. These complexes, termed dREAM (Drosophila RBF, E2F and Myb-interacting proteins) or Myb/MuvB, contain RBF1 or RBF2 (the homologs of mammalian pRB and p107/p130, respectively), dDP and dE2F2 (the only fly repressor E2F), CAF1p55 (the homolog of mammalian RbAp46), dMyb, as well as Myb-interacting proteins (Mips) Mip40, Mip120 and Mip130 [7,8]. dREAM-Myb/MuvB is involved in transcriptional repression: RNAi-mediated ablation of dREAM components leads to the transcriptional activation of genes regulated by dE2F factors, and the Mip130 and dE2F2 components directly bind their promoters. CAF1p55 has also been shown to be essential for the repression of certain E2F target genes [9]. However, this repressive effect of dREAM-Myb/MuvB appears to be specific to genes involved in development or sex-specific expression patterns [5]. These results suggest that a different dE2F-RBF complex may be required for the repression of cell cycle genes in this setting or that dREAM-Myb/MuvB represses cell cycle control targets under different conditions.
The dREAM complex was shown to bind exclusively to deacetylated histone H4, hinting at a mechanism whereby dREAM prevents acetylation of target gene promoters and maintains them in a repressed state. Initial deacetylation could presumably be mediated by a distinct dE2F-containing complex. Indeed, the Myb-MuvB complex incl udes dRpd3, the homolog of histone deacetylase HDAC1, and L(3)mbtl, a protein involved in triggering higher order compaction of the chromatin fiber (see below) [8]. The association of RBF with dMyb-containing complexes is noteworthy, given that both have been individually shown to be involved in the control of chorion gene duplication in flies [10,11].
An analogous complex has recently been purified from human cells [12-14]. Interestingly, Osterloh et al. found that the LIN-9 component is essential for cell cycle progression through the G2 phase and into mitosis. Its knock-down by RNAi prevents the induction of several genes that play important roles in mitosis, suggesting its involvement in their regulation. Intriguingly, LIN-9 can associate with E2F4 and p130 in G0 and with B-MYB in S phase, allowing it to bind the promoters of these genes during both the G2/M transition but also in quiescent (G0) cells (Figure 1). However, LIN-9 is not required for gene silencing in G0, but only for activation later in the G2 phase, like B-MYB. Therefore, it remains unclear how LIN-9 regulates gene expression. It could play a passive role in G0 through association with E2F4-p130, and presumably a more active role by association with B-MYB and recruitment of additional factors.
Figure 1. Speculative model describing co-factors that regulate E2F target genes during the cell cycle.
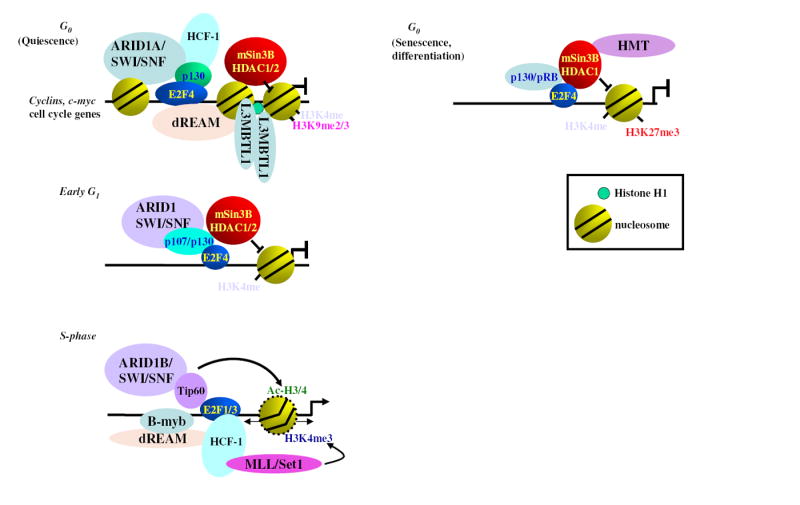
Major complexes discussed in this review are shown, but others may also exist. Interactions between distinct complexes are possible but not yet demonstrated. During cell cycle arrest, the DREAM complex associates with E2F4-p130 and E2F target promoters, whereas it associates with B-MYB in S phase. It remains to be determined if B-MYB-associated DREAM core-complex binds to B-MYB target promoters at this stage of the cycle. The HCF-1 complex promotes gene activation by binding activator E2Fs in S phase, recruiting the HMTase MLL/Set1 to E2F targets. The SWI/SNF complex also changes composition by incorporating ARID1A during quiescence but ARID1B in S phase, when E2F target genes are expressed. In the G0 phase, when cell cycle genes are not expressed, they may acquire a more compact state through the action of L(3)MBTL1. During senescence and terminal differentiation, characterized by an irreversible cell cycle arrest, specific histone marks may be involved, such as tri-methyl H3K27, which is deposited by the Polycomb Repressor Complex 2.
The biochemical composition of the complexes that exist in the G0 and in S phases [13] shows striking similarity between the human DREAM complex and its Drosophila counterpart. Unlike Drosophila dREAM, however, DREAM exhibits modular behavior in G0 and S-phase cells: in G0, a “core” complex (SynMuv class B proteins LIN9, LIN37, LIN52, LIN54 and RBBP4) associates with the p130/E2F4/DP1 repressor, whereas in S phase it binds B-MYB. The association between the core module and E2F4/p130 in quiescent cells is also supported by identification of overlapping LIN9, LIN54, E2F4 and p130 targets in genome-wide ChIP-on-chip experiments. Binding of the core complex to these loci in S-phase persists but is generally weaker and was ruled as insignificant by the authors. This discrepancy prompts further experiments to clarify the issue. In particular, it remains to be determined if binding of DREAM and B-MYB coincide in S-phase, as biochemical evidence and data from Osterloh et al. suggest. In line with an association with E2F4 and p130 in G0, DREAM complex components were found to be important for the repression of cell cycle control genes during quiescence, and their over-expression caused delays in cell cycle progression from G0 to S phase, although this result does not fully agree with those of Osterloh et al.
Given that no chromatin-remodeling activities were associated with the human DREAM, it remains to be determined how many complexes exist and how the core DREAM module can mediate repression in G0 and activation in S and G2 phases, and perhaps DNA replication.
Chromatin complexes that mediate E2F repression and compaction
Histone modifications provide a basis for modulating chromatin structure, and such modifications can be classified according to their association with repressed or active genes. Although there are exceptions, genes that are predominantly acetylated on histones H3 and H4 or methylated on lysine 4 of histone H3 (H3K4) have been activated, while genes that are methylated on histone H3 lysine 9 or 27 (H3K9 or H3K27) or histone H4 lysine 20 (H4K20) tend to be repressed. It is now clear that E2Fs and pocket proteins mediate the addition or removal of histone tail modifications. Recent studies suggest that methylation of H3K9, H3K27 and H4K20, modifications associated with heterochromatin and chromatin condensation, can be mediated or maintained by the E2F-Rb pathway.
Interestingly, pRB appears to mediate gene silencing by the Polycomb Repressor Complexes, PRC1 and PRC2, often associated with long-term repression [15,16]. Although their mechanism of action is not fully understood, PRC2 acts by marking targets with tri-methyl-H3K27 (H3K27Me3), whereupon PRC1 recognizes the H3K27Me3 mark and establishes a silent state. Precisely how PRC2 is recruited to specific genes in mammals is unknown, since unlike its Drosophila counterpart, mammalian PRC complexes do not contain site-specific DNA-binding activities, and no human Polycomb Response Element has been identified. Recently, Kotake et al. have shown that pRB is required for silencing of the p16INK4a locus by PRC1 components, suggesting that pRB, and possibly E2Fs, could direct the silencing activity of Polycomb complexes to specific genes [17]. This would represent an attractive mechanism for terminal gene silencing, and work from our laboratory (A.B. and B.D.D., unpublished observations) indicates that terminal silencing of cell cycle control genes during myogenic differentiation is accompanied by pRB-dependent H3K27 tri-methylation.
E2F, pRB, and chromatin compaction
Current models posit that the chromatin fiber is further compacted, partly in response to these histone modifications and other cues [18,19]. Here again, mechanisms for locus-specific compaction of chromatin await discovery. Recently, L3MBTL1, a human homolog of the Drosophila tumor suppressor protein (L(3)mbt) found in the dREAM complex, was shown to play a key role in this process. L3MBTL1 can interact with histone H3 and the linker histone H1 bearing specific covalent marks. L3MBTL1 can simultaneously recognize marks on two or more nucleosomes, imbuing it with the ability to compact nucleosomes in vitro, and thus, it may represent the long-sought trigger for compaction. L3MBTL1 also associates with heterochromatin protein HP1γ and pRB [20]. The latter observation is most interesting, as it is therefore possible that E2F-pRB complexes directly recruit chromatin compaction machinery to genes that must be silenced. It will be interesting to determine whether this is a general mechanism through which the E2F-pRB pathway functions to modulate gene expression during the cell cycle and whether other transcriptional repressors share this ability to recruit L3MBTL1. Interestingly, the fly tumor suppressor L(3)mbt was also identified in a genome-wide RNAi screen for genes required for E2F-dependent repression [4]. L(3)mbt was recruited to many target genes in an E2F-dependent, but RBF-independent, manner, suggesting that an E2F-L3MBT chromatin modification and repression pathway may be functionally conserved from flies to humans.
Gene activation by E2F: it’s all about choice
Histone modifications
Timely activation of proliferation-associated genes by E2Fs at the G1-S boundary is also important, yet relatively little is known regarding mechanisms that activate these genes. Recent studies have begun to shed light on the machinery required for E2F-dependent gene activation.
Recruitment of E2F1-3 to promoters was associated with the acquisition of activating marks (acetylation of histones H3 and H4), consistent with their ability to function as activators, and coincident with cell cycle-dependent gene expression [21,22]. Although the relevant machinery has not been fully identified, ectopic expression experiments have implicated p300/CBP and PCAF/GCN5 histone acetyltransferases (HATs), while more recent studies using ChIP have suggested that the Tip60 HAT could account for E2F-mediated acetylation, likely in a manner redundant with p300 and PCAF, [22].
In addition to acetylation of histones H3 and H4, gene activation is also associated with tri-methylation of histone H3 lysine 4 (H3K4me3). It is catalyzed by histone methyltransferases (HMTs) of the mixed lineage leukemia (MLL) and SET families and is thought to be influenced by the degree of acetylation of histone H3 [23]. Once more, a persistent challenge in this field is to discover how gene-specific recruitment of these chromatin modifying enzymes is achieved. One possible direct connection between E2Fs and H3K4 HMTs has been demonstrated, wherein E2F2, E2F4, and E2F6 bind MLL, facilitating H3K4 methylation and activation of E-type cyclins and other cell cycle genes [24]. More recently, multi-protein complexes containing human herpes simplex virus host cell factor-1 (HCF-1) have been shown to be involved in this process [25]. HCF-1 is important for proper cell cycle progression, and its disruption by herpes viral proteins blocks the proliferation of infected cells [26-28]. Interestingly, HCF-1 associates with E2F factors, and the composition of these complexes fluctuates during the cell cycle: in early G1 phase, HCF-1 associates only with E2F4, but E2F1 gradually replaces E2F4 as cells progress toward the G1-S boundary. E2F4 promoter occupancy has been linked to recruitment of a Sin3B co-repressor/HDAC complex, loss of acetylation, and gene repression in quiescent and early G1 phase cells [1,21,29]. Interestingly, the E2F4-containing HCF-1 complex in early G1 phase also contains the related Sin3A co-repressor protein, whereas the G1/S HCF-1 complex, containing E2F1, is associated with Ash2, a component of the MLL HMT activator complex. In line with a model whereby E2F1 recruits such a complex to activate the expression of its target genes in S phase, binding of MLL, Set1 and WDR5, and appearance of H3K4me3 peak at E2F target genes at the G1/S transition, and these events and target gene activation all depend on HCF-1. Therefore this represents another example of a multi-protein complex that undergoes a switch in composition that converts it from a repressor (E2F4/HCF-1/Sin3A) to an activator (E2F1/HCF-1/MLL-Set1). Interestingly, the E2F4/HCF-1 repressor complex does not contain pocket proteins, strengthening the notion that E2F4 regulates the expression of a subset of its target genes independently of this class of proteins [1].
E2F and chromatin remodeling
It is known that the SWI/SNF ATP-dependent chromatin remodeling complexes are implicated in repressing E2F targets during cell cycle arrest [30,31], but new evidence suggests that specific adaptors may control the choice of transcriptional fates. Several versions of this multi-protein complex exist, distinguished by alternative subunit compositions, and they are thought to have different activity. The non-catalytic ARID subunits, ARID1A and ARID1B, may dictate functional differences. Indeed, recent studies suggest that the ARID1A subunit of the SWI/SNF complex is required for cell cycle exit, whereas the ARID1B-containing complex is only necessary for cell cycle reentry [32]. Both proteins interact with repressor E2Fs, but only ARID1B interacts specifically with E2F1, an activator E2F. Interestingly, ARID1B is essential for the recruitment of both the SWI/SNF complex and E2F1 to E2F target genes in proliferating cells, while ARID1A recruitment was associated with repression of cell cycle-specific genes during quiescence. Interestingly, ARID1B specifically associates with a HAT activity, while ARID1A associates exclusively with HDAC1 and HDAC2, presumably through Sin3 recruitment. These studies suggest that E2F-mediated cell cycle-dependent gene repression and activation require histone modification, in particular acetylation, which is directly coordinated with chromatin remodeling through the actions of these two distinct SWI/SNF complexes. It remains to be seen whether a subunit-dependent switch in the activity of SWI/SNF is a general mechanism for E2F target gene activation or if it is restricted to certain promoters and cell types.
Conclusions and perspective
The last two decades of research on E2F have considered mostly “static” behaviors. It is now clear that the E2F and pRB families form scaffolds that recruit additional proteins with the ability to either activate or repress genes and thereby promote or suppress proliferation. As such, the E2F-pRB complex serves as a potent recruiter of enzymes that acetylate, methylate, remodel, and ultimately compact chromatin, reinforcing our understanding of its well-recognized role in human cancer. It will be essential to understand the full range of stimulatory and inhibitory E2F-mediated controls, the extent to which the diverse complexes are required during cell cycle progression, and the context in which the multitude of factors exhibit dynamic rearrangements.
| Of special Interest (one dot) |
| Tyagi et al. 2007. Mol Cell |
| The authors provide a mechanism whereby activator and repressor E2Fs can recruit the HMTs MLL and Set1 to target promoters during specific phases of the cell cycle to activate genes via H3K4 methylation. |
| Osterloh et al. 2007, EMBO J. |
| This represents the first characterization of a human complex similar to the drosophila dREAM complex. It was shown that a LIN-9 containing complex associates with E2F4-p130 in G0, but with B-MYB in S phase. |
| Litovchick et al., 2007, Mol Cell |
| The authors use genomic and proteomic approaches to identify the human DREAM complex, and they provide evidence suggesting that this multi-protein complex can repress E2F4-p130 target genes in quiescent cells. |
| Lu et al., 2007, PNAS USA |
| This study was based on a genome-wide RNAi screen in Drosophila to identify repressors of E2F-mediated gene activation. It led to the identification of L3mbt and Domino as important repressors. Intriguingly, the effect of l3mbt is postulated to be RBF-independent, although Trojer et al. suggest that it is targeted through pRB in human cells. |
| Of exceptional Interest (two dots) |
| Nagl et al. 2007 |
| The authors elegantly demonstrate that SWI/SNF complexes of varying composition associate with distinct E2F family members and regulate either cell cycle exit or proliferation. |
| Trojer et al. 2007 |
| This work presents biochemical evidence that L3MBTL1 has the capacity to impose higher-order chromatin compaction. The authors also demonstrate that L3MBTL1 and pRB participate together in a common complex, suggesting that this could represent a mechanism to impose chromatin condensation at E2F target genes. |
| Takeda et al., 2006 |
| Demonstration that Taspase1-mediated activation of MLL and MLL2 through proteolytic cleavage of these histone-methyl-transferases is required for proper E2F target gene expression and that MLL/MLL2 directly associate with various E2F family members to catalyze the tri-methylation of H3K4 at these loci. |
Acknowledgments
We apologize to the many colleagues whose work could not be cited owing to space constraints. Work in BD’s laboratory was supported by NIH grant CA77245-06.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Balciunaite E, Spektor A, Lents NH, Cam H, Te Riele H, Scime A, Rudnicki MA, Young R, Dynlacht BD. Pocket protein complexes are recruited to distinct targets in quiescent and proliferating cells. Mol Cell Biol. 2005;25:8166–8178. doi: 10.1128/MCB.25.18.8166-8178.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cam H, Balciunaite E, Blais A, Spektor A, Scarpulla RC, Young R, Kluger Y, Dynlacht BD. A common set of gene regulatory networks links metabolism and growth inhibition. Mol Cell. 2004;16:399–411. doi: 10.1016/j.molcel.2004.09.037. [DOI] [PubMed] [Google Scholar]
- 3.Bieda M, Xu X, Singer MA, Green R, Farnham PJ. Unbiased location analysis of E2F1-binding sites suggests a widespread role for E2F1 in the human genome. Genome Res. 2006;16:595–605. doi: 10.1101/gr.4887606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lu J, Ruhf ML, Perrimon N, Leder P. A genome-wide RNA interference screen identifies putative chromatin regulators essential for E2F repression. Proc Natl Acad Sci U S A. 2007;104:9381–9386. doi: 10.1073/pnas.0610279104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Dimova DK, Stevaux O, Frolov MV, Dyson NJ. Cell cycle-dependent and cell cycle-independent control of transcription by the Drosophila E2F/RB pathway. Genes Dev. 2003;17:2308–2320. doi: 10.1101/gad.1116703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ouellet J, Roy R. The lin-35/Rb and RNAi pathways cooperate to regulate a key cell cycle transition in C. elegans. BMC Dev Biol. 2007;7:38. doi: 10.1186/1471-213X-7-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Korenjak M, Taylor-Harding B, Binne UK, Satterlee JS, Stevaux O, Aasland R, White-Cooper H, Dyson N, Brehm A. Native E2F/RBF complexes contain Myb-interacting proteins and repress transcription of developmentally controlled E2F target genes. Cell. 2004;119:181–193. doi: 10.1016/j.cell.2004.09.034. [DOI] [PubMed] [Google Scholar]
- 8.Lewis PW, Beall EL, Fleischer TC, Georlette D, Link AJ, Botchan MR. Identification of a Drosophila Myb-E2F2/RBF transcriptional repressor complex. Genes Dev. 2004;18:2929–2940. doi: 10.1101/gad.1255204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Taylor-Harding B, Binne UK, Korenjak M, Brehm A, Dyson NJ. p55, the Drosophila ortholog of RbAp46/RbAp48, is required for the repression of dE2F2/RBF-regulated genes. Mol Cell Biol. 2004;24:9124–9136. doi: 10.1128/MCB.24.20.9124-9136.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bosco G, Du W, Orr-Weaver TL. DNA replication control through interaction of E2F-RB and the origin recognition complex. Nat Cell Biol. 2001;3:289–295. doi: 10.1038/35060086. [DOI] [PubMed] [Google Scholar]
- 11.Beall EL, Manak JR, Zhou S, Bell M, Lipsick JS, Botchan MR. Role for a Drosophila Myb-containing protein complex in site-specific DNA replication. Nature. 2002;420:833–837. doi: 10.1038/nature01228. [DOI] [PubMed] [Google Scholar]
- 12.Osterloh L, von Eyss B, Schmit F, Rein L, Hubner D, Samans B, Hauser S, Gaubatz S. The human synMuv-like protein LIN-9 is required for transcription of G2/M genes and for entry into mitosis. Embo J. 2007;26:144–157. doi: 10.1038/sj.emboj.7601478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Litovchick L, Sadasivam S, Florens L, Zhu X, Swanson SK, Velmurugan S, Chen R, Washburn MP, Liu XS, DeCaprio JA. Evolutionarily conserved multisubunit RBL2/p130 and E2F4 protein complex represses human cell cycle-dependent genes in quiescence. Mol Cell. 2007;26:539–551. doi: 10.1016/j.molcel.2007.04.015. [DOI] [PubMed] [Google Scholar]
- 14.Pilkinton M, Sandoval R, Colamonici OR. Mammalian Mip/LIN-9 interacts with either the p107, p130/E2F4 repressor complex or B-Myb in a cell cycle-phase-dependent context distinct from the Drosophila dREAM complex. Oncogene. 2007 doi: 10.1038/sj.onc.1210562. [DOI] [PubMed] [Google Scholar]
- 15.Ringrose L. Polycomb comes of age: genome-wide profiling of target sites. Curr Opin Cell Biol. 2007;19:290–297. doi: 10.1016/j.ceb.2007.04.010. [DOI] [PubMed] [Google Scholar]
- 16.Schwartz YB, Pirrotta V. Polycomb silencing mechanisms and the management of genomic programmes. Nat Rev Genet. 2007;8:9–22. doi: 10.1038/nrg1981. [DOI] [PubMed] [Google Scholar]
- 17.Kotake Y, Cao R, Viatour P, Sage J, Zhang Y, Xiong Y. pRB family proteins are required for H3K27 trimethylation and Polycomb repression complexes binding to and silencing p16INK4alpha tumor suppressor gene. Genes Dev. 2007;21:49–54. doi: 10.1101/gad.1499407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Luger K, Hansen JC. Nucleosome and chromatin fiber dynamics. Curr Opin Struct Biol. 2005;15:188–196. doi: 10.1016/j.sbi.2005.03.006. [DOI] [PubMed] [Google Scholar]
- 19.Kouzarides T. Chromatin modifications and their function. Cell. 2007;128:693–705. doi: 10.1016/j.cell.2007.02.005. [DOI] [PubMed] [Google Scholar]
- 20.Trojer P, Li G, Sims RJ, 3rd, Vaquero A, Kalakonda N, Boccuni P, Lee D, Erdjument-Bromage H, Tempst P, Nimer SD, et al. L3MBTL1, a histone-methylation-dependent chromatin lock. Cell. 2007;129:915–928. doi: 10.1016/j.cell.2007.03.048. [DOI] [PubMed] [Google Scholar]
- 21.Takahashi Y, Rayman JB, Dynlacht BD. Analysis of promoter binding by the E2F and pRB families in vivo: distinct E2F proteins mediate activation and repression. Genes Dev. 2000;14:804–816. [PMC free article] [PubMed] [Google Scholar]
- 22.Taubert S, Gorrini C, Frank SR, Parisi T, Fuchs M, Chan HM, Livingston DM, Amati B. E2F-dependent histone acetylation and recruitment of the Tip60 acetyltransferase complex to chromatin in late G1. Mol Cell Biol. 2004;24:4546–4556. doi: 10.1128/MCB.24.10.4546-4556.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nightingale KP, Gendreizig S, White DA, Bradbury C, Hollfelder F, Turner BM. Cross-talk between histone modifications in response to histone deacetylase inhibitors: MLL4 links histone H3 acetylation and histone H3K4 methylation. J Biol Chem. 2007;282:4408–4416. doi: 10.1074/jbc.M606773200. [DOI] [PubMed] [Google Scholar]
- 24.Takeda S, Chen DY, Westergard TD, Fisher JK, Rubens JA, Sasagawa S, Kan JT, Korsmeyer SJ, Cheng EH, Hsieh JJ. Proteolysis of MLL family proteins is essential for taspase1-orchestrated cell cycle progression. Genes Dev. 2006;20:2397–2409. doi: 10.1101/gad.1449406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tyagi S, Chabes AL, Wysocka J, Herr W. E2F activation of S phase promoters via association with HCF-1 and the MLL family of histone H3K4 methyltransferases. Mol Cell. 2007;27:107–119. doi: 10.1016/j.molcel.2007.05.030. [DOI] [PubMed] [Google Scholar]
- 26.Goto H, Motomura S, Wilson AC, Freiman RN, Nakabeppu Y, Fukushima K, Fujishima M, Herr W, Nishimoto T. A single-point mutation in HCF causes temperature-sensitive cell-cycle arrest and disrupts VP16 function. Genes Dev. 1997;11:726–737. doi: 10.1101/gad.11.6.726. [DOI] [PubMed] [Google Scholar]
- 27.Julien E, Herr W. Proteolytic processing is necessary to separate and ensure proper cell growth and cytokinesis functions of HCF-1. Embo J. 2003;22:2360–2369. doi: 10.1093/emboj/cdg242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.de Bruyn Kops A, Knipe DM. Formation of DNA replication structures in herpes virus-infected cells requires a viral DNA binding protein. Cell. 1988;55:857–868. doi: 10.1016/0092-8674(88)90141-9. [DOI] [PubMed] [Google Scholar]
- 29.Rayman JB, Takahashi Y, Indjeian VB, Dannenberg JH, Catchpole S, Watson RJ, te Riele H, Dynlacht BD. E2F mediates cell cycle-dependent transcriptional repression in vivo by recruitment of an HDAC1/mSin3B corepressor complex. Genes Dev. 2002;16:933–947. doi: 10.1101/gad.969202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Liu K, Luo Y, Lin FT, Lin WC. TopBP1 recruits Brg1/Brm to repress E2F1-induced apoptosis, a novel pRb-independent and E2F1-specific control for cell survival. Genes Dev. 2004;18:673–686. doi: 10.1101/gad.1180204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hendricks KB, Shanahan F, Lees E. Role for BRG1 in cell cycle control and tumor suppression. Mol Cell Biol. 2004;24:362–376. doi: 10.1128/MCB.24.1.362-376.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nagl NG, Jr, Wang X, Patsialou A, Van Scoy M, Moran E. Distinct mammalian SWI/SNF chromatin remodeling complexes with opposing roles in cell-cycle control. Embo J. 2007;26:752–763. doi: 10.1038/sj.emboj.7601541. [DOI] [PMC free article] [PubMed] [Google Scholar]