

Abstract
Hematopoietic stem cells (HSCs) are unique in their capacity to give rise to all mature cells of the immune system. For years, HSC transplantation has been used for treatment of genetic and neoplastic diseases of the hematopoietic and immune systems. The sourcing of HSCs from human umbilical cord blood has salient advantages over isolation from mobilized peripheral blood. However, poor sample yield has prompted development of methodologies to expand HSCs ex vivo. Cytokines, trophic factors, and small molecules have been variously used to promote survival and proliferation of HSCs in culture, whilst strategies to lower the concentration of inhibitors in the culture media have recently been applied to promote HSC expansion. In this paper, we outline strategies to expand HSCs in vitro, and to improve engraftment and reconstitution of human immune systems in immunocompromised mice. To the extent that these “humanized” mice are representative of the endogenous human immune system, they will be invaluable tools for both basic science and translational medicine.
1. Introduction
Hematopoietic stem cells (HSCs) were the first class of stem cells used for cell-based therapy in humans. Specifically, both autologous and allogeneic HSC transplantation (HSCT) have been practiced for decades to treat a variety of hematologic malignancies and congenital and autoimmune disorders [1, 2]. Fewer than 30% of patients requiring allogeneic HSCT have a histocompatible sibling, and it is exceedingly rare for patients to have an identical twin donor. Infectious complications and acute or chronic graft-versus-host disease (GVHD) remain the major obstacles affecting patient outcome after allogeneic HSCT [3]. Strikingly, GVHD occurs in approximately 20% to 50% of patients who receive stem cells from a human-leukocyte-antigen- (HLA-) identical sibling donor. Chances increase to 50–80% for those who receive stem cells from an HLA-mismatched sibling or even from an HLA-identical unrelated donor, while chronic GVHD occurs in less than 50% of long-term survivors [4]. Interestingly though, patients with acute GVHD have lower incidence of leukemia relapse, presumably owing to concurrent graft-versus-leukemia response [5]. This beneficial effect may not be limited to patients with leukemia, because something similar was observed in lymphoma patients that received allogeneic bone marrow transplantation [6].
Most commonly, HSCs are obtained by apheresis of adult peripheral blood after mobilization of bone marrow HSCs by granulocyte-colony stimulating factor (G-CSF) injections [7]. As an alternative, HSCs can be isolated from fresh or banked umbilical cord blood (CB) [8], which is highly enriched in HSCs compared to peripheral blood. Benefits of cord blood for transplantation include availability of banked samples, absence of risk to the donor, and low risk of transmitting infectious diseases. A specific advantage of cord blood over bone marrow-derived HSCs is reduced incidence of graft failure and acute or chronic GVHD, especially when cryopreserved CB is used for transplantation [9–11].
Despite having higher HSC concentration than peripheral blood, CB samples are insufficient to provide enough CD34+ cells for successful transplantations in adults and therefore require “pooling” samples from multiple donors. In addition to poor yield, shortage of HLA-matched cord blood samples has stimulated the development of methodologies to allow ex vivo HSC expansion. In principle, these methodologies would maintain self-renewal and inhibit differentiation during the course of expansion. The vast majority of strategies to expand HSCs in vitro have focused on regulation of stem cell renewal and survival of HSCs mediated by intrinsic factors (transcription factors and signaling molecules) and environmental cues (cytokines, chemokines, and adhesion molecules) [12].
An alternative application for HSCs has been to generate mice bearing human immune systems—so-called “humanized” mice. This experimental model was developed to address difficulties associated with studying human immune-related diseases in mice (this has been reviewed in [13–16]). Although a fully functional human immune system has not yet been achieved in the mouse, several strategies have been implemented with variable success.
In this review, we consider various methodologies for maintaining HSCs for the purpose of reconstituting mice with human immune systems.
2. Mouse Models of Hematopoietic Stem Cell Engraftment
The development of chimeric mice bearing human immune system components provides a valuable tool to study human immune responses using small animals. In terms of disease biology, humanized mice can be used to study infection with human-specific pathogens, human autoimmune diseases, and human-specific immune responses in many contexts. These unique models can be created by engraftment of immunodeficient mice with human CD34+ HSCs. A crucial step towards the creation of immunodeficient mice that efficiently accept xenografts was the crossing of nonobese diabetic (NOD) and severe combined immunodeficient (SCID) mouse strains [17]. These NOD-SCID mice display T, B, and NK cell immunodeficiency, in addition to being deficient for macrophages and protein complement. These compound immune deficient mice enable increased chimerism upon HSC transplantation compared to SCID mice [15]. However, these animals have poor human T and B cell maturation, which has limited their use in immunology research. Targeting of cytokine receptors with IL-2Rβ monoclonal antibody prior to transplantation of human HSCs has allowed for even greater engraftment efficiency and human T cell development in the NOD-SCID mouse thymus [18]. Concurrently, new strains of mice deficient for the common cytokine receptor γ-chain (Il2rγ) have been generated. These include NODLtSz-SCID Il2rγ null/null (NSG; Il2rγ is completely null), NODShi-SCID Il2rγ −/− (NOG; the Il2rγ chain lacks the intracytoplasmic domain) [19–22], and BALB/c Rag2−/− Il2rγ −/− mice (BRG) [23, 24]. These important immunocompromised mouse strains have become the most common vehicles for reconstitution of the human hematolymphoid system. Engraftment of CD34+ HSCs into these mice leads, under the right conditions, to differentiation and maintenance (for >6 months) of B and T lymphocytes, NK cells, dendritic cells, monocytes, erythrocytes, and platelets [20, 23, 24].
Myriad conditions contribute to the reconstitution success of engrafted immunodeficient mice. For example, the source of the HSCs plays an important role, with CD34+ HSCs derived from fetal liver or CB providing improved immune reconstitution compared to G-CSF-mobilized adult peripheral blood cells [25]. The age of the recipient immune compromised mouse is also critical, with neonatal recipient mice exhibiting enhanced engraftment compared to adults [26]. A third key factor is the genetic background of the recipient mouse strain. For example, NSG and BRG mice are equivalent in terms of generation of human B cells, dendritic cells, and platelets, whereas NSG mice are superior in supporting human T cell development [25, 26].
This salient difference on T cell development is based on a polymorphism in the gene encoding the signal-regulatory protein alpha (SIRPα) receptor [27]. SIRPα is a receptor expressed mainly in macrophages, granulocytes, and dendritic cells, but its ligand, CD47, is almost ubiquitously expressed. SIRPα binds to CD47 and generates an inhibitory signal to macrophages, which prevents phagocytosis of CD47-expressing cells. Mouse SIRPα interacts weakly with human CD47, with the upshot being phagocytosis and therefore rejection of transplanted human cells. However, NOD mice have a polymorphic allele of SIRPα that binds with high affinity to human CD47, preventing human cells from macrophage-mediated phagocytosis and leading to graft tolerance.
Although the presence of human cells can be detected in chimeric mice for 12 months, all hematopoietic subsets begin to decline around 6 months after transplantation [28, 29]. This effect is probably due to the inability of mouse cytokines to react with human receptors, leading to survival signal and trophic factor deprivation in transplanted human cells. One strategy to overcome this is supplementation with human cytokines; the concept is to create a more favorable immunologic environment for human cells within the mouse host. Another approach to transiently increasing hematopoietic cell lineages in humanized mice has been to inject recombinant proteins including interleukin (IL)-15 [30], IL-7 [31], B-cell activating factor [32], or hydrodynamic injection of a plasmid DNA mixture including IL-15 + Flt-3L and Flt-3L + granulocyte monocyte-CSF(GM-CSF) + IL-4 [33]. Human IL-7 has also been expressed in BRG mice by in vivo lentiviral gene delivery, and this led to stable but supraphysiological levels resulting in increased abundance of T cells [34]. Transgenic mice have also been used to stably increase expression of human cytokines. For example, forced expression of stem cell factor (SCF), GM-CSF, and IL-3 on the NOD-SCID mouse background (NS-SGM3) produced robust human hematopoietic reconstitution in blood, spleen, bone marrow, and liver and significantly increased myeloid cell numbers [35, 36]. Similarly, transgenic NSG mice expressing membrane-bound SCF exhibited a high degree of human CD45+ cell chimerism in irradiated [37] and nonirradiated [38] recipient pups.
A more radical strategy has been to engineer a knock-in mouse in which the genes encoding mouse cytokines have been replaced by their human counterparts. Though laborious, this strategy has major advantages including stable expression of physiological levels of cytokines and localization to the right organ(s). Thus far, three mice have been reported, including one that expresses human thrombopoietin (TPO) [39], another expressing human CSF-1 [40], and an animal that expresses both human IL-3 and GM-CSF [41]. The TPO knock-in mice demonstrated improved engraftment of human CD34+ hematopoietic and progenitor cells, especially in bone marrow, and long-term maintenance of chimerism for over 6 months. Interestingly though, generation of the myelomonocytic lineage was particularly favored in these mice compared with lymphoid lineages. Mice expressing human CSF-1 displayed increased frequency and more efficient differentiation of human fetal liver-derived HSCs into monocytes/macrophages in various organs and increased functional properties, such as migration, phagocytosis and activation, and response to LPS. Transplanted IL-3/GM-CSF knock-in mice had no significant improvement in human hematopoietic engraftment, although enhanced reconstitution with human alveolar macrophages was reported [41]. This specific effect on the lung macrophage subset makes IL-3/GM-CSF knock-in mice a unique model to study the involvement of the immune system in human lung pathologies. Yet, iatrogenic events have been reported in some of these mouse strains. For example, the TPO knock-in strain developed thrombocytopenia [39], despite the fact that human TPO can support murine thrombopoiesis. This effect is likely owed to levels of human TPO expressed in this mouse that were ~10-fold lower than the endogenous murine TPO. In knock-in mice expressing IL-3/GM-CSF, nonengrafted mice developed pulmonary alveolar proteinosis caused by the absence of mouse GM-CSF [41]. Figure 1 summarizes various approaches to the design of humanized mice.
Figure 1.
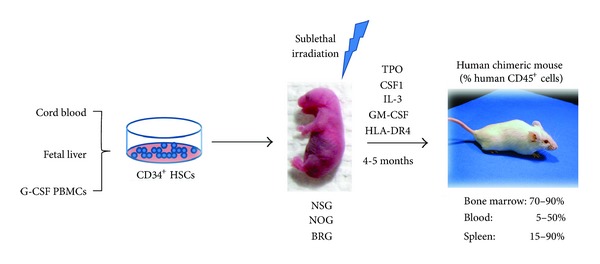
Generation of “humanized” mice. The reconstitution of the hematolymphoid compartment in immunocompromised mice is carried out by human CD34+ HSCs. Human HSCs can be isolated from umbilical cord blood (CB), fetal liver, or adult peripheral blood mononuclear cells (PBMCs) after mobilization of bone marrow HSCs by granulocyte-colony stimulating factor (G-CSF) injections. Although sublethally irradiated newborn NSG, NOG, or BRG immunocompromised mice are the most permissive recipients for engraftment of human CD34+ HSCs, exogenous supply of human cytokines or HLA class II transgenes creates a better environment for cell engraftment and improved development and function of the resultant differentiated immune cell lineages. Abbreviations used: BRG: BALB/c Rag2−/− Il2rγ −/−, CSF1: colony stimulating factor 1, GM-CSF: granulocyte-monocyte colony stimulating factor, HSCs: hematopoietic stem cells, IL-3: interleukin-3, NOG: NODShi-SCID Il2rγ −/−, NSG: NODLtSz-SCID Il2rγ null/null, TPO: thrombopoietin.
3. Human HLA Transgenic Mice
Humanized mice generated by transplantation of human HSCs have demonstrated long-term reconstitution and some degree of maturation of human T cells, evidenced by the presence of CD4/CD8 single-positive T cells in the spleen and peripheral blood [13, 15, 42], CD8+ T cells with effective cytotoxic activity against infection with Epstein-Barr virus (EBV) [24], and development of human B cells that produce antigen-specific IgM upon immunization with exogenous antigens [43, 44]. However, the extent of T and B cell maturation seems incomplete, leading to generation of cells that are not fully functional. For example, CD4+ T cells from humanized NOG mice responded poorly (compared with normal human T cells from healthy donors) to in vitro antigenic stimulation with anti-CD3 and anti-CD28 antibodies [45], and immunization of humanized mice with exogenous antigens was only able to induce a restricted immunoglobulin (Ig) G response [44–46].
HLA molecules are required for development of human T cells, and interactions between human B and T cells are essential to activate the molecular machinery responsible for B cell antibody class switching [47]. Thus, impaired human B and T cell function in humanized mice has been attributed to the absence of HLA in the mouse thymus [48]. In support of this notion, mice transplanted with human fetal liver and thymus under the kidney capsule and injected with HSCs (the bone marrow, liver, thymus (BLT) mouse model) had significantly improved human T and B cell function [49, 50]. Human HLA-DR (MHC class II equivalent) appears to play a more important role in T and B cell development and in T cell positive selection than HLA-A2 (MHC class I analog) in humanized mice. For example, while HLA-A2 transgenic mice elicit a slight improvement in human T cell reconstitution and function of T and B cells, HLA-DR4 or HLA-DR5 transgenic mice had significantly increased human cell reconstitution and better immune responses, including Ig class switching and elevated human IgG responses [51–54]. Furthermore, transgenic expression of human HLA-A2 in humanized NSG mice resulted in improved HLA-A2-restricted CD8+ T cell responses to both EBV and dengue virus infection [55–57]. It is noteworthy that in EBV-infected humanized HLA-A2 transgenic mice, T cell responses against lytic EBV antigens predominated over latent antigens, similar to what is observed in human EBV carriers [57].
4. Ex Vivo Expansion of Hematopoietic Stem Cells
It has become increasingly clear that developing better humanized mouse models will rely, at least in part, on increasing the quality of the HSC input material. Over the past few decades, the study of hematopoietic development and dynamic HSC interactions within the niche has shed light on signaling molecules that play roles in HSC self-renewal and lineage commitment. Rooted in this work, select cytokines and growth factors have been used to maintain and expand HSCs in culture, either alone or in combination [58]. Factors that have been used to promote expansion of human HSCs include Flt3 ligand [59], SCF [60–62], TPO [61, 63], IL-3 [64–66], IL-6 [65, 67], IL-11 [68, 69], and angiopoietin [70]. Although combinations of these cytokines and growth factors have been shown to promote in vitro proliferation of HSCs, the durability of this effect during short-term culture is limited as is the ability to maintain HSCs in an undifferentiated state. One explanation for this is the high sensitivity of HSCs to their microenvironment.
The role of cytokines, including IL-3 and IL-6, in the expansion of HSCs is somewhat controversial. On the one hand, there are reports of stimulatory effects on HSC ex vivo expansion and long-term repopulating capacity [64, 71, 72], whereas others have described an inhibitory effect [73, 74]. One strategy to address this has been to supply these factors by coculturing HSCs with stromal cells, although this technique has met with limited success [75], prompting investigation into other possible factors. Amongst these factors, pleiotrophin, the Notch receptor ligand Delta-1, angiopoietin-like protein 5 (Angptl5), hedgehog (Hh), p38 mitogen-activated protein kinase (MAPK) inhibition, prostaglandin E2 (PGE2), and StemRegenin (SR1) have been shown to stimulate human HSC proliferation and are further discussed below.
The neurite outgrowth factor pleiotrophin has been shown to promote in vitro expansion of both mouse and human HSCs [76]. This effect was observed on mouse bone marrow HSCs, where the protein caused a marked increase in numbers of long-term repopulating HSCs. Furthermore, treatment of human CB CD34+CD38−Lin− cells with pleiotrophin in serum-free media containing TPO, SCF, and Flt3 ligands induced modest ex vivo expansion, but the fraction of CD34+CD38−Lin−cells was significantly higher, indicating a selective effect on differentiation rather than proliferation. Additionally, studies of transplantation using limiting dilution showed increased short- and long-term repopulating capacity of pleiotrophin-treated human CB CD34+CD38−Lin− cells.
It has been proposed that the effect of pleiotrophin could be mediated by activation of phosphoinositide 3-kinase (PI3K)/AKT signaling in HSCs. This effect is most likely refereed by activation of Notch signaling, as antagonism of PI3K or Notch signaling pathways inhibits pleiotrophin-mediated expansion of HSCs in culture [76]. In fact, it is now widely appreciated that Notch signaling plays important roles in the regulation of proliferation and cell fate determination of HSCs. HSCs express Notch receptors, which bind to the transmembrane ligands Jagged-1, Jagged-2, and Delta [77]. Activation of these receptors on murine hematopoietic precursors by the immobilized extracellular domain of Delta1 fused to the Fc domain of human IgG1, [Delta1(ext-IgG)] resulted in marked proliferation of progenitors capable of short-term lymphoid and myeloid repopulation [67, 78]. Similarly, human HSCs cultured in serum-free conditions supplemented with SCF, Flt3-L, TPO, IL-3, and IL-6 were also responsive to activation by Notch ligands, and CD34+ cells underwent ~100-fold expansion in presence of Delta1 (ext-IgG) and exhibited enhanced repopulating ability in an immunodeficient mouse model [79, 80]. However, the effect on Notch signaling appears to be dependent on ligand abundance, since low-density Delta-1 enhances proliferation of CD34+ cells, whereas higher amounts induce apoptosis of CD34+ precursors [81].
One of the most pronounced amplifications of HSCs was achieved by supplementation with angiopoietin-like proteins (Angptls). These proteins were identified in mouse fetal liver CD3+ cells and may play a role in vivo in stimulating mouse fetal liver growth. In particular, Angptl2 and Angptl3 induced up to 30-fold expansion of long-term cultured mouse HSCs as determined by reconstitution analysis [82]. The same group found that another Angptl protein, Angptl5, acted more specifically on human HSCs and promoted ~20-fold net expansion of repopulating human CB HSCs when used in serum-free culture media containing SCF, TPO, fibroblast growth factor-1, and insulin-like growth factor binding protein 2 [83]. Although it appears that Angptls activate a distinct signaling pathway from other growth factors, the mechanism of action remains unclear.
The hedgehog (Hh) signaling pathway has been implicated in primitive and definitive hematopoiesis. In this regard, Bhardwajand and coworkers reported 60 to 80% increased proliferation of human CD34+CD38−Lin− cells at 7 or 12 days in culture following addition of exogenous Hh in presence of SCF, G-CSF, Flt3 ligand, IL-3, and IL-6. Under these conditions, the cells retained their capacity to engraft into immunocompromised NOD-SCID mice [84]. The Hh pathway may also play a role in acute regeneration by inducing cell cycling and expansion of HSCs [85]. There is, however, controversial evidence regarding the role of Hh signaling in hematopoiesis. For example, studies targeting gain or loss of function reported no apparent effect on adult hematopoiesis [86, 87]. This discrepancy may be related to differences in experimental systems (e.g., human, mouse, or zebrafish), approaches (e.g., transgenic models, ES cells, or in vitro culture systems), genetic approaches to removing the Hh activator, smoothened (Smo), in Smo conditional knockout mice (i.e., use of different promoters to induce recombination), source of HSCs (e.g., fetal liver versus adult HSCs), and associated changes in developmental schedules.
Regarding regulation of HSC self-renewal, p38 MAPK has been identified as a key intrinsic negative factor. Activation of p38 MAPK has been associated with induction of HSC senescence under different physiological and pathological conditions [88, 89]. In line with these observations, selective inhibition of p38 MAPK activity with the synthetic agent SB203580 promoted ex vivo expansion of mouse bone marrow and human CB HSCs [90, 91]. Human umbilical CB CD133+ cells expanded in the presence of the drug by about threefold versus vehicle and displayed better engraftment into NOD-SCID mice following transplantation [91]. Improved self-renewal of HSCs following p38 MAPK inhibition has generally been attributed to inhibition of glycogen synthase kinase 3beta (GSK3β) and activation of the Wnt signaling pathway as evidenced by upregulation of the downstream target gene, HOXB4 [92].
Recent investigations seeking to identify modulators of HSC proliferation and homeostasis have revealed new targets. For example, prostaglandin E2 (PGE2) was identified in zebrafish by high-throughput screening of bioactive compounds regulating HSC expansion [93]. Receptors for PGE2 were found in mouse and human HSCs, and short-term ex vivo exposure of HSCs to PGE2 enhanced their homing to bone marrow via the chemokine receptor CXCR4 when transplanted into lethally irradiated hosts. These PGE2-treated cells also demonstrated increased proliferation, resulting in 4-fold increased long-term repopulating cell and competitive repopulating unit frequency, and enhanced survival associated with increased expression of Survivin [94]. Although its mechanism of action is not known, PGE2 has been shown to interact with the Wnt pathway by stabilizing β-catenin [95], highlighting the pivotal and beneficial role of Wnt signaling in HSC biology.
Another high-throughput screen, in this case of a drug library, revealed a purine derivative compound named StemRegenin 1 (SR1) that was able to promote in vitro expansion of mobilized human peripheral blood CD34+ cells in serum-free media containing TPO, IL-6, Flt3 ligand, and SCF [96]. The proliferative effect of SR1 on CD34+ cells did not occur in the absence of cytokines; it was reversible and had an anti-proliferative effect at high concentration, indicating that SR1 enhanced cytokine-mediated signals within a defined dose range. Interestingly, there were species-specific differences on SR1 bioactivity, as the compound did not expand murine HSCs, but potently affected human, monkey, and dog bone marrow-derived CD34+ cells. Furthermore, umbilical CB-derived CD34+ cells cultured for 3 weeks in the presence of SR1 had striking 17-fold expansion that improved early and long-term in vivo repopulation capacity in immunocompromised mice, and these cells retained multilineage potential [96]. The mechanism by which SR1 induces proliferation of HSCs is via direct binding to and antagonism of the aryl hydrocarbon receptor.
Perhaps the most important among intrinsic factors are transcription factors, such as the members of the homeobox (HOX) gene family that have emerged as important regulators of hematopoietic cell proliferation and differentiation. In particular, HOXB4 and HOXA4 are potent ex vivo inductors of HSC expansion, as revealed by overexpression of these transcription factors in murine HSC culture experiments [97–99]. Human HSCs also appear to be responsive to HOX transcription factors. When cultured on stromal cells genetically engineered to secrete HOXB4, human long-term culture-initiating cells and NOD-SCID mouse repopulating cells expanded by more than 20- and 2.5-fold, respectively [100]. Likewise, HOXB4 overexpressing HSCs cells also displayed increased proliferation in culture [101]. In another report, Aurvray and coworkers showed that, in cocultures of HOXC4-producing stromal cells with human CD34+ HSCs, the HOXC4 homeoprotein expanded immature HSCs by 3 to 6 times in in vitro cloning assays and significantly improved in vivo engraftment in immunocompromised mice. Comparative transcriptome analyses of CD34+ cells subjected to HOXB4 or HOXC4 revealed that both homeoproteins regulated the same set of genes, indicating similar downstream effectors [102].
Wnt signaling is another pathway involved in development and function of HSCs. Forced expression of β-catenin enhanced ex vivo proliferation of murine HSCs by increasing HOXB4 and Notch-1 expression [103], and Wnt-3a induced self-renewal of mouse HSCs [104]. There is, however, controversial evidence from experiments based on constitutive activation of canonical Wnt signaling or β-catenin [105, 106]. Furthermore, no impairment of HSC function (e.g., self-renewal or reconstitution) was observed by inactivation of β- and γ-catenin [107, 108]. In experiments where the Wnt signaling pathway was modified by administration of GSK3β inhibitors or overexpression of Wnt5a, long-term repopulation was increased in mice transplanted with murine or human HSCs, but in vitro proliferation went unchanged [109, 110]. This controversy might be explained by differences in experimental systems or functional redundancy.
5. Inhibitory Signals and Control of Hematopoietic Stem Cell Expansion
Most of the strategies designed to expand HSCs in vitro have focused on identifying molecules that promote self-renewal of the stem cell population. Comparably less attention has been directed toward inhibitory signals generated by the differentiated progeny and accumulated during the course of culture. Many of these factors, including transforming growth factor-β (TGF-β), tumor necrosis factor-alpha [111–113], and chemokines such as CCL2, CCL3, CCL4, and CXCL10 [114–116] have been reported to negatively impact the expansion of human hematopoietic stem and progenitor cells. Most of these inhibitory cytokines and chemokines are produced by monocytes and interact in an antagonistic manner with stimulatory factors from megakaryocyte origin (epidermal growth factor, platelet-derived growth factor subunit B, vascular endothelial growth factor, and serotonin) to modulate progenitor expansion [116]. In addition, highly purified bone marrow-derived CD34+ cells also secrete detectable amounts of growth factors, cytokines, and chemokines, which could affect their proliferation in an autocrine or paracrine manner by exerting either stimulatory (kit ligand, Flt3 ligand, and thrombopoietin) or inhibitory influences (TGF-β1, TGF-β2, and platelet factor 4) [117]. Various strategies for maintaining HSCs in culture are outlined in Figure 2.
Figure 2.
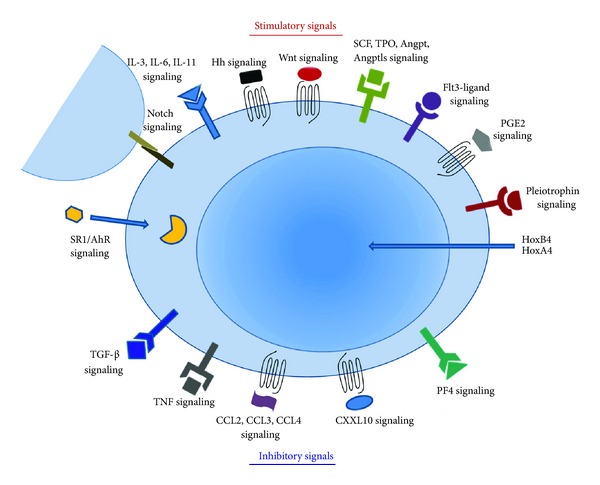
Diagram showing input from multiple signals that affect expansion of HSCs. Proliferation and differentiation of HSCs are regulated by the interplay of stimulatory and inhibitory modulators released by neighboring cells that inhabit the HSC niche within the bone marrow and other cell types within the hematolymphoid compartment. Synthetic molecules utilized to promote ex vivo expansion of HSCs are also depicted. Addition of stimulatory molecules combined with reduction in abundance of inhibitory factors represents a promising strategy to induce in vitro proliferation while maintaining HSCs in an undifferentiated state. Abbreviations used: AhR: aryl hydrocarbon receptor, Angpt: angiopoietin, Angptls: angiopoietin-like proteins, CCL: chemokine ligand, CXCL10: chemokine ligand 10, Hh: hedgehog, Hox: homeobox, IL: interleukin, PF4: platelet factor 4, PGE2: prostaglandin E2, SCF: stem cell factor, SR1: StemRegenin, TGF-β: transforming growth factor beta, TNF: tumor necrosis factor, TPO: thrombopoietin.
Recently, Csaszar and colleagues described an integrated computational and experimental strategy to enable reduction of inhibitory signals in HSC culture media [118]. Based on the effect that feedback signaling from differentiated cells has on stem and progenitor cell expansion, the authors developed a fed-batch media dilution system, consisting of an input stream that results in a continuous increase in culture volume and, consequently, dilution of inhibitory signals. When compared to other media change protocols, including full media change every four days or every day, half media change twice a day, or continuous perfusion, the fed-batch system achieved the most effective enhancement in stem and progenitor expansion. Specifically, it yielded an 11-fold increase in HSCs from human cord blood after 12 days culture, and these cells demonstrated self-renewing, multilineage repopulating ability [118].
Several new factors and molecules have been utilized that, when combined with the most commonly used cocktail of cytokines, and have been successful to varying degrees at improving in vitro HSC expansion. However, at present, combination of these new molecules aiming to optimize culture conditions has not yet been reported. Thus, whether these new factors will have additive or even synergistic effects remains to be proven. Considering the potential for in vitro expansion and long-term engraftment in immunocompromised mice, the most effective factors for expansion of human HSCs to date are SR1 and Angptl5. They have shown similar ability to induce proliferation in culture while preventing differentiation and long-term engraftment in immunocompromised mice.
6. Concluding Remarks
Efficient ex vivo expansion of HSCs still remains an elusive goal. The limited capacity of HSCs to self-renew in culture and their propensity to differentiate despite addition of cytokines and trophic factors represent significant hurdles that limit expansion and long-term engraftment potential of these cells. Nonetheless, significant advances have been made in this area. For example, the identification of new molecules that promote HSC proliferation and maintenance in culture and strategies designed to reduce the levels of inhibitors in the media represent important advances in this area.
With the creation of more sophisticated immunodeficient mice that exhibit improved reconstitution of the human hematolymphoid system, the application of HSCs to study human immune diseases in small animals has moved forward at a rapid pace. It deserves mentioning, however, that these models have limitations, such as the relatively short-term maintenance of chimerism and the poor reconstitution and function of T, B, and natural killer lymphocytes. The exogenous administration or endogenous expression of human growth factors or cytokines in these mice has improved both maintenance and reconstitution and holds future promise for the optimization of humanized mice.
Acknowledgment
This work was made possible by a Grant from the California Institute for Regenerative Medicine to T. Town (RM1-01735).
References
- 1.Appelbaum FR. Hematopoietic-cell transplantation at 50. The New England Journal of Medicine. 2007;357(15):1472–1475. doi: 10.1056/NEJMp078166. [DOI] [PubMed] [Google Scholar]
- 2.Preti RA, Lazarus HM, Winter J, et al. Tumor cell depletion of peripheral blood progenitor cells using positive and positive/negative selection in metastatic breast cancer. Cytotherapy. 2001;3(2):85–95. doi: 10.1080/14653240152584578. [DOI] [PubMed] [Google Scholar]
- 3.Tabbara IA, Zimmerman K, Morgan C, Nahleh Z. Allogeneic hematopoietic stem cell transplantation: complications and results. Archives of Internal Medicine. 2002;162(14):1558–1566. doi: 10.1001/archinte.162.14.1558. [DOI] [PubMed] [Google Scholar]
- 4.Ferrara JLM, Deeg HJ. Graft-versus-host disease. The New England Journal of Medicine. 1991;324(10):667–674. doi: 10.1056/NEJM199103073241005. [DOI] [PubMed] [Google Scholar]
- 5.Horowitz MM, Gale RP, Sondel PM, et al. Graft-versus-leukemia reactions after bone marrow transplantation. Blood. 1990;75(3):555–562. [PubMed] [Google Scholar]
- 6.Jones RJ, Ambinder RF, Piantadosi S, Santos GW. Evidence of a graft-versus-lymphoma effect associated with allogeneic bone marrow transplantation. Blood. 1991;77(3):649–653. [PubMed] [Google Scholar]
- 7.To LB, Haylock DN, Simmons PJ, Juttner CA. The biology and clinical uses of blood stem cells. Blood. 1997;89(7):2233–2258. [PubMed] [Google Scholar]
- 8.Gluckman E. Milestones in umbilical cord blood transplantation. Blood Reviews. 2011;25(6):255–259. doi: 10.1016/j.blre.2011.06.003. [DOI] [PubMed] [Google Scholar]
- 9.Rocha V, Wagner JE, Sobocinski KA, et al. Graft-versus-host disease in children who have received a cord blood or bone marrow transplant from an HLA-identical sibling. Eurocord and International Bone Marrow Transplant Registry Working Committee on Alternative Donor and Stem Cell Sources. The New England Journal of Medicine. 2000;342(25):1846–1854. doi: 10.1056/NEJM200006223422501. [DOI] [PubMed] [Google Scholar]
- 10.Wagner JE, Barker JN, DeFor TE, et al. Transplantation of unrelated donor umbilical cord blood in 102 patients with malignant and nonmalignant diseases: influence of CD34 cell dose and HLA disparity on treatment-related mortality and survival. Blood. 2002;100(5):1611–1618. doi: 10.1182/blood-2002-01-0294. [DOI] [PubMed] [Google Scholar]
- 11.Wagner JE, Rosenthal J, Sweetman R, et al. Successful transplantation of HLA-matched and HLA-mismatched umbilical cord blood from unrelated donors: analysis of engraftment and acute graft- versus-host disease. Blood. 1996;88(3):795–802. [PubMed] [Google Scholar]
- 12.Wilson A, Trumpp A. Bone-marrow haematopoietic-stem-cell niches. Nature Reviews Immunology. 2006;6(2):93–106. doi: 10.1038/nri1779. [DOI] [PubMed] [Google Scholar]
- 13.Ito R, Takahashi T, Katano I, Ito M. Current advances in humanized mouse models. Cellular & Molecular Immunology. 2012;9(3):208–214. doi: 10.1038/cmi.2012.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shultz LD, Brehm MA, Garcia-Martinez JV, Greiner DL. Humanized mice for immune system investigation: progress, promise and challenges. Nature Reviews Immunology. 2012;12(11):786–798. doi: 10.1038/nri3311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Shultz LD, Ishikawa F, Greiner DL. Humanized mice in translational biomedical research. Nature Reviews Immunology. 2007;7(2):118–130. doi: 10.1038/nri2017. [DOI] [PubMed] [Google Scholar]
- 16.Willinger T, Rongvaux A, Strowig T, Manz MG, Flavell RA. Improving human hemato-lymphoid-system mice by cytokine knock-in gene replacement. Trends in Immunology. 2011;32(7):321–327. doi: 10.1016/j.it.2011.04.005. [DOI] [PubMed] [Google Scholar]
- 17.Shultz LD, Schweitzer PA, Christianson SW, et al. Multiple defects in innate and adaptive immunologic function in NOD/LtSz- scid mice. Journal of Immunology. 1995;154(1):180–191. [PubMed] [Google Scholar]
- 18.Kerre TCC, De Smet G, De Smedt M, et al. Adapted NOD/SCID model supports development of phenotypically and functionally mature T cells from human umbilical cord blood CD34+ cells. Blood. 2002;99(5):1620–1626. doi: 10.1182/blood.v99.5.1620. [DOI] [PubMed] [Google Scholar]
- 19.Hiramatsu H, Nishikomori R, Heike T, et al. Complete reconstitution of human lymphocytes from cord blood CD34+ cells using the NOD/SCID/γcnull mice model. Blood. 2003;102(3):873–880. doi: 10.1182/blood-2002-09-2755. [DOI] [PubMed] [Google Scholar]
- 20.Ishikawa F, Yasukawa M, Lyons B, et al. Development of functional human blood and immune systems in NOD/SCID/IL2 receptor γ chainnull mice. Blood. 2005;106(5):1565–1573. doi: 10.1182/blood-2005-02-0516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ito M, Hiramatsu H, Kobayashi K, et al. NOD/SCID/γcnull mouse: an excellent recipient mouse model for engraftment of human cells. Blood. 2002;100(9):3175–3182. doi: 10.1182/blood-2001-12-0207. [DOI] [PubMed] [Google Scholar]
- 22.Shultz LD, Lyons BL, Burzenski LM, et al. Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2Rγnull mice engrafted with mobilized human hemopoietic stem cells. Journal of Immunology. 2005;174(10):6477–6489. doi: 10.4049/jimmunol.174.10.6477. [DOI] [PubMed] [Google Scholar]
- 23.Gimeno R, Weijer K, Voordouw A, et al. Monitoring the effect of gene silencing by RNA interference in human CD34+ cells injected into newborn RAG2-/- γc -/- mice: functional inactivation of p53 in developing T cells. Blood. 2004;104(13):3886–3893. doi: 10.1182/blood-2004-02-0656. [DOI] [PubMed] [Google Scholar]
- 24.Traggiai E, Chicha L, Mazzucchelli L, et al. Development of a human adaptive immune system in cord blood cell-transplanted mice. Science. 2004;304(5667):104–107. doi: 10.1126/science.1093933. [DOI] [PubMed] [Google Scholar]
- 25.Lepus CM, Gibson TF, Gerber SA, et al. Comparison of human fetal liver, umbilical cord blood, and adult blood hematopoietic stem cell engraftment in NOD-scid/γc-/-, Balb/c-Rag1-/-γc-/-, and C.B-17-scid/bg immunodeficient mice. Human Immunology. 2009;70(10):790–802. doi: 10.1016/j.humimm.2009.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Brehm MA, Cuthbert A, Yang C, et al. Parameters for establishing humanized mouse models to study human immunity: analysis of human hematopoietic stem cell engraftment in three immunodeficient strains of mice bearing the IL2rγnull mutation. Clinical Immunology. 2010;135(1):84–98. doi: 10.1016/j.clim.2009.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Takenaka K, Prasolava TK, Wang JCY, et al. Polymorphism in Sirpa modulates engraftment of human hematopoietic stem cells. Nature Immunology. 2007;8(12):1313–1323. doi: 10.1038/ni1527. [DOI] [PubMed] [Google Scholar]
- 28.André MC, Erbacher A, Gille C, et al. Long-term human CD34+ stem cell-engrafted nonobese diabetic/SCID/IL-2Rγnull mice show impaired CD8+ T cell maintenance and a functional arrest of immature NK cells. Journal of Immunology. 2010;185(5):2710–2720. doi: 10.4049/jimmunol.1000583. [DOI] [PubMed] [Google Scholar]
- 29.Watanabe S, Terashima K, Ohta S, et al. Hematopoietic stem cell-engrafted NOD/SCID/IL2Rγnull mice develop human lymphoid systems and induce long-lasting HIV-1 infection with specific humoral immune responses. Blood. 2007;109(1):212–218. doi: 10.1182/blood-2006-04-017681. [DOI] [PubMed] [Google Scholar]
- 30.Huntington ND, Alves NL, Legrand N, et al. IL-15 transpresentation promotes both human T-cell reconstitution and T-cell-dependent antibody responses in vivo. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(15):6217–6222. doi: 10.1073/pnas.1019167108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.van Lent AU, Dontje W, Nagasawa M, et al. IL-7 enhances thymic human T cell development in "human immune system" Rag2-/-IL-2Rγc-/- mice without affecting peripheral T cell homeostasis. Journal of Immunology. 2009;183(12):7645–7655. doi: 10.4049/jimmunol.0902019. [DOI] [PubMed] [Google Scholar]
- 32.Schmidt MR, Appel MC, Giassi LJ, Greiner DL, Shultz LD, Woodland RT. Human BLyS facilitates engraftment of human PBL derived B cells in immunodeficient mice. PLoS One. 2008;3(9, article e3192) doi: 10.1371/journal.pone.0003192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Chen Q, Khoury M, Chen J. Expression of human cytokines dramatically improves reconstitution of specific human-blood lineage cells in humanized mice. Proceedings of the National Academy of Sciences of the United States of America. 2009;106(51):21783–21788. doi: 10.1073/pnas.0912274106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.O’Connell RM, Balazs AB, Rao DS, Kivork C, Yang L, Baltimore D. Lentiviral vector delivery of human interleukin-7 (hiL-7) to human immune system (HIS) mice expands T lymphocyte populations. PLoS One. 2010;5(8, article e12009) doi: 10.1371/journal.pone.0012009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Billerbeck E, Barry WT, Mu K, Dorner M, Rice CM, Ploss A. Development of human CD4+FoxP3+ regulatory T cells in human stem cell factor-, granulocyte-macrophage colony-stimulating factor-, and interleukin-3-expressing NOD-SCID IL2Rγnull humanized mice. Blood. 2011;117(11):3076–3086. doi: 10.1182/blood-2010-08-301507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Nicolini FE, Cashman JD, Hogge DE, Humphries RK, Eaves CJ. NOD/SCID mice engineered to express human IL-3, GM-CSF and steel factor constitutively mobilize engrafted human progenitors and compromise human stem cell regeneration. Leukemia. 2004;18(2):341–347. doi: 10.1038/sj.leu.2403222. [DOI] [PubMed] [Google Scholar]
- 37.Takagi S, Saito Y, Hijikata A, et al. Membrane-bound human SCF/KL promotes in vivo human hematopoietic engraftment and myeloid differentiation. Blood. 2012;119(12):2768–2777. doi: 10.1182/blood-2011-05-353201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Brehm MA, Racki WJ, Leif J, et al. Engraftment of human HSCs in nonirradiated newborn NOD-scid IL2rgamma null mice is enhanced by transgenic expression of membrane-bound human SCF. Blood. 2012;119(12):2778–2788. doi: 10.1182/blood-2011-05-353243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Rongvaux A, Willinger T, Takizawa H, et al. Human thrombopoietin knockin mice efficiently support human hematopoiesis in vivo. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(6):2378–2383. doi: 10.1073/pnas.1019524108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Rathinam C, Poueymirou WT, Rojas J, et al. Efficient differentiation and function of human macrophages in humanized CSF-1 mice. Blood. 2011;118(11):3119–3128. doi: 10.1182/blood-2010-12-326926. [DOI] [PubMed] [Google Scholar]
- 41.Willinger T, Rongvaux A, Takizawa H, et al. Human IL-3/GM-CSF knock-in mice support human alveolar macrophage development and human immune responses in the lung. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(6):2390–2395. doi: 10.1073/pnas.1019682108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Legrand N, Weijer K, Spits H. Experimental models to study development and function of the human immune system in vivo. Journal of Immunology. 2006;176(4):2053–2058. doi: 10.4049/jimmunol.176.4.2053. [DOI] [PubMed] [Google Scholar]
- 43.Becker PD, Legrand N, van Geelen CMM, et al. Generation of human antigen-specific monoclonal IgM antibodies using vaccinated "human immune system" mice. PLoS One. 2010;5(10, article e13137) doi: 10.1371/journal.pone.0013137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kametani Y, Shiina M, Katano I, et al. Development of human-human hybridoma from anti-Her-2 peptide-producing B cells in immunized NOG mouse. Experimental Hematology. 2006;34(9):1240–1248. doi: 10.1016/j.exphem.2006.05.006. [DOI] [PubMed] [Google Scholar]
- 45.Watanabe Y, Takahashi T, Okajima A, et al. The analysis of the functions of human B and T cells in humanized NOD/shi-scid/γcnull (NOG) mice (hu-HSC NOG mice) International Immunology. 2009;21(7):843–858. doi: 10.1093/intimm/dxp050. [DOI] [PubMed] [Google Scholar]
- 46.Baenziger S, Tussiwand R, Schlaepfer E, et al. Disseminated and sustained HIV infection in CD34+ cord blood cell-transplanted Rag2-/-γc-/- mice. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(43):15951–15956. doi: 10.1073/pnas.0604493103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Fairfax KA, Kallies A, Nutt SL, Tarlinton DM. Plasma cell development: from B-cell subsets to long-term survival niches. Seminars in Immunology. 2008;20(1):49–58. doi: 10.1016/j.smim.2007.12.002. [DOI] [PubMed] [Google Scholar]
- 48.Takahama Y, Nitta T, Mat Ripen A, Nitta S, Murata S, Tanaka K. Role of thymic cortex-specific self-peptides in positive selection of T cells. Seminars in Immunology. 2010;22(5):287–293. doi: 10.1016/j.smim.2010.04.012. [DOI] [PubMed] [Google Scholar]
- 49.Lan P, Tonomura N, Shimizu A, Wang S, Yang YG. Reconstitution of a functional human immune system in immunodeficient mice through combined human fetal thymus/liver and CD34+ cell transplantation. Blood. 2006;108(2):487–492. doi: 10.1182/blood-2005-11-4388. [DOI] [PubMed] [Google Scholar]
- 50.Rajesh D, Zhou Y, Jankowska-Gan E, et al. Th1 and Th17 immunocompetence in humanized NOD/SCID/IL2rgammanull mice. Human Immunology. 2010;71(6):551–559. doi: 10.1016/j.humimm.2010.02.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Covassin L, Laning J, Abdi R, et al. Human peripheral blood CD4 T cell-engrafted non-obese diabetic-scid IL2rgamma(null) H2-Ab1 (tm1Gru) Tg (human leucocyte antigen D-related 4) mice: a mouse model of human allogeneic graft-versus-host disease. Clinical & Experimental Immunology. 2011;166(2):269–280. doi: 10.1111/j.1365-2249.2011.04462.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Danner R, Chaudhari SN, Rosenberger J, et al. Expression of HLA class II molecules in humanized NOD.Rag1KO.IL2RgcKO mice is critical for development and function of human T and B cells. PLoS One. 2011;6(5, article e19826) doi: 10.1371/journal.pone.0019826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sato Y, Nagata S, Takiguchi M. Effective elicitation of human effector CD8+ T Cells in HLA-B*51:01 transgenic humanized mice after infection with HIV-1. PLoS One. 2012;7(8, article e42776) doi: 10.1371/journal.pone.0042776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Suzuki M, Takahashi T, Katano I, et al. Induction of human humoral immune responses in a novel HLA-DR-expressing transgenic NOD/Shi-scid/gammacnull mouse. International Immunology. 2012;24(4):243–252. doi: 10.1093/intimm/dxs045. [DOI] [PubMed] [Google Scholar]
- 55.Jaiswal S, Pearson T, Friberg H, et al. Dengue virus infection and virus-specific HLA-A2 restricted immune responses in humanized NOD-scid IL2rγnull mice. PLoS One. 2009;4(10, article e7251) doi: 10.1371/journal.pone.0007251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Shultz LD, Saito Y, Najima Y, et al. Generation of functional human T-cell subsets with HLA-restricted immune responses in HLA class I expressing NOD/SCID/IL2rγnull humanized mice. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(29):13022–13027. doi: 10.1073/pnas.1000475107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Strowig T, Gurer C, Ploss A, et al. Priming of protective T cell responses against virus-induced tumors in mice with human immune system components. Journal of Experimental Medicine. 2009;206(6):1423–1434. doi: 10.1084/jem.20081720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Takizawa H, Schanz U, Manz MG. Ex vivo expansion of hematopoietic stem cells: mission accomplished? Swiss Medical Weekly. 2011;141, article w13316 doi: 10.4414/smw.2011.13316. [DOI] [PubMed] [Google Scholar]
- 59.Yonemura Y, Ku H, Lyman SD, Ogawa M. In vitro expansion of hematopoietic progenitors and maintenance of stem cells: comparison between FLT3/FLK-2 ligand and KIT ligand. Blood. 1997;89(6):1915–1921. [PubMed] [Google Scholar]
- 60.Nakauchi H, Sudo K, Ema H. Quantitative assessment of the stem cell self-renewal capacity. Annals of the New York Academy of Sciences. 2001;938:18–25. doi: 10.1111/j.1749-6632.2001.tb03570.x. [DOI] [PubMed] [Google Scholar]
- 61.Yagi M, Ritchie KA, Sitnicka E, Storey C, Roth GJ, Bartelmez S. Sustained ex vivo expansion of hematopoietic stem cells mediated by thrombopoietin. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(14):8126–8131. doi: 10.1073/pnas.96.14.8126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hoffman R, Tong J, Brandt J, et al. The in vitro and in vivo effects of stem cell factor on human hematopoiesis. Stem Cells. 1993;11(supplement 2):76–82. doi: 10.1002/stem.5530110813. [DOI] [PubMed] [Google Scholar]
- 63.Piacibello W, Sanavio F, Garetto L, et al. The role of c-Mpl ligands in the expansion of cord blood hematopoietic progenitors. Stem Cells. 1998;16(supplement 2):243–248. doi: 10.1002/stem.5530160727. [DOI] [PubMed] [Google Scholar]
- 64.Bryder D, Jacobsen SEW. Interleukin-3 supports expansion of long-term multilineage repopulating activity after multiple stem cell divisions in vitro. Blood. 2000;96(5):1748–1755. [PubMed] [Google Scholar]
- 65.Bordeaux-Rego P, Luzo A, Costa FF, Olalla Saad ST, Crosara-Alberto DP. Both Interleukin-3 and Interleukin-6 are necessary for better ex vivo expansion of CD133+ cells from umbilical cord blood. Stem Cells and Development. 2010;19(3):413–422. doi: 10.1089/scd.2009.0098. [DOI] [PubMed] [Google Scholar]
- 66.Roßmanith T, Schröder B, Bug G, et al. Interleukin 3 improves the ex vivo expansion of primitive human cord blood progenitor cells and maintains the engraftment potential of SCID repopulating cells. Stem Cells. 2001;19(4):313–320. doi: 10.1634/stemcells.19-4-313. [DOI] [PubMed] [Google Scholar]
- 67.Varnum-Finney B, Brashem-Stein C, Bernstein ID. Combined effects of Notch signaling and cytokines induce a multiple log increase in precursors with lymphoid and myeloid reconstituting ability. Blood. 2003;101(5):1784–1789. doi: 10.1182/blood-2002-06-1862. [DOI] [PubMed] [Google Scholar]
- 68.Lemieux ME, Chappel SM, Miller CL, Eaves CJ. Differential ability of flt3-ligand, interleukin-11, and steel factor to support the generation of B cell progenitors and myeloid cells from primitive murine fetal liver cells. Experimental Hematology. 1997;25(9):951–957. [PubMed] [Google Scholar]
- 69.van de Ven C, Ishizawa L, Law P, Cairo MS. IL-11 in combination with SLF and G-CSF or GM-CSF significantly increases expansion of isolated CD34+ cell population from cord blood vs. adult bone marrow. Experimental Hematology. 1995;23(12):1289–1295. [PubMed] [Google Scholar]
- 70.Nakamura Y, Yahata T, Muguruma Y, et al. Angiopoietin-1 supports induction of hematopoietic activity in human CD34− bone marrow cells. Experimental Hematology. 2007;35(12):1872–1883. doi: 10.1016/j.exphem.2007.08.007. [DOI] [PubMed] [Google Scholar]
- 71.Ottmann OG, Abboud M, Welte K, Souza LM, Pelus LM. Stimulation of human hematopoietic progenitor cell proliferation and differentiation by recombinant human interleukin 3. Comparison and interactions with recombinant human granulocyte-macrophage and granulocyte colony-stimulating factors. Experimental Hematology. 1989;17(2):191–197. [PubMed] [Google Scholar]
- 72.Saeland S, Caux C, Favre C, et al. Effects of recombinant human interleukin-3 on CD34-enriched normal hematopoietic progenitors and on myeloblastic leukemia cells. Blood. 1988;72(5):1580–1588. [PubMed] [Google Scholar]
- 73.Encabo A, Mateu E, Carbonell-Uberos F, Miñana MD. Interleukin-6 precludes the differentiation induced by interleukin-3 on expansion of CD34+ cells from cord blood. Haematologica. 2003;88(4):388–395. [PubMed] [Google Scholar]
- 74.Piacibello W, Gammaitoni L, Bruno S, et al. Negative influence of IL3 on the expansion of human cord blood in vivo long-term repopulating stem cells. Journal of Hematotherapy and Stem Cell Research. 2000;9(6):945–956. doi: 10.1089/152581600750062408. [DOI] [PubMed] [Google Scholar]
- 75.Bennaceur-Griscelli A, Tourino C, Izac B, Vainchenker W, Coulombel L. Murine stromal cells counteract the loss of long-term culture- initiating cell potential induced by cytokines in CD34+CD38low/neg human bone marrow cells. Blood. 1999;94(2):529–538. [PubMed] [Google Scholar]
- 76.Himburg HA, Muramoto GG, Daher P, et al. Pleiotrophin regulates the expansion and regeneration of hematopoietic stem cells. Nature Medicine. 2010;16(4):475–482. doi: 10.1038/nm.2119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Kojika S, Griffin JD. Notch receptors and hematopoiesis. Experimental Hematology. 2001;29(9):1041–1052. doi: 10.1016/s0301-472x(01)00676-2. [DOI] [PubMed] [Google Scholar]
- 78.Dallas MH, Varnum-Finney B, Martin PJ, Bernstein ID. Enhanced T-cell reconstitution by hematopoietic progenitors expanded ex vivo using the Notch ligand Delta1. Blood. 2007;109(8):3579–3587. doi: 10.1182/blood-2006-08-039842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Delaney C, Heimfeld S, Brashem-Stein C, Voorhies H, Manger RL, Bernstein ID. Notch-mediated expansion of human cord blood progenitor cells capable of rapid myeloid reconstitution. Nature Medicine. 2010;16(2):232–236. doi: 10.1038/nm.2080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ohishi K, Varnum-Finney B, Bernstein ID. Delta-1 enhances marrow and thymus repopulating ability of human CD34+CD38− cord blood cells. Journal of Clinical Investigation. 2002;110(8):1165–1174. doi: 10.1172/JCI16167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Delaney C, Varnum-Finney B, Aoyama K, Brashem-Stein C, Bernstein ID. Dose-dependent effects of the Notch ligand Delta1 on ex vivo differentiation and in vivo marrow repopulating ability of cord blood cells. Blood. 2005;106(8):2693–2699. doi: 10.1182/blood-2005-03-1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zhang CC, Kaba M, Ge G, et al. Angiopoietin-like proteins stimulate ex vivo expansion of hematopoietic stem cells. Nature Medicine. 2006;12(2):240–245. doi: 10.1038/nm1342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Zhang CC, Kaba M, Lizuka S, Huynh H, Lodish HF. Angiopoietin-like 5 and IGFBP2 stimulate ex vivo expansion of human cord blood hematopoietic stem cells as assayed by NOD/sCiD transplantation. Blood. 2008;111(7):3415–3423. doi: 10.1182/blood-2007-11-122119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Bhardwaj G, Murdoch B, Wu D, et al. Sonic hedgehog induces the proliferation of primitive human hematopoietic cells via BMP regulation. Nature Immunology. 2001;2(2):172–180. doi: 10.1038/84282. [DOI] [PubMed] [Google Scholar]
- 85.Trowbridge JJ, Scott MP, Bhatia M. Hedgehog modulates cell cycle regulators in stem cells to control hematopoietic regeneration. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(38):14134–14139. doi: 10.1073/pnas.0604568103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Gao J, Graves S, Koch U, et al. Hedgehog signaling is dispensable for adult hematopoietic stem cell function. Cell Stem Cell. 2009;4(6):548–558. doi: 10.1016/j.stem.2009.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Hofmann I, Stover EH, Cullen DE, et al. Hedgehog signaling is dispensable for adult murine hematopoietic stem cell function and hematopoiesis. Cell Stem Cell. 2009;4(6):559–567. doi: 10.1016/j.stem.2009.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Wada T, Penninger JM. Mitogen-activated protein kinases in apoptosis regulation. Oncogene. 2004;23(16):2838–2849. doi: 10.1038/sj.onc.1207556. [DOI] [PubMed] [Google Scholar]
- 89.Zarubin T, Han J. Activation and signaling of the p38 MAP kinase pathway. Cell Research. 2005;15(1):11–18. doi: 10.1038/sj.cr.7290257. [DOI] [PubMed] [Google Scholar]
- 90.Wang Y, Kellner J, Liu L, Zhou D. Inhibition of p38 mitogen-activated protein kinase promotes ex vivo hematopoietic stem cell expansion. Stem Cells and Development. 2011;20(7):1143–1152. doi: 10.1089/scd.2010.0413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Zou J, Zou P, Wang J, et al. Inhibition of p38 MAPK activity promotes ex vivo expansion of human cord blood hematopoietic stem cells. Annals of Hematology. 2012;91(6):813–823. doi: 10.1007/s00277-011-1397-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Bug G, Gül H, Schwarz K, et al. Valproic acid stimulates proliferation and self-renewal of hematopoietic stem cells. Cancer Research. 2005;65(7):2537–2541. doi: 10.1158/0008-5472.CAN-04-3011. [DOI] [PubMed] [Google Scholar]
- 93.North TE, Goessling W, Walkley CR, et al. Prostaglandin E2 regulates vertebrate haematopoietic stem cell homeostasis. Nature. 2007;447(7147):1007–1011. doi: 10.1038/nature05883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Hoggatt J, Singh P, Sampath J, Pelus LM. Prostaglandin E2 enhances hematopoietic stem cell homing, survival, and proliferation. Blood. 2009;113(22):5444–5455. doi: 10.1182/blood-2009-01-201335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Goessling W, North TE, Loewer S, et al. Genetic interaction of PGE2 and Wnt signaling regulates developmental specification of stem cells and regeneration. Cell. 2009;136(6):1136–1147. doi: 10.1016/j.cell.2009.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Boitano AE, Wang J, Romeo R, et al. Aryl hydrocarbon receptor antagonists promote the expansion of human hematopoietic stem cells. Science. 2010;329(5997):1345–1348. doi: 10.1126/science.1191536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Antonchuk J, Sauvageau G, Humphries RK. HOXB4-induced expansion of adult hematopoietic stem cells ex vivo. Cell. 2002;109(1):39–45. doi: 10.1016/s0092-8674(02)00697-9. [DOI] [PubMed] [Google Scholar]
- 98.Fournier M, Lebert-Ghali CÉ, Krosl G, Bijl JJ. HOXA4 induces expansion of hematopoietic stem cells in vitro and confers enhancement of pro-B-cells in vivo. Stem Cells and Development. 2012;21(1):133–142. doi: 10.1089/scd.2011.0259. [DOI] [PubMed] [Google Scholar]
- 99.Miyake N, Brun ACM, Magnusson M, Miyake K, Scadden DT, Karlsson S. HOXB4-induced self-renewal of hematopoietic stem cells is significantly enhanced by p21 deficiency. Stem Cells. 2006;24(3):653–661. doi: 10.1634/stemcells.2005-0328. [DOI] [PubMed] [Google Scholar]
- 100.Amsellem S, Pflumio F, Bardinet D, et al. Ex vivo expansion of human hematopoietic stem cells by direct delivery of the HOXB4 homeoprotein. Nature Medicine. 2003;9(11):1423–1427. doi: 10.1038/nm953. [DOI] [PubMed] [Google Scholar]
- 101.Friel J, Schiedlmeier B, Geldmacher M, Ostertag W. Stromal cells selectively reduce the growth advantage of human committed CD34+ hematopoietic cells ectopically expressing HOXB4. Growth Factors. 2006;24(2):97–105. doi: 10.1080/08977190600581269. [DOI] [PubMed] [Google Scholar]
- 102.Auvray C, Delahaye A, Pflumio F, et al. HOXC4 homeoprotein efficiently expands human hematopoietic stem cells and triggers similar molecular alterations as HOXB4. Haematologica. 2012;97(2):168–178. doi: 10.3324/haematol.2011.051235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Reya T, Duncan AW, Ailles L, et al. A role for Wnt signalling in self-renewal of haematopoietic stem cells. Nature. 2003;423(6938):409–414. doi: 10.1038/nature01593. [DOI] [PubMed] [Google Scholar]
- 104.Willert K, Brown JD, Danenberg E, et al. Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature. 2003;423(6938):448–452. doi: 10.1038/nature01611. [DOI] [PubMed] [Google Scholar]
- 105.Kirstetter P, Anderson K, Porse BT, Jacobsen SEW, Nerlov C. Activation of the canonical Wnt pathway leads to loss of hematopoietic stem cell repopulation and multilineage differentiation block. Nature Immunology. 2006;7(10):1048–1056. doi: 10.1038/ni1381. [DOI] [PubMed] [Google Scholar]
- 106.Scheller M, Huelsken J, Rosenbauer F, et al. Hematopoietic stem cell and multilineage defects generated by constitutive β-catenin activation. Nature Immunology. 2006;7(10):1037–1047. doi: 10.1038/ni1387. [DOI] [PubMed] [Google Scholar]
- 107.Cobas M, Wilson A, Ernst B, et al. β-catenin is dispensable for hematopoiesis and lymphopoiesis. Journal of Experimental Medicine. 2004;199(2):221–229. doi: 10.1084/jem.20031615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Koch U, Wilson A, Cobas M, Kemler R, MacDonald HR, Radtke F. Simultaneous loss of β- and γ-catenin does not perturb hematopoiesis or lymphopoiesis. Blood. 2008;111(1):160–164. doi: 10.1182/blood-2007-07-099754. [DOI] [PubMed] [Google Scholar]
- 109.Murdoch B, Chadwick K, Martin M, et al. Wnt-5A augments repopulating capacity and primitive hematopoietic development of human blood stem cells in vivo. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(6):3422–3427. doi: 10.1073/pnas.0130233100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Trowbridge JJ, Xenocostas A, Moon RT, Bhatia M. Glycogen synthase kinase-3 is an in vivo regulator of hematopoietic stem cell repopulation. Nature Medicine. 2006;12(1):89–98. doi: 10.1038/nm1339. [DOI] [PubMed] [Google Scholar]
- 111.Bonnet D, Lemoine FM, Najman A, Guigon M. Comparison of the inhibitory effect of AcSDKP, TNF-α, TGF-β, and MIP-1α on marrow-purified CD34+ progenitors. Experimental Hematology. 1995;23(6):551–556. [PubMed] [Google Scholar]
- 112.Fortunel NO, Hatzfeld A, Hatzfeld JA. Transforming growth factor-β pleiotropic role in the regulation of hematopoiesis. Blood. 2000;96(6):2022–2036. [PubMed] [Google Scholar]
- 113.Zhang Y, Harada A, Bluethmann H, et al. Tumor necrosis factor (TNF) is a physiologic regulator of hematopoietic progenitor cells: increase of early hematopoietic progenitor cells in TNF receptor p55-deficient mice in vivo and potent inhibition of progenitor cell proliferation by TNFα in vitro. Blood. 1995;86(8):2930–2937. [PubMed] [Google Scholar]
- 114.Broxmeyer HE, Kim CH. Regulation of hematopoiesis in a sea of chemokine family members with a plethora of redundant activities. Experimental Hematology. 1999;27(7):1113–1123. doi: 10.1016/s0301-472x(99)00045-4. [DOI] [PubMed] [Google Scholar]
- 115.Cashman JD, Eaves CJ, Sarris AH, Eaves AC. MCP-1, not MIP-1α, is the endogenous chemokine that cooperates with TGF-β to inhibit the cycling of primitive normal but not leukemic (CML) progenitors in long-term human marrow cultures. Blood. 1998;92(7):2338–2344. [PubMed] [Google Scholar]
- 116.Kirouac DC, Ito C, Csaszar E, et al. Dynamic interaction networks in a hierarchically organized tissue. Molecular Systems Biology. 2010;6, article 417 doi: 10.1038/msb.2010.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Majka M, Janowska-Wieczorek A, Ratajczak J, et al. Numerous growth factors, cytokines, and chemokines are secreted by human CD34+ cells, myeloblasts, erythroblasts, and megakaryoblasts and regulate normal hematopoiesis in an autocrine/paracrine manner. Blood. 2001;97(10):3075–3085. doi: 10.1182/blood.v97.10.3075. [DOI] [PubMed] [Google Scholar]
- 118.Csaszar E, Kirouac DC, Yu M, et al. Rapid expansion of human hematopoietic stem cells by automated control of inhibitory feedback signaling. Cell Stem Cell. 2012;10(2):218–229. doi: 10.1016/j.stem.2012.01.003. [DOI] [PubMed] [Google Scholar]