

Synopsis
The composition of human milk is the biologic norm for infant nutrition. Human milk also contains many hundreds to thousands of distinct bioactive molecules that protect against infection and inflammation and contribute to immune maturation, organ development, and healthy microbial colonization. Some of these molecules, e.g., lactoferrin, are being investigated as novel therapeutic agents. A dynamic, bioactive fluid, human milk changes in composition from colostrum to late lactation, and varies within feeds, diurnally, and between mothers. Feeding infants with expressed human milk is increasing. Pasteurized donor milk is now commonly provided to high risk infants and most mothers in the U.S. express and freeze their milk at some point in lactation for future infant feedings. Many milk proteins are degraded by heat treatment and freeze-thaw cycles may not have the same bioactivity after undergoing these treatments. This article provides an overview of the composition of human milk, sources of its variation, and its clinical relevance.
Keywords: human milk composition, breastfeeding, infant nutrition, pasteurization, bioactive factors
INTRODUCTION
Exclusive human milk feeding for the first 6 months of life, with continued breastfeeding for 1 to 2 years of life or longer, is recognized as the normative standard for infant feeding.1,2 Human milk is uniquely suited to the human infant, both in its nutritional composition and in the non-nutritive bioactive factors that promote survival and healthy development.3 Here, we briefly review the nutritional composition of human milk and provide an overview of its varied bioactive factors, which include cells, anti-infectious and anti-inflammatory agents, growth factors, and prebiotics. Unlike infant formula, which is standardized within a very narrow range of composition, human milk composition is dynamic, and varies within a feeding, diurnally, over lactation, and between mothers and populations. Influences on compositional differences of human milk include maternal and environmental factors and the expression and management of milk (e.g., its storage and pasteurization). Understanding human milk composition provides an important tool for management of infant feeding, particularly of fragile, high risk infants, and for understanding the potential impact of storage and pasteurization on milk components. Further, we note some bioactive components found in human milk that are being developed and tested for potential medical applications as prophylactic or therapeutic agents.
STAGES OF LACTATION
The first fluid produced by mothers after delivery is colostrum, which is distinct in volume, appearance and composition. Colostrum, produced in low quantities in the first few days postpartum, is rich in immunologic components such as secretory IgA, lactoferrin, leukocytes, as well as developmental factors such as epidermal growth factor.4-6 Colostrum also contains relatively low concentrations of lactose, indicating its primary functions to be immunologic and trophic rather than nutritional. Levels of sodium, chloride and magnesium are higher and levels of potassium and calcium are lower in colostrum than later milk.5,6 As tight junction closure occurs in the mammary epithelium, the sodium to potassium ratio declines and lactose concentration increases, indicating secretory activation and the production of transitional milk. The timing of secretory activation (lactogenesis stage II) varies among women, but typically occurs over the first few days postpartum. Delayed onset of lactogenesis is defined as onset >72 hours after delivery and appears to occur more often with preterm delivery and maternal obesity, and may be predicted by markers of metabolic health.7,8 Biochemical markers in early milk for onset of secretory activation include its sodium content, the sodium to potassium ratio, citrate, and lactose.9
Transitional milk shares some of the characteristics of colostrum but represents a period of “ramped up” milk production to support the nutritional and developmental needs of the rapidly growing infant, and typically occurs from 5 days to two weeks postpartum, after which milk is considered largely mature. By four to six weeks postpartum, human milk is considered fully mature. In contrast to the dramatic shift in composition observed in the first month of life, human milk remains relatively similar in composition, although subtle changes in milk composition do occur over the course of lactation.
STUDIES OF HUMAN MILK COMPOSITION
A Medline search using only the phrase “human milk composition” reveals a steady increase in publications since the 1960s, with new components still being identified in human milk, and the functionality of those components under active investigation in many laboratories worldwide. Many studies of human milk composition have been conducted in diverse populations using varied collection, storage and testing methods. The gold standard of milk collection involves sampling of all milk expressed over 24-hours, with collection on multiple occasions from the same individuals over time.10,11 This method, however, can be costly, limiting the number of participants. Alternatively, studies of milk composition can standardize collection at a specific time of day (e.g., morning) by emptying the entire breast, avoiding collection from a breast that was used for nursing within the past 2-3 hours, with collection on multiple occasions from the same individuals over time.12 However, most published studies involve non-standardized collection from donors to milk banks whose milk is collected at different times of day, at different timings within a feed or at diverse stages of lactation. Studies of human milk composition also vary in their attention to storage or treatment conditions, such as the number of freeze-thaw cycles, duration of storage, or pasteurization, which may sometimes explain differing study results.
NUTRITIONAL COMPONENTS OF HUMAN MILK
The nutritional components of human milk derive from three sources: Some of the nutrients of milk originate by synthesis in the lactocyte, some are dietary in origin, and some originate from maternal stores. Overall, the nutritional quality of human milk is highly conserved, but attention to maternal diet is important for some vitamins and the fatty acid composition of human milk (see article in this issue by Valentine and Wagner).
Macronutrients
The macronutrient composition of human milk varies within mothers and across lactation but is remarkably conserved across populations despite variations in maternal nutritional status.13 As shown in Table 1, the mean macronutrient composition of mature, term milk is estimated to be approximately 0.9 to 1.2 g/dL for protein, 3.2 to 3.6 g/dL for fat, and 6.7 to 7.8 g/dL for lactose. Energy estimates range from 65 to 70 kcal/dL, and are highly correlated with the fat content of human milk. Macronutrient composition differs between preterm and term milk, with preterm milk tending to be higher in protein and fat (Table 1). A study in Davis, California examined the association between maternal characteristics and the composition of human milk macronutrients10 and found that after 4 months postpartum, the macronutrient concentrations of human milk are associated with one or more of the following factors: Maternal body weight for height, protein intake, parity, return of menstruation, and nursing frequency. This study also found that mothers who produce higher quantities of milk tend to have lower milk concentrations of fat and protein but higher concentrations of lactose.
Table 1.
Macronutrient (g/dL) and energy (kcal/dL) composition of human milk from specified references
| Author (year), n | Protein | Fat | Lactose | Energy |
|---|---|---|---|---|
| Mean (± 2 SD) | Mean (± 2 SD) | Mean (± 2 SD) | Mean (± 2 SD) | |
|
| ||||
| Term infants, 24-hour collection, mature milk | ||||
|
| ||||
| Nommsen et al (1991), n=58 | 1.2 (0.9, 1.5) | 3.6 (2.2, 5.0) | 7.4 (7.2, 7.7) | 70 (57, 83) |
|
| ||||
| Donor human milk samples | ||||
|
| ||||
| Wojcik et al (2009), n=415 | 1.2 (0.7, 1.7) | 3.2 (1.2, 5.2) | 7.8 (6.0, 9.6) | 65 (43, 87) |
|
| ||||
| Michaelsen et al (1990), n=2553 | a 0.9 (0.6, 1.4) | a 3.6 (1.8, 8.9) | a 7.2 (6.4, 7.6) | a 67 (50,115) |
|
| ||||
| Representative values of mature milk, term infants | ||||
|
| ||||
| Reference standard | 0.9 | 3.5 | 6.7 | 65 to 70 |
|
| ||||
| Preterm, 24-hour collection, first 8 weeks of life | ||||
|
| ||||
| Bauer & Gerss (2011) | ||||
| Born <29 weeks, n=52 | 2.2 (1.3, 3.3) | 4.4 (2.6, 6.2) | 7.6 (6.4, 8.8) | 78 (61, 94) |
| Born 32-33 weeks, n=20 | 1.9 (1.3, 2.5) | 4.8 (2.8, 6.8) | 7.5 (6.5, 8.5) | 77 (64, 89) |
|
| ||||
| Preterm donor milk | ||||
|
| ||||
| Hartmann (2012), n=47 | 1.4 (0.8, 1.9) | 4.2 (2.4, 5.9) | 6.7 (5.5, 7.9) | 70 (53, 87) |
Median (lower bound)
Nommsen et al, 1991: Davis, CA mothers at 3 months postpartum
Wojcik et al, 2009, U.S. milk bank donors
Michaelsen et al, 1990, Danish milk bank donors
Reference standard based on expert review: Pediatric Nutrition Handbook, 6th edition, Table C-1, p. 1201 (Mature Milk)
Bauer & Gerss, 2011, preterm infants 23 to 33 weeks gestational age
Hartmann (2012), Australian donor mothers of preterm infants (see article this issue by Landers and Hartmann)
The proteins of human milk are divided into the whey and casein fractions or complexes, with each comprised by a remarkable array of specific proteins and peptides.14,15 The most abundant proteins are casein, α-lactalbumin, lactoferrin, secretory immunoglobulin IgA, lysozyme, and serum albumin.16,17 Non-protein nitrogen-containing compounds, including urea, uric acid, creatine, creatinine, amino acids, and nucleotides, comprise ~25% of human milk nitrogen. The protein content of milk obtained from mothers who deliver preterm is significantly higher than that of mothers who deliver at term (Table 1). Protein levels decrease in human milk over the first 4 to 6 weeks or more of life regardless of timing of delivery (Figure 1).11 For feeding preterm infants, the lower level of total protein and specific amino acids from donor (typically, term, late lactation) milk alone is limiting, and requires additional supplementation (see article in this issue by Underwood).18 Human milk protein concentration is not affected by maternal diet, but increases with maternal body weight for height, and decreases in mothers producing higher amounts of milk.10
Figure 1.
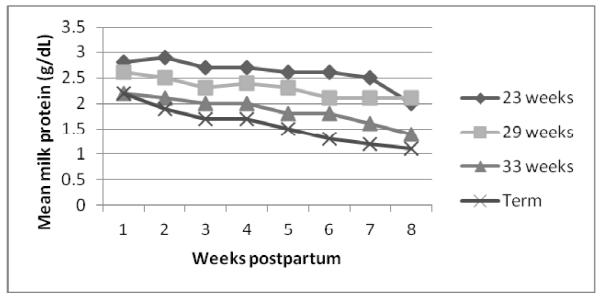
Milk protein concentrations, comparing milk from mothers who delivered preterm and term, by gestational age at delivery and weeks postpartum (adapted from Bauer & Gerss, 2011).
Human milk fat is characterized by high contents of palmitic and oleic acids, the former heavily concentrated in the 2-position and the latter in the 1- and 3-positions of the triglycerides. Fat is the most highly variable macronutrient of milk. Hindmilk, defined as the last milk of a feed, may contain two to three times the concentration of milk fat found in foremilk, defined as the initial milk of a feed.19 A study of milk from 71 mothers over a 24-hour period found that the milk fat content was significantly lower in night and morning feedings compared to afternoon or evening feedings.20 Another study found that ~25% of the variation in lipid concentration between mothers’ milk may be explained by maternal protein intake.10
The fatty acid profile of human milk varies in relation to maternal diet, particularly, in the long chain polyunsaturated fatty acids (LCPUFAs). LCPUFA intake in the Western world is skewed towards the omega-6 fatty acids, with sub-optimal intake of omega-3 fatty acids. The docosahexanoic acid (DHA) composition of human milk is particularly low in North American populations; supplementation should be considered for breastfeeding North American women with DHA limited diets.18,21,22
The principal sugar of human milk is the disaccharide lactose. The concentration of lactose in human milk is the least variable of the macronutrients, but higher concentrations of lactose are found in the milk of mothers producing higher quantities of milk.10 The other significant carbohydrates of human milk are the oligosaccharides, which comprise approximately 1 g/dL in human milk, depending on stage of lactation and maternal genetic factors.23-25 The oligosaccharides are among the non-nutritive bioactive factors discussed below.
The mean values for macronutrients provided here, while valid, belie the observed variability of individual milk samples, particularly in their fat and protein content. Individual donor milk samples from term mothers range at least from 0.6 to 1.4 g/dL for total protein, 1.8 to 8.9 g/dL for fat, 6.4 to 7.6 for lactose, and 50 to 115 kcal/dL for energy.26 Furthermore, the typical composition of preterm milk differs from that of term milk (Table 1). Thus, nutritional management of high risk infants using human milk requires individualized, adaptive or targeted, strategies for fortification, based on measurement of milk composition and growth monitoring.27
Micronutrients
Human milk provides the normative standard for infant nutrition. Nevertheless, many micronutrients vary in human milk depending on maternal diet and body stores (see article in this issue by Valentine and Wagner, 2012), including vitamins A, B1, B2, B6, B12, D, and iodine. The maternal diet is not always optimal, thus, continuing multi-vitamins during lactation is recommended.28,29 Regardless of maternal diet, Vitamin K is extremely low in human milk and thus, the American Academy of Pediatrics recommends an injection of this vitamin to avoid hemorrhagic disease of the newborn.28 Vitamin D also occurs in low quantity in human milk, particularly with low maternal exposure to sunshine, a circumstance now common in populations worldwide.30 While the impact of maternal supplementation with vitamin D on milk composition is under investigation, current pediatric recommendations target postnatal vitamin D supplementation of breastfed infants. Detailed review of the micronutrient composition of human milk is not possible here, but reviews and composition tables are available elsewhere.28,29,31
BIOACTIVE COMPONENTS AND THEIR SOURCES
Bioactive components of food are defined as elements that “affect biological processes or substrates and hence have an impact on body function or condition and ultimately health”.32 Bioactive components in human milk come from a variety of sources; some are produced and secreted by the mammary epithelium, some are produced by cells carried within the milk,33 while others are drawn from maternal serum and carried across the mammary epithelium by receptor-mediated transport. Further, the secretion of the milk fat globule (MFG) into milk by the mammary epithelium carries with it a diverse collection of membrane-bound proteins and lipids into the milk.34 Together these methods produce the variety of bioactive components in human milk. For example, in lactating women, antigen-specific B cells home to the mammary gland, where polymeric immunoglobulin receptors (pIgR) transport sIgA into the lumen of the duct.35 An alternative example is vascular endothelial growth factor (VEGF), which is found at concentrations significantly higher in milk than maternal serum, indicating a mammary gland source.36,37 Understanding the sources of bioactive components of milk also helps to explain the variability in milk concentrations that are observed following maternal use of specific medications (see article in this issue by Rowe, Baker and Hale).
What are the clinical implications of research on human milk bioactive factors? The depth of scientific evidence is such that in patient or public education, it is valid to clarify that human milk is not “merely nutrition.” Rather, human milk contains a variety of factors with medicinal qualities that have a profound role in infant survival and health. Thus, safe donor milk substitutes are needed for infants at medical risk when mother’s own milk is not available. Proteomic analysis has discovered thematic distinctions in the proteins that compose milk at differing stages of lactation, as well as differences between term and preterm milks.14,15 These studies suggest that when donor milk is needed, it should be matched to the developmental stage of the infant whenever feasible, although this is often difficult in practice. Furthermore, recognition of potent, bioactive human milk factors indicates the importance of preserving their biologic activity, to the extent possible, through the process of milk collection, storage, and pasteurization. Finally, recognition of the unique mechanisms by which human milk protects and enhances development provides models for new preventive and therapeutic approaches in medicine.
A complete characterization of bioactive factors of human milk is beyond the scope of this review. Here, we focus on a selected set of bioactive factors that vary between mothers of term and preterm infants, or over the course of lactation, and thus represent responsiveness to the changing needs of the infant (see Table 2). Many of these factors act synergistically, such that consumption of human milk is superior to supplementation with individual factors or their combinations.38
Table 2.
Major Bioactive Factors in Human Milk
| Component | Function | Reference |
|---|---|---|
| Cells | ||
| Macrophages | Protection against infection, T-cell activation |
Jarvinen, 2002, Yagi, 2010, Ichikawa, 2003 |
| Stem cells | Regeneration and repair | Indumathi, 2012 |
| Immunoglobulins | ||
| IgA/sIgA | Pathogen binding inhibition |
Van de Perre, 2003, Cianga, 1999; Brandtzaeg, 2010; Kadaoui, 2007; Corthësy, 2009; Hurley, 2011; Agarwal, 2010; Castellote, 2011 |
| IgG | Anti-microbial, activation of phagocytosis (IgG1, IgG2, IgG3); anti-inflammatory, response to allergens (IgG4) |
Cianga, 1999; Agarwal, 2010 |
| IgM | Agglutination, complement activation | Brandtzaeg, 2010; Van de Perre, 1993; Agarwal, 2010 |
| Cytokines | ||
| IL-6 | Stimulation of the acute phase response, B cell activation, pro-inflammatory |
Ustundag, 2005; Meki, 2003; Mizuno, 2012; Agarwal, 2010; Castellote, 2011 |
| IL-7 | Increased thymic size and output | Aspinall, 2011; Ngom, 2004 |
| IL-8 | Recruitment of neutrophils, pro- inflammatory |
Claud, 2003; Ustundag, 2005; Meki, 2003; Maheshwari, 2002; Maheshwari, 2003; Maheshwari, 2004; Hunt, 2012; Agarwal, 2010; Castellote, 2011; Mehta, 2011 |
| IL-10 | Repressing Th1-type inflammation, induction of antibody production, facilitation of tolerance |
Meki, 2003; Agarwal, 2010; Castellote, 2011; Mehta, 2011 |
| IFNγ | Pro-inflammatory, stimulates Th1 response | Hrdý, 2012; Agarwal, 2010 |
| TGFβ | Anti-inflammatory, stimulation of T cell phenotype switch |
Penttila, 2010; Kalliomäki, 1999; Saito, 1993; Nakamura, 2009; Letterio, 1994; Ando, 2007; Ozawa, 2009; Donnet-Hughes, 2000; Verhasselt, 2008; Verhasselt, 2010; Penttila, 2003; Mosconi, 2010; Okamoto, 2005; Penttila, 2006; Peroni, 2009; McPherson, 2001; Ewaschuk, 2011; Castellote, 2011 |
| TNFα | Stimulates inflammatory immune activation | Rudloff, 1992; Ustundag, 2005; Erbağci, 2005; Meki, 2003; Agarwal, 2010; Castellote, 2011 |
| Chemokines | ||
| G-CSF | Trophic factor in intestines | Gilmore, 1994; Gersting, 2003; Calhoun, 2003; Gersting, 2004; |
| MIF | Macrophage Migratory Inhibitory Factor: Prevents macrophage movement, increases anti-pathogen activity of macrophages |
Magi, 2002; Vigh, 2011 |
| Cytokine Inhibitors | ||
| TNFRI and II | Inhibition of TNFα, anti-inflammatory | Buescher, 1998; Buescher, 1996; Meki, 2003; Castellote, 2011 |
| Growth Factors | ||
| EGF | Stimulation of cell proliferation and maturation |
Patki, 2012; Kobata, 2008; Hirai, 2002; Wagner, 2008; Dvorak, 2003; Dvorak, 2004; Chang, 2002; Khailova, 2009; Coursodon, 2012; Clark, 2004; Castellote, 2011; Untalan, 2009 |
| HB-EGF | Protective against damage from hypoxia and ischemia |
Radulescu, 2011 |
| VEGF | Promotion of angiogenesis and tissue repair | Loui, 2012; Ozgurtas, 2011 |
| NGF | Promotion of neuron growth and maturation |
Rodrigues, 2011; Boesmans 2008; Sánchez 1996; Fichter, 2011; |
| IGF | Stimulation of growth and development, increased RBCs and hemoglobin |
Chellakooty, 2006; Blum, 2002; Burrin 1997; Philipps, 2002; Milsom, 2008; Prosser, 1996; Elmlinger, 2007; Peterson, 2000; Murali, 2005; Corpeleijn, 2008; Baregamian, 2006; Baregamian, 2012; Büyükkayhan, 2003; Philipps, 2000; Kling, 2006; |
| Erythropoietin | Erythropoiesis, intestinal development | Carbonell-Estrany 2000; Juul, 2003; Kling, 2008; Miller-Gilbert, 2001; Pasha, 2008; Soubasi, 1995; Shiou, 2011; Arsenault, 2010; Miller, 2002; Untalan, 2009 |
| Hormones | ||
| Calcitonin | Development of enteric neurons | Struck, 2002; Wookey, 2012 |
| Somatostatin | Regulation of gastric epithelial growth | Chen, 1999; Rao, 1999; Gama, 1996 |
| Anti-microbial | ||
| Lactoferrin | Acute phase protein, chelates iron, anti- bacterial, anti-oxidant |
Adamkin, 2012; Sherman, 2004; Manzoni, 2009; Hirotani, 2008; Buccigrossi, 2007; Velona, 1999 |
|
Lactadherin/
MFG E8 |
Anti-viral, prevents inflammation by enhancing phagocytosis of apoptotic cells |
Stubbs, 1990; Kusunoki, 2012; Aziz, 2011; Shi, 2004; Chogle, 2011; Baghdadi, 2012; Peterson, 1998; Newburg, 1998; Shah, 2012; Miksa, 2006; Komura, 2009; Miksa, 2009; Wu, 2012; Matsuda, 2011; Silvestre, 2005 |
| Metabolic hormones | ||
| Adiponectin | Reduction of infant BMI and weight, anti- inflammatory |
Martin, 2006; Newburg, 2010; Woo, 2009; Woo, 2012; Ley, 2011; Dundar 2010; Ozarda, 2012; Savino, 2008; Weyerman, 2006 |
| Leptin | Regulation of energy conversion and infant BMI, appetite regulation |
Savino, 2008; Savino, 2012a; Savino 2012b; Palou, 2009; Weyermann, 2006 |
| Ghrelin | Regulation of energy conversion and infant BMI |
Savino, 2008; Savino, 2012; Dundar 2010 |
| Oligosaccharides & glycans | ||
| HMOS | Prebiotic, stimulating beneficial colonization and reducing colonization with pathogens; reduced inflammation |
Newburg, 2005; Morrow, 2005; DeLeoz, 2012; Marcoba, 2012; Kunz, 2012; Ruhaak, 2012; Bode, 2012 |
| Gangliosides | Brain development; anti-infectious | Wang B, 2012 |
| Glycosaminoglycans | Anti-infectious | Coppa, 2012; Coppa 2011 |
| Mucins | ||
| MUC1 | Block infection by viruses and bacteria |
Ruvoen-Clouet, 2006; Liu, 2012; Sando, 2009; Saeland, 2009; Yolken, 1992 |
| MUC4 | Block infection by viruses and bacteria | Ruvoen-Clouet, 2006; Liu, 2012; Chaturvedi, 2008 |
GROWTH FACTORS
Human milk contains numerous growth factors that have wide-ranging effects on the intestinal tract, vasculature, nervous system, and endocrine system.
Intestinal maturation, and repair: Epidermal growth factor (EGF)
Found in both amniotic fluid and breast milk,38-40 EGF is critical to the maturation and healing of the intestinal mucosa. EGF is resistant to low pH and digestive enzymes, allowing it to pass through the stomach to the intestine, where it stimulates the enterocyte to increase DNA synthesis, cell division, absorbance of water and glucose, and protein synthesis.41,42 There are multiple protective mechanisms of action for EGF in the infant intestine, including inhibition of programmed cell death, and correcting alterations in intestinal and liver tight junction proteins induced by pro-inflammatory TNF-α.43 Heparin-binding growth factor (HB-EGF) is a member of the EGF family, and the primary growth factor responsible for damage resolution following hypoxia, ischemia-reperfusion injury, hemorrhagic shock/resuscitation injury, and necrotizing enterocolitis.44 EGF is highest in early milk and decreases over lactation.45,46 The average EGF level in colostrum is 2000-fold higher and in mature milk is 100-fold higher than in maternal serum.41 Further, preterm milk contains higher levels of EGF than term milk.45,46
Growth and development of the enteral nervous system: Neuronal growth factors
The immaturity of the newborn intestine extends to the enteral nervous system, which requires brain-derived neurotrophic factor (BDNF) and glial cell-line derived neurotrophic factor (GDNF) for its development.47 BDNF can enhance peristalsis, a function which is frequently impaired in the pre-term gut.48 Rodents who lack GDNF display a profound loss of neurons in the ENS.49 BDNF, GDNF, and a related protein, ciliary neurotophic factor (CNTF), are detected in human milk up to 90 days after birth.50,51 In human cells, breast milk-derived GDNF increases neuron survival and outgrowth.51
Tissue growth: The insulin-like growth factor (IGF) superfamily
IGF-I and IGF-II, as well as IGF binding proteins and IGF-specific proteases, are found in human milk.52-54 Levels are highest in colostrum, and steadily decline over the course of lactation.55,56 There are no significant differences between preterm and term milk, with the exception of IGF binding protein-2, which is higher in preterm milk.52,57 In rodents, administration of human IGF-I during surgical stress/total parental nutrition causes increased tissue growth,58 and attenuates intestinal atrophy,59 though no human trial has been conducted.60 IGF-1 may also play a role in the survival of enterocytes following intestinal damage from oxidative stress.57 Breastfed infants have higher circulating IGF-I in the serum.61 IGF can be taken up in a bioactive form by intestines, and transported into the blood.54,62 The function of absorbed IGF has not been fully detailed, but enteral administration of physiological levels of IGF-I stimulates erythropoiesis and increases hematocrit.63
Regulation of the vascular system: Vascular endothelial growth factor (VEGF)
Angiogenesis is regulated primarily by the relative expression of VEGF and its antagonists. VEGF concentration is highest in colostrum in both preterm and term human milk, with preterm milk containing less VEGF than term milk.64 In retinopathy of prematurity (ROP), it is thought that pulmonary immaturity, supplemental oxygen, and negative regulation of VEGF lead to dysregulated vascularization of the retina,65,66 suggesting a mechanism by which human milk may help reduce the burden of ROP.
Intestinal development and prevention of anemia: Erythropoietin (Epo)
Milk contains significant quantities of Epo, which is the primary hormone responsible for increasing red blood cells (RBC). Blood loss, intestinal pathology, and immaturity of the hematopoietic system all contribute to anemia of prematurity, which profoundly impacts growth and development,67 thus, some suggest that Epo may help prevent anemia of prematurity,68 but administration of Epo has shown mixed results.69 Administration of Epo in conjunction with iron, however, may increase hemoglobin and hematocrit levels.70 A small trial of enteral Epo in preterm infants showed increased serum and reticulocyte levels.71 In addition, Epo is an important trophic factor and tightens intestinal junctions.72 There is some evidence that Epo may help protect against mother-to-child transmission of HIV,73 and may help reduce risk of necrotizing enterocolitis.72,74
Growth-regulating hormones: Calcitonin and somatostatin
Calcitonin and its precursor procalcitonin are present in large quantities in milk.75 Enteric neurons express the calcitonin receptor-immunoreactivity (CTR-ir) from late gestation into infancy.76 Somatostatin is rapidly degraded in the jejunum and not transferred through the intestinal wall, but delivery with milk protects it from degradation and maintains bioactivity within the lumen.77 Somatostatin normally inhibits growth factors, but its role in human milk remains unclear.78
Regulating metabolism and body composition: Adiponectin and other hormones
Adiponectin is a large, multi-functional hormone that actively regulates metabolism and suppresses inflammation. Found in large quantities in human milk, adiponectin can cross the intestinal barrier, and appears to modify infant metabolism.79,80 Levels of adiponectin in milk correlate inversely with infant weight and BMI while exclusively breastfeeding, thus, some have proposed that adiponectin in human milk may contribute to reduced incidence of overweight and obesity in later life, though this remains to be determined.79,81 Other metabolism-regulating hormones found in effective quantities in human milk are leptin, resistin, and ghrelin, which appear to play an important role in regulating energy conversion, body composition, and appetite control.82-85
IMMUNOLOGICAL FACTORS
Feeding human milk protects against infection and inflammation,86 and early milk is enriched in immune factors that help to ensure infant survival.15 The specific protective components of human milk are so numerous and multi-functional, that science is just beginning to understand their functions.
Transfer of living protection and programming: Cells of human milk
Human milk contains a variety of cells, including macrophages, T cells, stem cells, and lymphocytes.87-93 In early lactation, the breastfed infant may consume as many as 1010 maternal leukocytes per day. The relative quantity of these cells differs among mothers and is reported to differ in the milk of infants who develop allergy.87 About 80% of the cells in early milk are breast milk macrophages, which originate as peripheral blood monocytes that exit the bloodstream and migrate into milk through the mammary epithelium. Phagocytosis of human milk components transforms these monocytes into potent breast milk macrophages with unique functional features, including the ability to differentiate into dendritic cells that stimulate infant T-cell activity.89,93 This capability provides broadly powerful protection against pathogens while stimulating development of the infant’s own immune system. In women infected with HIV-1 and HTLV-1, however, the activity of these cells unfortunately enables mother to infant viral transmission (see article by Lawrence in this issue). Stem cells have also been identified in human milk;88,90 their function is under investigation.
Communication between cells: Cytokines and chemokines
Cytokines are multi-functional peptides that act in autocrine/paracrine fashion.33 Chemokines are a special class of chemotactic cytokines that induce movement of other cells. Human milk cytokines can cross the intestinal barrier, where they “communicate” with cells to influence immune activity. While many cytokines and chemokines have multiple functions, milk-borne cytokines may be grouped broadly into those that enhance inflammation or defend against infection, and those that reduce inflammation. A complete review of the cytokines of human milk is available elsewhere,33,94-96
The TGF-β family constitutes the most abundant cytokines of human milk, and consists of three isoforms, of which TGF-β2 predominates.97-99 Milk-borne TGF-β regulates inflammation and wound repair, and helps prevent allergic diseases. TGF-β is converted to its active form by the low pH of the stomach.100 Only 15 minutes after TGF-β1-deficient mouse pups are suckled by heterozygous dams, TGF-β1 is measurable in the pup’s serum and tissue.101 Milk TGF-β has tolerance-enhancing activity in the intestinal tract,100,102-104 a proposed mechanism for its anti-allergenic effects.
Granulocyte-colony stimulating factor (G-CSF), identified in human milk decades ago,105 has beneficial effects on intestinal development and the treatment of sepsis. While milk-borne G-CSF survives transit through the stomach, it is not appreciably absorbed and acts at the intestinal surface ,106,107 where it increases villi, crypt depth, and cell proliferation.108 Other regulatory cytokines found in milk are IL-10 and IL-7;109 milk-borne IL-7 is known to cross the intestinal wall and influence thymic development.110
Pro-inflammatory cytokines TNF-α, IL-6, IL-8, and IFNγ are also found in mother’s milk, generally at lower levels, and decreasing over lactation.4,111 The levels of these cytokines in milk are also associated with the timing of delivery.4,111 Intriguingly, soluble TNF Receptors I and II, which neutralize TNFα, also are found in human milk., 112 thus, most TNFα may be bound and not freely active in milk.113 Both IL-8 and TNFα are modestly increased in mature milk of mothers who had preeclampsia.114 Levels of TNFα correlate positively with other inflammatory cytokines.115 Interleukin-6 (IL-6) is associated with systemic inflammation and fever.115 The role of the inflammatory cytokines found in human milk remains under investigation, but they are known to be engaged in the recruitment of neutrophils, enhance intestinal development, and IL-8 may help protect against TNFα-mediated damage.116,117,118 Higher levels of IL-6 and IL-8 occur in mastitis, with higher cytokine levels confined to only the affected lobes.119,120 Interferon gamma (IFNγ) has also been widely studied due to its pro-inflammatory role; IFNγ enhances the Th1/inflammation response while suppressing the Th2/allergic response.94 The colostrum of allergic mothers contains lower IFNγ but higher Th2 cytokines IL-4 and IL-13 compared to non-allergic mothers.121
Protection from infection: Acquired and innate factors
A recent study in Nepal found a 9-fold increase in risk of diarrhea in children who were not breastfed,122 consistent with previous global studies.123 Infants are born with immature acquired immunity, and rely on maternal antibodies for defense against pathogens.124 Human milk sIgA-antigen complexes are taken up and processed by intestinal dendritic cells, which allows for antigen recognition while maintaining a non-inflammatory environment.125 While sIgA is the predominant antibody of human milk, milk also contains IgM and IgG, the latter becoming more abundant in later lactation.15 A comprehensive review of human milk immunoglobulins is available elsewhere.124,126,127
A set of innate, multi-functional molecules also provide significant protection against infection. These molecules are typically highest in colostrums, and decrease over lactation. Among the most abundant of these is lactoferrin, an iron-binding glycoprotein belonging to the transferrin family,128,129 which is effective against many different bacteria, viruses, and fungi.130-134 Another glycoprotein, lactadherin, was initially identified in the milk fat globule.135 Milk-borne lactadherin survives transit through the stomach,136 and prevents rotaviral infection in the newborn.137 Following infection or damage, lactadherin mediates phagocytic uptake of apoptotic cells138,139 and stimulates a signaling cascade that results in decreased inflammation via blockade of TLR4 and NF-κB signaling.138,140 Lactadherin promotes healing during intestinal inflammation140,141 and a tolerogenic phenotype in intestinal dendritic cells and macrophages,138,142 which is important to maintaining gut health. Another multi-functional protein, bile salt stimulating lipase (BSSL)143 is a highly-glycosylated enzyme that breaks down milk fats, thereby releasing their energy for infant metabolism. Milk-borne BSSL also protects infants from viral infection, including Norwalk and HIV. BSSL binds to dendritic cells, preventing HIV trans-infection of CD4+ T cells.144,145 The bioactive region of BSSL is modified by the glycans present, which change with maternal genetics and over lactation.143,146
The milk fat globule (MFG) contains mucins (MUC1, MUC4, and potentially others) derived from the maternal plasma membrane.147 These mucins are multi-functional, but most importantly, protect infants from infection. For example, MUC1 blocks infection by HIV and rotavirus148,149 and both MUC1 and MUC4 block infection by Salmonella enterica serovar typhimurium and Norwalk virus.146,150,151
Selection for the growth of beneficial organisms: Oligosaccharides
The human milk oligosaccharides (HMOS) range from 3 to 32 sugars in size, and differ in composition from those of any other mammal.23,24 Though non-nutritive to the infant, HMOS constitute a remarkable quantity of human milk, similar to the quantity of total protein. These structures are synthesized glycosyltransferases, enzymes that also synthesize similar structures in other human secretions and on mucosal surfaces. The HMOS are “prebiotic” agents that selectively encourage the growth of beneficial (probiotic) organisms. In addition, the HMOS and their protein conjugates are recognized as pathogen-binding inhibitors that function as soluble “decoy” receptors for pathogens that have an affinity for binding to oligosaccharide receptors expressed on the infant’s intestinal surface. Mothers vary in the specific structures of HMOS in their milk as a result of genetic differences similar to blood group types.23-25 This variation in HMOS composition, unlike blood group types, does not create incompatability, so that all mothers may be considered “universal donors.” Rather, the variation in HMOS composition among mothers is thought to promote human survival as pathogens differ in their affinity for binding to specific oligosaccharides. Protection by some forms of HMOS but not others has been shown in relation to diarrhea caused by specific pathogens23,24 and HIV.152 The apparent differences in lactose and HMOS composition of preterm milk requires further investigation.25,153
It was long believed that human milk was sterile, but it is now recognized that human milk harbors a microbial community, the composition of which appears to change with maternal characteristics and over the course of lactation.154,155 The HMOS influence intestinal colonization and may also influence the bacterial community composition of milk.
IMPACT OF STORAGE AND PASTEURIZATION ON MILK COMPONENTS
Increasingly, feeding human milk occurs apart from feeding at the mother’s breast. Most lactating women in North America now express and store their milk at some time.156 Varying degrees of nutrient loss occur depending on the nutrient and the storage methods. For vitamin C, loss occurs rapidly, even during the process of feeding freshly expressed human milk by bottle.155 For multiple human milk components, however, significant degradation may only occur with long term storage and freeze-thaw cycles, which tend to reduce bactericidal capacity. The Academy of Breastfeeding Medicine has a protocol for home storage of human milk that can be used to guide mothers in these activities to optimize the integrity of expressed and stored milk (see article in this issue by Chantry and Howard, ABM Protocol #8).
Another major movement has been the development of milk sharing via donor human milk banks or alternatively, via internet milk sharing (see article in this issue by Landers and Hartmann, 2012). Due to the known or theoretical risk of transmitting pathogens via human milk, a variety of protocols have been developed for pasteurization of donor milk, including: High temperature short time (HTST) heating (72°C for 15 seconds); Holder pasteurization (62.5°C for 30 minutes, a low temperature long time [LTLT] method); and flash heating (a low-tech HTST method that involves heating a jar of milk in a water bath which is rapidly brought to a rolling boil, then milk is removed and rapidly cooled). Flash heating was developed with the goal of achieving effective pasteurization in a home environment in resource-poor areas of the world to prevent transmission of HIV.
Each pasteurization method has been investigated for its ability to eliminate pathogens while preserving as many bioactive components and nutrients as possible.157-159 Unfortunately, heat treatment of human milk reduces the concentration and functionality of its bioactive components, particularly in protein composition and function. Significant reductions have been demonstrated after pasteurization in sIgA, lysozyme, BSSL, cytokines, lipases, TGF-β, and adiponectin, among other proteins.160-162 Pasteurization damages some proteins more than others,162 but is particularly damaging when paired with the multiple freeze-thaw cycles that can occur with donated milk.163 The degree of impact appears to vary by pasteurization method. Boiling is likely the most damaging, while Holder pasteurization, the method utilized by the milk banks associated with the Human Milk Banking Association of North America, may be more destructive than HTST (see also in this issue, Landers & Hartmann). Though a low-tech method, flash-heat treatment appears to preserve the bacteriostatic activity of human milk.158 Much work remains to be done regarding the bioactivity of human milk components following milk treatment.162
HUMAN MILK COMPONENTS IN RELATION TO INFANT HEALTH
Management or augmentation of human milk factors
Recognition of the dynamic variability of human milk is important to the management of human milk feeding. For example, mothers of term infants are commonly advised to empty an entire breast prior to feeding from the other breast. As hindmilk is more energy dense due to its higher lipid content, this recommendation ensures that infant satiety and energy needs for growth are well met. Hindmilk has been successfully used to improve growth of very premature infants, and is recommended for their nutritional management.164 Furthermore, understanding the dynamic variability of human milk, donor milk banks have established protocols for “pooling” different milk donations to achieve a more uniform milk for distribution.
Some milk factors can be modified through dietary intake or exposures in order to optimize infant growth and health. For example, maternal diet influences the DHA content of milk, which is below recommended levels in many populations. Valentine et al have shown that maternal supplementation with 1 g of preformed DHA daily significantly increases milk DHA levels, which thereby improves the dietary DHA of breastfed infants.164 Another approach to modification of human milk is through maternal immunization, which is being actively pursued as a public health strategy. Trials of maternal immunization have demonstrated significant increases in the level of protective immunoglobulins in milk and dramatically reduced influenza in mothers and infants.165
Potential novel therapeutics based on human milk components
The important maternal act of breastfeeding provides many bioactive components to infants. Some of these components have been proposed as novel medical agents for prevention or treatment of disease, including lactoferrin, lactadherin, epidermal growth factor, erythropoietin, and human milk oligosaccharides. While various human milk components have demonstrated significant biologic activity in vitro or in vivo, only a few have been tested in clinical trials to improve specific health outcomes. One of the most studied of these factors is lactoferrin. Testing lactoferrin as a novel medical agent has been advanced by the use of recombinant lactoferrin or bovine lactoferrin, due to its high homology with human lactoferrin. An Italian study of pre-term infants found that supplementation with bovine lactoferrin, alone or in combination with Lactobacillus casei sps. rhamnosus (LGG), significantly reduced the risk of late-onset sepsis.133,134 A current phase 1/2 trail is evaluating the safety and efficacy of an oral lactoferrin solution to prevent infection in preterm infants.161 Many other individual milk components may be tested in future for specific applications, but human milk remains the unique and potent standard for feeding infants.
CONCLUSION
Human milk is a dynamic, multi-faceted fluid containing nutrients and bioactive factors needed for infant health and development. Its composition varies by stage of lactation and between term and preterm infants. While many studies of human milk composition have been conducted, components of human milk are still being identified. Standardized, multi-population studies of human milk composition are sorely needed to create a rigorous, comprehensive reference inclusive of nutrients and bioactive factors. Nevertheless, knowledge of human milk composition is increasing, leading to greater understanding of the role of human milk in infant health and development.166
Key points.
Human milk composition provides the standard for human infant nutrition, including the bioactive components that safeguard infant growth and development.
The composition of human milk is variable within feeds, diurnally, over lactation, and between mothers and populations. This variability has benefits for infant health and survival, but for high risk infants requiring close nutritional oversight, supplementation, and strategies for managing the variability of human milk feeds are needed.
The composition of human milk can be altered with treatment of expressed milk, including its storage and pasteurization. Thus, attention to management of expressed milk is important.
Acknowledgements
We gratefully acknowledge Donna Wuest for assistance with manuscript preparation. This work was supported in part by grants to ALM from the National Institute of Child Health and Human Development (P01 HD 13021, “The role of human milk in infant nutrition and health”) to study bioactive factors in human milk and from Mead Johnson Nutrition, Inc. to conduct a comprehensive study of human milk factors (“The Global Exploration of Human Milk [GEHM] Study”).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Disclosures: Dr. Morrow has grant funding from Mead Johnson Nutrition, Inc. to study the composition of human milk.
Contributor Information
Olivia Ballard, Center for Interdisciplinary Research in Human Milk and Lactation & Division of Immunobiology, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave., MLC 7009, Cincinnati, OH 45229. Olivia.Ballard@cchmc.org.
Ardythe L. Morrow, Center for Interdisciplinary Research in Human Milk and Lactation, Perinatal Institute, Cincinnati Children’s Hospital Medical Center, 3333 Burnet Ave., MLC 7009, Cincinnati, OH 45229. Ardythe.Morrow@cchmc.org.
REFERENCES
- 1.Breastfeeding and the use of human milk. Pediatrics. 2012;129(3):e827–841. doi: 10.1542/peds.2011-3552. [DOI] [PubMed] [Google Scholar]
- 2.WHO . Infant and young child nutrition. Geneva: 2003. [Google Scholar]
- 3.Oftedal OT. The evolution of milk secretion and its ancient origins. Animal : an international journal of animal bioscience. 2012;6(3):355–368. doi: 10.1017/S1751731111001935. [DOI] [PubMed] [Google Scholar]
- 4.Castellote C, Casillas R, Ramirez-Santana C, Perez-Cano FJ, Castell M, Moretones MG, Lopez-Sabater MC, Franch A. Premature delivery influences the immunological composition of colostrum and transitional and mature human milk. The Journal of nutrition. 2011;141(6):1181–1187. doi: 10.3945/jn.110.133652. [DOI] [PubMed] [Google Scholar]
- 5.Pang WW, Hartmann PE. Initiation of human lactation: secretory differentiation and secretory activation. Journal of mammary gland biology and neoplasia. 2007;12(4):211–221. doi: 10.1007/s10911-007-9054-4. [DOI] [PubMed] [Google Scholar]
- 6.Kulski JK, Hartmann PE. Changes in human milk composition during the initiation of lactation. Aust J Exp Biol Med Sci. 1981;59(1):101–114. doi: 10.1038/icb.1981.6. [DOI] [PubMed] [Google Scholar]
- 7.Henderson JJ, Hartmann PE, Newnham JP, Simmer K. Effect of preterm birth and antenatal corticosteroid treatment on lactogenesis II in women. Pediatrics. 2008;121(1):e92–100. doi: 10.1542/peds.2007-1107. [DOI] [PubMed] [Google Scholar]
- 8.Nommsen-Rivers LA, Dolan LM, Huang B. Timing of stage II lactogenesis is predicted by antenatal metabolic health in a cohort of primiparas. Breastfeed Med. 2012;7(1):43–49. doi: 10.1089/bfm.2011.0007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Cregan MD, De Mello TR, Kershaw D, McDougall K, Hartmann PE. Initiation of lactation in women after preterm delivery. Acta obstetricia et gynecologica Scandinavica. 2002;81(9):870–877. doi: 10.1034/j.1600-0412.2002.810913.x. [DOI] [PubMed] [Google Scholar]
- 10.Nommsen LA, Lovelady CA, Heinig MJ, Lonnerdal B, Dewey KG. Determinants of energy, protein, lipid, and lactose concentrations in human milk during the first 12 mo of lactation: the DARLING Study. The American journal of clinical nutrition. 1991;53(2):457–465. doi: 10.1093/ajcn/53.2.457. [DOI] [PubMed] [Google Scholar]
- 11.Bauer J, Gerss J. Longitudinal analysis of macronutrients and minerals in human milk produced by mothers of preterm infants. Clinical nutrition (Edinburgh, Scotland) 2011;30(2):215–220. doi: 10.1016/j.clnu.2010.08.003. [DOI] [PubMed] [Google Scholar]
- 12.Geraghty SR, Davidson BS, Warner BB, Sapsford AL, Ballard JL, List BA, Akers R, Morrow AL. The development of a research human milk bank. J Hum Lact. 2005;21(1):59–66. doi: 10.1177/0890334404273162. [DOI] [PubMed] [Google Scholar]
- 13.Prentice A. Regional Variations in the Composition of Human Milk. In: Jensen RG, editor. Handbook of Milk Composition. Academic Press, Inc.; San Diego, CA: 1995. p. 919. [Google Scholar]
- 14.Liao Y, Alvarado R, Phinney B, Lonnerdal B. Proteomic characterization of human milk whey proteins during a twelve-month lactation period. J Proteome Res. 2011;10(4):1746–1754. doi: 10.1021/pr101028k. [DOI] [PubMed] [Google Scholar]
- 15.Gao X, McMahon RJ, Woo JG, Davidson BS, Morrow AL, Zhang Q. Temporal changes in milk proteomes reveal developing milk functions. J Proteome Res. 2012;11(7):3897–3907. doi: 10.1021/pr3004002. [DOI] [PubMed] [Google Scholar]
- 16.Lonnerdal B. Human milk proteins: key components for the biological activity of human milk. Advances in experimental medicine and biology. 2004;554:11–25. [PubMed] [Google Scholar]
- 17.Jensen RG. Handbook of Milk Composition. Academic Press, Inc.; San Diego, CA: 1995. [Google Scholar]
- 18.Valentine CJ, Morrow G, Fernandez S, Gulati P, Bartholomew D, Long D, Welty SE, Morrow AL, Rogers LK. Docosahexaenoic Acid and Amino Acid Contents in Pasteurized Donor Milk are Low for Preterm Infants. The Journal of pediatrics. 2010;157(6):906–910. doi: 10.1016/j.jpeds.2010.06.017. [DOI] [PubMed] [Google Scholar]
- 19.Saarela T, Kokkonen J, Koivisto M. Macronutrient and energy contents of human milk fractions during the first six months of lactation. Acta Paediatr. 2005;94(9):1176–1181. doi: 10.1111/j.1651-2227.2005.tb02070.x. [DOI] [PubMed] [Google Scholar]
- 20.Kent JC, Mitoulas LR, Cregan MD, Ramsay DT, Doherty DA, Hartmann PE. Volume and frequency of breastfeedings and fat content of breast milk throughout the day. Pediatrics. 2006;117(3):e387–395. doi: 10.1542/peds.2005-1417. [DOI] [PubMed] [Google Scholar]
- 21.Valentine CJ, Morrow G, Pennell M, Morrow AL, Hodge A, Haban-Bartz A, Collins K, Rogers LK. Randomized Controlled Trial of Docosahexaenoic Acid Supplementation in Midwestern U.S. Human Milk Donors. Breastfeed Med. 2012 doi: 10.1089/bfm.2011.0126. EPub doi:10.1089/bfm.2011.0126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Martin MA, Lassek WD, Gaulin SJ, Evans RW, Woo JG, Geraghty SR, Davidson BS, Morrow AL, Kaplan HS, Gurven MD. Fatty acid composition in the mature milk of Bolivian forager-horticulturalists: controlled comparisons with a US sample. Matern Child Nutr. 2012;8(3):404–418. doi: 10.1111/j.1740-8709.2012.00412.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Newburg DS, Ruiz-Palacios GM, Morrow AL. Human milk glycans protect infants against enteric pathogens. Annual review of nutrition. 2005;25:37–58. doi: 10.1146/annurev.nutr.25.050304.092553. [DOI] [PubMed] [Google Scholar]
- 24.Morrow AL, Ruiz-Palacios GM, Jiang X, Newburg DS. Human-milk glycans that inhibit pathogen binding protect breast-feeding infants against infectious diarrhea. The Journal of nutrition. 2005;135(5):1304–1307. doi: 10.1093/jn/135.5.1304. [DOI] [PubMed] [Google Scholar]
- 25.Gabrielli O, Zampini L, Galeazzi T, Padella L, Santoro L, Peila C, Giuliani F, Bertino E, Fabris C, Coppa GV. Preterm milk oligosaccharides during the first month of lactation. Pediatrics. 2011;128(6):e1520–1531. doi: 10.1542/peds.2011-1206. [DOI] [PubMed] [Google Scholar]
- 26.Michaelsen KF, Skafte L, Badsberg JH, Jorgensen M. Variation in macronutrients in human bank milk: influencing factors and implications for human milk banking. Journal of pediatric gastroenterology and nutrition. 1990;11(2):229–239. doi: 10.1097/00005176-199008000-00013. [DOI] [PubMed] [Google Scholar]
- 27.Arslanoglu S, Moro GE, Ziegler EE, The Wapm Working Group On Nutrition Optimization of human milk fortification for preterm infants: new concepts and recommendations. Journal of perinatal medicine. 2010;38(3):233–238. doi: 10.1515/jpm.2010.073. [DOI] [PubMed] [Google Scholar]
- 28.Greer FR. Do breastfed infants need supplemental vitamins? Pediatric clinics of North America. 2001;48(2):415–423. doi: 10.1016/s0031-3955(08)70034-8. [DOI] [PubMed] [Google Scholar]
- 29.Allen LH. B vitamins in breast milk: relative importance of maternal status and intake, and effects on infant status and function. Adv Nutr. 2012;3(3):362–369. doi: 10.3945/an.111.001172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Dawodu A, Zalla L, Woo JG, Herbers PM, Davidson BS, Heubi JE, Morrow AL. Heightened attention to supplementation is needed to improve the vitamin D status of breastfeeding mothers and infants when sunshine exposure is restricted. Matern Child Nutr. 2012 doi: 10.1111/j.1740-8709.2012.00422.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pediatrics AAo. Pediatric Nutrition Handbook. 6th ed American Academy of Pediatrics; Elk Gove Village, IL: 2009. [Google Scholar]
- 32.Schrezenmeir J, Korhonen H, Williams C, Gill HS, Shah N. Foreword. The British journal of nutrition. 2000;84(S1):1. [Google Scholar]
- 33.Garofalo R. Cytokines in human milk. The Journal of pediatrics. 2010;156(2 Suppl):S36–40. doi: 10.1016/j.jpeds.2009.11.019. [DOI] [PubMed] [Google Scholar]
- 34.Cavaletto M, Giuffrida MG, Conti A. The proteomic approach to analysis of human milk fat globule membrane. Clinica chimica acta; international journal of clinical chemistry. 2004;347(1-2):41–48. doi: 10.1016/j.cccn.2004.04.026. [DOI] [PubMed] [Google Scholar]
- 35.Van de Perre P. Transfer of antibody via mother’s milk. Vaccine. 2003;21(24):3374–3376. doi: 10.1016/s0264-410x(03)00336-0. [DOI] [PubMed] [Google Scholar]
- 36.Kobata R, Tsukahara H, Ohshima Y, Ohta N, Tokuriki S, Tamura S, Mayumi M. High levels of growth factors in human breast milk. Early human development. 2008;84(1):67–69. doi: 10.1016/j.earlhumdev.2007.07.005. [DOI] [PubMed] [Google Scholar]
- 37.Patki S, Patki U, Patil R, Indumathi S, Kaingade P, Bulbule A, Nikam A, Pishte A. Comparison of the levels of the growth factors in umbilical cord serum and human milk and its clinical significance. Cytokine. 2012;59(2):305–308. doi: 10.1016/j.cyto.2012.04.010. [DOI] [PubMed] [Google Scholar]
- 38.Hirai C, Ichiba H, Saito M, Shintaku H, Yamano T, Kusuda S. Trophic effect of multiple growth factors in amniotic fluid or human milk on cultured human fetal small intestinal cells. JPGN. 2002;34:524–528. doi: 10.1097/00005176-200205000-00010. [DOI] [PubMed] [Google Scholar]
- 39.Chailler P, Menard D. Ontogeny of EGF receptors in the human gut. Frontiers in Biosci. 1999;4:87–101. doi: 10.2741/chailler. [DOI] [PubMed] [Google Scholar]
- 40.Wagner CL, Taylor SN, Johnson D. Host factors in amniotic fluid and breast milk that contribute to gut maturation. Clinic. Rev. Allerg. Immunol. 2008;34:191–204. doi: 10.1007/s12016-007-8032-3. [DOI] [PubMed] [Google Scholar]
- 41.Read LC, Upton FM, Francis GL, Wallace JC, Dahlenberg GW, Ballad FJ. Changes in the growth-promoting activity of human milk during lactation. Pediatr. Res. 1984;18(2):133–139. doi: 10.1203/00006450-198402000-00004. [DOI] [PubMed] [Google Scholar]
- 42.Chang C-Y, Chao JC-J. Effect of human milk and epidermal growth factor on growth of human intestinal caco-2 cells. JPGN. 2002;34:394–401. [PubMed] [Google Scholar]
- 43.Khailova L, Dvorak K, Arganbright KM, Williams CS, Halpern MD, Dvorak B. Changes in hepatic cell junctions structure during experimental necrotizing enterocolitis: effect of EGF treatment. Pediatr. Res. 2009;66(2):140–144. doi: 10.1203/PDR.0b013e3181aa3198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Radulescu A, Zhang H-Y, Chen C-L, Chen Y, Zhou Y, Yu X, Otabor I, Olson JK, Besner GE. Heparin-Binding EGF-Like Growth Factor promotes intestinal anastomotic healing. J. Surg. Res. 2011;171:540–550. doi: 10.1016/j.jss.2010.06.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Dvorak B, Fituch CC, Williams CS, Hurst NM, Schanler RJ. Increased epidermal growth factor levels in human milk of mothers with extremely premature infants. Pediatr. Res. 2003;54(1):15–19. doi: 10.1203/01.PDR.0000065729.74325.71. [DOI] [PubMed] [Google Scholar]
- 46.Dvorak B, Fituch CC, Williams CS, Hurst NM, Schanler RJ. Concentrations of epidermal growth factor and Transforming Growth Factor-Alpha in preterm milk. In: al. Pe., editor. Protecting Infants Through Human Milk. Kluwer Academic/Plenum Publishers; 2004. pp. 407–409. [DOI] [PubMed] [Google Scholar]
- 47.Rodrigues D, Li A, Nair D, Blennerhassett M. Glial cell line-derived neurotrophic factor is a key neurotrophin in the postnatal enteric nervous system. Neurogastroenterol. Motil. 2011;23:e44–e56. doi: 10.1111/j.1365-2982.2010.01626.x. [DOI] [PubMed] [Google Scholar]
- 48.Boesmans W, Gomes P, Janssens J, Tack J, Berghe PV. Brain-derived neurotrophic factor amplifies neurotransmitter responses and promotes synaptic communication in the enteric nervous system. Gut. 2008;57:314–322. doi: 10.1136/gut.2007.131839. [DOI] [PubMed] [Google Scholar]
- 49.Sánchez M, Silos-Santiago I, Frisén J, He B, Lira S, Barbacid M. Renal agenesis and the absence of enteric neurons in mice lacking GDNF. Nature. 1996;382(6586):70–73. doi: 10.1038/382070a0. [DOI] [PubMed] [Google Scholar]
- 50.Li R, Xia W, Zhang Z, Wu K. S100B protein, brain-derived neurotrophic factor, and glial cell line-derived neurotrophic factor in human milk. PloS ONE. 2011;6(6) doi: 10.1371/journal.pone.0021663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Fichter M, Klotz M, Hirschberg DL, Waldura B, Schofer O, Ehnert S, Schwarz LK, Ginneken CV, Schafer K-H. Breast milk contains relevant neurotrophic factors and cytokines for enteric nervous system development. Mol. Nutr. Food Res. 2011;55:1592–1596. doi: 10.1002/mnfr.201100124. [DOI] [PubMed] [Google Scholar]
- 52.Blum JW, Baumrucker CR. Colostral and milk insulin-like growth factors and related substances: mammary gland and neonatal (intestinal and systemic) targets. Dom. Anim. Endocrin. 2002;23:101–110. doi: 10.1016/s0739-7240(02)00149-2. [DOI] [PubMed] [Google Scholar]
- 53.Burrin DG. Is milk-borne insulin-like growth factor-I essential for neonatal development? J. Nutr. 1997;127:975S–979S. doi: 10.1093/jn/127.5.975S. [DOI] [PubMed] [Google Scholar]
- 54.Philipps AF, Kling PJ, Grille JG, Dvorak B. Intestinal transport of insulin-like growth factor-I (IGF-I) in the suckling rat. JPGN. 2002;35:539–544. doi: 10.1097/00005176-200210000-00015. [DOI] [PubMed] [Google Scholar]
- 55.Milsom SR, Blum WF, Gunn AJ. Temporal changes in insulin-like growth factors I and II and in insulin-like growth factor binding proteins 1, 2, and 3 in human milk. Horm. Res. 2008;69:307–311. doi: 10.1159/000114863. [DOI] [PubMed] [Google Scholar]
- 56.Prosser CG. Insulin-like growth factors in milk and mammary gland. J. Mammary Gland Biol. & Neoplas. 1996;1(3):297–306. doi: 10.1007/BF02018082. [DOI] [PubMed] [Google Scholar]
- 57.Elmlinger MW, Hochhaus F, Loui A, Frommer KW, Obladen M, Ranke MB. Insulin-like growth factors and binding proteins in early milk from mothers of preterm and term infants. Horm. Res. 2007;68:124–131. doi: 10.1159/000100488. [DOI] [PubMed] [Google Scholar]
- 58.Peterson CA, Gillingham MB, Mohapatra NK, Dahly EM, Adamo ML, Carey HV, Lund PK, Ney DM. Enterotrophic effect of insulin-like growth factor-I but not growth hormone and localized expression of insulin-like growth factor-I, insulin-like growth factor binding protein-3 and -5 mRNAs in jejunum of parenterally fed rats. JPEN. 2000;24(5):288–295. doi: 10.1177/0148607100024005288. [DOI] [PubMed] [Google Scholar]
- 59.Murali SG, Nelson DW, Draxler AK, Liu X, Ney DM. Insulin-like growth factor-I (IGF-I) attenuates jejunal atrophy in association with increased expression of IGF-I binding protein-5 in parenterally fed mice. J. Nutr. 2005;135(11):2553–2559. doi: 10.1093/jn/135.11.2553. [DOI] [PubMed] [Google Scholar]
- 60.Corpeleijn WE, Vliet Iv, Dana-Anne H, de Gast-Bakker, Schoor SRDvd, Alles MS, Hoijer M, Tibboel D, Goudoever JBv. Effect of enteral IGF-1 supplementation on feeding tolerance, growth, and gut permeability in enterally fed premature neonates. JPGN. 2008;46:184–190. doi: 10.1097/MPG.0b013e31815affec. [DOI] [PubMed] [Google Scholar]
- 61.Büyükkayhan D, Tanzer F, Erselcan T, Cinar Z, Yönem O. Umbilical serum insulin-like growth factor 1 (IGF-1) in newborns: effects of gestational age, postnatal age, and nutrition. Int J Vitam Nutr Res. 2003;73(5):343–346. doi: 10.1024/0300-9831.73.5.343. [DOI] [PubMed] [Google Scholar]
- 62.Philipps AF, Dvorak B, Kling PJ, Grille JG, Koldovsky O. Absorption of milk-borne insulin-like growth factor-I into portal blood of sucklin rats. JPGN. 2000;31:128–135. doi: 10.1097/00005176-200008000-00008. [DOI] [PubMed] [Google Scholar]
- 63.Kling PJ, Taing KM, Dvorak B, Woodward SS, Philipps AF. Insulin-like growth factor-I stimulates erythropoiesis when administered enterally. Growth Factors. 2006;24(3):218–223. doi: 10.1080/08977190600783162. [DOI] [PubMed] [Google Scholar]
- 64.Loui A, Eilers E, Strauss E, Pohl-Schickinger A, Obladen M, Koehne P. Vascular endothelial growth factor (VEGF) and soluble VEGF Receptor 1 (Sflt-1) levels in early and mature human milk from mothers of preterm versus term infants. J Hum. Lact. 2012 doi: 10.1177/0890334412447686. [DOI] [PubMed] [Google Scholar]
- 65.Reynolds JD. The management of Retinopathy of Prematurity. Paediatr Drugs. 2001;3(4):263–272. doi: 10.2165/00128072-200103040-00003. [DOI] [PubMed] [Google Scholar]
- 66.DiBiasie A. Evidence-based review of Retinopathy of Prematurity prevention in VLBW and ELBW Infants. Neonat. Network. 2006;25(6):393–403. doi: 10.1891/0730-0832.25.6.393. [DOI] [PubMed] [Google Scholar]
- 67.Kett JC. Anemia in infancy. Pediatr. Rev. 2012;33(4):186–187. doi: 10.1542/pir.33-4-186. [DOI] [PubMed] [Google Scholar]
- 68.Soubasi V, Kremenopoulos G, Diamanti E, Tsantali C, Sarafidis K, Tsakiris D. Follow-up of very low birth weight infants after erythropoietin treatment to prevent anemia of prematurity. J. Pediatr. 1995;127:291–297. doi: 10.1016/s0022-3476(95)70313-6. [DOI] [PubMed] [Google Scholar]
- 69.Carbonell-Estrany X, Figueras-Aloy J, Alvarez E. Erythropoietin and prematurity – where do we stand? J. Perinat. Med. 2005;33:277–286. doi: 10.1515/JPM.2005.054. [DOI] [PubMed] [Google Scholar]
- 70.Kling PJ, Willeitner A, Dvorak B, Blohowiak SE. Enteral erythropoietin and iron stimulate erythropoiesis in suckling rats. JPGN. 2008;46:202–207. doi: 10.1097/MPG.0b013e3181559e8b. [DOI] [PubMed] [Google Scholar]
- 71.Pasha YZ, Ahmadpour-Kacho M, Hajiahmadi M, Hosseini M. Enteral erythropoietin increases plasma erythropoietin level in preterm infants: a randomized controlled trial. Indian Pediatr. 2008;45:25–28. [PubMed] [Google Scholar]
- 72.Shiou S-R, Yu Y, Chen S, Ciancio MJ, Petrof EO, Sun J, Claud EC. Erythropoietin protects intestinal epithelial barrier function and lowers the incidence of experimental neonatal necrotizing enterocolitis. J. Biol. Chem. 2011;286(14):12123–12132. doi: 10.1074/jbc.M110.154625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Arsenault JE, Webb AL, Koulinska IN, Aboud S, Fawzi WW, Villamor E. Association between breast milk erythropoietin and reduced risk of mother-to-child transmission of HIV. JID. 2010;202(3):370–373. doi: 10.1086/653706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Claud EC, Savidge T, Walker WA. Modulation of human intestinal epithelial cell IL-8 secretion by human milk factors. Pediatr. Res. 2003;53:419–425. doi: 10.1203/01.PDR.0000050141.73528.AD. [DOI] [PubMed] [Google Scholar]
- 75.Struck J, Almeida Pd, Bergmann A, Morgenthaler NG. High Concentrations of Procalcitonin but Not Mature Calcitonin in Normal Human Milk. Horm. Metab. Res. 2002;34:460–465. doi: 10.1055/s-2002-33596. [DOI] [PubMed] [Google Scholar]
- 76.Wookey PJ, Turner K, Furness JB. Transient expression of the calcitonin receptor by enteric neurons of the embryonic and early post-natal mouse. Cell Tissue Res. 2012;347:311–317. doi: 10.1007/s00441-011-1303-6. [DOI] [PubMed] [Google Scholar]
- 77.Rao RK, Davis TP, Williams C, Koldovsky O. Effect of milk on somatostatin degradation in suckling rat jejunum in vivo. JPGN. 1999;28(1):84–94. doi: 10.1097/00005176-199901000-00019. [DOI] [PubMed] [Google Scholar]
- 78.Gama P, Alvares EP. LHRH and somatostatin effects on the cell proliferation of the gastric epithelium of suckling and weaning rats. Reg. Peptides. 1996;63:73–78. doi: 10.1016/0167-0115(96)00013-4. [DOI] [PubMed] [Google Scholar]
- 79.Newburg DS, Woo JG, Morrow AL. Characteristics and potential functions of human milk adiponectin. J. Pediatr. 2010;156:S41–S46. doi: 10.1016/j.jpeds.2009.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Martin LJ, Woo JG, Geraghty SR, Altaye M, Davidson BS, Banach W, Dolan LM, Ruiz-Palacios GM, Morrow AL. Adiponectin is present in human milk and is associated with maternal factors. Am. J. Clin. Nutr. 2006;83:1106–1111. doi: 10.1093/ajcn/83.5.1106. [DOI] [PubMed] [Google Scholar]
- 81.Woo JG, Guerrero ML, Guo F, Martin LJ, Davidson BS, Ortega H, Ruiz-Palacios GM, Morrow AL. Human milk adiponectin affects infant weight trajectory during the second year of life. Journal of pediatric gastroenterology and nutrition. 2012;54(4):532–539. doi: 10.1097/MPG.0b013e31823fde04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Savino F, Sorrenti M, Benetti S, Lupica MM, Liguori SA, Oggero R. Resistin and leptin in breast milk and infants in early life. Early Hum. Devel. 2012;88:779–782. doi: 10.1016/j.earlhumdev.2012.05.004. [DOI] [PubMed] [Google Scholar]
- 83.Savino F, Liguori SA. Update on breast milk hormones: Leptin, ghrelin and adiponectin. Clin. Nutr. 2008;27:42–47. doi: 10.1016/j.clnu.2007.06.006. [DOI] [PubMed] [Google Scholar]
- 84.Palou A, Sánchez J, Picó C. Nutrient–gene interactions in early life programming: leptin in breast milk prevents obesity later on in life. In: Koletzko B, editor. Early Nutrition Programming and Health: Outcomes in Later Life: Obesity and Beyond. Springer Science + Business Media B.V.; 2009. pp. 95–104. [DOI] [PubMed] [Google Scholar]
- 85.Dündar NO, Dündar B, Cesur G, Yılmaz N, Sütçu R, özgüner F. Ghrelin and adiponectin levels in colostrum, cord blood and maternal serum. Pediatr. Int. 2010;52:622–625. doi: 10.1111/j.1442-200X.2010.03100.x. [DOI] [PubMed] [Google Scholar]
- 86.Goldman AS. Modulation of the Gastrointestinal Tract of Infants by Human Milk. Interfaces and Interactions. An Evolutionary Perspective1. J. Nutr. 2000;130:426S–431S. doi: 10.1093/jn/130.2.426S. [DOI] [PubMed] [Google Scholar]
- 87.Jarvinen KM, Suomalainen H. Leucocytes in human milk and lymphocyte subsets in cow’s milk-allergic infants. Pediatr Allergy Immunol. 2002;13(4):243–254. doi: 10.1034/j.1399-3038.2002.00087.x. [DOI] [PubMed] [Google Scholar]
- 88.Patki S, Kadam S, Chandra V, Bhonde R. Human breast milk is a rich source of multipotent mesenchymal stem cells. Human cell. 2010;23(2):35–40. doi: 10.1111/j.1749-0774.2010.00083.x. [DOI] [PubMed] [Google Scholar]
- 89.Ichikawa M, Sugita M, Takahashi M, Satomi M, Takeshita T, Araki T, Takahashi H. Breast milk macrophages spontaneously produce granulocyte-macrophage colony-stimulating factor and differentiate into dendritic cells in the presence of exogenous interleukin-4 alone. Immunology. 2003;108(2):189–195. doi: 10.1046/j.1365-2567.2003.01572.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Indumathi S, Dhanasekaran M, Rajkumar JS, Sudarsanam D. Exploring the stem cell and non-stem cell constituents of human breast milk. Cytotechnology. 2012 doi: 10.1007/s10616-012-9492-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Riskin A, Almog M, Peri R, Halasz K, Srugo I, Kessel A. Changes in immunomodulatory constituents of human milk in response to active infection in the nursing infant. Pediatric research. 2012;71(2):220–225. doi: 10.1038/pr.2011.34. [DOI] [PubMed] [Google Scholar]
- 92.Sabbaj S, Ibegbu CC, Kourtis AP. Cellular immunity in breast milk: implications for postnatal transmission of HIV-1 to the infant. Advances in experimental medicine and biology. 2012;743:161–169. doi: 10.1007/978-1-4614-2251-8_11. [DOI] [PubMed] [Google Scholar]
- 93.Yagi Y, Watanabe E, Watari E, Shinya E, Satomi M, Takeshita T, Takahashi H. Inhibition of DC-SIGN-mediated transmission of human immunodeficiency virus type 1 by Toll-like receptor 3 signalling in breast milk macrophages. Immunology. 2010;130(4):597–607. doi: 10.1111/j.1365-2567.2010.03264.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Agarwal S, Karmaus W, Davis S, Gangur V. Immune markers in breast milk and fetal and maternal body fluids: a systematic review of perinatal concentrations. J Hum Lact. 2011;27(2):171–186. doi: 10.1177/0890334410395761. [DOI] [PubMed] [Google Scholar]
- 95.Groer MW, Beckstead JW. Multidimensional scaling of multiplex data: human milk cytokines. Biological research for nursing. 2011;13(3):289–296. doi: 10.1177/1099800411402055. [DOI] [PubMed] [Google Scholar]
- 96.Kverka M, Burianova J, Lodinova-Zadnikova R, Kocourkova I, Cinova J, Tuckova L, Tlaskalova-Hogenova H. Cytokine profiling in human colostrum and milk by protein array. Clinical chemistry. 2007;53(5):955–962. doi: 10.1373/clinchem.2006.077107. [DOI] [PubMed] [Google Scholar]
- 97.Penttila IA. Milk-derived transforming growth factor-b and the infant immune response. J. Pediatr. 2010;156:S21–S25. doi: 10.1016/j.jpeds.2009.11.016. [DOI] [PubMed] [Google Scholar]
- 98.Kalliomäki M, Ouwehand A, Arvilommi H, Kero P, Isolauri E. Transforming growth factor-β in breast milk: A potential regulator of atopic disease at an early age. J. Allergy Clin. Immunol. 1999:1251–1257. doi: 10.1016/s0091-6749(99)70021-7. [DOI] [PubMed] [Google Scholar]
- 99.Saito S, Yoshida M, Ichijo M, Ishizaka S, Tsujii T. Transforming growth factor-beta (TGF-f) in human milk. Clin. Exp. Immunol. 1993;94:220–224. doi: 10.1111/j.1365-2249.1993.tb06004.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Nakamura Y, Miyata M, Ando T, Shimokawa N, Ohnuma Y, Katoh R, Ogawa H, Okumura K, Nakao A. The latent form of transforming growth factor-b administered orally is activated by gastric acid in mice. J. Nutr. 2009;139:1463–1468. doi: 10.3945/jn.109.108761. [DOI] [PubMed] [Google Scholar]
- 101.Letterio JJ, Geiser AG, Kulkarni Ashok B., Roche NS, Sporn MB, Roberts AB. Maternal rescue of transforming growth factor-b1 null mice. Science. 1994;264:1936–1938. doi: 10.1126/science.8009224. [DOI] [PubMed] [Google Scholar]
- 102.Ando T, Hatsushika K, Wako M, Ohba T, Koyama K, Ohnuma Y, Katoh R, Ogawa H, Okumura K, Luo J, Wyss-Coray T, Nakao A. Orally administered TGF-b is biologically active in the intestinal mucosa and enhances oral tolerance. J. Allergy Clin. Immunol. 2007;120:916–923. doi: 10.1016/j.jaci.2007.05.023. [DOI] [PubMed] [Google Scholar]
- 103.Ozawa T, Miyata M, Nishimura M, Ando T, Ouyang Y, Ohba T, Shimokawa N, Ohnuma Y, Katoh R, Ogawa H, Nakao A. Transforming growth factor-b activity in commercially available pasteurized cow milk provides protection against inflammation in mice. J. Nutr. 2009;139:69–75. doi: 10.3945/jn.108.092528. [DOI] [PubMed] [Google Scholar]
- 104.Donnet-Hughes A, Duc N, Serrant P, Vidal K, Schiffrin EJ. Bioactive molecules in milk and their role in health and disease: The role of transforming growth factor-b. Immunol. Cell Biol. 2000;78:74–79. doi: 10.1046/j.1440-1711.2000.00882.x. [DOI] [PubMed] [Google Scholar]
- 105.Gilmore W, McKelvey-Martin V, Rutherford S, Strain J, Loane P, Kell M, Millar S. Human milk contains granulocyte colony stimulating factor. Eur J Clin Nutr. 1994;48(3):222–224. [PubMed] [Google Scholar]
- 106.Gersting JA, Kotto-Komeb CA, Dua Y, Christensen RD, Calhoun DA. Bioavailability of granulocyte colony-stimulating factor administered enterally to suckling mice. Pharmacol. Res. 2003;48:643–647. doi: 10.1016/s1043-6618(03)00249-4. [DOI] [PubMed] [Google Scholar]
- 107.Calhoun DA, Maheshwari A, Christensen RD. Recombinant Granulocyte Colony-Stimulating Factor administered enterally to neonates is not absorbed. Pediatrics. 2003;112(2):421–423. doi: 10.1542/peds.112.2.421. [DOI] [PubMed] [Google Scholar]
- 108.Gersting JA, Christensen RD, Calhoun DA. Effects of enterally administering granulocyte colony-stimulating factor to suckling mice. Pediatr Res. 2004;55(5):802–806. doi: 10.1203/01.PDR.0000117846.51197.7C. [DOI] [PubMed] [Google Scholar]
- 109.Ngom P, Collinson A, Pido-Lopez J, Henson S, Prentice A, Aspinall R. Improved thymic function in exclusively breastfed infants is associated with higher interleukin 7 concentrations in their mothers’ breast milk. Am. J. Clin. Nutr. 2004;80(3):722–728. doi: 10.1093/ajcn/80.3.722. [DOI] [PubMed] [Google Scholar]
- 110.Aspinall R, Prentice AM, Ngom PT. Interleukin 7 from maternal milk crosses the intestinal barrier and modulates T-cell development in offspring. PloS ONE. 2011;6(6) doi: 10.1371/journal.pone.0020812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Ustundag B, Yilmaz E, Dogan Y, Akarsu S, Canatan H, Halifeoglu I, Cikim G, Aygun AD. Levels of cytokines (IL-1beta, IL-2, IL-6, IL-8, TNF-alpha) and trace elements (Zn, Cu) in breast milk from mothers of preterm and term infants. Mediators of inflammation. 2005;2005(6):331–336. doi: 10.1155/MI.2005.331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Buescher ES, McWilliams-Koeppen P. Soluble Tumor Necrosis Factor-α (TNF-α) Receptors in human colostrum and milk bind to TNF-α and neutralize TNF-α bioactivity. Pediatr Res. 1998;44:37–42. doi: 10.1203/00006450-199807000-00006. [DOI] [PubMed] [Google Scholar]
- 113.Buescher E, Malinowska I. Soluble receptors and cytokine antagonists in human milk. Pediatr Res. 1996;40(6):839–844. doi: 10.1203/00006450-199612000-00011. [DOI] [PubMed] [Google Scholar]
- 114.Erbağci A, Cekmen M, Balat O, Balat A, Aksoy F, M. MT. Persistency of high proinflammatory cytokine levels from colostrum to mature milk in preeclampsia. Clin. Biochem. 2005;38(8):712–716. doi: 10.1016/j.clinbiochem.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 115.Meki A-RMA, Saleem TH, Al-Ghazali MH, Sayed AA. Interleukins -6, -8 and -10 and tumor necrosis factor alpha and its soluble receptor I in human milk at different periods of lactation. Nutr. Res. 2003;23:845–855. [Google Scholar]
- 116.Maheshwari A, Lu W, Lacson A, Barleycorn AA, Nolan S, Christensen RD, Calhoun DA. Effects of Interleukin-8 on the developing human intestine. Cytokine. 2002;20(6):256–267. doi: 10.1006/cyto.2002.1996. [DOI] [PubMed] [Google Scholar]
- 117.Maheshwari A, Lacson A, Lu W, Fox SE, Barleycorn AA, Christensen RD, Calhoun DA. Interleukin-8/CXCL8 forms an autocrine loop in fetal intestinal mucosa. Pediatr Res. 2004;56(2):240–249. doi: 10.1203/01.PDR.0000133196.25949.98. [DOI] [PubMed] [Google Scholar]
- 118.Maheshwari A, Christensen RD, Calhoun DA. ELR+ CXC chemokines in human milk. Cytokine. 2003;24:91–102. doi: 10.1016/j.cyto.2003.07.002. [DOI] [PubMed] [Google Scholar]
- 119.Mizuno K, Hatsuno M, Aikawa K, Takeichi H, Himi T, Kaneko A, Kodaira K, Takahashi H, Itabashi K. Mastitis is associated with IL-6 Levels and milk fat globule size in breast milk. J. Hum. Lact. 2012 doi: 10.1177/0890334412455946. [DOI] [PubMed] [Google Scholar]
- 120.Hunt KM, Williams JE, Shafii B, Hunt MK, Behre R, Ting R, McGuire MK, McGuire MA. Mastitis Is Associated with Increased Free Fatty Acids, Somatic Cell Count, and Interleukin-8 Concentrations in Human Milk. Breastfeeding Med. 2012 doi: 10.1089/bfm.2011.0141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Hrdý J, Novotná O, Kocourková I, Prokeśová L. Cytokine expression in the colostral cells of healthy and allergic mothers. Folia Microbiologica. 2012;57(3):215–219. doi: 10.1007/s12223-012-0112-y. [DOI] [PubMed] [Google Scholar]
- 122.Strand TA, Sharma PR, Gjessing HK, Ulak M, Chandyo RK, Adhikari RK, Sommerfelt H. Risk factors for extended duration of acute diarrhea in young children. PloS ONE. 2012;7(5) doi: 10.1371/journal.pone.0036436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Lamberti LM, Walker CLF, Noiman A, Victora C, Black RE. Breastfeeding and the risk for diarrhea morbidity and mortality. BMC Pub. Health. 2011;11(Supp. 3) doi: 10.1186/1471-2458-11-S3-S15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Brandtzaeg P. The mucosal immune system and its integration with the mammary glands. The Journal of pediatrics. 2010;156(2 Suppl):S8–15. doi: 10.1016/j.jpeds.2009.11.014. [DOI] [PubMed] [Google Scholar]
- 125.Kadaoui KA, Corthésy B. Secretory IgA mediates bacterial translocation to dendritic cells in mouse peyer’s patches with restriction to mucosal compartment. J. Immunol. 2007;179:7751–7757. doi: 10.4049/jimmunol.179.11.7751. [DOI] [PubMed] [Google Scholar]
- 126.Corthesy B. Secretory immunoglobulin A: well beyond immune exclusion at mucosal surfaces. Immunopharmacology and immunotoxicology. 2009;31(2):174–179. doi: 10.1080/08923970802438441. [DOI] [PubMed] [Google Scholar]
- 127.Hurley WL, Theil PK. Perspectives on immunoglobulins in colostrum and milk. Nutrients. 2011;3(4):442–474. doi: 10.3390/nu3040442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Velona T, Abbiati L, Beretta B, Gaiaschi A, Flauto U, Tagliabue P, Galli CL, Restani P. Protein profiles in breast milk from mothers delivering term and preterm babies. Pediatr Res. 1999;45(5):658–663. doi: 10.1203/00006450-199905010-00008. [DOI] [PubMed] [Google Scholar]
- 129.Adamkin DH. Mother’s milk, feeding strategies, and lactoferrin to prevent necrotizing enterocolitis. JPEN. 2012:36. doi: 10.1177/0148607111420158. [DOI] [PubMed] [Google Scholar]
- 130.Beljaars L, van der Strate BW, Bakker HI, Reker-Smit C, van Loenen-Weemaes AM, Wiegmans FC, Harmsen MC, Molema G, Meijer DK. Inhibition of cytomegalovirus infection by lactoferrin in vitro and in vivo. Antiviral Res. 2004;63(3):197–208. doi: 10.1016/j.antiviral.2004.05.002. [DOI] [PubMed] [Google Scholar]
- 131.Kuipers ME, de Vries HG, Eikelboom MC, Meijer DK, Swart PJ. Synergistic fungistatic effects of lactoferrin in combination with antifungal drugs against clinical Candida isolates. Antimicrobial agents and chemotherapy. 1999;43(11):2635–2641. doi: 10.1128/aac.43.11.2635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Leitch EC, Willcox MD. Lactoferrin increases the susceptibility of S. epidermidis biofilms to lysozyme and vancomycin. Current eye research. 1999;19(1):12–19. doi: 10.1076/ceyr.19.1.12.5342. [DOI] [PubMed] [Google Scholar]
- 133.Manzoni P, Rinaldi M, Cattani S, Pugni L, Romeo MG, Messner H, Stolfi I, Decembrino L, Laforgia N, Vagnarelli F, Memo L, Bordignon L, Saia OS, Maule M, Gallo E, Mostert M, Magnani C, Quercia M, Bollani L, Pedicino R, Renzullo L, Betta P, Mosca F, Ferrari F, Magaldi R, Stronati M, Farina D. Bovine lactoferrin supplementation for prevention of late-onset sepsis in very low-birth-weight neonates: a randomized trial. Jama. 2009;302(13):1421–1428. doi: 10.1001/jama.2009.1403. [DOI] [PubMed] [Google Scholar]
- 134.Sherman MP, Bennett SH, Hwang FF, Yu C. Neonatal small bowel epithelia: enhancing anti-bacterial defense with lactoferrin and Lactobacillus GG. Biometals. 2004;17(3):285–289. doi: 10.1023/b:biom.0000027706.51112.62. [DOI] [PubMed] [Google Scholar]
- 135.Stubbs J, Lekutis C, Singer K, Bui A, Yuzuki D, Srinivasan U, Parry G. cDNA cloning of a mouse mammary epithelial cell surface protein reveals the existence of epidermal growth factor-like domains linked to factor VIII-like sequences. PNAS. 1990;87(21):8417–8421. doi: 10.1073/pnas.87.21.8417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Peterson J, Hamosh M, Scallan C, Ceriani R, Henderson T, Mehta N, Armand M, Hamosh P. Milk fat globule glycoproteins in human milk and in gastric aspirates of mother’s milk-fed preterm infants. Pediatr Res. 1998;44(4):499–506. doi: 10.1203/00006450-199810000-00006. [DOI] [PubMed] [Google Scholar]
- 137.Newburg D, Peterson J, Ruiz-Palacios G, Matson D, Morrow A, Shults J, Guerrero M, Chaturvedi P, Newburg S, Scallan C, MRTaylor, Ceriani R, Pickering L. Role of human-milk lactadherin in protection against symptomatic rotavirus infection. The Lancet. 1998;351(9110):1160–1164. doi: 10.1016/s0140-6736(97)10322-1. [DOI] [PubMed] [Google Scholar]
- 138.Aziz M, Jacob A, Matsuda A, Wang P. Review: milk fat globule-EGF factor 8 expression, function and plausible signal transduction in resolving inflammation. Apoptosis. 2011;16:1077–1086. doi: 10.1007/s10495-011-0630-0. [DOI] [PubMed] [Google Scholar]
- 139.Shi J, Heegaard CW, Rasmussen JT, Gilbert GE. Lactadherin binds selectively to membranes containing phosphatidyl-l-serine and increased curvature. Biochim Biophys Acta. 2004;1667:82–90. doi: 10.1016/j.bbamem.2004.09.006. [DOI] [PubMed] [Google Scholar]
- 140.Kusunoki R, Ishihara S, Aziz M, Oka A, Tada Y, Kinoshita Y. Roles of Milk Fat Globule-Epidermal Growth Factor 8 in intestinal inflammation. Digestion. 2012;85:103–107. doi: 10.1159/000334679. [DOI] [PubMed] [Google Scholar]
- 141.Chogle A, Bu H-F, Wang X, Brown JB, Chou PM, Tan X-D. Milk Fat Globule–EGF Factor 8 is a critical protein for healing of dextran sodium sulfate–induced acute colitis in mice. Mol. Med. 2011;17(5):502–207. doi: 10.2119/molmed.2010.00074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Baghdadi M, Chiba S, Yamashina T, Yoshiyama H, Jinushi M. MFG-E8 regulates the immunogenic potential of dendritic cells primed with necrotic cell-mediated inflammatory signals. PloS ONE. 2012;7(6) doi: 10.1371/journal.pone.0039607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Landberg E, Huang Y, Stromqvist M, Mechref Y, Hansson L, Lundblad A, Novotny MV, Påhlsson P. Changes in glycosylation of human Bile-Salt-Stimulated Lipase during lactation. Arch. Biochem. Biophys. 2000;377(2):246–254. doi: 10.1006/abbi.2000.1778. [DOI] [PubMed] [Google Scholar]
- 144.Stax MJ, Naarding MA, Tanck MWT, Lindquist S, Hernell O, Lyle R, Brandtzaeg P, Eggesbø M, Pollakis G, Paxton WA. Binding of human milk to pathogen receptor DC-SIGN varies with bile salt-stimulated lipase (BSSL) gene polymorphism. PloS ONE. 2011;6(2) doi: 10.1371/journal.pone.0017316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Naarding MA, Dirac AM, Ludwig IS, Speijer D, Lindquist S, Vestman E-L, Stax MJ, Geijtenbeek TBH, Pollakis G, Hernell O, Paxton WA. Bile Salt-Stimulated Lipase from human milk binds DC-SIGN and inhibits Human Immunodeficiency Virus type 1 transfer to CD4+ T cells Antimicrob. Agents Chemother. 2006;50(10):3367–3374. doi: 10.1128/AAC.00593-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Ruvoen-Clouet N, Mas E, Mariounneau S, Guillon P, Lombardo D, Pendu JL. Bile-salt-stimulated lipase and mucins from milk of ‘secretor’ mothers inhibit the binding of Norwalk virus capsids to their carbohydrate ligands. Biochem J. 2006;393:627–634. doi: 10.1042/BJ20050898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Hettinga K, Valenberg Hv, Vries Sd, Boeren S, Hooijdonk Tv, Arendonk Jv, Vervoort J. The host defense proteome of human and bovine milk. PloS ONE. 2011;6(4) doi: 10.1371/journal.pone.0019433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Saeland E, Jong MAWPd, Nabatov AA, Kalay H, Geijtenbeek TBH, Kooyk Yv. MUC1 in human milk blocks transmission of human immunodeficiency virus from dendritic cells to T cells. Mol. Immunol. 2009;46:2309–2316. doi: 10.1016/j.molimm.2009.03.025. [DOI] [PubMed] [Google Scholar]
- 149.Yolken RH, Peterson JA, Vonderfecht SL, Fouts ET, Midthun K, Newburg DS. Human milk mucin inhibits rotavirus replication and prevents experimental gastroenteritis. J. Clin. Invest. 1992;90:1984–1987. doi: 10.1172/JCI116078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Parker P, Sando L, Pearson R, Kongsuwan K, Tellam RL, Smith S. Bovine Muc1 inhibits binding of enteric bacteria to Caco-2 cells. Glycoconj. 2010;27:89–97. doi: 10.1007/s10719-009-9269-2. [DOI] [PubMed] [Google Scholar]
- 151.Liu B, Yu Z, Chen C, Kling DE, Newburg DS. Human milk mucin 1 and mucin 4 inhibit Salmonella enterica serovar typhimurium invasion of human intestinal epithelial cells in vitro. J. Nutr. 2012;142:1504–1509. doi: 10.3945/jn.111.155614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Bode L, Kuhn L, Kim HY, Hsiao L, Nissan C, Sinkala M, Kankasa C, Mwiya M, Thea DM, Aldrovandi GM. Human milk oligosaccharide concentration and risk of postnatal transmission of HIV through breastfeeding. The American journal of clinical nutrition. 2012;96(4):831–839. doi: 10.3945/ajcn.112.039503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.De Leoz ML, Gaerlan SC, Strum JS, Dimapasoc LM, Mirmiran M, Tancredi DJ, Smilowitz JT, Kalanetra KM, Mills DA, German JB, Lebrilla CB, Underwood MA. Lacto-N-tetraose, fucosylation, and secretor status are highly variable in human milk oligosaccharides from women delivering preterm. J Proteome Res. 2012;11(9):4662–4672. doi: 10.1021/pr3004979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Hunt KM, Foster JA, Forney LJ, Schutte UM, Beck DL, Abdo Z, Fox LK, Williams JE, McGuire MK, McGuire MA. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE. 2011;6(6):e21313. doi: 10.1371/journal.pone.0021313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Cabrera-Rubio R, Collado MC, Laitinen K, Salminen S, Isolauri E, Mira A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. The American journal of clinical nutrition. 2012;96(3):544–551. doi: 10.3945/ajcn.112.037382. [DOI] [PubMed] [Google Scholar]
- 156.Rasmussen KM, Geraghty SR. The quiet revolution: breastfeeding transformed with the use of breast pumps. Am J Public Health. 2011;101(8):1356–1359. doi: 10.2105/AJPH.2011.300136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Baro C, Giribaldi M, Arslanoglu S, Giuffrida MG, Dellavalle G, Conti A, Tonetto P, Biasini A, Coscia A, Fabris C, Moro GE, Cavallarin L, Bertino E. Effect of two pasteurization methods on the protein content of human milk. Front Biosci (Elite Ed) 2011;3:818–829. doi: 10.2741/e289. [DOI] [PubMed] [Google Scholar]
- 158.Chantry CJ, Wiedeman J, Buehring G, Peerson JM, Hayfron K, K’Aluoch O, Lonnerdal B, Israel-Ballard K, Coutsoudis A, Abrams B. Effect of flash-heat treatment on antimicrobial activity of breastmilk. Breastfeed Med. 2011;6(3):111–116. doi: 10.1089/bfm.2010.0078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Israel-Ballard K, Donovan R, Chantry C, Coutsoudis A, Sheppard H, Sibeko L, Abrams B. Flash-heat inactivation of HIV-1 in human milk: a potential method to reduce postnatal transmission in developing countries. J Acquir Immune Defic Syndr. 2007;45(3):318–323. doi: 10.1097/QAI.0b013e318074eeca. [DOI] [PubMed] [Google Scholar]
- 160.Peroni D, Piacentini G, Bodini A, Pigozzi R, Boner A. Transforming growth factor-b1 is elevated in unpasteurized cow’s milk. Pediatr. Allergy Immunol. 2009;20:42–44. doi: 10.1111/j.1399-3038.2008.00737.x. [DOI] [PubMed] [Google Scholar]
- 161. [August 23, 2012];Agennix. Phase 1/2 Study of Talactoferrin Oral Solution ofr Nosocomial Infection in Preterm Infants. http://clinicaltrials.gov/ct2/show/NCT00854633.
- 162.Ewaschuk JB, Unger S, O’Connor DL, Stone D, Harvey S, Clandinin MT, Field CJ. Effect of pasteurization on selected immune components of donated human breast milk. J Perinatol. 2011;31(9):593–598. doi: 10.1038/jp.2010.209. [DOI] [PubMed] [Google Scholar]
- 163.Akinbi H, Meinzen-Derr J, Auer C, Ma Y, Pullum D, Kusano R, Reszka KJ, Zimmerly K. Alterations in the host defense properties of human milk following prolonged storage or pasteurization. Journal of pediatric gastroenterology and nutrition. 2010;51(3):347–352. doi: 10.1097/MPG.0b013e3181e07f0a. [DOI] [PubMed] [Google Scholar]
- 164.Valentine CJ. Optimizing Humn Milk Fortification for the Preterm Infant. PNPG Building Block for Life. 2011;34(4):9–11. [Google Scholar]
- 165.Steinhoff MC, Omer SB. A review of fetal and infant protection associated with antenatal influenza immunization. American journal of obstetrics and gynecology. 2012;207(3 Suppl):S21–27. doi: 10.1016/j.ajog.2012.06.071. [DOI] [PubMed] [Google Scholar]
- 166.Neville MC, Anderson SM, McManaman JL, Badger TM, Bunik M, Contractor N, Crume T, Dabelea D, Donovan SM, Forman N, Frank DN, Friedman JE, German JB, Goldman A, Hadsell D, Hambidge M, Hinde K, Horseman ND, Hovey RC, Janoff E, Krebs NF, Lebrilla CB, Lemay DG, Maclean PS, Meier P, Morrow AL, Neu J, Nommsen-Rivers LA, Raiten DJ, Rijnkels M, Seewaldt V, Shur BD, Vanhouten J, Williamson P. Lactation and neonatal nutrition: defining and refining the critical questions. Journal of mammary gland biology and neoplasia. 2012;17(2):167–188. doi: 10.1007/s10911-012-9261-5. [DOI] [PMC free article] [PubMed] [Google Scholar]