

Abstract
Interventions to effect therapeutic cardiomyocyte renewal have received considerable interest of late. Such interventions, if successful, could give rise to myocardial regeneration in diseased hearts. Regenerative interventions fall into two broad categories, namely approaches based on promoting renewal of pre-existing cardiomyocytes and approaches based on cardiomyogenic stem cell activity. The latter category can be further subdivided into approaches promoting differentiation of endogenous cardiomyogenic stem cells, approaches wherein cardiomyogenic stem cells are harvested, amplified or enriched ex vivo, and subsequently engrafted into the heart, and approaches wherein an exogenous stem cell is induced to differentiate in vitro, and the resulting cardiomyocytes are engrafted into the heart. There is disagreement in the literature regarding the degree to which cardiomyocyte renewal occurs in the normal and injured heart, the mechanism(s) by which this occurs, and the degree to which therapeutic interventions can enhance regenerative growth. This review discusses several caveats which are encountered when attempting to measure cardiomyocyte renewal in vivo which likely contribute, at least in part, to the disagreement regarding the levels at which this occurs in normal, injured and treated hearts.
Keywords: cardiac regeneration, cardiomyogensis, cardiomyocyte proliferation
The regenerative potential of the adult myocardium in cold-blooded animals is well established. Indeed, the initial observation suggesting regenerative growth following injury, in frogs, was reported in 1875 [1]. A substantive body of work, largely by the Rumyantsev (summarized in [2]) and Oberpriller [3–7] laboratories in the 1970s–1980s, further characterized this phenomenon, providing evidence that cardiomyocyte proliferation was a major component of the phenomenon. More recent work established a similar regenerative phenotype in zebrafish [8], a particularly important observation given the suitability of that species for mutational screens. Importantly, the magnitude of cardiomyocyte renewal in lower invertebrates and reptiles is sufficiently high so as to be readily detected by simple histologic analysis at various time points post-injury.
More recent studies examining potential therapeutic interventions to promote cardiac regeneration speak of challenging “the old dogma” that the adult mammalian heart lacks regenerative potential. In fact, the notion of on-going cardiomyocyte renewal in the mammalian heart is not a new idea. Many studies have examined cardiomyocyte cell cycle activity in normal and injured adult hearts, and the presence of ongoing cardiomyocyte renewal has been long accepted. Rather, the debate in the literature resides in the magnitude at which this occurs. For example, in the uninjured adult rat heart, values for ventricular cardiomyocyte cell cycle activity (which is often used as a surrogate marker for cardiomyocyte renewal) varied from 0 to 3.15% (reviewed in [9]). This variation likely reflects a number of factors, including the duration of the cell cycle marker being scored. For example, the duration of S-phase is much greater than M-phase, and as such assays monitoring cardiomyocyte DNA synthesis (i.e., tritiated thymidine or bromodeoxyuridine incorporation) would by default give rise to a proportionally higher renewal rate than assays monitoring the presence of mitotic figures. Other markers (such as Proliferating Cell Nuclear Antigen or Ki67 immune reactivity) which are expressed throughout much of the cell cycle would give rise to even higher renewal rates.
Another factor contributing to the high level of variation reported for baseline cardiomyocyte cell cycle activity is the accuracy with which cardiomyocytes (and in particular cardiomyocyte nuclei) are identified. It has been argued that the use of confocal microscopy, in combination with immune fluorescence visualization of a cytoplasmic marker (as for example, Troponin T) and a cell membrane marker (as for example, wheat germ agglutinin or laminin) can be used to unequivocally identify cardiomyocyte nuclei in tissue sections [10]. In practice, the z-axis resolution of confocal microscopy is insufficient to resolve non-cardiomyocyte nuclei in close (ie <0.5 microns) proximity to cardiomyocyte cytoplasm [11, 12], thus precluding accurate cardiomyocyte nucleus identification in some instances. This is an important consideration, since although cardiomyocytes comprise more than 90% of the mass of the heart, they constitute only 20% of the total cell number.
This caveat was nicely illustrated in experiments using a transgenic mouse wherein the cardiomyocyte-restricted alpha myosin heavy chain promoter [13] targeted expression of a nuclear localized beta-galactosidase reporter (the transgene is depicted in Figure 1A). These mice (designated MHC-nLAC mice [14]), can be used to identify cardiomyocyte nuclei in tissue sections by simple reaction with a chromogenic beta-galactosidase substrate (i.e., X-GAL; Figure 1B, blue signal) or by beta-galactosidase immune fluorescence (Figure 1C, green signal). Sections from these hearts were processed for beta-galactosidase (to identify cardiomyocyte nuclei) and Troponin T (to identify cardiomyocyte cytoplasm) immune reactivity, and stained with wheat germ agglutinin (to identify cell membranes) and Hoechst (to all identify nuclei). The sections were then imaged via confocal microscopy using a high numerical aperture objective, and Z-axis stacks comprising 42 steps at intervals of 0.24 μm for each imaged volume were generated. Observers were supplied with image volumes containing the Troponin T, wheat germ agglutinin and Hoechst signals, and were asked to identify the origin (i.e., cardiomyocyte or non-cardiomyocyte) of each nucleus. These results were then directly compared to those obtained with beta-galactosidase immune fluorescence within the same tissue volume. Significant error rates were observed [15], underscoring the subjectivity of traditional confocal microscopic analyses for cardiomyocyte nuclear identification.
Figure 1.
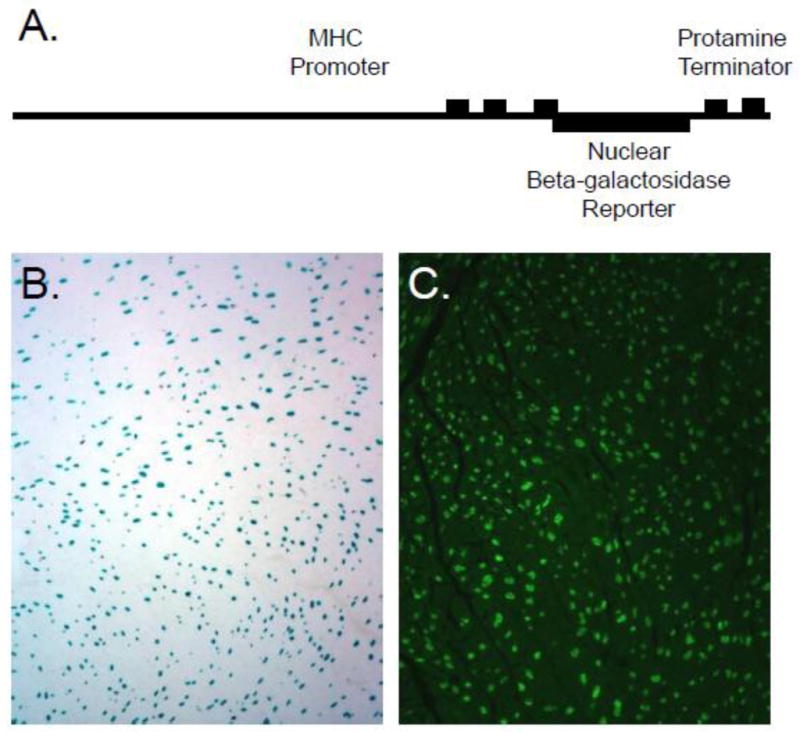
Detection of cardiomyocyte nuclei in MHC-nLAC transgenic mice. A. Schematic depiction of the MHC-nLAC transgene. The MHC promoter consists of 4500 bp of 5′ flanking sequence plus Exons 1, 2 and the non-coding region of Exon 3. The protamine terminator was inserted down-stream of the beta-galactosidase sequence to ensure proper processing of transgene-encoded transcripts. B. Low power image of a section from an adult MHC-nLAC transgenic heart following reaction with X-GAL, a chromogenic beta-galactosidase substrate. Cardiomyocyte nuclei appear blue. C. Low power image of a section from an adult MHC-nLAC transgenic heart processed for beta-galactosidase immune reactivity. Cardiomyocyte nuclei are identified by green fluorescence.
We have used the MHC-nLAC mice to monitor cardiomyocyte cell cycle activity in normal adult hearts. For these experiments, the mice received a single injection of tritiated thymidine, and were sacrificed four hours later. The hearts were harvested, sectioned, and processed for X-GAL reaction (to identify cardiomyocyte nuclei) and autoradiography (to identify S-phase nuclei). A typical cardiomyocyte in S-phase is shown in Figure 2A; using this approach only 0.0005% of the cardiomyocyte nuclei in adult hearts were thymidine positive [16]. While this is a relatively low rate of cell cycle activity, it is important to note that it represents only those cardiomyocytes which were synthesizing DNA during the 4 hour labeling session. If one assumes a linear relationship, multiplying by 6 would provide a daily cell cycle rate (i.e., 24 hours/4 hours), and multiplying further by 365 would provide a yearly rate. This would predict a cardiomyocyte renewal rate of 1.09% per year in the uninjured adult mouse heart, assuming that the DNA synthesis events culminated in cytokinesis. This value is remarkably close to the human cardiomyocyte renewal rate calculated by the Frisén laboratory [17]. These later experiments utilized a carbon dating approach that exploited the spike in atmospheric radioactive carbon resulting from numerous above-ground nuclear tests in the 1950s and 1960s, in combination with a cardiomyocyte nuclear-specific marker, and predicted an annual cardiomyocyte renewal rate of 1% per year in young adults.
Figure 2.
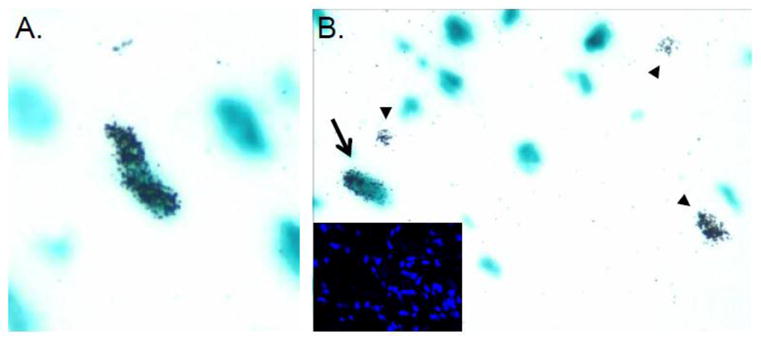
Use of tritiated thymidine incorporation in conjuction with the MHC-nLAC reporter transgene to detect cardiomyocyte cell cycle activity in normal and injured hearts. A. Cardiomyocyte DNA synthesis in a normal heart from a mouse receiving a single injection of tritiated thymidine. The image shows a section following X-GAL reaction and autoradiography. An S-phase cardiomyocyte nucleus is identified by the presence of silver grains over blue signal. B. Detection of cardiomyocyte (arrow) and non-cardiomyocyte (arrowheads) DNA synthesis at the peri-infarct zone in a mouse carrying the MHC-nLAC reporter transgene following a single injection of tritiated thymidine. Insert shows the Hoechst epifluorescence signal from the same field.
Monitoring cardiomyocyte DNA synthesis per se does not necessarily discriminate between renewal via proliferation of pre-existing cardiomyocytes vs. cardiomyogenic differentiation of stem cells. For example, the human experiment was cumulative in nature, and could detect DNA synthesis events in pre-existing cardiomyocytes as well as in stem cells which subsequently differentiated into cardiomyocytes. In contrast, the labeling period in the mouse experiment was too short to detect a de novo cardiomyogenic events; it could only detect DNA synthesis events in pre-existing cardiomyocytes. The fact that the normalized mouse data agreed well with the cumulative human data provides circumstantial data suggesting that the primary mechanism for cardiomyocyte renewal in the uninjured adult heart is via proliferation of pre-existing cardiomyocytes.
There are also many reports examining cardiomyocyte renewal following myocardial injury in mammalian hearts. Once again, the reported levels of cardiomyocyte cell cycle re-entry, and the degree to which this results in cardiomyocyte renewal, varied greatly between laboratories and appeared to be greatly influenced by the age of the animal at the time of injury, the method used to monitor cell cycle entry, the anatomical position of the cells studied, and the timing of the analyses post-injury (reviewed in [9]). With regards to age at the time of injury, studies in rats from the Gerdes laboratory clearly demonstrated that high levels of cardiomyocyte renewal could be induced during early post-natal life [18], a finding that was recently confirmed by surgical interventions in mice [19]. In mice, this potential for renewal was markedly decreased by post-natal day 7, which coincides with the cessation of developmental cardiomyocyte proliferation in normal mice [20].
With regards to the method used to monitor cardiomyocyte renewal, all of the caveats raised above regarding the nuances of a given cell cycle marker, as well as the identification of cardiomyocyte nuclei, in normal tissue are also applicable when examining injured tissue. Indeed, since injury results in a massive influx of immune cells, as well as a pronounced fibrotic response which is accompanied by the amplification of myofibroblasts, nuclear identification becomes even more problematic. Once again this is nicely illustrated using the MHC-nLAC mice. Figure 2B shows an image of the peri-infarct zone of an MHC-nLAC mouse which received a single injection of tritiated thymidine 2 weeks following permanent coronary artery ligation. The heart was harvested four hours later and sectioned, and the section was reacted with X-GAL and processed for autoradiography. A single S-phase cardiomyocyte nucleus (arrow) and multiple S-phase non-cardiomyocyte nuclei (arrowheads) are seen. A vast increase in the number of non-myocytes is evident in the Hoechst image from the same field (insert). Retrospective analysis from our laboratory revealed an average border zone (defined arbitrarily as 500 microns from the myocardium/scar interface) cardiomyocyte nuclear labeling index of roughly 0.1% using this approach.
Given the relatively low levels of renewal, analyses based on cumulative label incorporation have the advantage of circumventing errors resulting from sampling size. Bromodeoxyuridine, delivered in drinking water or via implanted osmotic mini-pumps, provides a suitable approach. The MHC-nLAC mice are also useful for assessment of bromodeoxyuridine incorporation. Figure 3A shows images from an MHC-nLAC mouse following a single injection of bromodeoxyuridine; the heart was harvested four hours later and sectioned. The sections were then processed for beta-galactosidase (green signal, left panel) and bromodeoxyuridine (red signal, middle panel) immune reactivity. The same field was also imaged using a dual-channel filter (right panel); the S-phase cardiomyocyte nucleus appears yellow due to the overlay of green beta-galactosidase and red bromodeoxyuridine signals (indicated by the arrow in all three panels). When infarcted MHC-nLAC mice were subjected to cumulative labeling analyses (7 day bromodeoxyuridine infusion using an osmotic mini-pump), a proportionate increase in the cardiomyocyte labeling index at the infarct border zone was observed. However, a dramatic increase in the non-cardiomyocyte labeling index was also observed (Figure 3B; image was captured with a dual-channel filter, S-phase cardiomyocyte nuclei are yellow, S-phase non-myocyte nuclei are red), further underscoring the need to have a non-subjective assay to identify cardiomyocyte nuclei, particularly when employing cumulative label assays.
Figure 3.

Use of bromodeoxyuridine incorporation in conjuction with the MHC-nLAC reporter transgene to detect cardiomyocyte cell cycle activity in normal and injured hearts. A. Cardiomyocyte DNA synthesis in a normal heart from a mouse receiving a single injection of bromodeoxyuridine. The images show the same field of a section following processing for beta-galactosidase (green fluorescence, left panel) and bromodeoxyuridine (red fluorescence, middle panel) immune reactivity. The image in the right panel was captured using a dual-channel filter; the S-phase cardiomyocyte nucleus (indicated by the arrow in all three panels) appears yellow due to the overlay of green and red fluorescence. B. Detection of cardiomyocyte (yellow signal) and non-cardiomyocyte (red signal) DNA synthesis at the peri-infarct zone in a mouse carrying the MHC-nLAC reporter transgene following 7 days of bromodeoxyuridine infusion. Cardiomyocyte nuclei which did not enter S-phase during the labeling period appear green.
Early studies recognized that the elevated level of myocardial renewal observed in injured hearts was largely limited to the region of injury, and that the remote myocardium was not markedly affected (summarized in [2]). Thus, experiments which examine only the infarct and/or a limited proximal region of the border zone will by default report higher renewal rates as compared to experiments which include more distal regions in the analyses. This in turn contributes to variation in the reported renewal rates in injured hearts. The timing of the analyses is also an important consideration, as experiments in rats clearly demonstrated that cell cycle activity reached maximal levels at two weeks following cryoinjury and then rapidly subsided [21]. The degree to which cell cycle activation (as evidenced by S-phase entry) culminates in cytokinesis is also subject to some debate. This is perhaps best illustrated by examples from infarcted human hearts, where markedly different conclusions were drawn based on the inclusion [22] or exclusion [23] of assays to monitor nuclear envelop breakdown. There are two interesting exceptions to the low rates of post-injury cardiomyocyte renewal. The Rumyantsev and Oberpriller laboratories clearly demonstrated atrial cardiomyocyte renewal following injury to the ventricular myocardium in rats [2, 24–27]. Furthermore, Rumyanstev clearly documented cell cycle induction in conduction system cells following myocardial injury (summarized in [2]). These finding have not been extensively pursued, presumably because amplification of working ventricular cardiomyocytes is the more relevant clinical target.
As indicated above, short pulse-chase experiments cannot detect stem-cell mediated cardiomyogenic events (unless the resulting cardiomyocytes retain cell cycle activity). In contrast, cumulative label incorporation assays have the potential to detect stem cell mediated events. Indeed, the notion that cardiomyogenic stem cells persist in the adult heart has received considerable interest over the past ten years. Ironically, the major studies supporting myocardial renewal via extrinsic [28] or cardiac-resident [29] stem cell-mediated cardiomyogenic activity (and which served as a major impetus in promoting the area of research) have proven to be difficult to reproduce, at least by some groups [30–33].
Perhaps the strongest data supporting the presence of cardiomyogenic stem cell activity in the adult heart was generated by the Lee laboratory. These studies used a genetic fate-mapping system comprised of a transgene encoding a conditional, tamoxifen-dependent Cre recombinase under the regulation of the cardiomyocyte-restricted myosin heavy chain promoter in conjunction with a ubiquitously-expressed reporter transgene [34]. Transient exposure to tamoxifen resulted in the generation of adult mice which expressed enhanced green fluorescent protein (EGFP) in terminally differentiated cardiomyocytes, and beta-galactosidase in all other cells. If stem cells undergo a cardiomyogenic event in these animals, it would give rise to beta-galactosidase expressing cardiomyocytes which could be identified in histologic sections. Using this approach, Lee and colleagues were able to demonstrate a progressive increase in beta-galactosidase expressing cardiomyocytes following myocardial infarction. Although these data provide strong evidence for cardiomyogenic stem cell activity, it should be noted that the conversion rate to activate EGFP reporter expression was only ca. 70%, with the remaining adult cardiomyocytes retaining beta-galactosidase activity. Consequently, any differential impact of EGFP vs. beta-galactosidase expression on cardiomyocyte proliferation or survival (i.e., apoptosis sensitivity) would impact interpretation of the data. Unfortunately, this incomplete conversion also renders the sensitivity of the model too low to detect a 1% annual cardiomyocyte renewal rate, as was reported above for non-injured adult mice.
The utility of genetic tracking systems to identify the origins of “new” cardiomyocytes in vivo ultimately is dependent upon the fidelity of the reporter systems. A priori, it seems obvious that the more complex the reporter system, the greater the potential for ambiguous results. For example, earlier studies talking advantage of differential folding rates (and consequently differential activation of epifluorescence) of cardiac-restricted EGFP and nuclear red fluorescent protein reporters suggested that zebrafish regeneration resulted via a stem cell-based mechanism [35]. In contrast, data from a more simple reporter system suggested cell cycle-based regeneration [36]; subsequent studies validated this as the main mechanism for myocardial renewal in zebrafish [37]. As indicated above, incomplete cre-mediated conversion in the bigenic mouse system used by Lee and colleagues complicated data interpretation. Moreover, the suggestion that the MHC promoter may be active in undifferentiated progenitor cells in some studies [38, 39] raises further concerns with this approach. Despite these concerns and caveats, genetic tracking systems as described above offer the best hope for establishing the molecular basis for myocardial renewal. The onus is on the investigators to employ the judicious use of controls to establish the limits of their reporter systems, and to restrict data interpretation within these limits.
Concluding thoughts
The presence of cardiomyocyte renewal in the adult heart has been recognized for over a century. The main disagreements in the field pertain to the level at which this occurs in the mammalian heart, and the underlying mechanism. It is hoped that this review provides some explanation for the basis of these disagreements. It is encouraging to note that as assays and reagents improve, a consensus with regards to the magnitude of renewal in normal and injured hearts is beginning to emerge. It is also likely that consensus will be reached regarding the underlying mechanism(s) for renewal. In that regard, it is sobering to note that the mechanism of post-injury myocardial renewal in zebrafish (which occurs at a very high level and is easily detected by histology) remained contested for many years.
Highlights.
It is well accepted that cardiomyocyte renewal occurs in the adult mammalian heart
The level of cardiomyocyte renewal is subject to debate
The mechanism of cardiomyocyte renewal is subject to debate
Issues contributing to these differences are discussed
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Zielonko J. Pathologische-anatomische und experimentelle studien uber hypertrophie des herzens. Virchows Arch Pathol Anat Physio, klin Med. 1875;62:29–57. [Google Scholar]
- 2.Rumyantsev PP. Growth and hyperplasia of cardiac muscle cells. Harwood Academic Publishers; London, U.K.; New York, N.Y., U.S.A: 1991. [Google Scholar]
- 3.Bader D, Oberpriller J. Autoradiographic and electron microscopic studies of minced cardiac muscle regeneration in the adult newt, notophthalmus viridescens. J Exp Zool. 1979;208:177–193. doi: 10.1002/jez.1402080206. [DOI] [PubMed] [Google Scholar]
- 4.Bader D, Oberpriller JO. Repair and reorganization of minced cardiac muscle in the adult newt (Notophthalmus viridescens) J Morphol. 1978;155:349–357. doi: 10.1002/jmor.1051550307. [DOI] [PubMed] [Google Scholar]
- 5.Oberpriller J, Oberpriller JC. Cell division in adult newt cardiac myocytes. In: Oberpriller J, Oberpriller JC, Maruro A, editors. The development and regenerative potential of cardiac muscle. Harwood Academic Publishers; Chur: 1991. pp. 293–312. [Google Scholar]
- 6.Oberpriller J, Oberpriller JC. Mitosis in adult newt ventricle. J Cell Biol. 1971;49:560–563. doi: 10.1083/jcb.49.2.560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Oberpriller JO, Oberpriller JC. Response of the adult newt ventricle to injury. J Exp Zool. 1974;187:249–253. doi: 10.1002/jez.1401870208. [DOI] [PubMed] [Google Scholar]
- 8.Poss KD, Wilson LG, Keating MT. Heart regeneration in zebrafish. Science. 2002;298:2188–2190. doi: 10.1126/science.1077857. [DOI] [PubMed] [Google Scholar]
- 9.Soonpaa MH, Field LJ. Survey of studies examining mammalian cardiomyocyte DNA synthesis. Circ Res. 1998;83:15–26. doi: 10.1161/01.res.83.1.15. [DOI] [PubMed] [Google Scholar]
- 10.Leri A, Kajstura J, Anversa P. Cardiac stem cells and mechanisms of myocardial regeneration. Physiol Rev. 2005;85:1373–1416. doi: 10.1152/physrev.00013.2005. [DOI] [PubMed] [Google Scholar]
- 11.Carter D. Practical considerations for collecting confocal images. Methods Mol Biol. 1999;122:35–57. doi: 10.1385/1-59259-722-x:35. [DOI] [PubMed] [Google Scholar]
- 12.Centonze VE, White JG. Multiphoton excitation provides optical sections from deeper within scattering specimens than confocal imaging. Biophys J. 1998;75:2015–2024. doi: 10.1016/S0006-3495(98)77643-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gulick J, Subramaniam A, Neumann J, Robbins J. Isolation and characterization of the mouse cardiac myosin heavy chain genes. J Biol Chem. 1991;266:9180–9185. [PubMed] [Google Scholar]
- 14.Soonpaa MH, Koh GY, Klug MG, Field LJ. Formation of nascent intercalated disks between grafted fetal cardiomyocytes and host myocardium. Science. 1994;264:98–101. doi: 10.1126/science.8140423. [DOI] [PubMed] [Google Scholar]
- 15.Ang KL, Shenje LT, Reuter S, Soonpaa MH, Rubart M, Field LJ, Galinanes M. Limitations of conventional approaches to identify myocyte nuclei in histologic sections of the heart. Am J Physiol Cell Physiol. 2010;298:C1603–1609. doi: 10.1152/ajpcell.00435.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Soonpaa MH, Field LJ. Assessment of cardiomyocyte DNA synthesis in normal and injured adult mouse hearts. Am J Physiol. 1997;272:H220–226. doi: 10.1152/ajpheart.1997.272.1.H220. [DOI] [PubMed] [Google Scholar]
- 17.Bergmann O, Bhardwaj RD, Bernard S, Zdunek S, Barnabe-Heider F, Walsh S, Zupicich J, Alkass K, Buchholz BA, Druid H, Jovinge S, Frisen J. Evidence for cardiomyocyte renewal in humans. Science. 2009;324:98–102. doi: 10.1126/science.1164680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bai SL, Campbell SE, Moore JA, Morales MC, Gerdes AM. Influence of age, growth, and sex on cardiac myocyte size and number in rats. The Anatomical record. 1990;226:207–212. doi: 10.1002/ar.1092260210. [DOI] [PubMed] [Google Scholar]
- 19.Porrello ER, Mahmoud AI, Simpson E, Hill JA, Richardson JA, Olson EN, Sadek HA. Transient regenerative potential of the neonatal mouse heart. Science. 2011;331:1078–1080. doi: 10.1126/science.1200708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Soonpaa MH, Kim KK, Pajak L, Franklin M, Field LJ. Cardiomyocyte DNA synthesis and binucleation during murine development. Am J Physiol. 1996;271:H2183–2189. doi: 10.1152/ajpheart.1996.271.5.H2183. [DOI] [PubMed] [Google Scholar]
- 21.van Amerongen MJ, Harmsen MC, Petersen AH, Popa ER, van Luyn MJ. Cryoinjury: a model of myocardial regeneration. Cardiovasc Pathol. 2008;17:23–31. doi: 10.1016/j.carpath.2007.03.002. [DOI] [PubMed] [Google Scholar]
- 22.Meckert PC, Rivello HG, Vigliano C, Gonzalez P, Favaloro R, Laguens R. Endomitosis and polyploidization of myocardial cells in the periphery of human acute myocardial infarction. Cardiovasc Res. 2005;67:116–123. doi: 10.1016/j.cardiores.2005.02.017. [DOI] [PubMed] [Google Scholar]
- 23.Leads from the MMWR. Premature mortality due to congenital anomalies--United States. Journal of the American Medical Association. 1988;260:1685. [PubMed] [Google Scholar]
- 24.Oberpriller JO, Ferrans VJ, Carroll RJ. Changes in DNA content number of nuclei and cellular dimensions of young rat atrial myocytes in response to left coronary artery ligation. J Mol Cell Cardiol. 1983;15:31–42. doi: 10.1016/0022-2828(83)90305-x. [DOI] [PubMed] [Google Scholar]
- 25.Oberpriller JO, Ferrans VJ, Carroll RJ. DNA synthesis in rat atrial myocytes as a response to left ventricular infarction. An autoradiographic study of enzymatically dissociated myocytes. J Mol Cell Cardiol. 1984;16:1119–1126. doi: 10.1016/s0022-2828(84)80038-3. [DOI] [PubMed] [Google Scholar]
- 26.Oberpriller JO, Oberpriller JC, Aafedt BC. Changes in binucleation and cellular dimensions of rat left atrial myocytes after induced left ventricular infarction. The American journal of anatomy. 1987;179:285–290. doi: 10.1002/aja.1001790310. [DOI] [PubMed] [Google Scholar]
- 27.Rumyantsev PP. Interrelations of the proliferation and differentiation processes during cardiact myogenesis and regeneration. Int Rev Cytol. 1977;51:186–273. [PubMed] [Google Scholar]
- 28.Orlic D, Kajstura J, Chimenti S, Jakoniuk I, Anderson SM, Li B, Pickel J, McKay R, Nadal-Ginard B, Bodine DM, Leri A, Anversa P. Bone marrow cells regenerate infarcted myocardium. Nature. 2001;410:701–705. doi: 10.1038/35070587. [DOI] [PubMed] [Google Scholar]
- 29.Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, Kasahara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J, Nadal-Ginard B, Anversa P. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell. 2003;114:763–776. doi: 10.1016/s0092-8674(03)00687-1. [DOI] [PubMed] [Google Scholar]
- 30.Balsam LB, Wagers AJ, Christensen JL, Kofidis T, Weissman IL, Robbins RC. Haematopoietic stem cells adopt mature haematopoietic fates in ischaemic myocardium. Nature. 2004;428:668–673. doi: 10.1038/nature02460. [DOI] [PubMed] [Google Scholar]
- 31.Murry CE, Soonpaa MH, Reinecke H, Nakajima H, Nakajima HO, Rubart M, Pasumarthi KB, Virag JI, Bartelmez SH, Poppa V, Bradford G, Dowell JD, Williams DA, Field LJ. Haematopoietic stem cells do not transdifferentiate into cardiac myocytes in myocardial infarcts. Nature. 2004;428:664–668. doi: 10.1038/nature02446. [DOI] [PubMed] [Google Scholar]
- 32.Nygren JM, Jovinge S, Breitbach M, Sawen P, Roll W, Hescheler J, Taneera J, Fleischmann BK, Jacobsen SE. Bone marrow-derived hematopoietic cells generate cardiomyocytes at a low frequency through cell fusion, but not transdifferentiation. Nat Med. 2004;10:494–501. doi: 10.1038/nm1040. [DOI] [PubMed] [Google Scholar]
- 33.Zaruba MM, Soonpaa M, Reuter S, Field LJ. Cardiomyogenic potential of C-kit(+)-expressing cells derived from neonatal and adult mouse hearts. Circulation. 2010;121:1992–2000. doi: 10.1161/CIRCULATIONAHA.109.909093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Hsieh PC, Segers VF, Davis ME, MacGillivray C, Gannon J, Molkentin JD, Robbins J, Lee RT. Evidence from a genetic fate-mapping study that stem cells refresh adult mammalian cardiomyocytes after injury. Nat Med. 2007;13:970–974. doi: 10.1038/nm1618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lepilina A, Coon AN, Kikuchi K, Holdway JE, Roberts RW, Burns CG, Poss KD. A dynamic epicardial injury response supports progenitor cell activity during zebrafish heart regeneration. Cell. 2006;127:607–619. doi: 10.1016/j.cell.2006.08.052. [DOI] [PubMed] [Google Scholar]
- 36.Raya A, Koth CM, Buscher D, Kawakami Y, Itoh T, Raya RM, Sternik G, Tsai HJ, Rodriguez-Esteban C, Izpisua-Belmonte JC. Activation of Notch signaling pathway precedes heart regeneration in zebrafish. Proc Natl Acad Sci U S A. 2003;100(Suppl 1):11889–11895. doi: 10.1073/pnas.1834204100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kikuchi K, Holdway JE, Werdich AA, Anderson RM, Fang Y, Egnaczyk GF, Evans T, Macrae CA, Stainier DY, Poss KD. Primary contribution to zebrafish heart regeneration by gata4(+) cardiomyocytes. Nature. 2010;464:601–605. doi: 10.1038/nature08804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zelarayan LC, Noack C, Sekkali B, Kmecova J, Gehrke C, Renger A, Zafiriou MP, van der Nagel R, Dietz R, de Windt LJ, Balligand JL, Bergmann MW. Beta-Catenin downregulation attenuates ischemic cardiac remodeling through enhanced resident precursor cell differentiation. Proc Natl Acad Sci U S A. 2008;105:19762–19767. doi: 10.1073/pnas.0808393105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bailey B, Izarra A, Alvarez R, Fischer KM, Cottage CT, Quijada P, Diez-Juan A, Sussman MA. Cardiac stem cell genetic engineering using the alphaMHC promoter. Regenerative medicine. 2009;4:823–833. doi: 10.2217/rme.09.51. [DOI] [PMC free article] [PubMed] [Google Scholar]