

Abstract
Hydroethidine (or dihydroethidium) (HE) is the most popular fluorogenic probe used for detecting intracellular superoxide radical anion. The reaction between superoxide and HE generates a highly specific red fluorescent product, 2-hydroxyethidium (2-OH-E+). In biological systems, another red fluorescent product, ethidium (E+), is also formed, usually at a much higher concentration than 2-OH-E+. In this article, we have reviewed the methods to selectively detect the superoxide-specific product (2-OH-E+) and the factors affecting its levels in cellular and biological systems. The most important conclusion of the present review is that it is nearly impossible to assess the intracellular levels of the superoxide specific product, 2-OH-E+, using the confocal microscopy or other fluorescence-based microscopic assays and that it is essential to measure by HPLC the intracellular HE and other oxidation products of HE, in addition to 2-OH-E+, in order to fully understand the origin of red fluorescence. The chemical reactivity of mitochondria-targeted hydroethidine (Mito-HE, MitoSOX Red ®) with superoxide is similar to the reactivity of HE with superoxide and therefore, all of the limitations attributed to the HE assay are applicable to Mito-HE (or Mito-SOX) as well.
Introduction
The discovery of the enzyme superoxide dismutase (SOD) has revolutionized our understanding of the role of reactive oxygen species (ROS) in biology and medicine (1-5). Once viewed as a deleterious byproduct of oxygen metabolism, superoxide radical anion (O2•-) is now considered to play a role in modulating signaling by reacting with •NO, a second messenger and as a precursor for H2O2, another second messenger. Despite tremendous progress in the understanding of the biological reactions and physiological signaling functions of superoxide, intracellular detection and quantitation of O2•- remains a challenge. Direct detection of intracellular superoxide is virtually impossible due to its short half-life and rapid intracellular scavenging. This has led to the development of many probes (e.g., luminol, nitro blue tetrazolium, hydroethidine) which can react with O2•-, producing easily detectable, relatively stable products that accumulate with time [6-10]. However, only a few of those probes yield products that are specific for superoxide. These probes include: lucigenin (a chemiluminescent probe, which forms N-methylacridone in excited state via an unstable dioxetane), hydroethidine (which forms a fluorescent product - 2-hydroxyethidium) and cyclic nitrone spin traps (which form spin adducts exhibiting characteristic spectra of superoxide spin adducts detectable by electron paramagnetic resonance (EPR) spectrometry). As discussed in previous reviews, lucigenin undergoes redox-cycling (self-generating O2•-) [11-13] and nitrone spin traps react slowly with superoxide, forming a relatively unstable superoxide adduct [14-16]; consequently, hydroethidine (HE, also known as dihydroethidium (DHE), Figure 1) has become a probe of choice for the detection of intracellular superoxide. The method based on the detection of 2-hydroxyethidium was described as a “gold standard” for superoxide detection in cardiovascular research [7].
Figure 1.
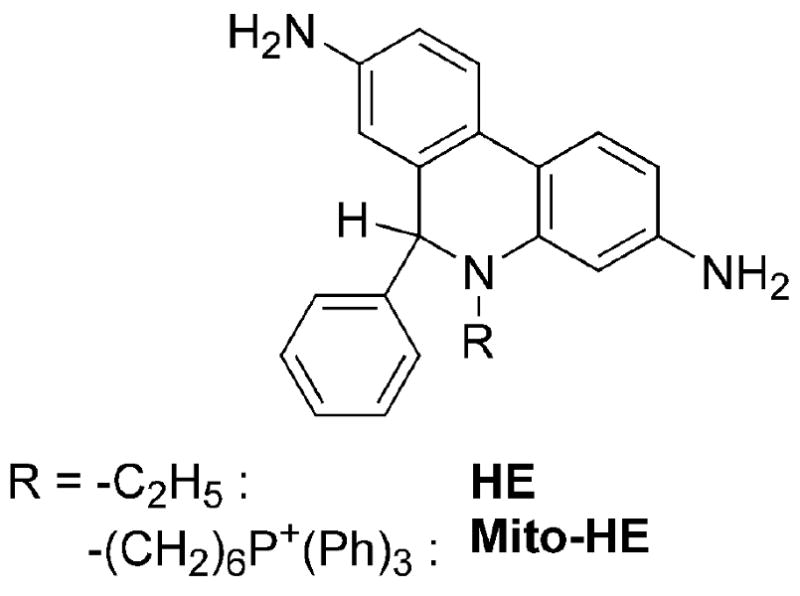
Structure of HE and Mito-HE.
Over the last 20 years, hydroethidine has been used to detect superoxide produced in a variety of biological systems, ranging from intracellular organelles, whole cells to whole organs in live animals. Hydroethidine and its mitochondria-targeted analog (Mito-HE or Mito-SOX) have been used to detect O2•- formed during oxidative burst in leukocytes, pro- and antioxidative action of cellular endogenous compounds and xenobiotics, during apoptosis and anti- and pro-apoptotic action of drugs, neurosignaling, neurodegeneration and neuroprotection, ischemia and reperfusion, hypertension, mechanical stress induced oxidant production, vascular signaling and pathology, renal function and pathology, mitochondrial and radiation-induced damage. These studies provide evidence for the involvement of superoxide in many physiological and pathophysiological conditions and contribute to the understanding of the molecular mechanisms of many diseases, potentially allowing for development of new strategies for treatment. In almost all studies, the “red fluorescence” generated from HE, attributed to DNA-bound ethidium, was used as a marker of superoxide formation. In this review, we discuss the reactions of HE with superoxide and other oxidants, some of which generate oxidative red fluorescent product (different from that of the superoxide and HE reaction product) and other oxidative non-fluorescent dimeric products. In addition, we emphasize the need to monitor the levels of HE and its oxidation products in order to correctly interpret the 2-hydroxyethidium data obtained from HPLC-based studies.
Evolution of HE from a vital dye to superoxide probe
HE – product of reduction of E+
Hydroethidine, a product of two-electron reduction of ethidium cation (E+, Figure 2), was initially synthesized by reacting ethidium bromide with sodium borohydride [17]. HE does not avidly bind to DNA as it lacks a positive charge and has a non-planar geometry of the phenanthridine moiety. Reduction of E+ to HE was proposed as a way to release E+ from DNA [17]. The ultraviolet-visible (UV-Vis) absorption spectrum of an aqueous solution of HE at neutral pH consists of several absorption bands in the range of 200 – 400 nm with the maxima at 224, 265 and 345 nm (Figure 3A), which can be utilized for quantitative analysis of the probe in solutions [17,18].
Figure 2.
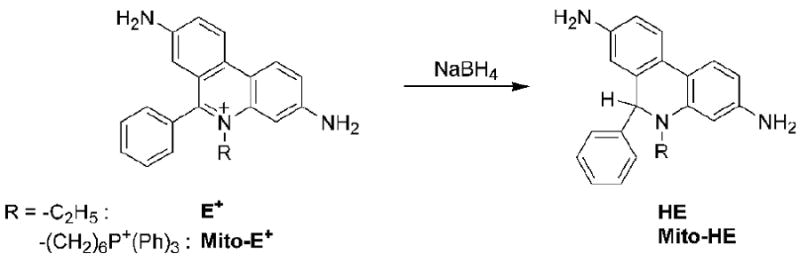
Reduction of E+ and Mito-E+ by NaBH4.
Figure 3.
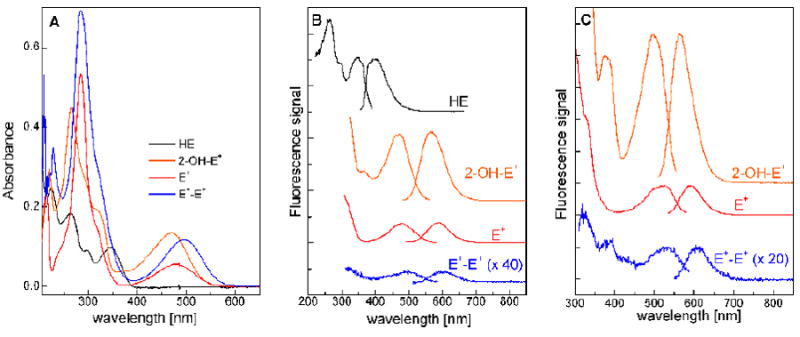
Spectroscopic properties of HE and its oxidation products. (A) UV-Vis absorption spectra; (B) Fluorescence spectra; (C) Fluorescence spectra in the presence of DNA.
HE as a vital dye
HE has been initially used as a vital dye because it is cell-permeable [19-23]. The “blue fluorescence” after incubation of live cells with HE was observed predominantly in the cytoplasm of various tumor cells [19]. Depending on the cell line, HE was distributed uniformly over the cytoplasm or was localized in small spots of bluish-white fluorescence. Moreover, the extent of HE uptake and accumulation may be dependent not only on the dye concentration and incubation time, but also on the metabolic state of the cells [19]. HE was used in the investigation of antiproliferative compounds [24-26] and in other biological studies as a vital stain [27-31].
HE as a probe for cellular redox status
The “blue fluorescence” detected in cells following incubation with HE was attributed to the unreacted HE whereas the “red fluorescence” in cells was attributed to the oxidized form of HE, namely the ethidium cation that was bound to the nuclear DNA [19,26]. Because of its charge, the ethidium cation binds to DNA and remains trapped inside the cells [19,22,32]. The “red fluorescence” staining of the cells observed upon incubation with HE [33-39] was used to follow the effector-target interaction [21,40-43] and to visualize the DNA structures in intact cells [44]. However, it was also shown that E+ was rapidly cleared by doxorubicin-resistant fibrosarcoma cells [20]. The extent of the conversion of HE into E+ was attributed to the redox state of the cells. It was proposed that HE oxidation is dependent upon the availability of NADP+ coenzyme and can be used to distinguish between the hypoxic and normoxic cells [45]. It was also shown that the intensity of “red fluorescence” of the cells is dependent on HE uptake and cell oxygenation [45]. An additional factor affecting the rate of intracellular oxidation of HE to E+ was the cell type. For example, the extent of HE oxidation was greater in monocytes as compared to neutrophils [46].
HE as a probe for intracellular ROS
The intracellular oxidation of HE to E+ was used to visualize the reactive oxygen species (ROS) generated upon stimulation of macrophages and monocytes [25,47-49]. Incubation of the cells with xanthine and xanthine oxidase (generating both superoxide and hydrogen peroxide) and HE caused an increase in the fluorescence intensity that was inhibitable by SOD and catalase. It was suggested that HE can react both with superoxide and H2O2 [25,47,48]. Reports also indicate that HE can be oxidized by other oxidants, including ONOO- and the Fenton's reagent, forming a fluorescent product, suggesting that HE is not specific for detecting the superoxide radical anion [50]. These reports concluded that HE is a non-specific probe for ROS [51,52]. However, further research, as outlined below, indicated that this appears not to be the case.
HE is a specific probe for superoxide
Several reports indicated that HE does not react readily with H2O2 and was oxidized intracellularly to E+, most likely by superoxide [46,53,54]. Support for this conclusion came from experiments using the SOD linked to polyethylene glycol (PEG-SOD) [55] or high concentrations of extracellularly added SOD [56]. The increase in the HE-derived fluorescence observed after treatment of the cells with H2O2 was attributed to intracellular stimulation of superoxide production by H2O2 [53,54,57-59]. Based on these results, the HE-derived red fluorescence became widely used for the detection and quantitation of intracellularly generated superoxide [60-68].
2-Hydroxyethidium – the product of the reaction between HE and superoxide
We first reported that the reaction between HE and superoxide formed a product that was structurally different from the ethidium cation, E+ [69,70]. The mass spectrometric analysis indicated that an oxygen atom was incorporated into ethidium cation. Results also showed that this particular product was unique to superoxide and that other oxidants (peroxynitrite, hydrogen peroxide, and hydroxyl radical) did not form the same product upon reaction with HE. Soon after this discovery, other investigators confirmed the formation of the same product in cells, as determined by high performance liquid chromatography (HPLC) analysis of the extracts of cells and blood vessels incubated with HE [71,72]. Detailed nuclear magnetic resonance (NMR) and mass spectral (MS) analyses of the product indicated that an oxygen atom was attached at the carbon atom position 2 of the phenanthridine ring resulting in the formation of the product, 2-hydroxyethidium (2-OH-E+, Figure 4) [72,73]. Several studies have since confirmed that 2-OH-E+ is the only product of the reaction of HE with superoxide generated by xanthine/xanthine oxidase [74-76].
Figure 4.
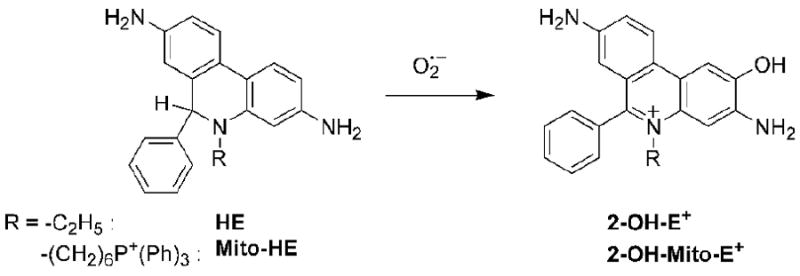
Formation of 2-hydroxyethidium.
Targeting HE to mitochondria
In order to detect superoxide generated in the mitochondria, the chemical structure was modified by conjugating HE to the triphenylphosphonium moiety (Fig. 1). The attachment of the triphenylphosphonium moiety to HE increased the accumulation of the compound (Mito-HE, Figure 1) into mitochondria [77]. The reaction between Mito-HE and superoxide yielded the corresponding hydroxylated ethidium analog (hydroxy-Mito-ethidium), similar to that formed from the reaction between HE and superoxide [73,77]. Based on NMR analyses, two different structures were proposed: hydroxyl group attachment at C-9 position [77] or at C-2 position [73]. We favor the structure with the hydroxyl group attached at C-2 position (as shown in Figure 4) [73].
HE toxicity
HE was shown to be cytotoxic to protoplast at relatively higher concentrations (45 μM), and the toxicity was increased in cells exposed to the UV light [78]. An inverse correlation between HE uptake and cell survival was observed in intact cells [45]. Intravenous injection of HE, however, protected against the injury caused by transient focal cerebral ischemia, which was attributed to scavenging of superoxide by HE [79].
Reaction between HE and oxidants
From reviewing the previous literature, it is evident that despite its extensive use in biological systems, the basic chemical reactivity of HE remains poorly understood. Thus, we feel it is necessary to present a short overview of the state-of-the-art of HE oxidation/reduction chemistry.
Two-electron oxidation to E+
HE can be readily converted to the ethidium cation by transferring the hydride anion (one proton and two electrons) to a suitable acceptor. In this regard, HE resembles the reduced form of the nicotinamide adenine dinucleotide (NADH). In fact, both HE and NADH can be converted to their oxidized forms (E+ and NAD+) by reacting with chloranil (tetrachloro-p-benzoquinone) [80]. Treatment of the cells with chloranil converted HE extensively into E+; however, it is not clear whether the conversion is solely non-enzymatic, or whether any cellular enzyme is involved in this conversion [81]. In addition, the one-electron oxidation of HE also leads to the formation of the ethidium cation (E+) along with other (dimeric) products under these conditions (see below) [73]. The proposed mechanism involves a disproportionation reaction of the radical intermediate (HE•+ or HE(NH•)), along with the formation of the 2-electron oxidation product (E+).
Superoxide radical anion
The reactivity of HE with superoxide was initially deduced based on the inhibitory effect of SOD in potassium superoxide (KO2) –induced oxidation of HE [46,53,82] and on the increased fluorescence intensity derived from HE incorporated into mitochondria in the presence of xanthine (X) and xanthine oxidase (XO) [47]. Many reports have since confirmed this finding [53,69,83-94]. The decrease in the fluorescence intensity observed in the presence of higher concentrations of KO2 was attributed to the reaction between E+ and O2•- [53]. However, as neither E+ nor 2-OH-E+ reacts with O2•- [71], the loss of fluorescence may be due to the reaction with the species other than O2•-. Based on the fluorescence kinetics changes (monitored after mixing of HE with KO2), formation of a transient fluorescent intermediate was also proposed [95].
In order to quantify the amount of superoxide trapped based on the yield of the HE-superoxide reaction product, the stoichiometry of this reaction needs to be determined. By analogy with the HE/Fremy's salt reaction, the stoichiometry for HE/O2•- reaction was postulated to be 1:2, i.e. two molecules of superoxide are consumed per one molecule of hydroethidine to form the fluorescent product [18]. However, this did not turn out to be the case using the X/XO system. It was suggested that HE may cause the dismutation of superoxide anion, thus lowering the yield of the oxidation product detected in the X/XO system [83]. The amount of 2-OH-E+ formed during oxidation of HE in X/XO system was nearly 3.6-fold lower as compared to superoxide generation [71]. In other studies, a 1:1 stoichiometry for HE reaction with O2•- was suggested at lower fluxes of superoxide [84,96,97]. The reported stoichiometry of 1:60 for HE/O2•- reaction in dimethyl sulfoxide (DMSO) [98] is also puzzling. Clearly, we still do not exactly know the mechanism and stoichiometry of 2-OH-E+ formation and additional investigation is necessary for establishing the exact mechanism and accurate stoichiometry of the HE/O2•- reaction.
Synthesis of 2-OH-E+: Reaction between nitrosodisulfonate radical dianion (Fremy's salt) and HE
We showed that HE reacts with nitrosodisulfonate radical dianion (NDS, a hydrolysis product of Fremy's salt) to form the 2-hydroxyethidium [18]. This reaction pathway is a facile route to synthesizing the authentic standard of 2-OH-E+. The stoichiometry of the reaction (leading to 2-OH-E+ formation) is 1:2 (HE:NDS). Based on the determined kinetic isotope effect (kH/kD) of 1.9, we proposed the hydrogen (deuterium) atom abstraction from one of the aromatic amine groups by NDS as the first step of the reaction. This is analogous to the initial hydrogen abstraction reaction between HE and O2•- forming a transient aromatic aminyl radical of HE, followed by the formation of the hydroperoxide and quinone imine forms of HE (Figure 5) [18]. Although the HE-derived radical was implicated in the reaction of HE with NDS, no effect of oxygen on the product yield was observed. Thus, it has been concluded that the radical formed is unreactive towards oxygen, in contrast to the radical species formed from other widely used fluorogenic probes, including dichlorodihydrofluorescein and dihydrorhodamine.
Figure 5.
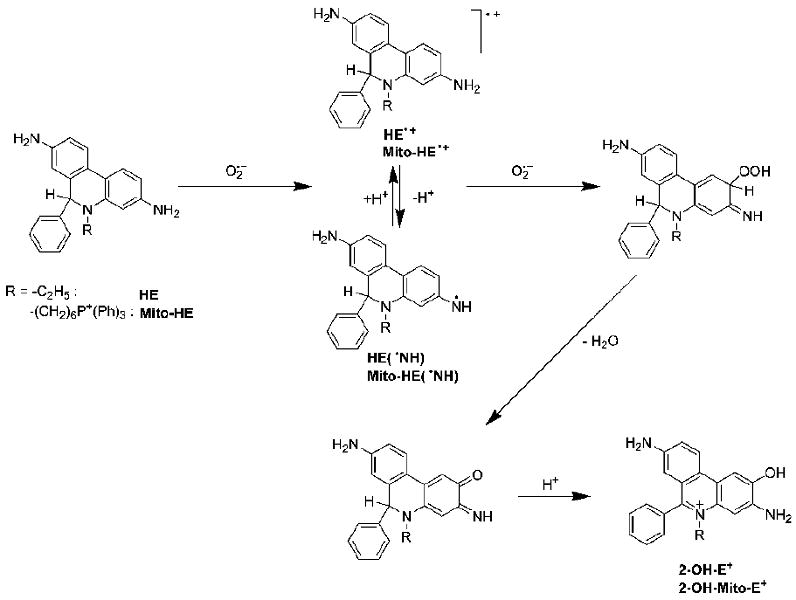
Hypothetical mechanism of the reaction of HE with superoxide radical anion.
Reaction between manganese porphyrin complexes and HE
It was reported that Mn(III) tetrakis(1-methyl-4-pyridyl)porphyrin pentachloride could quench the “DHE fluorescence” [81]. This suggests that the manganese-porphyrin complex can react with HE in the ground and/or the excited state. It was also reported that HE can react with another manganese-porphyrin complex, namely the manganese(III) tetrakis(4-benzoic acid)porphyrin (MnTBAP) [76]. The HPLC analysis of the reaction product(s) revealed the formation of E+. Mn(III)TBAP treatment was also shown to cause the depletion of HE in cells [76]. Thus, the decrease in the amount of superoxide-specific product may be due to lower intracellular availability of the probe. This is particularly worrisome as some porphyrin complexes are used as SOD mimetics to verify the intermediacy of intracellular superoxide using the HE probe.
Oxidation of HE by ferricyanide anion
It has been reported that HE reacts with ferricyanide anion ([Fe(CN)6]3-) leading to the formation of product(s) with fluorescence properties similar to those of ethidium, although the fluorescence intensity was significantly lower [87]. This report also suggests that the stoichiometry of the reaction is 1:1, irrespective of the molar ratio between the reagents, and that a single product is formed. That observation led to the conclusion that a dimer was formed via the recombination of radicals formed from the one-electron oxidation of HE [87]. The HPLC analysis of the reaction products, however, indicated that several products are formed including E+ and the dimeric products of different redox state (HE-HE, HE-E+, E+-E+, Figure 6) [73]. Ferricyanide-mediated oxidation of Mito-HE resulted in the formation of Mito-E+ and the corresponding dimeric products. The reaction of HE (or Mito-HE) with ferricyanide has been proposed as a facile route for synthesis of the diethidium (E+-E+ or Mito-E+-Mito-E+) standard [99]. Measurement of dimeric productsof HE can be used as a diagnostic quantitative marker for oxidant formation in cells.
Figure 6.
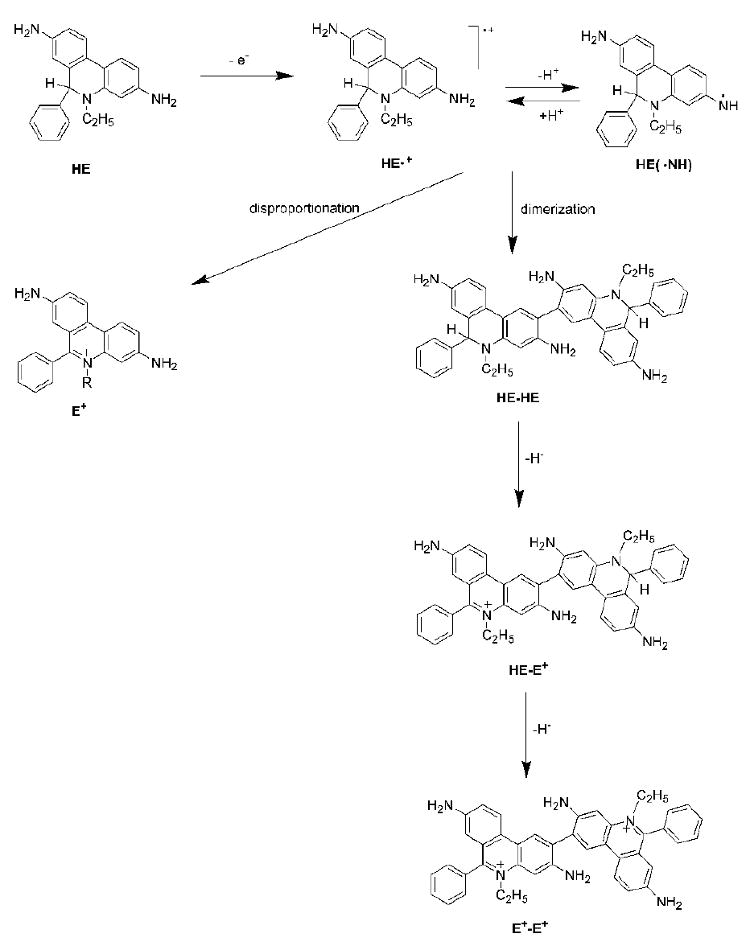
Transformations of HE upon one-electron oxidation.
Reaction between HE and H2O2 and organic peroxides?
Most previous reports indicate that HE does not react readily with hydrogen peroxide (H2O2) in cell-free solution [46,50,53,54,74,82,83,93]. Yet it has been reported that organic peroxide, tert-butyl hydroperoxide was able to oxidize HE with the formation of a fluorescent product [50]. However, this reaction appears to be highly unlikely in the absence of redox active metal ions or other catalyst. The presence of low-molecular weight complexes of iron and/or heme proteins (e.g. cytochromes) or in the cellular systems, may facilitate the reaction of HE with peroxides, as discussed below.
Cytochromes
Reports indicate that ferricytochrome c (cyt c3+) can react with HE to form the ferrocytochrome c (cyt c2+) [83]. As one molecule of HE could cause the reduction of up to 4 molecules of cyt c3+ it was suggested that in addition to E+, the dimeric products are most likely formed [83]. Moreover, the kinetics of this reaction is biphasic, indicating a complex reaction mechanism [73,83,87]. Depending on the molar ratio between HE and cyt c3+, different products were formed as monitored by the UV-Vis absorption and fluorescence techniques [87]. Also, no oxygen consumption during the reaction between cyt c3+ and HE was observed [87]. Thus, one can conclude that the reaction intermediate(s) and products formed during this reaction are inert towards molecular oxygen. HPLC analysis of the reaction mixtures containing HE and cyt c3+ revealed the formation of several products including E+, E+-E+ (diethidium) and two additional products assigned to the dimers, HE-HE and HE-E+ (Figure 6) [73]. Addition of cyt c3+ also catalyzed the reaction between HE and H2O2 giving rise to the same products. The reaction of Mito-HE with cyt c3+ results in the formation of Mito-E+ as well as the dimeric products [73]. The rate constant for the reaction of HE and Mito-HE with cyt c3+ was estimated as 104 – 105 M-1s-1, indicating the likelihood of a rapid consumption of these probes in the presence of excess cyt c3+ [73]. This type of reactivity of HE is not limited to cytochrome c, and other cytochromes present in mitochondria could cause the oxidation of HE as well [87]. Incubation of HE and Mito-HE with mitochondria yielded the same products (albeit in different ratios) as detected in the case of cyt c3+ [73]. Depletion of mitochondrial cytochrome c caused an increase of the probes' stability, suggesting that cytochrome c is largely responsible for the oxidation of HE or Mito-HE in the mitochondria.
HE oxidation by heme proteins
HE can be oxidized by horseradish peroxidase (HRP) and H2O2 with the formation of E+ [46,74,86] along with non-fluorescent products [46]. Other studies showed that the product(s) of HE oxidation had absorption spectral characteristics similar to those of E+ [88]. The stoichiometry of the reaction was determined to be 1:1 (i.e., 1 mol of HE oxidized per 1 mol of H2O2) [88].
Both hemoglobin and myoglobin react with HE to form the fluorescent products [87]. In both cases, a mixture of oxidation products was detected, similar to those formed during the reaction between HE and cytochrome c. In contrast to cytochrome c, methemoglobin and metmyoglobin-dependent oxidation of HE required the presence of oxygen [87].
Fenton's reagent
Several reports showed that HE can be oxidized by H2O2 in the presence of iron or copper [50,86] via the hydroxyl radical (•OH) or higher oxidants of iron [50]. However, in most of these studies the stock solution of HE was prepared in DMSO or in dimethylacetamide. Thus, the actual oxidants could be the solvent-derived radicals, as both DMSO and dimethylacetamide could scavenge •OH radicals. Moreover the HPLC analysis of the products of the incubation of HE with Fe2+/EDTA/H2O2 revealed the formation of both 2-OH-E+ and E+ [74]. Recently, it has been reported that the formation of 2-OH-E+ during oxidation of HE by Fenton's reagent is SOD-inhibitable, and so the formation of 2-OH-E+ in this system was attributed to superoxide formation [80]. The results show that the metal chelator-derived radical formed from the oxidizing species generated in the Fenton reaction could form the superoxide anion. The addition of ethanol or formate anion to the Fenton's system also increased the yield of 2-OH-E+ via superoxide formation [80]. Thus, the notion that hydroxyl radical or other oxidants derived from the Fenton system can form 2-OH-E+ from HE without the intermediacy of superoxide has not been substantiated.
Hydroxyl radical
Previous reports showed that •OH radical does not oxidize HE to a fluorescent product [53]. However, other studies showed the formation of a fluorescent product during irradiation of HE solutions, which has been attributed to the product of reaction of HE with hydroxyl radical [93,96]. It was suggested, based on the fluorescence spectral characteristics, that 2-OH-E+ was one of the products of the reaction of HE with hydroxyl radical [100]. However, without the HPLC analysis of the products formed, these conclusions remain speculative. Direct monitoring of the reaction of HE with hydroxyl radical by pulse radiolysis indicated the formation of HE radical cation (see below), and the rate constant value of 7 × 109 M-1s-1 was determined for this reaction [80].
Nitrogen oxides
Published reports consistently showed that •NO does not react with HE [53,86,94]. However, when HE was incubated with •NO in the presence of oxygen, a fluorescent product was detected [86]. In addition, it was also shown that the nitrogen dioxide radical (•NO2) could oxidize HE to a fluorescent product [86]. Structures of the products derived from HE were not, however, determined.
Peroxynitrite
Early on, it was reported that •NO or ONOO- was unreactive towards HE. This conclusion was based on the lack of increase of the ethidium fluorescence [53]. It was then postulated that •NO can compete with HE for O2•- thus inhibiting oxidation of HE [53,69,91,101]. However, it was also shown that incubation of SIN-1 (that slowly releases ONOO- in the presence of oxygen) or a bolus amount of ONOO- with HE led to the formation of a fluorescent product [50,86,91], and that reaction can be inhibited by the addition of glutathione [91]. Product(s) formed from the reaction between HE and peroxynitrite were totally different from 2-OH-E+ [69]. In another study it was postulated that, whereas peroxynitrite alone does not oxidize HE to 2-OH-E+ nor E+, the presence of CO2 stimulated oxidation of HE by ONOO- [74]. Moreover, the authors reported that, while no fluorescent product was formed during the reaction of HE with ONOO-, a product absorbing at 430 nm was detected [74]. These studies clearly indicate that in the presence of ONOO-, HE is oxidized to the products different from 2-OH-E+.
Hypochlorous acid
It has been reported that HE undergoes oxidation by HOCl to form a product with fluorescence characteristics similar to that of the ethidium cation [86,96]. Another study indicated that several products of similar spectral (UV-Vis absorption and fluorescence) characteristics were formed during HOCl-mediated oxidation of HE [88].
Photooxidation
In a cell-free system, HE was relatively stable, when exposed to an excitation light corresponding to that of ethidium (at 510 nm) [53]. On the other hand, when irradiated with UV light, HE undergoes oxidation yielding fluorescent product(s) [53,102]. The HPLC analysis of the products of HE photolysis revealed the formation of 2-OH-E+ and, with continued irradiation with the visible light, E+ was also formed [76]. The photooxidation of HE was shown to be oxygen-dependent, as after deoxygenation of the solution the process was inhibited [76]. It was also reported that the exposure of acetonitrile cell extracts to ambient light caused an increase in the levels of 2-OH-E+ [74].
We have previously shown that in the presence of 2-OH-E+, HE can undergo oxidation under the exposure to visible light, with the formation of E+ as a final product [76]. The extent of HE oxidation was proportional to the concentration of 2-OH-E+, and the photo-sensitizing effect of 2-OH-E+ was abrogated by deoxygenation of the reaction mixture, suggesting the involvement of molecular oxygen in the reaction. However, azide, a well-known scavenger of singlet oxygen, did not inhibit HE photooxidation, thus excluding the involvement of the singlet oxygen in the reaction [76].
Singlet oxygen
Although previous reports indicate that HE does not react with singlet oxygen (1O2) [53,96], a more recent report suggests that HE scavenges 1O2, with a rate constant of 1.1 × 109 M-1s-1 in acetonitrile [103]. The product analysis of the reaction between HE and singlet oxygen has not been, however, reported.
Ultrasound
An increase in HE-derived fluorescence intensity was observed after a shock wave treatment of cell suspension or a cell-free system containing HE [104,105]. It was shown that the increase in HE-derived fluorescence after treating cells with shock waves correlates directly with the cell membrane damage and increased membrane permeability, thus confounding the interpretation of the results of the flow cytometry measurements [106]. The sonication of HE aqueous solution causes a rapid consumption of HE along with the formation of 2-OH-E+ and probably other products [76]. The formation of 2-OH-E+ has been interpreted in terms of superoxide formation in the solution during sonication, as the addition of SOD inhibited the formation of 2-OH-E+ [76]. It was, however, reported that the sonication of the cell extract in acetonitrile did not cause formation of E+ or 2-OH-E+ [74]. These findings can be rationalized by the protective effects of the cell components present in the extract and/or the different sonochemistry of acetonitrile as compared to aqueous solution. Nevertheless, care should be taken when exposing HE to ultrasound [107-113].
Other reactions
The six-membered nitroxide, Tempol (4-Hydroxy-2,2,6,6-tetramethylpiperidine 1-oxyl), was shown to increase the HE-derived fluorescence in cells, when both compounds were co-incubated [114]. This indicates the possibility of a direct reaction between the two compounds. The acidity of the solution may affect HE stability. Reports indicate an increase in HE stability due to protonation or enhanced oxidation of HE, as has been reported for trichloroacetic acid [18, 76,87].
Intermediates formed during oxidation of HE
The intermediate radical formed during the one-electron oxidation of HE has been characterized spectroscopically and kinetically using the pulse radiolysis technique [80]. One electron oxidation of HE forms a radical cation (HE•+) with the absorption spectral maxima at 455 nm and 700 nm. It has been shown that the hydroxyl radical can quantitatively oxidize HE to the radical cation, HE•+. During the reaction between HE and superoxide, HE•+ is presumably formed which reacts rapidly with another superoxide molecule (k ≈ 2 × 109 M-1s-1). This reaction is responsible for the specific formation of 2-OH-E+ during HE reaction with superoxide. During the reaction between HE and nitrosodisulfonate radical dianion, another intermediate product, with the absorption maximum at 460 nm and 670 nm, has been detected and assigned tentatively to the corresponding iminoquinone form of oxidized hydroethidine [18].
Physico-chemical analyses of HE-derived products
Acid-base properties
At neutral pH, the ethidium cation exists as a single, positively charged species. However, in acidic solution, it undergoes double protonation with the pKa values of 0.4-0.8 and 2.0-2.1 for the protonation of aromatic amine groups at positions 3 and 8, respectively [115,116]. In the case of 2-OH-E+, an additional acid-base equilibrium is expected – with the involvement of the aromatic hydroxyl group [18,116]. In fact, the pKa value of 7.3, next to 0.5 and 2.2, has been reported for this cation [116]. This indicates that at pH 7.4, both cationic (2-OH-E+) and zwitter-ionic (2-O--E+) forms of the molecule should be present in almost equal amounts. Similar pH equilibria have been observed for mitochondria-targeted compounds, with pKa values of 0.2 and 2.0 for Mito-E+ and 0.4, 1.9 and 7.2 for 2-OH-Mito-E+ [116].
Binding to DNA
The binding of ethidium cation to polyanions, including DNA, is well known [117,118]. Ethidium has been used as a fluorescent stain for DNA and RNA, as the fluorescence intensity of DNA-bound ethidium is ca. 40 times higher than of unbound E+ [119,120]. 2-OH-E+ also binds to DNA with a concomitant increase in the quantum yield of fluorescence [69,72,116]. No DNA binding capabilities have been reported for the dimeric (HE-HE and HE-E+) oxidation products of HE; however, the dimer E+-E+ binds to DNA, as reflected by the red shift of its UV-Vis absorption spectrum [73]. The oxidation products of Mito-HE show enhancement in the fluorescence intensity upon binding to DNA [77]. The binding to DNA causes changes in the spectroscopic as well as in the physico-chemical properties of the cations [117,121]. For example, the pKa value of the hydroxyl group of 2-OH-Mito-E+ in aqueous solutions containing DNA is 10.4 [77], as compared to the value of 7.2 in the absence of DNA [73].
Binding to the cation-exchange column
The cationic species, E+ and 2-OH-E+, can also bind to the cation exchange columns, and this property has been utilized for purification of HE stock solutions from contaminating E+ [122]. This property can be also used for isolation of E+ and 2-OH-E+ from cell and tissue extracts [84,98].
Redox reactions
Ethidium is reduced to HE via a hydride anion transfer (Figure 2). This reaction has been used to synthesize HE which is accomplished by a two-electron reduction of E+ to HE using sodium borohydride [17]. It was reported previously that E+ can be reduced enzymatically to HE by glucose oxidase [83]. Although the possibilities of an enzymatic reduction of E+ to HE in cellular systems have not yet been explored, a recent report suggests that 2-OH-E+ is formed from E+ in endothelial cells via reduction of E+ to HE [71]. The electrochemical reduction of E+ has also been studied and HE was reported as the final product [123]. Two reduction peaks (at -0.232 V and -556 V vs. Ag/AgCl) have been identified during E+ reduction. Moreover, the product formed by the reduction of E+ at the potentials between -0.3 V and -0.45 V, which was assigned to a carbon-centered ethidium radical (E•), was highly fluorescent (λexc = 260 nm, λemi = 450 nm) [123]. The radical intermediate formed from the one-electron reduction of ethidium cation has been characterized by pulse radiolysis and based on the differences in electronic absorption spectra, it was concluded that the intermediate formed is structurally different from the species formed upon one-electron oxidation of HE [80]. 2-OH-E+ was shown to be unreactive towards the biologically relevant reductants, involving ascorbic acid and glutathione [71].
In the presence of strong oxidants (e.g. dibromine radical anion, azidyl radical), ethidium could be oxidized to form the radical dication E•2+ that exists in equilibrium with its deprotonated form, ethidium radical cation, E(•NH)+ (pKa = 8.2) [121]. However, a recent study showed that while 2-OH-E+ can be oxidized by horseradish peroxidase enzyme (HRP) and hydrogen peroxide, E+ was unreactive under those conditions. This feature enabled quantification of 2-OH-E+ and E+ by measuring the fluorescence intensity of the mixture before and after incubation with HRP and H2O2 [84,98].
Other considerations affecting HE-derived fluorescence
The ethidium cation, a two-electron oxidation product of HE, was shown to interact with antimycin, a mitochondrial complex III inhibitor, thereby causing an increase in the fluorescence intensity [124]. Mn(III)TBAP has been reported to react with E+ and 2-OH-E+, which can potentially complicate the interpretation of results of its use as an SOD–mimetic agent [76]. Moreover, the cationic porphyrins reportedly quench the fluorescence of ethidium bound to DNA, but not that of ethidium in DNA-free solution [125]. Thus, in addition to HE consumption, the metal porphyrin complexes may also decrease the fluorescence intensity observed by physical and/or chemical interaction with the fluorescent products. The ethidium cation is known to undergo self-association [126-128], but the possibility of self- or hetero-association processes of 2-OH-E+ is not known.
Methods for detecting HE and its oxidation products in cells and tissues
As ethidium has long been assumed to be the sole product of HE oxidation, the quantification of HE oxidation was based on the changes in the fluorescence intensity. With increased understanding of the oxidative chemistry of HE, it became apparent that even for qualitative analysis, the selective detection of the specific products is critical. The fluorescence settings with greater sensitivity for 2-OH-E+ than for E+ have been established to achieve more selective detection of superoxide-specific products. However, to obtain the whole profile of HE oxidation products, the methods based on separation of HE and its products, including HPLC techniques, have been established. The ability to detect the extent of HE consumption and monitor the formation of the oxidation product(s) is a critical factor in HE-based assay for superoxide. Each detection method described below has its own advantages as well as limitations that should be taken into consideration.
Fluorescence-based techniques
The red fluorescence (excitation ≈ 500 nm, emission ≈ 600 nm, Figure 3) formed from HE has been used in the detection of intracellular superoxide for the last two decades. As HE is also fluorescent (excitation at 350 nm, emission at 400 nm, Figure 3B), the blue fluorescence intensity due to HE has been used to monitor its intracellular uptake. The major advantages of fluorescence detection are the possibility of “real-time” monitoring of the probe oxidation and the lack of cumbersome sample processing. Moreover, using the plate reader, large number of samples can be analyzed simultaneously. The fluorescence microscopy enables the determination of the intracellular/tissue localization of the fluorescent products of HE oxidation and HE distribution. The use of fluorescence detection methods for superoxide measurements was deemed to be inappropriate [87,99,116,129] due to several limitations, including the confounding effects of the non-specific oxidation product(s) of HE contributing to the total fluorescence intensity and light-sensitivity of HE. Selective detection of 2-OH-E+ using the excitation light at 396 nm was recently suggested [77,130]. The excitation band between 350 nm and 400 nm was shown to be present in the case of 2-OH-E+ (and 2-OH-Mito-E+) but not E+ (or Mito-E+) (Figure 3C). However, it should be emphasized that even at that wavelength, E+ will still absorb the excitation light and emit the fluorescence, although to a much lower extent than 2-OH-E+. Given that the levels of E+ detected in the biological systems are nearly ten-fold higher than that of 2-OH-E+, E+ could significantly contribute to the fluorescence intensity [116]. As additional oxidative products of HE in cells continue to be discovered, the interpretation of the changes in the fluorescence intensity should be made with caution.
The determination of the intracellular levels of HE using the fluorescence assay remains problematic. Although HE can be detected by fluorescence with a high sensitivity in cell-free systems, using the excitation around 350 nm and emission above 400 nm, the interfering autofluorescence due to the reduced forms of NAD(P) coenzymes in cells makes it difficult to determine intracellular HE levels. The changes in the cellular levels of NAD(P)H can complicate data interpretation. For example, it has been shown that rotenone treatment causes an increase in NAD(P)H autofluorescence [131], which could be mistaken for increased intracellular HE levels.
The fluorescent properties (excitation/emission maxima, quantum yields of fluorescence) of HE oxidation products can be affected by their intracellular localization. Besides binding to polyanionic macromolecules such as DNA, the hydrophobic environment can also affect the fluorescence of E+ and 2-OH-E+. For example, the partitioning of 2-OH-E+ into n-octanol phase, or binding to DNA is accompanied by a shift in the maxima of excitation and emission of fluorescence, resulting in the appearance of an additional excitation band between 350 nm and 400 nm [116].
Another drawback of the fluorescence-based detection is the photosensitivity of HE (see above), which can affect the formation of the fluorescent products, especially during constant monitoring of intracellular HE. Moreover, the excitation of the products of HE oxidation may stimulate further HE oxidation, as has been shown in the case of 2-OH-E+ [76].
HPLC separation of HE oxidation products
Clearly, there are many drawbacks with the fluorescence approaches for detection of superoxide in intact cells or tissue. The HPLC-based methods allow for detection of both HE and the whole range of the oxidation products. In addition to the analytes' resolution based on the different retention times, the detectors coupled with HPLC systems typically offer the additional selectivity options via different detector settings, including different absorption wavelengths (UV-Vis absorption detectors), excitation/emission wavelengths (fluorescence detectors), different electric potentials (electrochemical detectors) or different mass/charge values (MS detectors). The major drawbacks of the HPLC methods include the lengthy analysis time (typically 30-75 min per sample) and the need to optimize the sample extraction procedures. The development of new HPLC technology (ultra-performance liquid chromatography, UPLC) aimed at shortening the analysis time by decreasing the column length and particle size (while retaining the resolution characteristic for longer columns) may pave the way for faster analysis of HE and its oxidation products. Different extraction/sample preparation procedures from cells/tissues have been described, but no systematic studies have been reported on the actual efficiency of the extraction together with the stability of HE and its products during sample preparation. Although the HPLC-based separation and detection methods have been most popular, other interesting approaches include the low-pressure chromatography and electrophoresis-based separation.
HPLC with UV-Vis absorption and/or fluorescence detection
The HPLC method coupled with fluorescence detection has been employed to selectively detect and quantify 2-OH-E+ and E+ both in cell-free and cellular systems [18,69,71-74,80,81,99,116,132-149]. The absorption detection at 290 nm and 500 nm has also been used for monitoring the HE oxidation products [80,99,116], and detection at 245 and/or 350-370 nm for monitoring HE levels [71,74,99,116,148]. Recently, the fluorescence detection of HE during HPLC analysis has been shown to be highly sensitive, and the method of programming of the fluorescence detector to detect both HE and its oxidation products during a single HPLC run has been proposed [99].
With Mito-HE, the HPLC methods with absorption/fluorescence detection have been described [73,77,99,116]; however no data on HPLC-based assay for Mito-HE and its oxidation products extracted from cell cultures or tissue have been published to date.
HPLC with electrochemical detection
To increase the sensitivity of the assay, the HPLC system equipped with electrochemical detection has been used to quantify HE and its oxidation products [76]. The electrochemical coulometric detection method for 2-OH-E+ is at least an order of magnitude more sensitive than the fluorescence detection [76]. This may, however, be dependent on the instrumental setup [99]. The reported HPLC method with amperometric detection of HE and 2-OH-E+ requires shorter time for sample analysis [150]. The electrochemical detection methods have already been applied to investigate HE oxidation in cell-free systems [73,76,80,99,151], cell culture systems [99,116,150-154] and in tissues from in vivo experiments [155]. The analysis of Mito-HE and its oxidation products by HPLC with electrochemical detection has been also described [73,99,116].
HPLC with mass spectrometric detection
As 2-OH-E+, E+ and E+-E+ are ionic species, their detection is easily achieved using mass spectral analysis [99]. Other analytes (HE, HE-HE and HE-E+) can be also monitored using mass spectrometric detectors, but in the case of HE and HE-HE, only the protonated forms can be detected [116]. To date, there are several reports utilizing HPLC-MS method for qualitative and/or quantitative analysis of HE oxidation products in cell-free and cellular systems [18,69,84,99,116,156]. Despite the selectivity of the MS detection due to different masses of the analytes, it needs to be emphasized that resolution based only on m/z differences may be not adequate for quantitative measurements. This is due to the overlap of the signals obtained from different protonation/isotopic forms of the analytes. For example, at m/z value of 316, in addition to the singly-protonated HE form, the isotopic peak of E+ is also present; while at m/z value of 314 (used for E+ quantification), the protonated form of HE-E+ can also be detected [116]. Despite the inherent sensitivity afforded by the additional positive charge localized on the triphenylphosphonium moiety, no reports on the HPLC-MS detection of Mito-HE and its oxidation products have been published so far.
Electrophoresis-based separation
The separation of the charged oxidation products of HE has also been obtained using the micellar electrokinetic capillary chromatography technique, and the fluorescent products have been quantified using the laser-induced fluorescence detection system [75,157]. The method has also been used to detect 2-OH-Mito-E+ and Mito-E+ [158]. This is the most sensitive method that is currently available (detection limit of 0.15 amol for 2-OH-E+) [75]. An additional advantage of this method is a short analysis time, as it takes less than three minutes to analyze a sample. The major drawback of the published method is that only the fluorescent cations (2-OH-E+ and E+, and their mitochondria-targeted analogs formed from Mito-HE) have been detected. Thus the probe (HE or Mito-HE) uptake into cells remains unknown, complicating the interpretation of the results.
Microcolumn/HRP assay
The separation of HE and its oxidation products has been also achieved using the sequential cation exchange and hydrophobic microcolumn low pressure chromatography [84,98]. After separation, a mixture of 2-OH-E+ and E+ was obtained and after adding DNA, the fluorescence intensity of the mixture was measured. Subsequently 2-OH-E+ was converted into a non-fluorescent product by HRP/H2O2 system, and the fluorescence intensity was measured again. From the difference in the fluorescence intensities, the amount of 2-OH-E+ was calculated [84,97,98]. The major advantage of this assay is the possibility of being able to perform parallel processing of a number of samples, thus decreasing the total analysis time. However, this method is not any more sensitive than the typical HPLC method with fluorescent or electrochemical detection [76,84]. The major drawback of the assay is that 2-OH-E+ is not quantified directly, but its amount is calculated from the difference in the fluorescence intensities measured before and after incubation with HRP/H2O2 [98]. Another disadvantage is the lack of detection of additional (e.g., dimeric) oxidation products of HE.
Factors affecting the yield of 2-hydroxyethidium in cells and tissues
The question is: is HE the ideal intracellular probe for measuring superoxide? Intracellularly, HE has to compete with SOD (k(SOD + O2•-) = 2 × 109 M-1s-1), which is present at a micromolar concentration in the cytosol. To outcompete SOD, HE levels should reach millimolar concentrations in cells, which may be cytotoxic. Thus, although 2-hydroxyethium, a specific marker for superoxide radical anion, has been used to monitor changes in superoxide levels, the amount of 2-OH-E+ produced is determined not only by intracellular superoxide generation, but also by other variables, e.g., HE uptake, HE consumption rate, and its availability and efficiency in the competition for superoxide with SOD, and intracellular peroxidatic activities [116]. The intracellular uptake of HE by cells is a function of its concentration and stability in the medium and the efficiency of transport from the extracellular medium into the cells.
Stability of HE in the tissue culture media
In a typical cell culture medium (e.g., Dulbecco's Modified Eagle Medium, DMEM), lacking HE-reactive additives (for example Mn(III)TBAP [76]), HE is relatively stable and virtually constant steady-state concentration of extracellular HE can be achieved. As a part of the experimental procedure, the stability of HE in the specific media has to be tested. HE has been used in the quantification of superoxide production by cellular organelles isolated from animal tissues or cultured cells, including microsomes [159,160], mitochondria [161-163] or cell membranes [74]. In all these instances, no data concerning the stability of HE in the incubation medium were provided.
While maintaining a constant HE level in the extracellular medium to assure the continuous availability of the probe is a relatively easy task in cell culture experiments, the situation is much more complicated in ex vivo tissue culture experiments, where the probe has to be transported to the site of superoxide production through multiple cellular compartments. Examples of the use of HE in ex vivo experiments include the detection of in situ production of superoxide in brain tissue [53,164-173], spinal cord sections[174-176], blood vessels [74,91,138,139,141,145,177-270], diaphragms [52,85,271-273], various components of kidney [94,274-280], heart [201,281-291], islets of Langerhans [292], penis sections [293], liver sections [294,295], lungs [296,297], retinal tissue sections [298], eye sections [299-301] and prostate tissue [302]. HE has been also used to detect superoxide in tissue homogenates [207,279,303-306] as well as in whole blood [307-311]. Although in some cases HE was constantly infused during the experiment, the availability of the probe for the cellular uptake remains unknown. Even more challenging is the task of delivering and maintaining a constant supply of HE to the specific organ in the in vivo experiments. The transport of HE from the site of injection to the organ of interest via the circulatory system is limited by the high reactivity of the probe with the heme proteins to form the ethidium cation (see above). Nonetheless the application of HE to detect in vivo production of superoxide has been reported [84,97,107-113,155,164,312-339]. It has been postulated that after an intravenous injection, HE is translocated to the brain [108-111,312,316-318,321-325,328,329,331], kidney [327] and eyes [315,319]. When HE was administered subcutaneously, 2-OH-E+ has been detected of in the heart, brain, kidney, liver, lungs and spleen [84]. After the intraperitoneal injection of HE solution, the red fluorescence has been detected in animals' brains [97,112,113,314,320] and hearts [334]. With a few exceptions ([97,155]), the estimation of transport efficiency of HE to the targeted organ in most of those reports was based on the fluorescence intensity of the oxidation products (E+ and 2-OH-E+), which does not provide any information concerning the availability of HE for reaction with superoxide. Moreover, as no reports have been reported on the distribution of E+ (or 2-OH-E+) in in vivo experiments, it remains to be established whether the fluorescence signals observed are due to oxidation of HE or transport of the oxidized product(s) formed elsewhere to the target organ.
Transport efficiency through cell membranes
The transport of HE from the extracellular environment into the cytosol is thought to occur by passive diffusion, although it remains to be established how different incubation conditions affect the efficiency of the cellular uptake of HE. Under various conditions (including oxidative stress), the permeability of the cell membranes changes [340-342]. It is yet to be established whether this would cause changes in the uptake of HE. With increased intracellular uptake of HE, one would expect an increased level of HE oxidation products, including 2-OH-E+, even in the absence of increased steady-state level of superoxide (see below).
Intracellular stability of HE
Intracellularly, HE is consumed via both superoxide-dependent and- independent pathways. Although the oxidation products that are not specific for superoxide (E+, dimers) are usually formed in higher yields than the superoxide-specific product, 2-OH-E+ in biological systems, the identity of the superoxide-independent pathways still remains to be established. It has been postulated that HE undergoes NADP+-dependent enzymatic dehydrogenation to produce E+ [20,22,45]; however other pathways, including oxidation by heme proteins [73] may be also involved. While the intracellular HE concentration is clearly an important factor controlling the amount of 2-OH-E+ produced, its actual concentration inside the cells is usually not measured. Based on the published data for intracellular HE levels in endothelial cells (ca. 0.2 nmol/mg protein after treatment of the cells with 10 μM HE [99]) and the endothelial cell volume reported (1 – 7 μl/mg protein, depending on the cellular environment [343]), one can estimate the average concentration of intracellular HE in the low micromolar to sub-millimolar range. In view of the high reactivity of SOD towards superoxide and its intracellular concentration within the micromolar range, the majority of superoxide is probably scavenged by SOD. Thus, the amount of superoxide trapped by HE does not reflect its total intracellular level, and the fraction of superoxide scavenged by HE should be a function of the ratio of HE to SOD levels. Instead of using the absolute fluorescence intensity of the oxidized product, the ratio of the fluorescence intensities of the oxidized product to unreacted HE has been used as a marker of superoxide (or ROS) production [227,260,275,277,344-348]. In some reports, the amount of 2-OH-E+ and/or E+ has been “normalized” to the amount of HE consumed, thus giving the fraction of HE converted to the specific product [74,134]. Also the ratio of the amount of 2-OH-E+ to the intracellular level of E+ formed has been used to “normalize” the amount of 2-OH-E+. This is based on the assumption that the concentration of E+ is not influenced by the treatments used, and thus reflects mainly the intracellular uptake of HE [135]. To normalize the data to the intracellular concentration of HE, however, one has to measure the HE levels during the experiment. Assuming that the intracellular steady-state concentration of HE is reached quickly, as compared to the duration of the assay, and is maintained until the end of incubation at a consistent level, one can analyze the concentration of HE in cell lysate after the incubation. This value may be used to “normalize” the amount of 2-OH-E+ to intracellular HE availability and to determine its efficiency in competition with SOD for superoxide. While the straightforward way to “normalize” to HE level is to divide the intracellular amount of 2-OH-E+ by the intracellular amount of HE [150], one has to keep in mind that even in a simple competition between scavengers A and B (two pseudo-first-order reactions, with the second-order rate constants kA and kB, respectively), the concentration of the specific product (cA), is given by the equation:
where c0 is the concentration of the common reactant, for which the compounds A and B compete. This equation can be simplified to the linear dependence of concentration cA on [A]/[B] value:
only, when kA· [A] ≪ kB· [B]. In case of the competition between HE and SOD for superoxide, this requirement would mean that the amount of superoxide scavenged by HE will be the linear function of HE only when the concentration (and activity) of SOD is constant, with SOD scavenging most of the intracellular pool of superoxide. Although it can be expected that in many cases these conditions are met, it is not clear whether the intracellular competition between HE and SOD can be adequately described by this simple model.
Intracellular levels of superoxide radical anion
It is now established that 2-OH-E+ is the sole product of the reaction of superoxide with HE in well-defined chemical and enzymatic systems. However, it remained to be established whether the product is also formed in cellular systems and can be used as a diagnostic marker for intracellular superoxide. Several investigators have shown that modulation of intracellular superoxide level causes changes in the amount of 2-OH-E+ formed. As the steady-state concentration of superoxide is dependent on both the rates of its formation and decay, stimulation of superoxide production or its scavenging has been used to validate the utility of the assay. Incubation of cells with menadione, a well known redox-cycling agent, has been shown to increase the level of 2-OH-E+ formed in isolated mitochondria [75], endothelial cells [71,72,84], spermatozoa [81], ovary cells [150], intact aortic segments [71] and in various mouse organs [84], and this effect was inhibited by treatment of cells with PEG-SOD [71], MnTBAP [72] or Tempol [84], potential scavengers of superoxide.
Results from these studies suggest that 2-OH-E+ can be used as the diagnostic marker product of superoxide radical. It has been shown that other redox active agents (e.g., paraquat [137] and mitoquinone [146]), cause an increase of the amount of intracellular 2-OH-E+. Angiotensin II, a physiologically-relevant stimulus, increased the amount of 2-OH-E+, presumably by activating the NADPH oxidase. The separation and quantitation of the HE oxidation products revealed the increased formation of 2-OH-E+ or both 2-OH-E+ and E+ following angiotensin II treatment [71,134,136,138,145]. The bacterial endotoxin, lipopolysaccharide (LPS), has been reported to increase the formation of intracellular 2-OH-E+ [74], which was attributed to activation of NADPH oxidase by LPS. Other physiological stimuli that can potentially stimulate superoxide production and their signaling pathway, including ceramide [72] and arachidonic acid [156], increase the intracellular level of 2-OH-E+ as well. The drug phenylephrine, α1-adrenergic receptor agonist, has also been reported to increase the amount of 2-OH-E+, as measured by HPLC [139]. 1-Methyl-4-phenylpyridinium cation (MPP+), a neurotoxin used in the cellular models of Parkinson disease, stimulated the formation of 2-OH-E+ in neuroblastoma cells [142].
The manipulation of the cellular antioxidant system has also been shown to affect the level of 2-OH-E+ detected. For example, depletion of intracellular GSH levels by incubating cells with buthionine sulfoximine has been shown to increase the intracellular concentration of 2-OH-E+ [71]. On the other hand, overexpression of intracellular SOD1 has been shown to inhibit angiotensin II-stimulated 2-OH-E+ formation [74]. The transfection studies should, however, be done with appropriate care as it has been shown that an empty vector can affect intracellular 2-OH-E+ and E+ levels [74]. Similarly, while incubating cells with PEG-SOD causes a decrease in 2-OH-E+ level, PEG alone and PEG-catalase have also been shown to exhibit the inhibitory effects [74]. These effects have been attributed to PEG-induced changes in the cell membrane fluidity, which may cause changes in the transmembrane transport of HE and/or its oxidation products.
The steady-state level of HE-derived radical
According to the proposed mechanism of the reaction of HE with superoxide (Figure 5), HE undergoes oxidation to the HE radical form (HE•+ or HE(•NH)) during the first step of the reaction. HE-derived radical reacts with another molecular of superoxide leading to the formation of 2-OH-E+. The first step – one electron oxidation of HE may be achieved by a variety of oxidants [18,73,80,87,88], and thus one would predict that superoxide-independent oxidation of HE to its radical can increase the yield of 2-OH-E+ at a constant flux of superoxide. This hypothesis, however, needs to be verified experimentally. With HE-based cell-free assay for superoxide, in the absence of other superoxide scavengers, one-electron oxidants could change the stoichiometry of the reaction from 1:2 (HE:O2•-) to 1:1, thus doubling the amount of 2-OH-E+. In the presence of other scavengers (e.g. SOD in cells), the efficiency of trapping superoxide by HE can be increased even more, as HE-derived radical reacts with superoxide much faster than HE (estimated rate constant of HE-derived radical with O2•-, k ∼ 2 × 109 M-1s-1 [80]). For example, the oxidation of HE by cytochrome c may account the increased red fluorescence intensity observed during the release of cytochrome c from mitochondria into cytosol during apoptosis [349]. The occurrence of one-electron oxidation of HE in cells can be verified by monitoring the levels of HE-derived dimeric products using HPLC-based assays. The presence of the compounds capable of reacting with HE-derived radical may cause a decrease in the amount of 2-OH-E+. Reduction of the radical back to the parent molecule (HE) is another complicating factor. The reactivity of HE-derived radical with biologically relevant reductants remains to be determined.
Stability and intracellular retention of 2-OH-E+
The use 2-OH-E+ for the (semi)quantitative analysis of superoxide formation inside the cells is based on the assumption that 2-OH-E+ formed intracellularly is retained inside the cells and does not undergo metabolic conversion to other species during the course of the experiment. Thus, the intracellular amount of 2-OH-E+ should linearly increase with incubation time and superoxide flux. Due to the destructive nature of the HPLC-based methods, each sample can be analyzed only for a single timepoint. In most reports, the analysis of the samples has been done using a single incubation time, without investigating the time-course of the 2-OH-E+ accumulation. As 2-OH-E+ binds to DNA, it is generally assumed that its transport out of the cells is negligible. It has been reported that after loading the endothelial cells with 2-OH-E+, its intracellular levels monitored for an hour remained unchanged [71]. In addition, incubation of 2-OH-E+ with the cell homogenate did not induce metabolic changes to the compound [71]. These results indicate the general stability of 2-OH-E+ in endothelial cells. However, the possibility of its metabolism or transport in other types of cells remains to be explored. Reports indicate that in some biological samples, peroxidase-catalyzed oxidation of 2-OH-E+ may decrease its levels, and hence the use of a non-specific peroxidase inhibitor, potassium cyanide, was recommended to overcome such problems [98].
HE-based localization of the sources of superoxide
The HE-based fluorescence assay has been used to identify the localization of superoxide source in tissues and cells. Those experiments are based on several assumptions:
The effectiveness of scavenging of superoxide by HE is similar in different cellular compartments, i.e. HE is distributed uniformly over the whole cell or tissue.
The fluorescent compound formed from HE oxidation remains stationary at the site of its formation, i.e. no intracellular or intercellular transport of the fluorescent product occurs during the experiment.
The fluorescent product formed is due to the reaction of HE with superoxide, i.e. the fluorescence intensity originates mostly from 2-OH-E+.
The differences in the fluorescence quantum yields of 2-OH-E+ in different environments cannot account for the observed differences in the fluorescence intensity.
These aspects are critically important for proper interpretation of results. HE distribution (and its kinetic parameters) in the whole organ or even in intact tissues has not been established. Even if it were to distribute evenly, the fluorescent products derived from HE are likely to translocate inside the cells. Due to the extreme affinity of E+ to DNA, the prevailing view is that the fluorescent oxidation product of HE relocates to the nucleus and binds to DNA. It has been shown that HE-derived red fluorescence colocalizes with the blue fluorescence of the nucleus-specific fluorescent dye – 4,6-diamino-2-phenylindole (DAPI) [252]. With regard to mitochondria, binding to mitochondrial nucleic acids has been assumed. It has been reported that HE and Mito-HE oxidation products can bind to the isolated mitochondria [73,350,351]. Moreover, E+ has been widely used to delete mitochondrial DNA in intact cells [352]. Thus, it is likely that the fluorescent products of HE oxidation may bind to mitochondrial DNA after translocation from the cytosol.
However, reports also suggest that E+ formed in mitochondria can exit out of the mitochondria after the addition of an uncoupling agent, thus causing a change in the fluorescence intensity [353-355]. Similar findings were reported using an antimicrobial peptide histatin 5 in yeast. Here the increase in HE-derived fluorescence intensity has been attributed to the release of the ethidium cation from the mitochondrial matrix rather than to oxidation of HE [356]. The most overlooked aspect of HE-derived fluorescence is the relative contribution from 2-OH-E+, the fluorescent product of HE and superoxide, to the total fluorescence intensity. In the absence of information regarding the identity of the product formed, it is virtually impossible to deduce the nature of the actual oxidant and its site of production.
Summary
Currently, HE is the most commonly used ROS-specific probe, especially for detecting intracellular superoxide. There is an ever increasing number of reports on the detection of superoxide with HE and Mito-HE, its mitochondria-targeted analog. These reports often lack any information with respect to superoxide reaction with the probes [129]. It has been nearly six years after publication of the initial report suggesting that HE/superoxide reaction forms 2-hydroxyethidium but not ethidium as a major product. However, many publications still claim that ethidium is the major product of HE and superoxide [357]. This review emphasizes the various factors that can affect the yield of 2-hydroxyethidium, the diagnostic marker of superoxide and HE reaction. The major conclusion is that the knowledge of the whole profile of HE oxidation products together with the intracellular levels of HE is required to make any conclusion regarding the amount of intracellular superoxide and/or the effect of the specific inhibitors. The use of a HPLC-based methodology is currently the only way of fulfilling this requirement. HE-based fluorescence microscopy alone will not be able to provide trustworthy information on intracellular superoxide formation. That said, both HE and Mito-HE (Mito-SOX) could, however, serve as ideal probes for measuring intracellular oxidant formation. As radicals derived from HE and Mito-HE do not react with oxygen to form additional superoxide and H2O2, disproportionation and radical recombination products derived from HE (ethidium, 2-hydroxyethidium, and dimers) and Mito-HE (Mito-ethidium, 2-hydroxy-mito-ethidium, and the corresponding dimers) yield a quantitative measurement of oxidants formed in the cytosolic and mitochondrial regions.
Acknowledgments
This work was supported by National Institutes of Health Grants NS40494, HL073056, HL063119
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Reference List
- 1.Fridovich I. Superoxide dismutases. An adaptation to a paramagnetic gas. J Biol Chem. 1989;264:7761–7764. [PubMed] [Google Scholar]
- 2.Fridovich I. Superoxide anion radical (O2-.), superoxide dismutases, and related matters. J Biol Chem. 1997;272:18515–18517. doi: 10.1074/jbc.272.30.18515. [DOI] [PubMed] [Google Scholar]
- 3.Fridovich I. Oxygen toxicity: a radical explanation. J Exp Biol. 1998;201:1203–1209. doi: 10.1242/jeb.201.8.1203. [DOI] [PubMed] [Google Scholar]
- 4.Fridovich I. Superoxide radical and superoxide dismutase. Acc Chem Res. 1972;5:321–326. [Google Scholar]
- 5.McCord JM, Fridovich I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein) J Biol Chem. 1969;244:6049–6055. [PubMed] [Google Scholar]
- 6.Bartosz G. Use of spectroscopic probes for detection of reactive oxygen species. Clin Chim Acta. 2006;368:53–76. doi: 10.1016/j.cca.2005.12.039. [DOI] [PubMed] [Google Scholar]
- 7.Dikalov S, Griendling KK, Harrison DG. Measurement of reactive oxygen species in cardiovascular studies. Hypertension. 2007;49:717–727. doi: 10.1161/01.HYP.0000258594.87211.6b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Halliwell B, Whiteman M. Measuring reactive species and oxidative damage in vivo and in cell culture: how should you do it and what do the results mean? Br J Pharmacol. 2004;142:231–255. doi: 10.1038/sj.bjp.0705776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tarpey MM, Fridovich I. Methods of detection of vascular reactive species: nitric oxide, superoxide, hydrogen peroxide, and peroxynitrite. Circ Res. 2001;89:224–236. doi: 10.1161/hh1501.094365. [DOI] [PubMed] [Google Scholar]
- 10.Tarpey MM, Wink DA, Grisham MB. Methods for detection of reactive metabolites of oxygen and nitrogen: in vitro and in vivo considerations. Am J Physiol Regul Integr Comp Physiol. 2004;286:R431–R444. doi: 10.1152/ajpregu.00361.2003. [DOI] [PubMed] [Google Scholar]
- 11.Spasojevic I, Liochev SI, Fridovich I. Lucigenin: redox potential in aqueous media and redox cycling with O-(2) production. Arch Biochem Biophys. 2000;373:447–450. doi: 10.1006/abbi.1999.1579. [DOI] [PubMed] [Google Scholar]
- 12.Vasquez-Vivar J, Hogg N, Pritchard KA, Jr, Martasek P, Kalyanaraman B. Superoxide anion formation from lucigenin: an electron spin resonance spin-trapping study. FEBS Lett. 1997;403:127–130. doi: 10.1016/s0014-5793(97)00036-7. [DOI] [PubMed] [Google Scholar]
- 13.Wardman P, Burkitt MJ, Patel KB, Lawrence A, Jones CM, Everett SA, Vojnovic B. Pitfalls in the use of common luminescent probes for oxidative and nitrosative stress. J Fluoresc. 2002;12:65–68. [Google Scholar]
- 14.Khan N, Wilmot CM, Rosen GM, Demidenko E, Sun J, Joseph J, O'Hara J, Kalyanaraman B, Swartz HM. Spin traps: in vitro toxicity and stability of radical adducts. Free Radic Biol Med. 2003;34:1473–1481. doi: 10.1016/s0891-5849(03)00182-5. [DOI] [PubMed] [Google Scholar]
- 15.Shi H, Timmins G, Monske M, Burdick A, Kalyanaraman B, Liu Y, Clement JL, Burchiel S, Liu KJ. Evaluation of spin trapping agents and trapping conditions for detection of cell-generated reactive oxygen species. Arch Biochem Biophys. 2005;437:59–68. doi: 10.1016/j.abb.2005.02.028. [DOI] [PubMed] [Google Scholar]
- 16.Villamena FA, Zweier JL. Detection of reactive oxygen and nitrogen species by EPR spin trapping. Antioxid Redox Signal. 2004;6:619–629. doi: 10.1089/152308604773934387. [DOI] [PubMed] [Google Scholar]
- 17.Thomas G, Roques B. Proton magnetic resonance studies of ethidium bromide and its sodium borohydride reduced derivative. FEBS Lett. 1972;26:169–175. [Google Scholar]
- 18.Zielonka J, Zhao H, Xu Y, Kalyanaraman B. Mechanistic similarities between oxidation of hydroethidine by Fremy's salt and superoxide: stopped-flow optical and EPR studies. Free Radic Biol Med. 2005;39:853–863. doi: 10.1016/j.freeradbiomed.2005.05.001. [DOI] [PubMed] [Google Scholar]
- 19.Bucana C, Saiki I, Nayar R. Uptake and accumulation of the vital dye hydroethidine in neoplastic cells. J Histochem Cytochem. 1986;34:1109–1115. doi: 10.1177/34.9.2426339. [DOI] [PubMed] [Google Scholar]
- 20.Bucana CD, Giavazzi R, Nayar R, O'Brian CA, Seid C, Earnest LE, Fan D. Retention of vital dyes correlates inversely with the multidrug-resistant phenotype of adriamycin-selected murine fibrosarcoma variants. Exp Cell Res. 1990;190:69–75. doi: 10.1016/0014-4827(90)90145-z. [DOI] [PubMed] [Google Scholar]
- 21.Cavarec L, Quillet-Mary A, Fradelizi D, Conjeaud H. An improved double fluorescence flow cytometry method for the quantification of killer cell/target cell conjugate formation. J Immunol Methods. 1990;130:251–261. doi: 10.1016/0022-1759(90)90055-z. [DOI] [PubMed] [Google Scholar]
- 22.Gallop PM, Paz MA, Henson E, Latt SA. Dynamic Approaches to the Delivery of Reporter Reagents into Living Cells. BioTechniques. 1984;1:32–36. [Google Scholar]
- 23.Kuypers TW, Koenderman L, Weening RS, Verhoeven AJ, Roos D. Continuous cell activation is necessary for stable interaction of complement receptor type 3 with its counter-structure in the aggregation response of human neutrophils. Eur J Immunol. 1990;20:501–508. doi: 10.1002/eji.1830200307. [DOI] [PubMed] [Google Scholar]
- 24.Bowles AP, Jr, Pantazis CG, Wansley W, Allen MB. Chemosensitivity testing of human gliomas using a fluorescent microcarrier technique. J Neurooncol. 1990;8:103–112. doi: 10.1007/BF00177832. [DOI] [PubMed] [Google Scholar]
- 25.Fidler IJ, Heicappell R, Saiki I, Grutter MG, Horisberger MA, Nuesch J. Direct antiproliferative effects of recombinant human interferon-alpha B/D hybrids on human tumor cell lines. Cancer Res. 1987;47:2020–2027. [PubMed] [Google Scholar]
- 26.Saiki I, Bucana CD, Tsao JY, Fidler IJ. Quantitative fluorescent microassay for identification of antiproliferative compounds. J Natl Cancer Inst. 1986;77:1235–1240. [PubMed] [Google Scholar]
- 27.Ericsson SA, Garner DL, Redelman D, Ahmad K. Assessment of the viability and fertilizing potential of cryopreserved bovine spermatozoa using dual fluorescent staining and two-flow cytometric systems. Gamete Res. 1989;22:355–368. doi: 10.1002/mrd.1120220402. [DOI] [PubMed] [Google Scholar]
- 28.Bicalho KA, Ribeiro MF, Martins-Filho OA. Molecular fluorescent approach to assessing intraerythrocytic hemoprotozoan Babesia canis infection in dogs. Vet Parasitol. 2004;125:221–235. doi: 10.1016/j.vetpar.2004.08.009. [DOI] [PubMed] [Google Scholar]
- 29.Evangelista V, Manarini S, Coller BS, Smyth SS. Role of P-selectin, beta2-integrins, and Src tyrosine kinases in mouse neutrophil-platelet adhesion. J Thromb Haemost. 2003;1:1048–1054. doi: 10.1046/j.1538-7836.2003.00214.x. [DOI] [PubMed] [Google Scholar]
- 30.Harvey RW, Mayberry N, Kinner NE, Metge DW, Novarino F. Effect of growth conditions and staining procedure upon the subsurface transport and attachment behaviors of a groundwater protist. Appl Environ Microbiol. 2002;68:1872–1881. doi: 10.1128/AEM.68.4.1872-1881.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pattanapanyasat K, Walsh DS, Yongvanitchit K, Piyawatthanasakul N, Wanachiwanawin W, Webster HK. Robust in vitro replication of Plasmodium falciparum in glycosyl-phosphatidylinositol-anchored membrane glycoprotein-deficient red blood cells. Am J Trop Med Hyg. 2003;69:360–365. [PubMed] [Google Scholar]
- 32.Horobin RW, Rashid F. Interactions of molecular probes with living cells and tissues. Part 1. Some general mechanistic proposals, making use of a simplistic Chinese box model. Histochemistry. 1990;94:205–209. doi: 10.1007/BF02440189. [DOI] [PubMed] [Google Scholar]
- 33.Ashley N, Harris D, Poulton J. Detection of mitochondrial DNA depletion in living human cells using PicoGreen staining. Exp Cell Res. 2005;303:432–446. doi: 10.1016/j.yexcr.2004.10.013. [DOI] [PubMed] [Google Scholar]
- 34.Coetzee JF, Apley MD, Kocan KM, Jones DE. Flow cytometric evaluation of selected antimicrobial efficacy for clearance of Anaplasma marginale in short-term erythrocyte cultures. J Vet Pharmacol Ther. 2006;29:173–183. doi: 10.1111/j.1365-2885.2006.00734.x. [DOI] [PubMed] [Google Scholar]
- 35.Davis WC, Wyatt CR, Hamilton MJ, Goff WL. A rapid, reliable method of evaluating growth and viability of intraerythrocytic protozoan hemoparasites using fluorescence flow cytometry. Mem Inst Oswaldo Cruz. 1992;87(3):235–239. doi: 10.1590/s0074-02761992000700039. [DOI] [PubMed] [Google Scholar]
- 36.Heller R. Spectrofluorometric assay for the quantitation of cell-tissue electrofusion. Anal Biochem. 1992;202:286–292. doi: 10.1016/0003-2697(92)90107-i. [DOI] [PubMed] [Google Scholar]
- 37.Jouin H, Daher W, Khalife J, Ricard I, Puijalon OM, Capron M, Dive D. Double staining of Plasmodium falciparum nucleic acids with hydroethidine and thiazole orange for cell cycle stage analysis by flow cytometry. Cytometry A. 2004;57:34–38. doi: 10.1002/cyto.a.10110. [DOI] [PubMed] [Google Scholar]
- 38.Shi T, Eaton AM, Ring DB. Selection of hybrid hybridomas by flow cytometry using a new combination of fluorescent vital stains. J Immunol Methods. 1991;141:165–175. doi: 10.1016/0022-1759(91)90143-4. [DOI] [PubMed] [Google Scholar]
- 39.Zucker RM, Keshaviah AP, Price OT, Goldman JM. Confocal laser scanning microscopy of rat follicle development. J Histochem Cytochem. 2000;48:781–791. doi: 10.1177/002215540004800607. [DOI] [PubMed] [Google Scholar]
- 40.Callewaert DM, Radcliff G, Waite R, LeFevre J, Poulik MD. Characterization of effector-target conjugates for cloned human natural killer and human lymphokine activated killer cells by flow cytometry. Cytometry. 1991;12:666–676. doi: 10.1002/cyto.990120711. [DOI] [PubMed] [Google Scholar]
- 41.Debout C, Birebent B, Griveau AM, Izard J. In vitro cytotoxic effect of guinea-pig natural killer cells (Kurloff cells) on homologous leukemic cells (L2C) Leukemia. 1993;7:733–735. [PubMed] [Google Scholar]
- 42.Radcliff G, Waite R, LeFevre J, Poulik MD, Callewaert DM. Quantification of effector/target conjugation involving natural killer (NK) or lymphokine activated killer (LAK) cells by two-color flow cytometry. J Immunol Methods. 1991;139:281–292. doi: 10.1016/0022-1759(91)90199-p. [DOI] [PubMed] [Google Scholar]
- 43.Zanyk MJ, Banerjee D, McFarlane DL. A flow cytometric assay for target binding by NKH1A+ cells using a single laser system. Anal Cell Pathol. 1990;2:241–251. [PubMed] [Google Scholar]
- 44.Tarin JJ, Cano A. Distribution of 5-chloromethylfluorescein diacetate staining during meiotic maturation and fertilization in vitro of mouse oocytes. J Reprod Fertil. 1998;114:211–218. doi: 10.1530/jrf.0.1140211. [DOI] [PubMed] [Google Scholar]
- 45.Olive PL. Hydroethidine: a fluorescent redox probe for locating hypoxic cells in spheroids and murine tumours. Br J Cancer. 1989;60:332–338. doi: 10.1038/bjc.1989.281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rothe G, Valet G. Flow cytometric analysis of respiratory burst activity in phagocytes with hydroethidine and 2′,7′-dichlorofluorescin. J Leukoc Biol. 1990;47:440–448. [PubMed] [Google Scholar]
- 47.Kobzik L, Godleski JJ, Brain JD. Oxidative metabolism in the alveolar macrophage: analysis by flow cytometry. J Leukoc Biol. 1990;47:295–303. [PubMed] [Google Scholar]
- 48.Kobzik L, Godleski JJ, Brain JD. Selective down-regulation of alveolar macrophage oxidative response to opsonin-independent phagocytosis. J Immunol. 1990;144:4312–4319. [PubMed] [Google Scholar]
- 49.Perticarari S, Presani G, Mangiarotti MA, Banfi E. Simultaneous flow cytometric method to measure phagocytosis and oxidative products by neutrophils. Cytometry. 1991;12:687–693. doi: 10.1002/cyto.990120713. [DOI] [PubMed] [Google Scholar]
- 50.Al-Mehdi AB, Shuman H, Fisher AB. Intracellular generation of reactive oxygen species during nonhypoxic lung ischemia. Am J Physiol. 1997;272:L294–L300. doi: 10.1152/ajplung.1997.272.2.L294. [DOI] [PubMed] [Google Scholar]
- 51.Bradner JR, Nevalainen KM. Metabolic activity in filamentous fungi can be analysed by flow cytometry. J Microbiol Methods. 2003;54:193–201. doi: 10.1016/s0167-7012(03)00043-5. [DOI] [PubMed] [Google Scholar]
- 52.Nethery D, Stofan D, Callahan L, DiMarco A, Supinski G. Formation of reactive oxygen species by the contracting diaphragm is PLA(2) dependent. J Appl Physiol. 1999;87:792–800. doi: 10.1152/jappl.1999.87.2.792. [DOI] [PubMed] [Google Scholar]
- 53.Bindokas VP, Jordan J, Lee CC, Miller RJ. Superoxide production in rat hippocampal neurons: selective imaging with hydroethidine. J Neurosci. 1996;16:1324–1336. doi: 10.1523/JNEUROSCI.16-04-01324.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Carter WO, Narayanan PK, Robinson JP. Intracellular hydrogen peroxide and superoxide anion detection in endothelial cells. J Leukoc Biol. 1994;55:253–258. doi: 10.1002/jlb.55.2.253. [DOI] [PubMed] [Google Scholar]
- 55.Suzuki Y, Tanigaki T, Heimer D, Wang WZ, Ross WG, Sussman HH, Raffin TA. Polyethylene glycol-conjugated superoxide dismutase attenuates septic lung injury in guinea pigs. Am Rev Respir Dis. 1992;145:388–393. doi: 10.1164/ajrccm/145.2_Pt_1.388. [DOI] [PubMed] [Google Scholar]
- 56.Takeuchi T, Nakajima M, Morimoto K. Relationship between the intracellular reactive oxygen species and the induction of oxidative DNA damage in human neutrophil-like cells. Carcinogenesis. 1996;17:1543–1548. doi: 10.1093/carcin/17.8.1543. [DOI] [PubMed] [Google Scholar]
- 57.Moriguchi T, Seres T, Ravichandran V, Sasada M, Johnston RB., Jr Diamide primes neutrophils for enhanced release of superoxide anion: relationship to S-thiolation of cellular proteins. J Leukoc Biol. 1996;60:191–198. doi: 10.1002/jlb.60.2.191. [DOI] [PubMed] [Google Scholar]
- 58.Rojo AI, Salinas M, Martin D, Perona R, Cuadrado A. Regulation of Cu/Zn-superoxide dismutase expression via the phosphatidylinositol 3 kinase/Akt pathway and nuclear factor-kappaB. J Neurosci. 2004;24:7324–7334. doi: 10.1523/JNEUROSCI.2111-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Scanlon JM, Aizenman E, Reynolds IJ. Effects of pyrroloquinoline quinone on glutamate-induced production of reactive oxygen species in neurons. Eur J Pharmacol. 1997;326:67–74. doi: 10.1016/s0014-2999(97)00137-4. [DOI] [PubMed] [Google Scholar]
- 60.Andres D, Sanz N, Zaragoza A, Alvarez AM, Cascales M. Changes in antioxidant defence systems induced by cyclosporine A in cultures of hepatocytes from 2- and 12-month-old rats. Biochem Pharmacol. 2000;59:1091–1100. doi: 10.1016/s0006-2952(00)00233-1. [DOI] [PubMed] [Google Scholar]
- 61.Barbacanne MA, Souchard JP, Darblade B, Iliou JP, Nepveu F, Pipy B, Bayard F, Arnal JF. Detection of superoxide anion released extracellularly by endothelial cells using cytochrome c reduction, ESR, fluorescence and lucigenin-enhanced chemiluminescence techniques. Free Radic Biol Med. 2000;29:388–396. doi: 10.1016/s0891-5849(00)00336-1. [DOI] [PubMed] [Google Scholar]
- 62.Carriedo SG, Yin HZ, Sensi SL, Weiss JH. Rapid Ca2+ entry through Ca2+-permeable AMPA/Kainate channels triggers marked intracellular Ca2+ rises and consequent oxygen radical production. J Neurosci. 1998;18:7727–7738. doi: 10.1523/JNEUROSCI.18-19-07727.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gunnett CA, Heistad DD, Berg DJ, Faraci FM. IL-10 deficiency increases superoxide and endothelial dysfunction during inflammation. Am J Physiol Heart Circ Physiol. 2000;279:H1555–H1562. doi: 10.1152/ajpheart.2000.279.4.H1555. [DOI] [PubMed] [Google Scholar]
- 64.Sensi SL, Yin HZ, Carriedo SG, Rao SS, Weiss JH. Preferential Zn2+ influx through Ca2+-permeable AMPA/kainate channels triggers prolonged mitochondrial superoxide production. Proc Natl Acad Sci U S A. 1999;96:2414–2419. doi: 10.1073/pnas.96.5.2414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Sensi SL, Yin HZ, Weiss JH. Glutamate triggers preferential Zn2+ flux through Ca2+ permeable AMPA channels and consequent ROS production. Neuroreport. 1999;10:1723–1727. doi: 10.1097/00001756-199906030-00018. [DOI] [PubMed] [Google Scholar]
- 66.Sensi SL, Yin HZ, Weiss JH. AMPA/kainate receptor-triggered Zn2+ entry into cortical neurons induces mitochondrial Zn2+ uptake and persistent mitochondrial dysfunction. Eur J Neurosci. 2000;12:3813–3818. doi: 10.1046/j.1460-9568.2000.00277.x. [DOI] [PubMed] [Google Scholar]
- 67.Sha SH, Schacht J. Formation of reactive oxygen species following bioactivation of gentamicin. Free Radic Biol Med. 1999;26:341–347. doi: 10.1016/s0891-5849(98)00207-x. [DOI] [PubMed] [Google Scholar]
- 68.Takeuchi T, Kato N, Watanabe K, Morimoto K. Mechanism of oxidative DNA damage induction in a strict anaerobe, Prevotella melaninogenica. FEMS Microbiol Lett. 2000;192:133–138. doi: 10.1111/j.1574-6968.2000.tb09371.x. [DOI] [PubMed] [Google Scholar]
- 69.Zhao H, Kalivendi S, Zhang H, Joseph J, Nithipatikom K, Vasquez-Vivar J, Kalyanaraman B. Superoxide reacts with hydroethidine but forms a fluorescent product that is distinctly different from ethidium: potential implications in intracellular fluorescence detection of superoxide. Free Radic Biol Med. 2003;34:1359–1368. doi: 10.1016/s0891-5849(03)00142-4. [DOI] [PubMed] [Google Scholar]
- 70.Fridovich I. Editorial commentary on “Superoxide reacts with hydroethidine but forms a fluorescent product that is distinctly different from ethidium: potential implications in intracellular fluorescence detection of superoxide” by H. Zhao et al. Free Radic Biol Med. 2003;34:1357–1358. doi: 10.1016/s0891-5849(03)00174-6. [DOI] [PubMed] [Google Scholar]
- 71.Fink B, Laude K, McCann L, Doughan A, Harrison DG, Dikalov S. Detection of intracellular superoxide formation in endothelial cells and intact tissues using dihydroethidium and an HPLC-based assay. Am J Physiol Cell Physiol. 2004;287:C895–C902. doi: 10.1152/ajpcell.00028.2004. [DOI] [PubMed] [Google Scholar]
- 72.Zhao H, Joseph J, Fales HM, Sokoloski EA, Levine RL, Vasquez-Vivar J, Kalyanaraman B. Detection and characterization of the product of hydroethidine and intracellular superoxide by HPLC and limitations of fluorescence. Proc Natl Acad Sci U S A. 2005;102:5727–5732. doi: 10.1073/pnas.0501719102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Zielonka J, Srinivasan S, Hardy M, Ouari O, Lopez M, Vasquez-Vivar J, Avadhani NG, Kalyanaraman B. Cytochrome c-mediated oxidation of hydroethidine and mito-hydroethidine in mitochondria: identification of homo- and heterodimers. Free Radic Biol Med. 2008;44:835–846. doi: 10.1016/j.freeradbiomed.2007.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Fernandes DC, Wosniak J, Jr, Pescatore LA, Bertoline MA, Liberman M, Laurindo FR, Santos CX. Analysis of DHE-derived oxidation products by HPLC in the assessment of superoxide production and NADPH oxidase activity in vascular systems. Am J Physiol Cell Physiol. 2007;292:C413–C422. doi: 10.1152/ajpcell.00188.2006. [DOI] [PubMed] [Google Scholar]
- 75.Meany DL, Thompson L, Arriaga EA. Simultaneously monitoring the superoxide in the mitochondrial matrix and extramitochondrial space by micellar electrokinetic chromatography with laser-induced fluorescence. Anal Chem. 2007;79:4588–4594. doi: 10.1021/ac062252+. [DOI] [PubMed] [Google Scholar]
- 76.Zielonka J, Vasquez-Vivar J, Kalyanaraman B. The confounding effects of light, sonication, and Mn(III)TBAP on quantitation of superoxide using hydroethidine. Free Radic Biol Med. 2006;41:1050–1057. doi: 10.1016/j.freeradbiomed.2006.04.017. [DOI] [PubMed] [Google Scholar]
- 77.Robinson KM, Janes MS, Pehar M, Monette JS, Ross MF, Hagen TM, Murphy MP, Beckman JS. Selective fluorescent imaging of superoxide in vivo using ethidium-based probes. Proc Natl Acad Sci U S A. 2006;103:15038–15043. doi: 10.1073/pnas.0601945103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Harman GE, Stasz TE. Fluorescent vital stains for complementary labelling of protoplasts from Trichoderma spp. Stain Technol. 1988;63:241–247. doi: 10.3109/10520298809107190. [DOI] [PubMed] [Google Scholar]
- 79.Yu F, Sugawara T, Chan PH. Treatment with dihydroethidium reduces infarct size after transient focal cerebral ischemia in mice. Brain Res. 2003;978:223–227. doi: 10.1016/s0006-8993(03)02775-6. [DOI] [PubMed] [Google Scholar]
- 80.Zielonka J, Sarna T, Roberts JE, Wishart JF, Kalyanaraman B. Pulse radiolysis and steady-state analyses of the reaction between hydroethidine and superoxide and other oxidants. Arch Biochem Biophys. 2006;456:39–47. doi: 10.1016/j.abb.2006.09.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.De Iuliis GN, Wingate JK, Koppers AJ, McLaughlin EA, Aitken RJ. Definitive evidence for the nonmitochondrial production of superoxide anion by human spermatozoa. J Clin Endocrinol Metab. 2006;91:1968–1975. doi: 10.1210/jc.2005-2711. [DOI] [PubMed] [Google Scholar]
- 82.Nguyen SM, Alexejun CN, Levin LA. Amplification of a reactive oxygen species signal in axotomized retinal ganglion cells. Antioxid Redox Signal. 2003;5:629–634. doi: 10.1089/152308603770310293. [DOI] [PubMed] [Google Scholar]
- 83.Benov L, Sztejnberg L, Fridovich I. Critical evaluation of the use of hydroethidine as a measure of superoxide anion radical. Free Radic Biol Med. 1998;25:826–831. doi: 10.1016/s0891-5849(98)00163-4. [DOI] [PubMed] [Google Scholar]
- 84.Georgiou CD, Papapostolou I, Patsoukis N, Tsegenidis T, Sideris T. An ultrasensitive fluorescent assay for the in vivo quantification of superoxide radical in organisms. Anal Biochem. 2005;347:144–151. doi: 10.1016/j.ab.2005.09.013. [DOI] [PubMed] [Google Scholar]
- 85.Nethery D, DiMarco A, Stofan D, Supinski G. Sepsis increases contraction-related generation of reactive oxygen species in the diaphragm. J Appl Physiol. 1999;87:1279–1286. doi: 10.1152/jappl.1999.87.4.1279. [DOI] [PubMed] [Google Scholar]
- 86.Palazzolo-Ballance AM, Suquet C, Hurst JK. Pathways for intracellular generation of oxidants and tyrosine nitration by a macrophage cell line. Biochemistry. 2007;46:7536–7548. doi: 10.1021/bi700123s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Papapostolou I, Patsoukis N, Georgiou CD. The fluorescence detection of superoxide radical using hydroethidine could be complicated by the presence of heme proteins. Anal Biochem. 2004;332:290–298. doi: 10.1016/j.ab.2004.06.022. [DOI] [PubMed] [Google Scholar]
- 88.Patsoukis N, Papapostolou I, Georgiou CD. Interference of non-specific peroxidases in the fluorescence detection of superoxide radical by hydroethidine oxidation: a new assay for H2O2. Anal Bioanal Chem. 2005;381:1065–1072. doi: 10.1007/s00216-004-2999-x. [DOI] [PubMed] [Google Scholar]
- 89.Peshavariya HM, Dusting GJ, Selemidis S. Analysis of dihydroethidium fluorescence for the detection of intracellular and extracellular superoxide produced by NADPH oxidase. Free Radic Res. 2007;41:699–712. doi: 10.1080/10715760701297354. [DOI] [PubMed] [Google Scholar]
- 90.Shao ZH, Li CQ, Vanden Hoek TL, Becker LB, Schumacker PT, Wu JA, Attele AS, Yuan CS. Extract from Scutellaria baicalensis Georgi attenuates oxidant stress in cardiomyocytes. J Mol Cell Cardiol. 1999;31:1885–1895. doi: 10.1006/jmcc.1999.1021. [DOI] [PubMed] [Google Scholar]
- 91.Stepp DW, Ou J, Ackerman AW, Welak S, Klick D, Pritchard KA., Jr Native LDL and minimally oxidized LDL differentially regulate superoxide anion in vascular endothelium in situ. Am J Physiol Heart Circ Physiol. 2002;283:H750–H759. doi: 10.1152/ajpheart.00029.2002. [DOI] [PubMed] [Google Scholar]
- 92.Takano K, Sasada M, Harakawa N, Nogawa M, Asagoe K, Yamamoto K, Okuma M. Demonstration of functionally distinct human polymorphonuclear leukocyte fractions by simultaneous measurement of phagocytosis and oxygen radical generation. Int J Hematol. 1997;66:69–78. doi: 10.1016/s0925-5710(97)00577-x. [DOI] [PubMed] [Google Scholar]
- 93.Vanden Hoek TL, Li C, Shao Z, Schumacker PT, Becker LB. Significant levels of oxidants are generated by isolated cardiomyocytes during ischemia prior to reperfusion. J Mol Cell Cardiol. 1997;29:2571–2583. doi: 10.1006/jmcc.1997.0497. [DOI] [PubMed] [Google Scholar]
- 94.Zou AP, Li N, Cowley AW., Jr Production and actions of superoxide in the renal medulla. Hypertension. 2001;37:547–553. doi: 10.1161/01.hyp.37.2.547. [DOI] [PubMed] [Google Scholar]
- 95.Hawkins BJ, Madesh M, Kirkpatrick CJ, Fisher AB. Superoxide flux in endothelial cells via the chloride channel-3 mediates intracellular signaling. Mol Biol Cell. 2007;18:2002–2012. doi: 10.1091/mbc.E06-09-0830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Bizyukin A, Korkina L, Velichkovskii B. Comparative Use of 2,7-dichlorofluorescin diacetate, dihydrorhodamine 123, and hydroethidine to study oxidative metabolism in phagocytic cells. Bull Exp Biol Med. 1995;119:347–351. [PubMed] [Google Scholar]
- 97.Assimakopoulos SF, Mavrakis AG, Grintzalis K, Papapostolou I, Zervoudakis G, Konstantinou D, Chroni E, Vagianos CE, Georgiou C. Superoxide radical formation in diverse organs of rats with experimentally induced obstructive jaundice. Redox Rep. 2008;13:179–184. doi: 10.1179/135100008X308902. [DOI] [PubMed] [Google Scholar]
- 98.Georgiou CD, Papapostolou I, Grintzalis K. Superoxide radical detection in cells, tissues, organisms (animals, plants, insects, microorganisms) and soils. Nat Protoc. 2008;3:1679–1692. doi: 10.1038/nprot.2008.155. [DOI] [PubMed] [Google Scholar]
- 99.Zielonka J, Hardy M, Kalyanaraman B. HPLC study of oxidation products of hydroethidine in chemical and biological systems: ramifications in superoxide measurements. Free Radic Biol Med. 2009;46:329–338. doi: 10.1016/j.freeradbiomed.2008.10.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Posen Y, Kalchenko V, Seger R, Brandis A, Scherz A, Salomon Y. Manipulation of redox signaling in mammalian cells enabled by controlled photogeneration of reactive oxygen species. J Cell Sci. 2005;118:1957–1969. doi: 10.1242/jcs.02323. [DOI] [PubMed] [Google Scholar]
- 101.Germano G, Sanguigni V, Pignatelli P, Caccese D, Lenti L, Ragazzo M, Lauro R, Violi F. Enhanced platelet release of superoxide anion in systemic hypertension: role of AT1 receptors. J Hypertens. 2004;22:1151–1156. doi: 10.1097/00004872-200406000-00016. [DOI] [PubMed] [Google Scholar]
- 102.Swannell RPJ, Caplin R, Nedwell DB, Williamson FA. An investigation of hydroethidine as a fluorescent vital stain for prokaryotes. FEMS Microbiol Lett. 1992;101:173–182. [Google Scholar]
- 103.Bilski PJ, Karriker B, Chignell CF. Quenching and generation of singlet oxygen by hydroethidine and related chromophores. Chem Phys Lett. 2009;475:116–119. [Google Scholar]
- 104.Suhr D, Brummer F, Hulser DF. Cavitation-generated free radicals during shock wave exposure: investigations with cell-free solutions and suspended cells. Ultrasound Med Biol. 1991;17:761–768. doi: 10.1016/0301-5629(91)90158-s. [DOI] [PubMed] [Google Scholar]
- 105.Suhr D, Brummer F, Irmer U, Schlachter M, Hulser DF. Reduced cavitation-induced cellular damage by the antioxidative effect of vitamin E. Ultrasonics. 1994;32:301–307. doi: 10.1016/0041-624x(94)90010-8. [DOI] [PubMed] [Google Scholar]
- 106.Endl E, Steinbach P, Hofstadter F. Flow cytometric analysis of cell suspensions exposed to shock waves in the presence of the radical sensitive dye hydroethidine. Ultrasound Med Biol. 1995;21:569–577. doi: 10.1016/0301-5629(94)00133-x. [DOI] [PubMed] [Google Scholar]
- 107.Arzuaga X, Wassenberg D, Di GR, Elskus A. The chlorinated AHR ligand 3,3′,4,4′,5-pentachlorobiphenyl (PCB126) promotes reactive oxygen species (ROS) production during embryonic development in the killifish (Fundulus heteroclitus) Aquat Toxicol. 2006;76:13–23. doi: 10.1016/j.aquatox.2005.07.013. [DOI] [PubMed] [Google Scholar]
- 108.Grzeschik SM, Maier CM, Chan PH. Effects of cold injury-induced trauma in manganese superoxide dismutase-deficient mice. J Neurotrauma. 2003;20:571–581. doi: 10.1089/089771503767168492. [DOI] [PubMed] [Google Scholar]
- 109.Kim GW, Noshita N, Sugawara T, Chan PH. Early decrease in dna repair proteins, Ku70 and Ku86, and subsequent DNA fragmentation after transient focal cerebral ischemia in mice. Stroke. 2001;32:1401–1407. doi: 10.1161/01.str.32.6.1401. [DOI] [PubMed] [Google Scholar]
- 110.Kim GW, Gasche Y, Grzeschik S, Copin JC, Maier CM, Chan PH. Neurodegeneration in striatum induced by the mitochondrial toxin 3-nitropropionic acid: role of matrix metalloproteinase-9 in early blood-brain barrier disruption? J Neurosci. 2003;23:8733–8742. doi: 10.1523/JNEUROSCI.23-25-08733.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Lewen A, Sugawara T, Gasche Y, Fujimura M, Chan PH. Oxidative cellular damage and the reduction of APE/Ref-1 expression after experimental traumatic brain injury. Neurobiol Dis. 2001;8:380–390. doi: 10.1006/nbdi.2001.0396. [DOI] [PubMed] [Google Scholar]
- 112.Wang J, Tsirka SE. Neuroprotection by inhibition of matrix metalloproteinases in a mouse model of intracerebral haemorrhage. Brain. 2005;128:1622–1633. doi: 10.1093/brain/awh489. [DOI] [PubMed] [Google Scholar]
- 113.Wang J, Tsirka SE. Tuftsin fragment 1-3 is beneficial when delivered after the induction of intracerebral hemorrhage. Stroke. 2005;36:613–618. doi: 10.1161/01.STR.0000155729.12931.8f. [DOI] [PubMed] [Google Scholar]
- 114.Burnaugh L, Sabeur K, Ball BA. Generation of superoxide anion by equine spermatozoa as detected by dihydroethidium. Theriogenology. 2007;67:580–589. doi: 10.1016/j.theriogenology.2006.07.021. [DOI] [PubMed] [Google Scholar]
- 115.Luedtke NW, Liu Q, Tor Y. On the electronic structure of ethidium. Chemistry-A European Journal. 2005;11:495–508. doi: 10.1002/chem.200400559. [DOI] [PubMed] [Google Scholar]
- 116.Zielonka J, Vasquez-Vivar J, Kalyanaraman B. Detection of 2-hydroxyethidium in cellular systems: a unique marker product of superoxide and hydroethidine. Nat Protoc. 2008;3:8–21. doi: 10.1038/nprot.2007.473. [DOI] [PubMed] [Google Scholar]
- 117.Greenstock CL, Ruddock GW. Interaction of ethidium bromide with DNA as studied by kinetic spectrophotometry. Chem Biol Interact. 1975;11:441–447. doi: 10.1016/0009-2797(75)90011-3. [DOI] [PubMed] [Google Scholar]
- 118.Meyer-Almes FJ, Porschke D. Mechanism of intercalation into the DNA double helix by ethidium. Biochemistry. 1993;32:4246–4253. doi: 10.1021/bi00067a012. [DOI] [PubMed] [Google Scholar]
- 119.Bonasera V, Alberti S, Sacchetti A. Protocol for high-sensitivity/long linear-range spectrofluorimetric DNA quantification using ethidium bromide. BioTechniques. 2007;43:173–4. 176. doi: 10.2144/000112500. [DOI] [PubMed] [Google Scholar]
- 120.Olmsted J, III, Kearns DR. Mechanism of ethidium bromide fluorescence enhancement on binding to nucleic acids. Biochemistry. 1977;16:3647–3654. doi: 10.1021/bi00635a022. [DOI] [PubMed] [Google Scholar]
- 121.Atherton SJ, Beaumont PC. The reaction of ethidium bromide with oxidising free radicals: Pulse radiolysis in water and DNA. Intl J Radiat Appl Instrum Part C Radiat Phys Chem. 1990;36:819–824. [Google Scholar]
- 122.Castilho RF, Ward MW, Nicholls DG. Oxidative stress, mitochondrial function, and acute glutamate excitotoxicity in cultured cerebellar granule cells. J Neurochem. 1999;72:1394–1401. doi: 10.1046/j.1471-4159.1999.721394.x. [DOI] [PubMed] [Google Scholar]
- 123.Hu X, Wang Q, He P, Fang Y. Spectroelectrochemistry study on the electrochemical reduction of ethidium bromide. Anal Sci. 2002;18:645–650. doi: 10.2116/analsci.18.645. [DOI] [PubMed] [Google Scholar]
- 124.Tollefson KE, Kroczynski J, Cutaia MV. Time-dependent interactions of oxidant-sensitive fluoroprobes with inhibitors of cellular metabolism. Lab Invest. 2003;83:367–375. doi: 10.1097/01.lab.0000059934.53602.4f. [DOI] [PubMed] [Google Scholar]
- 125.Pasternack RF, Caccam M, Keogh B, Stephenson TA, Williams AP, Gibbs EJ. Long-range fluorescence quenching of ethidium ion by cationic porphyrins in the presence of DNA. J Am Chem Soc. 1991;113:6835–6840. [Google Scholar]
- 126.Reinhardt CG, Krugh TR. A comparative study of ethidium bromide complexes with dinucleotides and DNA: direct evidence for intercalation and nucleic acid sequence preferences. Biochemistry. 1978;17:4845–4854. doi: 10.1021/bi00616a001. [DOI] [PubMed] [Google Scholar]
- 127.Davies DB, Veselkov DA, Djimant LN, Veselkov AN. Hetero-association of caffeine and aromatic drugs and their competitive binding with a DNA oligomer. Eur Biophys J. 2001;30:354–366. doi: 10.1007/s002490100150. [DOI] [PubMed] [Google Scholar]
- 128.Davies DB, Veselkov DA, Veselkov AN. NMR determination of the hetero-association of phenanthridines with daunomycin and their competitive binding to a DNA oligomer. Eur Biophys J. 2002;31:153–162. doi: 10.1007/s00249-001-0199-6. [DOI] [PubMed] [Google Scholar]
- 129.Zielonka J, Kalyanaraman B. “ROS-generating mitochondrial DNA mutations can regulate tumor cell metastasis”--a critical commentary. Free Radic Biol Med. 2008;45:1217–1219. doi: 10.1016/j.freeradbiomed.2008.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Robinson KM, Janes MS, Beckman JS. The selective detection of mitochondrial superoxide by live cell imaging. Nat Protoc. 2008;3:941–947. doi: 10.1038/nprot.2008.56. [DOI] [PubMed] [Google Scholar]
- 131.Verkaart S, Koopman WJ, Cheek J, van Emst-de Vries SE, van den Heuvel LW, Smeitink JA, Willems PH. Mitochondrial and cytosolic thiol redox state are not detectably altered in isolated human NADH:ubiquinone oxidoreductase deficiency. Biochim Biophys Acta. 2007;1772:1041–1051. doi: 10.1016/j.bbadis.2007.05.004. [DOI] [PubMed] [Google Scholar]
- 132.Aitken RJ, Wingate JK, De Iuliis GN, Koppers AJ, McLaughlin EA. Cis-unsaturated fatty acids stimulate reactive oxygen species generation and lipid peroxidation in human spermatozoa. J Clin Endocrinol Metab. 2006;91:4154–4163. doi: 10.1210/jc.2006-1309. [DOI] [PubMed] [Google Scholar]
- 133.Al-Enezi KS, Alkhalaf M, Benov LT. Glycolaldehyde induces growth inhibition and oxidative stress in human breast cancer cells. Free Radic Biol Med. 2006;40:1144–1151. doi: 10.1016/j.freeradbiomed.2005.10.065. [DOI] [PubMed] [Google Scholar]
- 134.Ceravolo GS, Fernandes L, Munhoz CD, Fernandes DC, Tostes RC, Laurindo FR, Scavone C, Fortes ZB, Carvalho MH. Angiotensin II chronic infusion induces B1 receptor expression in aorta of rats. Hypertension. 2007;50:756–761. doi: 10.1161/HYPERTENSIONAHA.107.094706. [DOI] [PubMed] [Google Scholar]
- 135.Clavreul N, Bachschmid MM, Hou X, Shi C, Idrizovic A, Ido Y, Pimentel D, Cohen RA. S-glutathiolation of p21ras by peroxynitrite mediates endothelial insulin resistance caused by oxidized low-density lipoprotein. Arterioscler Thromb Vasc Biol. 2006;26:2454–2461. doi: 10.1161/01.ATV.0000242791.28953.4c. [DOI] [PubMed] [Google Scholar]
- 136.Doughan AK, Harrison DG, Dikalov SI. Molecular mechanisms of angiotensin II-mediated mitochondrial dysfunction: linking mitochondrial oxidative damage and vascular endothelial dysfunction. Circ Res. 2008;102:488–496. doi: 10.1161/CIRCRESAHA.107.162800. [DOI] [PubMed] [Google Scholar]
- 137.Gray JP, Heck DE, Mishin V, Smith PJ, Hong JY, Thiruchelvam M, Cory-Slechta DA, Laskin DL, Laskin JD. Paraquat increases cyanide-insensitive respiration in murine lung epithelial cells by activating an NAD(P)H:paraquat oxidoreductase: identification of the enzyme as thioredoxin reductase. J Biol Chem. 2007;282:7939–7949. doi: 10.1074/jbc.M611817200. [DOI] [PubMed] [Google Scholar]
- 138.Guzik TJ, Hoch NE, Brown KA, McCann LA, Rahman A, Dikalov S, Goronzy J, Weyand C, Harrison DG. Role of the T cell in the genesis of angiotensin II induced hypertension and vascular dysfunction. J Exp Med. 2007;204:2449–2460. doi: 10.1084/jem.20070657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Hao L, Nishimura T, Wo H, Fernandez-Patron C. Vascular responses to alpha1-adrenergic receptors in small rat mesenteric arteries depend on mitochondrial reactive oxygen species. Arterioscler Thromb Vasc Biol. 2006;26:819–825. doi: 10.1161/01.ATV.0000204344.90301.7c. [DOI] [PubMed] [Google Scholar]
- 140.Huh J, Liepins A, Zielonka J, Andrekopoulos C, Kalyanaraman B, Sorokin A. Cyclooxygenase 2 rescues LNCaP prostate cancer cells from sanguinarine-induced apoptosis by a mechanism involving inhibition of nitric oxide synthase activity. Cancer Res. 2006;66:3726–3736. doi: 10.1158/0008-5472.CAN-05-4033. [DOI] [PubMed] [Google Scholar]
- 141.Nath KA, d'Uscio LV, Juncos JP, Croatt AJ, Manriquez MC, Pittock ST, Katusic ZS. An analysis of the DOCA-salt model of hypertension in HO-1-/- mice and the Gunn rat. Am J Physiol Heart Circ Physiol. 2007;293:H333–H342. doi: 10.1152/ajpheart.00870.2006. [DOI] [PubMed] [Google Scholar]
- 142.Shang T, Kotamraju S, Zhao H, Kalivendi SV, Hillard CJ, Kalyanaraman B. Sepiapterin attenuates 1-methyl-4-phenylpyridinium-induced apoptosis in neuroblastoma cells transfected with neuronal NOS: role of tetrahydrobiopterin, nitric oxide, and proteasome activation. Free Radic Biol Med. 2005;39:1059–1074. doi: 10.1016/j.freeradbiomed.2005.05.022. [DOI] [PubMed] [Google Scholar]
- 143.Slane BG, Aykin-Burns N, Smith BJ, Kalen AL, Goswami PC, Domann FE, Spitz DR. Mutation of succinate dehydrogenase subunit C results in increased O2.-, oxidative stress, and genomic instability. Cancer Res. 2006;66:7615–7620. doi: 10.1158/0008-5472.CAN-06-0833. [DOI] [PubMed] [Google Scholar]
- 144.Whitsett J, Martasek P, Zhao H, Schauer DW, Hatakeyama K, Kalyanaraman B, Vasquez-Vivar J. Endothelial cell superoxide anion radical generation is not dependent on endothelial nitric oxide synthase-serine 1179 phosphorylation and endothelial nitric oxide synthase dimer/monomer distribution. Free Radic Biol Med. 2006;40:2056–2068. doi: 10.1016/j.freeradbiomed.2006.02.001. [DOI] [PubMed] [Google Scholar]
- 145.Widder JD, Guzik TJ, Mueller CF, Clempus RE, Schmidt HH, Dikalov SI, Griendling KK, Jones DP, Harrison DG. Role of the multidrug resistance protein-1 in hypertension and vascular dysfunction caused by angiotensin II. Arterioscler Thromb Vasc Biol. 2007;27:762–768. doi: 10.1161/01.ATV.0000259298.11129.a2. [DOI] [PubMed] [Google Scholar]
- 146.Doughan AK, Dikalov SI. Mitochondrial redox cycling of mitoquinone leads to superoxide production and cellular apoptosis. Antioxid Redox Signal. 2007;9:1825–1836. doi: 10.1089/ars.2007.1693. [DOI] [PubMed] [Google Scholar]
- 147.Fernandes DC, Manoel AH, Wosniak J, Jr, Laurindo FR. Protein disulfide isomerase overexpression in vascular smooth muscle cells induces spontaneous preemptive NADPH oxidase activation and Nox1 mRNA expression: effects of nitrosothiol exposure. Arch Biochem Biophys. 2009;484:197–204. doi: 10.1016/j.abb.2009.01.022. [DOI] [PubMed] [Google Scholar]
- 148.Laurindo FR, Fernandes DC, Santos CX. Assessment of superoxide production and NADPH oxidase activity by HPLC analysis of dihydroethidium oxidation products. Methods Enzymol. 2008;441:237–260. doi: 10.1016/S0076-6879(08)01213-5. [DOI] [PubMed] [Google Scholar]
- 149.Sampath V, Radish AC, Eis AL, Broniowska K, Hogg N, Konduri GG. Attenuation of lipopolysaccharide-induced oxidative stress and apoptosis in fetal pulmonary artery endothelial cells by hypoxia. Free Radic Biol Med. 2009;46:663–671. doi: 10.1016/j.freeradbiomed.2008.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Maghzal GJ, Stocker R. Improved analysis of hydroethidine and 2-hydroxyethidium by HPLC and electrochemical detection. Free Radic Biol Med. 2007;43:1095–1096. doi: 10.1016/j.freeradbiomed.2007.06.023. [DOI] [PubMed] [Google Scholar]
- 151.Vasquez-Vivar J, Whitsett J, Ionova I, Konorev E, Zielonka J, Kalyanaraman B, Shi Y, Pieper GM. Cytokines and lipopolysaccharides induce inducible nitric oxide synthase but not enzyme activity in adult rat cardiomyocytes. Free Radic Biol Med. 2008;45:994–1001. doi: 10.1016/j.freeradbiomed.2008.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Whitsett J, Picklo MJ, Sr, Vasquez-Vivar J. 4-Hydroxy-2-nonenal increases superoxide anion radical in endothelial cells via stimulated GTP cyclohydrolase proteasomal degradation. Arterioscler Thromb Vasc Biol. 2007;27:2340–2347. doi: 10.1161/ATVBAHA.107.153742. [DOI] [PubMed] [Google Scholar]
- 153.Maghzal GJ, Thomas SR, Hunt NH, Stocker R. Cytochrome b5, not superoxide anion radical, is a major reductant of indoleamine 2,3-dioxygenase in human cells. J Biol Chem. 2008;283:12014–12025. doi: 10.1074/jbc.M710266200. [DOI] [PubMed] [Google Scholar]
- 154.Amour J, Brzezinska AK, Weihrauch D, Billstrom AR, Zielonka J, Krolikowski JG, Bienengraeber MW, Warltier DC, Pratt PF, Jr, Kersten JR. Role of heat shock protein 90 and endothelial nitric oxide synthase during early anesthetic and ischemic preconditioning. Anesthesiology. 2009;110:317–325. doi: 10.1097/ALN.0b013e3181942cb4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Tang PC, Qin L, Zielonka J, Zhou J, Matte-Martone C, Bergaya S, van RN, Shlomchik WD, Min W, Sessa WC, Pober JS, Tellides G. MyD88-dependent, superoxide-initiated inflammation is necessary for flow-mediated inward remodeling of conduit arteries. J Exp Med. 2008;205:3159–3171. doi: 10.1084/jem.20081298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Rossary A, Arab K, Steghens JP. Polyunsaturated fatty acids modulate NOX 4 anion superoxide production in human fibroblasts. Biochem J. 2007;406:77–83. doi: 10.1042/BJ20061009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Meany DL, Poe BG, Navratil M, Moraes CT, Arriaga EA. Superoxide released into the mitochondrial matrix. Free Radic Biol Med. 2006;41:950–959. doi: 10.1016/j.freeradbiomed.2006.06.003. [DOI] [PubMed] [Google Scholar]
- 158.Xu X, Arriaga EA. Qualitative determination of superoxide release at both sides of the mitochondrial inner membrane by capillary electrophoretic analysis of the oxidation products of triphenylphosphonium hydroethidine. Free Radic Biol Med. 2009;46:905–913. doi: 10.1016/j.freeradbiomed.2008.12.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Schlezinger JJ, White RD, Stegeman JJ. Oxidative inactivation of cytochrome P-450 1A (CYP1A) stimulated by 3,3′,4,4′-tetrachlorobiphenyl: production of reactive oxygen by vertebrate CYP1As. Mol Pharmacol. 1999;56:588–597. doi: 10.1124/mol.56.3.588. [DOI] [PubMed] [Google Scholar]
- 160.Schlezinger JJ, Stegeman JJ. Induction and suppression of cytochrome P450 1A by 3,3′,4,4′,5-pentachlorobiphenyl and its relationship to oxidative stress in the marine fish scup (Stenotomus chrysops) Aquat Toxicol. 2001;52:101–115. doi: 10.1016/s0166-445x(00)00141-7. [DOI] [PubMed] [Google Scholar]
- 161.Daiber A, Oelze M, August M, Wendt M, Sydow K, Wieboldt H, Kleschyov AL, Munzel T. Detection of superoxide and peroxynitrite in model systems and mitochondria by the luminol analogue L-012. Free Radic Res. 2004;38:259–269. doi: 10.1080/10715760410001659773. [DOI] [PubMed] [Google Scholar]
- 162.Helmerhorst EJ, Troxler RF, Oppenheim FG. The human salivary peptide histatin 5 exerts its antifungal activity through the formation of reactive oxygen species. Proc Natl Acad Sci U S A. 2001;98:14637–14642. doi: 10.1073/pnas.141366998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Muller FL, Liu Y, Van RH. Complex III releases superoxide to both sides of the inner mitochondrial membrane. J Biol Chem. 2004;279:49064–49073. doi: 10.1074/jbc.M407715200. [DOI] [PubMed] [Google Scholar]
- 164.Chan PH, Kawase M, Murakami K, Chen SF, Li Y, Calagui B, Reola L, Carlson E, Epstein CJ. Overexpression of SOD1 in transgenic rats protects vulnerable neurons against ischemic damage after global cerebral ischemia and reperfusion. J Neurosci. 1998;18:8292–8299. doi: 10.1523/JNEUROSCI.18-20-08292.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Girouard H, Park L, Anrather J, Zhou P, Iadecola C. Angiotensin II attenuates endothelium-dependent responses in the cerebral microcirculation through nox-2-derived radicals. Arterioscler Thromb Vasc Biol. 2006;26:826–832. doi: 10.1161/01.ATV.0000205849.22807.6e. [DOI] [PubMed] [Google Scholar]
- 166.Iwanami J, Mogi M, Okamoto S, Gao XY, Li JM, Min LJ, Ide A, Tsukuda K, Iwai M, Horiuchi M. Pretreatment with eplerenone reduces stroke volume in mouse middle cerebral artery occlusion model. Eur J Pharmacol. 2007;566:153–159. doi: 10.1016/j.ejphar.2007.03.043. [DOI] [PubMed] [Google Scholar]
- 167.Kamada H, Yu F, Nito C, Chan PH. Influence of hyperglycemia on oxidative stress and matrix metalloproteinase-9 activation after focal cerebral ischemia/reperfusion in rats: relation to blood-brain barrier dysfunction. Stroke. 2007;38:1044–1049. doi: 10.1161/01.STR.0000258041.75739.cb. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Kovacs R, Schuchmann S, Gabriel S, Kardos J, Heinemann U. Ca2+ signalling and changes of mitochondrial function during low-Mg2+-induced epileptiform activity in organotypic hippocampal slice cultures. Eur J Neurosci. 2001;13:1311–1319. doi: 10.1046/j.0953-816x.2001.01505.x. [DOI] [PubMed] [Google Scholar]
- 169.Mehta SH, Webb RC, Ergul A, Tawfik A, Dorrance AM. Neuroprotection by tempol in a model of iron-induced oxidative stress in acute ischemic stroke. Am J Physiol Regul Integr Comp Physiol. 2004;286:R283–R288. doi: 10.1152/ajpregu.00446.2002. [DOI] [PubMed] [Google Scholar]
- 170.Mogi M, Iwai M, Chen R, Iwanami J, Ide A, Tsukuda K, Yoshii T, Horiuchi M. Amlodipine treatment reduces stroke size in apolipoprotein E-deficient mice. Am J Hypertens. 2006;19:1144–1149. doi: 10.1016/j.amjhyper.2006.04.009. [DOI] [PubMed] [Google Scholar]
- 171.Mogi M, Tsukuda K, Li JM, Iwanami J, Min LJ, Sakata A, Fujita T, Iwai M, Horiuchi M. Inhibition of cognitive decline in mice fed a high-salt and cholesterol diet by the angiotensin receptor blocker, olmesartan. Neuropharmacology. 2007;53:899–905. doi: 10.1016/j.neuropharm.2007.08.020. [DOI] [PubMed] [Google Scholar]
- 172.Shichinohe H, Kuroda S, Yasuda H, Ishikawa T, Iwai M, Horiuchi M, Iwasaki Y. Neuroprotective effects of the free radical scavenger Edaravone (MCI-186) in mice permanent focal brain ischemia. Brain Res. 2004;1029:200–206. doi: 10.1016/j.brainres.2004.09.055. [DOI] [PubMed] [Google Scholar]
- 173.Uchida K, Samejima M, Okabe A, Fukuda A. Neuroprotective effects of melatonin against anoxia/aglycemia stress, as assessed by synaptic potentials and superoxide production in rat hippocampal slices. J Pineal Res. 2004;37:215–222. doi: 10.1111/j.1600-079X.2004.00159.x. [DOI] [PubMed] [Google Scholar]
- 174.Luo J, Li N, Robinson JP, Shi R. The increase of reactive oxygen species and their inhibition in an isolated guinea pig spinal cord compression model. Spinal Cord. 2002;40:656–665. doi: 10.1038/sj.sc.3101363. [DOI] [PubMed] [Google Scholar]
- 175.Sanchez-Carbente MR, Castro-Obregon S, Covarrubias L, Narvaez V. Motoneuronal death during spinal cord development is mediated by oxidative stress. Cell Death Differ. 2005;12:279–291. doi: 10.1038/sj.cdd.4401560. [DOI] [PubMed] [Google Scholar]
- 176.Wu DC, Re DB, Nagai M, Ischiropoulos H, Przedborski S. The inflammatory NADPH oxidase enzyme modulates motor neuron degeneration in amyotrophic lateral sclerosis mice. Proc Natl Acad Sci U S A. 2006;103:12132–12137. doi: 10.1073/pnas.0603670103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Al-Benna S, Hamilton CA, McClure JD, Rogers PN, Berg GA, Ford I, Delles C, Dominiczak AF. Low-density lipoprotein cholesterol determines oxidative stress and endothelial dysfunction in saphenous veins from patients with coronary artery disease. Arterioscler Thromb Vasc Biol. 2006;26:218–223. doi: 10.1161/01.ATV.0000193626.22269.45. [DOI] [PubMed] [Google Scholar]
- 178.Andersson IJ, Johansson ME, Wickman A, Bohlooly Y, Klintland N, Caidahl K, Gustafsson M, Boren J, Gan LM, Bergstrom G. Endothelial dysfunction in growth hormone transgenic mice. Clin Sci (Lond) 2006;110:217–225. doi: 10.1042/CS20050281. [DOI] [PubMed] [Google Scholar]
- 179.August M, Wingerter O, Oelze M, Wenzel P, Kleschyov AL, Daiber A, Mulsch A, Munzel T, Tsilimingas N. Mechanisms underlying dysfunction of carotid arteries in genetically hyperlipidemic rabbits. Nitric Oxide. 2006;15:241–251. doi: 10.1016/j.niox.2006.02.003. [DOI] [PubMed] [Google Scholar]
- 180.Azumi H, Inoue N, Ohashi Y, Terashima M, Mori T, Fujita H, Awano K, Kobayashi K, Maeda K, Hata K, Shinke T, Kobayashi S, Hirata K, Kawashima S, Itabe H, Hayashi Y, Imajoh-Ohmi S, Itoh H, Yokoyama M. Superoxide generation in directional coronary atherectomy specimens of patients with angina pectoris: important role of NAD(P)H oxidase. Arterioscler Thromb Vasc Biol. 2002;22:1838–1844. doi: 10.1161/01.atv.0000037101.40667.62. [DOI] [PubMed] [Google Scholar]
- 181.Banes-Berceli AK, Ogobi S, Tawfik A, Patel B, Shirley A, Pollock DM, Fulton D, Marrero MB. Endothelin-1 activation of JAK2 in vascular smooth muscle cells involves NAD(P)H oxidase-derived reactive oxygen species. Vascul Pharmacol. 2005;43:310–319. doi: 10.1016/j.vph.2005.08.024. [DOI] [PubMed] [Google Scholar]
- 182.Bleeke T, Zhang H, Madamanchi N, Patterson C, Faber JE. Catecholamine-induced vascular wall growth is dependent on generation of reactive oxygen species. Circ Res. 2004;94:37–45. doi: 10.1161/01.RES.0000109412.80157.7D. [DOI] [PubMed] [Google Scholar]
- 183.Brandes RP, Miller FJ, Beer S, Haendeler J, Hoffmann J, Ha T, Holland SM, Gorlach A, Busse R. The vascular NADPH oxidase subunit p47phox is involved in redox-mediated gene expression. Free Radic Biol Med. 2002;32:1116–1122. doi: 10.1016/s0891-5849(02)00789-x. [DOI] [PubMed] [Google Scholar]
- 184.Bubolz AH, Li H, Wu Q, Liu Y. Enhanced oxidative stress impairs cAMP-mediated dilation by reducing Kv channel function in small coronary arteries of diabetic rats. Am J Physiol Heart Circ Physiol. 2005;289:H1873–H1880. doi: 10.1152/ajpheart.00357.2005. [DOI] [PubMed] [Google Scholar]
- 185.Cai S, Khoo J, Mussa S, Alp NJ, Channon KM. Endothelial nitric oxide synthase dysfunction in diabetic mice: importance of tetrahydrobiopterin in eNOS dimerisation. Diabetologia. 2005;48:1933–1940. doi: 10.1007/s00125-005-1857-5. [DOI] [PubMed] [Google Scholar]
- 186.Chamseddine AH, Miller FJ., Jr Gp91phox contributes to NADPH oxidase activity in aortic fibroblasts but not smooth muscle cells. Am J Physiol Heart Circ Physiol. 2003;285:H2284–H2289. doi: 10.1152/ajpheart.00459.2003. [DOI] [PubMed] [Google Scholar]
- 187.Chen C, Korshunov VA, Massett MP, Yan C, Berk BC. Impaired vasorelaxation in inbred mice is associated with alterations in both nitric oxide and super oxide pathways. J Vasc Res. 2007;44:504–512. doi: 10.1159/000106751. [DOI] [PubMed] [Google Scholar]
- 188.Christensen FH, Stankevicius E, Hansen T, Jorgensen MM, Valverde VL, Simonsen U, Buus NH. Flow- and acetylcholine-induced dilatation in small arteries from rats with renovascular hypertension--effect of tempol treatment. Eur J Pharmacol. 2007;566:160–166. doi: 10.1016/j.ejphar.2007.03.058. [DOI] [PubMed] [Google Scholar]
- 189.Collin B, Busseuil D, Zeller M, Perrin C, Barthez O, Duvillard L, Vergely C, Bardou M, Dumas M, Cottin Y, Rochette L. Increased superoxide anion production is associated with early atherosclerosis and cardiovascular dysfunctions in a rabbit model. Mol Cell Biochem. 2007;294:225–235. doi: 10.1007/s11010-006-9263-y. [DOI] [PubMed] [Google Scholar]
- 190.Conklin BS, Fu W, Lin PH, Lumsden AB, Yao Q, Chen C. HIV protease inhibitor ritonavir decreases endothelium-dependent vasorelaxation and increases superoxide in porcine arteries. Cardiovasc Res. 2004;63:168–175. doi: 10.1016/j.cardiores.2004.03.020. [DOI] [PubMed] [Google Scholar]
- 191.Csiszar A, Labinskyy N, Smith K, Rivera A, Orosz Z, Ungvari Z. Vasculoprotective effects of anti-tumor necrosis factor-alpha treatment in aging. Am J Pathol. 2007;170:388–398. doi: 10.2353/ajpath.2007.060708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.d'Uscio LV, Baker TA, Mantilla CB, Smith L, Weiler D, Sieck GC, Katusic ZS. Mechanism of endothelial dysfunction in apolipoprotein E-deficient mice. Arterioscler Thromb Vasc Biol. 2001;21:1017–1022. doi: 10.1161/01.atv.21.6.1017. [DOI] [PubMed] [Google Scholar]
- 193.De AA, Rinaldi B, Capuano A, Rossi F, Filippelli A. Indomethacin potentiates acetylcholine-induced vasodilation by increasing free radical production. Br J Pharmacol. 2004;142:1233–1240. doi: 10.1038/sj.bjp.0705877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Dick GM, Katz PS, Farias M, III, Morris M, James J, Knudson JD, Tune JD. Resistin impairs endothelium-dependent dilation to bradykinin, but not acetylcholine, in the coronary circulation. Am J Physiol Heart Circ Physiol. 2006;291:H2997–H3002. doi: 10.1152/ajpheart.01035.2005. [DOI] [PubMed] [Google Scholar]
- 195.Didion SP, Ryan MJ, Baumbach GL, Sigmund CD, Faraci FM. Superoxide contributes to vascular dysfunction in mice that express human renin and angiotensinogen. Am J Physiol Heart Circ Physiol. 2002;283:H1569–H1576. doi: 10.1152/ajpheart.00079.2002. [DOI] [PubMed] [Google Scholar]
- 196.Didion SP, Ryan MJ, Didion LA, Fegan PE, Sigmund CD, Faraci FM. Increased superoxide and vascular dysfunction in CuZnSOD-deficient mice. Circ Res. 2002;91:938–944. doi: 10.1161/01.res.0000043280.65241.04. [DOI] [PubMed] [Google Scholar]
- 197.Ding H, Hashem M, Triggle C. Increased oxidative stress in the streptozotocin-induced diabetic apoE-deficient mouse: changes in expression of NADPH oxidase subunits and eNOS. Eur J Pharmacol. 2007;561:121–128. doi: 10.1016/j.ejphar.2006.12.034. [DOI] [PubMed] [Google Scholar]
- 198.Duncan ER, Walker SJ, Ezzat VA, Wheatcroft SB, Li JM, Shah AM, Kearney MT. Accelerated endothelial dysfunction in mild prediabetic insulin resistance: the early role of reactive oxygen species. Am J Physiol Endocrinol Metab. 2007;293:E1311–E1319. doi: 10.1152/ajpendo.00299.2007. [DOI] [PubMed] [Google Scholar]
- 199.Elmarakby AA, Loomis ED, Pollock JS, Pollock DM. NADPH oxidase inhibition attenuates oxidative stress but not hypertension produced by chronic ET-1. Hypertension. 2005;45:283–287. doi: 10.1161/01.HYP.0000153051.56460.6a. [DOI] [PubMed] [Google Scholar]
- 200.Ergul A, Johansen JS, Stromhaug C, Harris AK, Hutchinson J, Tawfik A, Rahimi A, Rhim E, Wells B, Caldwell RW, Anstadt MP. Vascular dysfunction of venous bypass conduits is mediated by reactive oxygen species in diabetes: role of endothelin-1. J Pharmacol Exp Ther. 2005;313:70–77. doi: 10.1124/jpet.104.078105. [DOI] [PubMed] [Google Scholar]
- 201.Fiordaliso F, Cuccovillo I, Bianchi R, Bai A, Doni M, Salio M, De AN, Ghezzi P, Latini R, Masson S. Cardiovascular oxidative stress is reduced by an ACE inhibitor in a rat model of streptozotocin-induced diabetes. Life Sci. 2006;79:121–129. doi: 10.1016/j.lfs.2005.12.036. [DOI] [PubMed] [Google Scholar]
- 202.Fresquet F, Pourageaud F, Leblais V, Brandes RP, Savineau JP, Marthan R, Muller B. Role of reactive oxygen species and gp91phox in endothelial dysfunction of pulmonary arteries induced by chronic hypoxia. Br J Pharmacol. 2006;148:714–723. doi: 10.1038/sj.bjp.0706779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Frisbee JC, Stepp DW. Impaired NO-dependent dilation of skeletal muscle arterioles in hypertensive diabetic obese Zucker rats. Am J Physiol Heart Circ Physiol. 2001;281:H1304–H1311. doi: 10.1152/ajpheart.2001.281.3.H1304. [DOI] [PubMed] [Google Scholar]
- 204.Frisbee JC, Maier KG, Stepp DW. Oxidant stress-induced increase in myogenic activation of skeletal muscle resistance arteries in obese Zucker rats. Am J Physiol Heart Circ Physiol. 2002;283:H2160–H2168. doi: 10.1152/ajpheart.00379.2002. [DOI] [PubMed] [Google Scholar]
- 205.Gunnett CA, Heistad DD, Faraci FM. Interleukin-10 protects nitric oxide-dependent relaxation during diabetes: role of superoxide. Diabetes. 2002;51:1931–1937. doi: 10.2337/diabetes.51.6.1931. [DOI] [PubMed] [Google Scholar]
- 206.Guzik TJ, Sadowski J, Guzik B, Jopek A, Kapelak B, Przybylowski P, Wierzbicki K, Korbut R, Harrison DG, Channon KM. Coronary artery superoxide production and nox isoform expression in human coronary artery disease. Arterioscler Thromb Vasc Biol. 2006;26:333–339. doi: 10.1161/01.ATV.0000196651.64776.51. [DOI] [PubMed] [Google Scholar]
- 207.Haruna Y, Morita Y, Yada T, Satoh M, Fox DA, Kashihara N. Fluvastatin reverses endothelial dysfunction and increased vascular oxidative stress in rat adjuvant-induced arthritis. Arthritis Rheum. 2007;56:1827–1835. doi: 10.1002/art.22632. [DOI] [PubMed] [Google Scholar]
- 208.Hatoum OA, Binion DG, Phillips SA, O'Loughlin C, Komorowski RA, Gutterman DD, Otterson MF. Radiation induced small bowel “web” formation is associated with acquired microvascular dysfunction. Gut. 2005;54:1797–1800. doi: 10.1136/gut.2005.073734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Hatoum OA, Otterson MF, Kopelman D, Miura H, Sukhotnik I, Larsen BT, Selle RM, Moulder JE, Gutterman DD. Radiation induces endothelial dysfunction in murine intestinal arterioles via enhanced production of reactive oxygen species. Arterioscler Thromb Vasc Biol. 2006;26:287–294. doi: 10.1161/01.ATV.0000198399.40584.8c. [DOI] [PubMed] [Google Scholar]
- 210.Herrmann J, Saguner AM, Versari D, Peterson TE, Chade A, Olson M, Lerman LO, Lerman A. Chronic proteasome inhibition contributes to coronary atherosclerosis. Circ Res. 2007;101:865–874. doi: 10.1161/CIRCRESAHA.107.152959. [DOI] [PubMed] [Google Scholar]
- 211.Iglarz M, Touyz RM, Viel EC, Amiri F, Schiffrin EL. Involvement of oxidative stress in the profibrotic action of aldosterone. Interaction wtih the renin-angiotension system. Am J Hypertens. 2004;17:597–603. [PubMed] [Google Scholar]
- 212.Kimura S, Zhang GX, Nishiyama A, Nagai Y, Nakagawa T, Miyanaka H, Fujisawa Y, Miyatake A, Nagai T, Tokuda M, Abe Y. D-allose, an all-cis aldo-hexose, suppresses development of salt-induced hypertension in Dahl rats. J Hypertens. 2005;23:1887–1894. doi: 10.1097/01.hjh.0000182523.29193.e3. [DOI] [PubMed] [Google Scholar]
- 213.Kin H, Zhao ZQ, Sun HY, Wang NP, Corvera JS, Halkos ME, Kerendi F, Guyton RA, Vinten-Johansen J. Postconditioning attenuates myocardial ischemia-reperfusion injury by inhibiting events in the early minutes of reperfusion. Cardiovasc Res. 2004;62:74–85. doi: 10.1016/j.cardiores.2004.01.006. [DOI] [PubMed] [Google Scholar]
- 214.Lai CF, Seshadri V, Huang K, Shao JS, Cai J, Vattikuti R, Schumacher A, Loewy AP, Denhardt DT, Rittling SR, Towler DA. An osteopontin-NADPH oxidase signaling cascade promotes pro-matrix metalloproteinase 9 activation in aortic mesenchymal cells. Circ Res. 2006;98:1479–1489. doi: 10.1161/01.RES.0000227550.00426.60. [DOI] [PubMed] [Google Scholar]
- 215.Lam CF, Liu YC, Tseng FL, Sung YH, Huang CC, Jiang MJ, Tsai YC. High-dose morphine impairs vascular endothelial function by increased production of superoxide anions. Anesthesiology. 2007;106:532–537. doi: 10.1097/00000542-200703000-00018. [DOI] [PubMed] [Google Scholar]
- 216.Landmesser U, Cai H, Dikalov S, McCann L, Hwang J, Jo H, Holland SM, Harrison DG. Role of p47(phox) in vascular oxidative stress and hypertension caused by angiotensin II. Hypertension. 2002;40:511–515. doi: 10.1161/01.hyp.0000032100.23772.98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Laufs U, Wassmann S, Czech T, Munzel T, Eisenhauer M, Bohm M, Nickenig G. Physical inactivity increases oxidative stress, endothelial dysfunction, and atherosclerosis. Arterioscler Thromb Vasc Biol. 2005;25:809–814. doi: 10.1161/01.ATV.0000158311.24443.af. [DOI] [PubMed] [Google Scholar]
- 218.Li H, Witte K, August M, Brausch I, Godtel-Armbrust U, Habermeier A, Closs EI, Oelze M, Munzel T, Forstermann U. Reversal of endothelial nitric oxide synthase uncoupling and up-regulation of endothelial nitric oxide synthase expression lowers blood pressure in hypertensive rats. J Am Coll Cardiol. 2006;47:2536–2544. doi: 10.1016/j.jacc.2006.01.071. [DOI] [PubMed] [Google Scholar]
- 219.Lodi F, Cogolludo A, Duarte J, Moreno L, Coviello A, Peral De BM, Vera R, Galisteo M, Jimenez R, Tamargo J, Perez-Vizcaino F. Increased NADPH oxidase activity mediates spontaneous aortic tone in genetically hypertensive rats. Eur J Pharmacol. 2006;544:97–103. doi: 10.1016/j.ejphar.2006.06.028. [DOI] [PubMed] [Google Scholar]
- 220.Loomis ED, Sullivan JC, Osmond DA, Pollock DM, Pollock JS. Endothelin mediates superoxide production and vasoconstriction through activation of NADPH oxidase and uncoupled nitric-oxide synthase in the rat aorta. J Pharmacol Exp Ther. 2005;315:1058–1064. doi: 10.1124/jpet.105.091728. [DOI] [PubMed] [Google Scholar]
- 221.Lund DD, Chu Y, Brooks RM, Faraci FM, Heistad DD. Effects of a common human gene variant of extracellular superoxide dismutase on endothelial function after endotoxin in mice. J Physiol. 2007;584:583–590. doi: 10.1113/jphysiol.2007.140830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Matsui R, Xu S, Maitland KA, Hayes A, Leopold JA, Handy DE, Loscalzo J, Cohen RA. Glucose-6 phosphate dehydrogenase deficiency decreases the vascular response to angiotensin II. Circulation. 2005;112:257–263. doi: 10.1161/CIRCULATIONAHA.104.499095. [DOI] [PubMed] [Google Scholar]
- 223.Miller AA, Drummond GR, Schmidt HH, Sobey CG. NADPH oxidase activity and function are profoundly greater in cerebral versus systemic arteries. Circ Res. 2005;97:1055–1062. doi: 10.1161/01.RES.0000189301.10217.87. [DOI] [PubMed] [Google Scholar]
- 224.Miller FJ, Jr, Gutterman DD, Rios CD, Heistad DD, Davidson BL. Superoxide production in vascular smooth muscle contributes to oxidative stress and impaired relaxation in atherosclerosis. Circ Res. 1998;82:1298–1305. doi: 10.1161/01.res.82.12.1298. [DOI] [PubMed] [Google Scholar]
- 225.Miller FJ, Jr, Sharp WJ, Fang X, Oberley LW, Oberley TD, Weintraub NL. Oxidative stress in human abdominal aortic aneurysms: a potential mediator of aneurysmal remodeling. Arterioscler Thromb Vasc Biol. 2002;22:560–565. doi: 10.1161/01.atv.0000013778.72404.30. [DOI] [PubMed] [Google Scholar]
- 226.Mitchell BM, Dorrance AM, Ergul A, Webb RC. Sepiapterin decreases vasorelaxation in nitric oxide synthase inhibition-induced hypertension. J Cardiovasc Pharmacol. 2004;43:93–98. doi: 10.1097/00005344-200401000-00014. [DOI] [PubMed] [Google Scholar]
- 227.Nurkiewicz TR, Boegehold MA. High salt intake reduces endothelium-dependent dilation of mouse arterioles via superoxide anion generated from nitric oxide synthase. Am J Physiol Regul Integr Comp Physiol. 2007;292:R1550–R1556. doi: 10.1152/ajpregu.00703.2006. [DOI] [PubMed] [Google Scholar]
- 228.Oelze M, Daiber A, Brandes RP, Hortmann M, Wenzel P, Hink U, Schulz E, Mollnau H, von SA, Kleschyov AL, Mulsch A, Li H, Forstermann U, Munzel T. Nebivolol inhibits superoxide formation by NADPH oxidase and endothelial dysfunction in angiotensin II-treated rats. Hypertension. 2006;48:677–684. doi: 10.1161/01.HYP.0000239207.82326.29. [DOI] [PubMed] [Google Scholar]
- 229.Oelze M, Warnholtz A, Faulhaber J, Wenzel P, Kleschyov AL, Coldewey M, Hink U, Pongs O, Fleming I, Wassmann S, Meinertz T, Ehmke H, Daiber A, Munzel T. NADPH oxidase accounts for enhanced superoxide production and impaired endothelium-dependent smooth muscle relaxation in BKbeta1-/- mice. Arterioscler Thromb Vasc Biol. 2006;26:1753–1759. doi: 10.1161/01.ATV.0000231511.26860.50. [DOI] [PubMed] [Google Scholar]
- 230.Okuda M, Inoue N, Azumi H, Seno T, Sumi Y, Hirata K, Kawashima S, Hayashi Y, Itoh H, Yodoi J, Yokoyama M. Expression of glutaredoxin in human coronary arteries: its potential role in antioxidant protection against atherosclerosis. Arterioscler Thromb Vasc Biol. 2001;21:1483–1487. doi: 10.1161/hq0901.095550. [DOI] [PubMed] [Google Scholar]
- 231.Orosz Z, Csiszar A, Labinskyy N, Smith K, Kaminski PM, Ferdinandy P, Wolin MS, Rivera A, Ungvari Z. Cigarette smoke-induced proinflammatory alterations in the endothelial phenotype: role of NAD(P)H oxidase activation. Am J Physiol Heart Circ Physiol. 2007;292:H130–H139. doi: 10.1152/ajpheart.00599.2006. [DOI] [PubMed] [Google Scholar]
- 232.Ou J, Ou Z, Jones DW, Holzhauer S, Hatoum OA, Ackerman AW, Weihrauch DW, Gutterman DD, Guice K, Oldham KT, Hillery CA, Pritchard KA., Jr L-4F, an apolipoprotein A-1 mimetic, dramatically improves vasodilation in hypercholesterolemia and sickle cell disease. Circulation. 2003;107:2337–2341. doi: 10.1161/01.CIR.0000070589.61860.A9. [DOI] [PubMed] [Google Scholar]
- 233.Oudot A, Martin C, Busseuil D, Vergely C, Demaison L, Rochette L. NADPH oxidases are in part responsible for increased cardiovascular superoxide production during aging. Free Radic Biol Med. 2006;40:2214–2222. doi: 10.1016/j.freeradbiomed.2006.02.020. [DOI] [PubMed] [Google Scholar]
- 234.Ozumi K, Tasaki H, Takatsu H, Nakata S, Morishita T, Koide S, Yamashita K, Tsutsui M, Okazaki M, Sasaguri Y, Adachi T, Nakashima Y. Extracellular superoxide dismutase overexpression reduces cuff-induced arterial neointimal formation. Atherosclerosis. 2005;181:55–62. doi: 10.1016/j.atherosclerosis.2005.01.051. [DOI] [PubMed] [Google Scholar]
- 235.Pannirselvam M, Wiehler WB, Anderson T, Triggle CR. Enhanced vascular reactivity of small mesenteric arteries from diabetic mice is associated with enhanced oxidative stress and cyclooxygenase products. Br J Pharmacol. 2005;144:953–960. doi: 10.1038/sj.bjp.0706121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Paravicini TM, Gulluyan LM, Dusting GJ, Drummond GR. Increased NADPH oxidase activity, gp91phox expression, and endothelium-dependent vasorelaxation during neointima formation in rabbits. Circ Res. 2002;91:54–61. doi: 10.1161/01.res.0000024106.81401.95. [DOI] [PubMed] [Google Scholar]
- 237.Park L, Anrather J, Forster C, Kazama K, Carlson GA, Iadecola C. Abeta-induced vascular oxidative stress and attenuation of functional hyperemia in mouse somatosensory cortex. J Cereb Blood Flow Metab. 2004;24:334–342. doi: 10.1097/01.WCB.0000105800.49957.1E. [DOI] [PubMed] [Google Scholar]
- 238.Qamirani E, Ren Y, Kuo L, Hein TW. C-reactive protein inhibits endothelium-dependent NO-mediated dilation in coronary arterioles by activating p38 kinase and NAD(P)H oxidase. Arterioscler Thromb Vasc Biol. 2005;25:995–1001. doi: 10.1161/01.ATV.0000159890.10526.1e. [DOI] [PubMed] [Google Scholar]
- 239.Rey FE, Cifuentes ME, Kiarash A, Quinn MT, Pagano PJ. Novel competitive inhibitor of NAD(P)H oxidase assembly attenuates vascular O(2)(-) and systolic blood pressure in mice. Circ Res. 2001;89:408–414. doi: 10.1161/hh1701.096037. [DOI] [PubMed] [Google Scholar]
- 240.Rice JB, Stoll LL, Li WG, Denning GM, Weydert J, Charipar E, Richenbacher WE, Miller FJ, Jr, Weintraub NL. Low-level endotoxin induces potent inflammatory activation of human blood vessels: inhibition by statins. Arterioscler Thromb Vasc Biol. 2003;23:1576–1582. doi: 10.1161/01.ATV.0000081741.38087.F9. [DOI] [PubMed] [Google Scholar]
- 241.Rice KM, Preston DL, Walker EM, Blough ER. Aging influences multiple incidices of oxidative stress in the aortic media of the Fischer 344/NNiaxBrown Norway/BiNia rat. Free Radic Res. 2006;40:185–197. doi: 10.1080/10715760500464957. [DOI] [PubMed] [Google Scholar]
- 242.Roghair RD, Miller FJ, Jr, Scholz TD, Lamb FS, Segar JL. Endothelial Superoxide Production Is Altered in Sheep Programmed by Early Gestation Dexamethasone Exposure. Neonatology. 2007;93:19–27. doi: 10.1159/000105521. [DOI] [PubMed] [Google Scholar]
- 243.Ryan MJ, Didion SP, Mathur S, Faraci FM, Sigmund CD. Angiotensin II-induced vascular dysfunction is mediated by the AT1A receptor in mice. Hypertension. 2004;43:1074–1079. doi: 10.1161/01.HYP.0000123074.89717.3d. [DOI] [PubMed] [Google Scholar]
- 244.Sanchez M, Lodi F, Vera R, Villar IC, Cogolludo A, Jimenez R, Moreno L, Romero M, Tamargo J, Perez-Vizcaino F, Duarte J. Quercetin and isorhamnetin prevent endothelial dysfunction, superoxide production, and overexpression of p47phox induced by angiotensin II in rat aorta. J Nutr. 2007;137:910–915. doi: 10.1093/jn/137.4.910. [DOI] [PubMed] [Google Scholar]
- 245.Sarr M, Chataigneau M, Martins S, Schott C, El BJ, Oak MH, Muller B, Chataigneau T, Schini-Kerth VB. Red wine polyphenols prevent angiotensin II-induced hypertension and endothelial dysfunction in rats: role of NADPH oxidase. Cardiovasc Res. 2006;71:794–802. doi: 10.1016/j.cardiores.2006.05.022. [DOI] [PubMed] [Google Scholar]
- 246.Sarr M, Chataigneau M, Etienne-Selloum N, Diallo AS, Schott C, Geffard M, Stoclet JC, Schini-Kerth VB, Muller B. Targeted and persistent effects of NO mediated by S-nitrosation of tissue thiols in arteries with endothelial dysfunction. Nitric Oxide. 2007;17:1–9. doi: 10.1016/j.niox.2007.04.003. [DOI] [PubMed] [Google Scholar]
- 247.Sartoretto JL, Oliveira MA, Nigro D, Carvalho MH, Tostes RC, Fortes ZB. Constrictor responses to noradrenaline, hemodynamic profile, and superoxide levels measured by hydroethidine oxidation in diabetic rats. Biol Pharm Bull. 2007;30:1938–1942. doi: 10.1248/bpb.30.1938. [DOI] [PubMed] [Google Scholar]
- 248.Sato K, Komaru T, Shioiri H, Takeda S, Takahashi K, Kanatsuka H, Nakayama M, Shirato K. Hypercholesterolemia impairs transduction of vasodilator signals derived from ischemic myocardium: myocardium-microvessel cross-talk. Arterioscler Thromb Vasc Biol. 2004;24:2034–2039. doi: 10.1161/01.ATV.0000143387.58166.c0. [DOI] [PubMed] [Google Scholar]
- 249.Sorescu D, Weiss D, Lassegue B, Clempus RE, Szocs K, Sorescu GP, Valppu L, Quinn MT, Lambeth JD, Vega JD, Taylor WR, Griendling KK. Superoxide production and expression of nox family proteins in human atherosclerosis. Circulation. 2002;105:1429–1435. doi: 10.1161/01.cir.0000012917.74432.66. [DOI] [PubMed] [Google Scholar]
- 250.Sutliff RL, Dikalov S, Weiss D, Parker J, Raidel S, Racine AK, Russ R, Haase CP, Taylor WR, Lewis W. Nucleoside reverse transcriptase inhibitors impair endothelium-dependent relaxation by increasing superoxide. Am J Physiol Heart Circ Physiol. 2002;283:H2363–H2370. doi: 10.1152/ajpheart.00151.2002. [DOI] [PubMed] [Google Scholar]
- 251.Sylvester FA, Stepp DW, Frisbee JC, Lombard JH. High-salt diet depresses acetylcholine reactivity proximal to NOS activation in cerebral arteries. Am J Physiol Heart Circ Physiol. 2002;283:H353–H363. doi: 10.1152/ajpheart.00127.2002. [DOI] [PubMed] [Google Scholar]
- 252.Symons JD, Rutledge JC, Simonsen U, Pattathu RA. Vascular dysfunction produced by hyperhomocysteinemia is more severe in the presence of low folate. Am J Physiol Heart Circ Physiol. 2006;290:H181–H191. doi: 10.1152/ajpheart.00765.2005. [DOI] [PubMed] [Google Scholar]
- 253.Szocs K, Lassegue B, Wenzel P, Wendt M, Daiber A, Oelze M, Meinertz T, Munzel T, Baldus S. Increased superoxide production in nitrate tolerance is associated with NAD(P)H oxidase and aldehyde dehydrogenase 2 downregulation. J Mol Cell Cardiol. 2007;42:1111–1118. doi: 10.1016/j.yjmcc.2007.03.904. [DOI] [PubMed] [Google Scholar]
- 254.Takahashi K, Komaru T, Takeda S, Takeda M, Koshida R, Nakayama M, Kokusho Y, Kawakami Y, Yamaguchi N, Miyazawa T, Shimokawa H, Shirato K. gamma-tocopherol, but not alpha-tocopherol, potently inhibits neointimal formation induced by vascular injury in insulin resistant rats. J Mol Cell Cardiol. 2006;41:544–554. doi: 10.1016/j.yjmcc.2006.06.010. [DOI] [PubMed] [Google Scholar]
- 255.Takaya T, Kawashima S, Shinohara M, Yamashita T, Toh R, Sasaki N, Inoue N, Hirata K, Yokoyama M. Angiotensin II type 1 receptor blocker telmisartan suppresses superoxide production and reduces atherosclerotic lesion formation in apolipoprotein E-deficient mice. Atherosclerosis. 2006;186:402–410. doi: 10.1016/j.atherosclerosis.2005.08.009. [DOI] [PubMed] [Google Scholar]
- 256.Troncoso Brindeiro CM, da Silva AQ, Allahdadi KJ, Youngblood V, Kanagy NL. Reactive oxygen species contribute to sleep apnea-induced hypertension in rats. Am J Physiol Heart Circ Physiol. 2007;293:H2971–H2976. doi: 10.1152/ajpheart.00219.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Ungvari Z, Csiszar A, Edwards JG, Kaminski PM, Wolin MS, Kaley G, Koller A. Increased superoxide production in coronary arteries in hyperhomocysteinemia: role of tumor necrosis factor-alpha, NAD(P)H oxidase, and inducible nitric oxide synthase. Arterioscler Thromb Vasc Biol. 2003;23:418–424. doi: 10.1161/01.ATV.0000061735.85377.40. [DOI] [PubMed] [Google Scholar]
- 258.Villamor E, Kessels CG, Fischer MA, Bast A, de Mey JG, Blanco CE. Role of superoxide anion on basal and stimulated nitric oxide activity in neonatal piglet pulmonary vessels. Pediatr Res. 2003;54:372–381. doi: 10.1203/01.PDR.0000077481.15081.C8. [DOI] [PubMed] [Google Scholar]
- 259.Vinh A, Widdop RE, Drummond GR, Gaspari TA. Chronic angiotensin IV treatment reverses endothelial dysfunction in ApoE-deficient mice. Cardiovasc Res. 2007 doi: 10.1093/cvr/cvm021. [DOI] [PubMed] [Google Scholar]
- 260.Wang D, Jose P, Wilcox CS. beta(1) Receptors protect the renal afferent arteriole of angiotensin-infused rabbits from norepinephrine-induced oxidative stress. J Am Soc Nephrol. 2006;17:3347–3354. doi: 10.1681/ASN.2006030212. [DOI] [PubMed] [Google Scholar]
- 261.Xi H, Akishita M, Nagai K, Yu W, Hasegawa H, Eto M, Kozaki K, Toba K. Potent free radical scavenger, edaravone, suppresses oxidative stress-induced endothelial damage and early atherosclerosis. Atherosclerosis. 2007;191:281–289. doi: 10.1016/j.atherosclerosis.2006.05.040. [DOI] [PubMed] [Google Scholar]
- 262.Xu H, Fink GD, Galligan JJ. Nitric oxide-independent effects of tempol on sympathetic nerve activity and blood pressure in DOCA-salt rats. Am J Physiol Heart Circ Physiol. 2002;283:H885–H892. doi: 10.1152/ajpheart.00134.2002. [DOI] [PubMed] [Google Scholar]
- 263.Yun MR, Kim JJ, Im DS, Yang SD, Kim CD. Involvement of NAD(P)H oxidase in the enhanced expression of cell adhesion molecules in the aorta of diabetic mice. Life Sci. 2004;75:2463–2472. doi: 10.1016/j.lfs.2004.04.041. [DOI] [PubMed] [Google Scholar]
- 264.Yzydorczyk C, Gobeil F, Jr, Cambonie G, Lahaie I, Le NL, Samarani S, Ahmad A, Lavoie JC, Oligny LL, Pladys P, Hardy P, Nuyt AM. Exaggerated vasomotor response to ANG II in rats with fetal programming of hypertension associated with exposure to a low-protein diet during gestation. Am J Physiol Regul Integr Comp Physiol. 2006;291:R1060–R1068. doi: 10.1152/ajpregu.00798.2005. [DOI] [PubMed] [Google Scholar]
- 265.Zhang C, Hein TW, Wang W, Kuo L. Divergent roles of angiotensin II AT1 and AT2 receptors in modulating coronary microvascular function. Circ Res. 2003;92:322–329. doi: 10.1161/01.res.0000056759.53828.2c. [DOI] [PubMed] [Google Scholar]
- 266.Zhang DX, Zou AP, Li PL. Ceramide reduces endothelium-dependent vasodilation by increasing superoxide production in small bovine coronary arteries. Circ Res. 2001;88:824–831. doi: 10.1161/hh0801.089604. [DOI] [PubMed] [Google Scholar]
- 267.Zhang DX, Zou AP, Li PL. Ceramide-induced activation of NADPH oxidase and endothelial dysfunction in small coronary arteries. Am J Physiol Heart Circ Physiol. 2003;284:H605–H612. doi: 10.1152/ajpheart.00697.2002. [DOI] [PubMed] [Google Scholar]
- 268.Zhou J, Werstuck GH, Lhotak S, de Koning AB, Sood SK, Hossain GS, Moller J, Ritskes-Hoitinga M, Falk E, Dayal S, Lentz SR, Austin RC. Association of multiple cellular stress pathways with accelerated atherosclerosis in hyperhomocysteinemic apolipoprotein E-deficient mice. Circulation. 2004;110:207–213. doi: 10.1161/01.CIR.0000134487.51510.97. [DOI] [PubMed] [Google Scholar]
- 269.Zhu J, Mori T, Huang T, Lombard JH. Effect of high-salt diet on NO release and superoxide production in rat aorta. Am J Physiol Heart Circ Physiol. 2004;286:H575–H583. doi: 10.1152/ajpheart.00331.2003. [DOI] [PubMed] [Google Scholar]
- 270.Zhu J, Huang T, Lombard JH. Effect of high-salt diet on vascular relaxation and oxidative stress in mesenteric resistance arteries. J Vasc Res. 2007;44:382–390. doi: 10.1159/000102955. [DOI] [PubMed] [Google Scholar]
- 271.Supinski G, Nethery D, Stofan D, DiMarco A. Extracellular calcium modulates generation of reactive oxygen species by the contracting diaphragm. J Appl Physiol. 1999;87:2177–2185. doi: 10.1152/jappl.1999.87.6.2177. [DOI] [PubMed] [Google Scholar]
- 272.Zuo L, Christofi FL, Wright VP, Liu CY, Merola AJ, Berliner LJ, Clanton TL. Intra- and extracellular measurement of reactive oxygen species produced during heat stress in diaphragm muscle. Am J Physiol Cell Physiol. 2000;279:C1058–C1066. doi: 10.1152/ajpcell.2000.279.4.C1058. [DOI] [PubMed] [Google Scholar]
- 273.Zuo L, Christofi FL, Wright VP, Bao S, Clanton TL. Lipoxygenase-dependent superoxide release in skeletal muscle. J Appl Physiol. 2004;97:661–668. doi: 10.1152/japplphysiol.00096.2004. [DOI] [PubMed] [Google Scholar]
- 274.Brasen JH, Nieminen-Kelha M, Markmann D, Malle E, Schneider W, Neumayer HH, Budde K, Luft FC, Dragun D. Lectin-like oxidized low-density lipoprotein (LDL) receptor (LOX-1)-mediated pathway and vascular oxidative injury in older-age rat renal transplants. Kidney Int. 2005;67:1583–1594. doi: 10.1111/j.1523-1755.2005.00240.x. [DOI] [PubMed] [Google Scholar]
- 275.Hong NJ, Garvin JL. Flow increases superoxide production by NADPH oxidase via activation of Na-K-2Cl cotransport and mechanical stress in thick ascending limbs. Am J Physiol Renal Physiol. 2007;292:F993–F998. doi: 10.1152/ajprenal.00383.2006. [DOI] [PubMed] [Google Scholar]
- 276.Li N, Yi FX, Spurrier JL, Bobrowitz CA, Zou AP. Production of superoxide through NADH oxidase in thick ascending limb of Henle's loop in rat kidney. Am J Physiol Renal Physiol. 2002;282:F1111–F1119. doi: 10.1152/ajprenal.00218.2001. [DOI] [PubMed] [Google Scholar]
- 277.Liu R, Garvin JL, Ren Y, Pagano PJ, Carretero OA. Depolarization of the macula densa induces superoxide production via NAD(P)H oxidase. Am J Physiol Renal Physiol. 2007;292:F1867–F1872. doi: 10.1152/ajprenal.00515.2006. [DOI] [PubMed] [Google Scholar]
- 278.Robles HV, Romo E, Sanchez-Mendoza A, Rios A, Soto V, vila-Casado MC, Medina A, Escalante B. Lead exposure effect on angiotensin II renal vasoconstriction. Hum Exp Toxicol. 2007;26:499–507. doi: 10.1177/0960327106077597. [DOI] [PubMed] [Google Scholar]
- 279.Satoh M, Fujimoto S, Haruna Y, Arakawa S, Horike H, Komai N, Sasaki T, Tsujioka K, Makino H, Kashihara N. NAD(P)H oxidase and uncoupled nitric oxide synthase are major sources of glomerular superoxide in rats with experimental diabetic nephropathy. Am J Physiol Renal Physiol. 2005;288:F1144–F1152. doi: 10.1152/ajprenal.00221.2004. [DOI] [PubMed] [Google Scholar]
- 280.Sharma K, Cook A, Smith M, Valancius C, Inscho EW. TGF-beta impairs renal autoregulation via generation of ROS. Am J Physiol Renal Physiol. 2005;288:F1069–F1077. doi: 10.1152/ajprenal.00345.2004. [DOI] [PubMed] [Google Scholar]
- 281.Bao N, Minatoguchi S, Kobayashi H, Yasuda S, Kawamura I, Iwasa M, Yamaki T, Sumi S, Misao Y, Arai M, Nishigaki K, Takemura G, Fujiwara T, Fujiwara H. Pravastatin reduces myocardial infarct size via increasing protein kinase C-dependent nitric oxide, decreasing oxyradicals and opening the mitochondrial adenosine triphosphate-sensitive potassium channels in rabbits. Circ J. 2007;71:1622–1628. doi: 10.1253/circj.71.1622. [DOI] [PubMed] [Google Scholar]
- 282.Camara AK, Aldakkak M, Heisner JS, Rhodes SS, Riess ML, An J, Heinen A, Stowe DF. ROS scavenging before 27 degrees C ischemia protects hearts and reduces mitochondrial ROS, Ca2+ overload, and changes in redox state. Am J Physiol Cell Physiol. 2007;292:C2021–C2031. doi: 10.1152/ajpcell.00231.2006. [DOI] [PubMed] [Google Scholar]
- 283.Hunter AL, Bai N, Laher I, Granville DJ. Cytochrome p450 2C inhibition reduces post-ischemic vascular dysfunction. Vascul Pharmacol. 2005;43:213–219. doi: 10.1016/j.vph.2005.07.005. [DOI] [PubMed] [Google Scholar]
- 284.Kai H, Mori T, Tokuda K, Takayama N, Tahara N, Takemiya K, Kudo H, Sugi Y, Fukui D, Yasukawa H, Kuwahara F, Imaizumi T. Pressure overload-induced transient oxidative stress mediates perivascular inflammation and cardiac fibrosis through angiotensin II. Hypertens Res. 2006;29:711–718. doi: 10.1291/hypres.29.711. [DOI] [PubMed] [Google Scholar]
- 285.Lebrecht D, Geist A, Ketelsen UP, Haberstroh J, Setzer B, Walker UA. Dexrazoxane prevents doxorubicin-induced long-term cardiotoxicity and protects myocardial mitochondria from genetic and functional lesions in rats. Br J Pharmacol. 2007;151:771–778. doi: 10.1038/sj.bjp.0707294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 286.Manintveld OC, Te Lintel HM, van den Bos EJ, Suurenbroek GM, Dekkers DH, Verdouw PD, Lamers JM, Duncker DJ. Cardiac effects of postconditioning depend critically on the duration of index ischemia. Am J Physiol Heart Circ Physiol. 2007;292:H1551–H1560. doi: 10.1152/ajpheart.00151.2006. [DOI] [PubMed] [Google Scholar]
- 287.Nilakantan V, Zhou X, Hilton G, Shi Y, Baker JE, Khanna AK, Pieper GM. Antagonizing reactive oxygen by treatment with a manganese (III) metalloporphyrin-based superoxide dismutase mimetic in cardiac transplants. J Thorac Cardiovasc Surg. 2006;131:898–906. doi: 10.1016/j.jtcvs.2005.11.015. [DOI] [PubMed] [Google Scholar]
- 288.Onogi H, Minatoguchi S, Chen XH, Bao N, Kobayashi H, Misao Y, Yasuda S, Yamaki T, Maruyama R, Uno Y, Arai M, Takemura G, Fujiwara H. Edaravone reduces myocardial infarct size and improves cardiac function and remodelling in rabbits. Clin Exp Pharmacol Physiol. 2006;33:1035–1041. doi: 10.1111/j.1440-1681.2006.04483.x. [DOI] [PubMed] [Google Scholar]
- 289.Sayen MR, Gustafsson AB, Sussman MA, Molkentin JD, Gottlieb RA. Calcineurin transgenic mice have mitochondrial dysfunction and elevated superoxide production. Am J Physiol Cell Physiol. 2003;284:C562–C570. doi: 10.1152/ajpcell.00336.2002. [DOI] [PubMed] [Google Scholar]
- 290.Shi Y, Hutchins WC, Su J, Siker D, Hogg N, Pritchard KA, Jr, Keszler A, Tweddell JS, Baker JE. Delayed cardioprotection with isoflurane: role of reactive oxygen and nitrogen. Am J Physiol Heart Circ Physiol. 2005;288:H175–H184. doi: 10.1152/ajpheart.00494.2004. [DOI] [PubMed] [Google Scholar]
- 291.Williams IA, Allen DG. The role of reactive oxygen species in the hearts of dystrophin-deficient mdx mice. Am J Physiol Heart Circ Physiol. 2007;293:H1969–H1977. doi: 10.1152/ajpheart.00489.2007. [DOI] [PubMed] [Google Scholar]
- 292.Bindokas VP, Kuznetsov A, Sreenan S, Polonsky KS, Roe MW, Philipson LH. Visualizing superoxide production in normal and diabetic rat islets of Langerhans. J Biol Chem. 2003;278:9796–9801. doi: 10.1074/jbc.M206913200. [DOI] [PubMed] [Google Scholar]
- 293.Bivalacqua TJ, Armstrong JS, Biggerstaff J, bdel-Mageed AB, Kadowitz PJ, Hellstrom WJ, Champion HC. Gene transfer of extracellular SOD to the penis reduces O2-* and improves erectile function in aged rats. Am J Physiol Heart Circ Physiol. 2003;284:H1408–H1421. doi: 10.1152/ajpheart.00770.2002. [DOI] [PubMed] [Google Scholar]
- 294.Minamiyama Y, Takemura S, Toyokuni S, Imaoka S, Funae Y, Hirohashi K, Yoshikawa T, Okada S. CYP3A induction aggravates endotoxemic liver injury via reactive oxygen species in male rats. Free Radic Biol Med. 2004;37:703–712. doi: 10.1016/j.freeradbiomed.2004.05.022. [DOI] [PubMed] [Google Scholar]
- 295.Zhou Z, Wang L, Song Z, Saari JT, McClain CJ, Kang YJ. Zinc supplementation prevents alcoholic liver injury in mice through attenuation of oxidative stress. Am J Pathol. 2005;166:1681–1690. doi: 10.1016/S0002-9440(10)62478-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 296.Grobe AC, Wells SM, Benavidez E, Oishi P, Azakie A, Fineman JR, Black SM. Increased oxidative stress in lambs with increased pulmonary blood flow and pulmonary hypertension: role of NADPH oxidase and endothelial NO synthase. Am J Physiol Lung Cell Mol Physiol. 2006;290:L1069–L1077. doi: 10.1152/ajplung.00408.2005. [DOI] [PubMed] [Google Scholar]
- 297.Zhang Q, Matsuzaki I, Chatterjee S, Fisher AB. Activation of endothelial NADPH oxidase during normoxic lung ischemia is KATP channel dependent. Am J Physiol Lung Cell Mol Physiol. 2005;289:L954–L961. doi: 10.1152/ajplung.00210.2005. [DOI] [PubMed] [Google Scholar]
- 298.Al-Shabrawey M, Bartoli M, El-Remessy AB, Platt DH, Matragoon S, Behzadian MA, Caldwell RW, Caldwell RB. Inhibition of NAD(P)H oxidase activity blocks vascular endothelial growth factor overexpression and neovascularization during ischemic retinopathy. Am J Pathol. 2005;167:599–607. doi: 10.1016/S0002-9440(10)63001-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 299.Chen P, Guo AM, Edwards PA, Trick G, Scicli AG. Role of NADPH oxidase and ANG II in diabetes-induced retinal leukostasis. Am J Physiol Regul Integr Comp Physiol. 2007;293:R1619–R1629. doi: 10.1152/ajpregu.00290.2007. [DOI] [PubMed] [Google Scholar]
- 300.Sicard P, Acar N, Gregoire S, Lauzier B, Bron AM, Creuzot-Garcher C, Bretillon L, Vergely C, Rochette L. Influence of rosuvastatin on the NAD(P)H oxidase activity in the retina and electroretinographic response of spontaneously hypertensive rats. Br J Pharmacol. 2007;151:979–986. doi: 10.1038/sj.bjp.0707322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 301.Yadav UC, Srivastava SK, Ramana KV. Aldose reductase inhibition prevents endotoxin-induced uveitis in rats. Invest Ophthalmol Vis Sci. 2007;48:4634–4642. doi: 10.1167/iovs.07-0485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Lim SD, Sun C, Lambeth JD, Marshall F, Amin M, Chung L, Petros JA, Arnold RS. Increased Nox1 and hydrogen peroxide in prostate cancer. Prostate. 2005;62:200–207. doi: 10.1002/pros.20137. [DOI] [PubMed] [Google Scholar]
- 303.Cao X, Dai X, Parker LM, Kreulen DL. Differential regulation of NADPH oxidase in sympathetic and sensory Ganglia in deoxycorticosterone acetate salt hypertension. Hypertension. 2007;50:663–671. doi: 10.1161/HYPERTENSIONAHA.107.089748. [DOI] [PubMed] [Google Scholar]
- 304.Haruna Y, Morita Y, Komai N, Yada T, Sakuta T, Tomita N, Fox DA, Kashihara N. Endothelial dysfunction in rat adjuvant-induced arthritis: vascular superoxide production by NAD(P)H oxidase and uncoupled endothelial nitric oxide synthase. Arthritis Rheum. 2006;54:1847–1855. doi: 10.1002/art.21891. [DOI] [PubMed] [Google Scholar]
- 305.Taylor NE, Glocka P, Liang M, Cowley AW., Jr NADPH oxidase in the renal medulla causes oxidative stress and contributes to salt-sensitive hypertension in Dahl S rats. Hypertension. 2006;47:692–698. doi: 10.1161/01.HYP.0000203161.02046.8d. [DOI] [PubMed] [Google Scholar]
- 306.Yan S, Wang X, Chai H, Wang H, Yao Q, Chen C. Secretoneurin increases monolayer permeability in human coronary artery endothelial cells. Surgery. 2006;140:243–251. doi: 10.1016/j.surg.2006.04.001. [DOI] [PubMed] [Google Scholar]
- 307.Bjorkqvist M, Jurstrand M, Bodin L, Fredlund H, Schollin J. Defective neutrophil oxidative burst in preterm newborns on exposure to coagulase-negative staphylococci. Pediatr Res. 2004;55:966–971. doi: 10.1203/01.pdr.0000127018.44938.89. [DOI] [PubMed] [Google Scholar]
- 308.Conrad A, Hansmann C, Engels I, Daschner FD, Frank U. Extract of Pelargonium sidoides (EPs 7630) improves phagocytosis, oxidative burst, and intracellular killing of human peripheral blood phagocytes in vitro. Phytomedicine. 2007;14(6):46–51. doi: 10.1016/j.phymed.2006.11.016. [DOI] [PubMed] [Google Scholar]
- 309.Holthe MR, Staff AC, Berge LN, Lyberg T. Leukocyte adhesion molecules and reactive oxygen species in preeclampsia. Obstet Gynecol. 2004;103:913–922. doi: 10.1097/01.AOG.0000124806.39111.ba. [DOI] [PubMed] [Google Scholar]
- 310.Nielsen HG, Hagberg IA, Lyberg T. Marathon running leads to partial exhaustion of ROS-generating capacity in leukocytes. Med Sci Sports Exerc. 2004;36:68–73. doi: 10.1249/01.MSS.0000106168.12113.95. [DOI] [PubMed] [Google Scholar]
- 311.Nielsen HG, Opstad PK, Lyberg T. LeuCAM and reactive oxygen species during long-term exercise combined with sleep and energy deficiency. Med Sci Sports Exerc. 2007;39:275–282. doi: 10.1249/01.mss.0000241648.07382.7c. [DOI] [PubMed] [Google Scholar]
- 312.Andrews ZB, Horvath B, Barnstable CJ, Elsworth J, Yang L, Beal MF, Roth RH, Matthews RT, Horvath TL. Uncoupling protein-2 is critical for nigral dopamine cell survival in a mouse model of Parkinson's disease. J Neurosci. 2005;25:184–191. doi: 10.1523/JNEUROSCI.4269-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Bao F, Dekaban GA, Weaver LC. Anti-CD11d antibody treatment reduces free radical formation and cell death in the injured spinal cord of rats. J Neurochem. 2005;94:1361–1373. doi: 10.1111/j.1471-4159.2005.03280.x. [DOI] [PubMed] [Google Scholar]
- 314.Choi SH, Lee DY, Kim SU, Jin BK. Thrombin-induced oxidative stress contributes to the death of hippocampal neurons in vivo: role of microglial NADPH oxidase. J Neurosci. 2005;25:4082–4090. doi: 10.1523/JNEUROSCI.4306-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315.Cingolani C, Rogers B, Lu L, Kachi S, Shen J, Campochiaro PA. Retinal degeneration from oxidative damage. Free Radic Biol Med. 2006;40:660–669. doi: 10.1016/j.freeradbiomed.2005.09.032. [DOI] [PubMed] [Google Scholar]
- 316.Kim GW, Lewen A, Copin J, Watson BD, Chan PH. The cytosolic antioxidant, copper/zinc superoxide dismutase, attenuates blood-brain barrier disruption and oxidative cellular injury after photothrombotic cortical ischemia in mice. Neuroscience. 2001;105:1007–1018. doi: 10.1016/s0306-4522(01)00237-8. [DOI] [PubMed] [Google Scholar]
- 317.Kim GW, Kondo T, Noshita N, Chan PH. Manganese superoxide dismutase deficiency exacerbates cerebral infarction after focal cerebral ischemia/reperfusion in mice: implications for the production and role of superoxide radicals. Stroke. 2002;33:809–815. doi: 10.1161/hs0302.103745. [DOI] [PubMed] [Google Scholar]
- 318.Manabe Y, Anrather J, Kawano T, Niwa K, Zhou P, Ross ME, Iadecola C. Prostanoids, not reactive oxygen species, mediate COX-2-dependent neurotoxicity. Ann Neurol. 2004;55:668–675. doi: 10.1002/ana.20078. [DOI] [PubMed] [Google Scholar]
- 319.Rogers BS, Symons RC, Komeima K, Shen J, Xiao W, Swaim ME, Gong YY, Kachi S, Campochiaro PA. Differential sensitivity of cones to iron-mediated oxidative damage. Invest Ophthalmol Vis Sci. 2007;48:438–445. doi: 10.1167/iovs.06-0528. [DOI] [PubMed] [Google Scholar]
- 320.Wu DC, Teismann P, Tieu K, Vila M, Jackson-Lewis V, Ischiropoulos H, Przedborski S. NADPH oxidase mediates oxidative stress in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine model of Parkinson's disease. Proc Natl Acad Sci U S A. 2003;100:6145–6150. doi: 10.1073/pnas.0937239100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 321.Endo H, Nito C, Kamada H, Yu F, Chan PH. Reduction in oxidative stress by superoxide dismutase overexpression attenuates acute brain injury after subarachnoid hemorrhage via activation of Akt/glycogen synthase kinase-3beta survival signaling. J Cereb Blood Flow Metab. 2007;27:975–982. doi: 10.1038/sj.jcbfm.9600399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 322.Ruocco A, Santillo M, Cicale M, Seru R, Cuda G, Anrather J, Iadecola C, Postiglione A, Avvedimento EV, Paterno R. Farnesyl transferase inhibitors induce neuroprotection by inhibiting Ha-Ras signalling pathway. Eur J Neurosci. 2007;26:3261–3266. doi: 10.1111/j.1460-9568.2007.05935.x. [DOI] [PubMed] [Google Scholar]
- 323.Saito A, Hayashi T, Okuno S, Ferrand-Drake M, Chan PH. Overexpression of copper/zinc superoxide dismutase in transgenic mice protects against neuronal cell death after transient focal ischemia by blocking activation of the Bad cell death signaling pathway. J Neurosci. 2003;23:1710–1718. doi: 10.1523/JNEUROSCI.23-05-01710.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Murakami K, Kondo T, Kawase M, Li Y, Sato S, Chen SF, Chan PH. Mitochondrial susceptibility to oxidative stress exacerbates cerebral infarction that follows permanent focal cerebral ischemia in mutant mice with manganese superoxide dismutase deficiency. J Neurosci. 1998;18:205–213. doi: 10.1523/JNEUROSCI.18-01-00205.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 325.Kawase M, Murakami K, Fujimura M, Morita-Fujimura Y, Gasche Y, Kondo T, Scott RW, Chan PH. Exacerbation of delayed cell injury after transient global ischemia in mutant mice with CuZn superoxide dismutase deficiency. Stroke. 1999;30:1962–1968. doi: 10.1161/01.str.30.9.1962. [DOI] [PubMed] [Google Scholar]
- 326.Tanaka K, Weihrauch D, Kehl F, Ludwig LM, LaDisa JF, Jr, Kersten JR, Pagel PS, Warltier DC. Mechanism of preconditioning by isoflurane in rabbits: a direct role for reactive oxygen species. Anesthesiology. 2002;97:1485–1490. doi: 10.1097/00000542-200212000-00021. [DOI] [PubMed] [Google Scholar]
- 327.Schneider R, Raff U, Vornberger N, Schmidt M, Freund R, Reber M, Schramm L, Gambaryan S, Wanner C, Schmidt HH, Galle J. L-Arginine counteracts nitric oxide deficiency and improves the recovery phase of ischemic acute renal failure in rats. Kidney Int. 2003;64:216–225. doi: 10.1046/j.1523-1755.2003.00063.x. [DOI] [PubMed] [Google Scholar]
- 328.Kunz A, Anrather J, Zhou P, Orio M, Iadecola C. Cyclooxygenase-2 does not contribute to postischemic production of reactive oxygen species. J Cereb Blood Flow Metab. 2007;27:545–551. doi: 10.1038/sj.jcbfm.9600369. [DOI] [PubMed] [Google Scholar]
- 329.Zhao H, Sapolsky RM, Steinberg GK. Interrupting reperfusion as a stroke therapy: ischemic postconditioning reduces infarct size after focal ischemia in rats. J Cereb Blood Flow Metab. 2006;26:1114–1121. doi: 10.1038/sj.jcbfm.9600348. [DOI] [PubMed] [Google Scholar]
- 330.Dantas AP, Franco MC, Silva-Antonialli MM, Tostes RC, Fortes ZB, Nigro D, Carvalho MH. Gender differences in superoxide generation in microvessels of hypertensive rats: role of NAD(P)H-oxidase. Cardiovasc Res. 2004;61:22–29. doi: 10.1016/j.cardiores.2003.10.010. [DOI] [PubMed] [Google Scholar]
- 331.Erdos B, Broxson CS, King MA, Scarpace PJ, Tumer N. Acute pressor effect of central angiotensin II is mediated by NAD(P)H-oxidase-dependent production of superoxide in the hypothalamic cardiovascular regulatory nuclei. J Hypertens. 2006;24:109–116. doi: 10.1097/01.hjh.0000198026.99600.59. [DOI] [PubMed] [Google Scholar]
- 332.Dantas AP, Tostes RC, Fortes ZB, Costa SG, Nigro D, Carvalho MH. In vivo evidence for antioxidant potential of estrogen in microvessels of female spontaneously hypertensive rats. Hypertension. 2002;39:405–411. doi: 10.1161/hy0202.102993. [DOI] [PubMed] [Google Scholar]
- 333.Camara AK, Riess ML, Kevin LG, Novalija E, Stowe DF. Hypothermia augments reactive oxygen species detected in the guinea pig isolated perfused heart. Am J Physiol Heart Circ Physiol. 2004;286:H1289–H1299. doi: 10.1152/ajpheart.00811.2003. [DOI] [PubMed] [Google Scholar]
- 334.Lund AK, Peterson SL, Timmins GS, Walker MK. Endothelin-1-mediated increase in reactive oxygen species and NADPH Oxidase activity in hearts of aryl hydrocarbon receptor (AhR) null mice. Toxicol Sci. 2005;88:265–273. doi: 10.1093/toxsci/kfi284. [DOI] [PubMed] [Google Scholar]
- 335.Makino A, Skelton MM, Zou AP, Roman RJ, Cowley AW., Jr Increased renal medullary oxidative stress produces hypertension. Hypertension. 2002;39:667–672. doi: 10.1161/hy0202.103469. [DOI] [PubMed] [Google Scholar]
- 336.Makino A, Skelton MM, Zou AP, Cowley AW., Jr Increased renal medullary H2O2 leads to hypertension. Hypertension. 2003;42:25–30. doi: 10.1161/01.HYP.0000074903.96928.91. [DOI] [PubMed] [Google Scholar]
- 337.Taylor NE, Maier KG, Roman RJ, Cowley AW., Jr NO synthase uncoupling in the kidney of Dahl S rats: role of dihydrobiopterin. Hypertension. 2006;48:1066–1071. doi: 10.1161/01.HYP.0000248751.11383.7c. [DOI] [PubMed] [Google Scholar]
- 338.McGuigan RM, Mullenix P, Norlund LL, Ward D, Walts M, Azarow K. Acute lung injury using oleic acid in the laboratory rat: establishment of a working model and evidence against free radicals in the acute phase. Curr Surg. 2003;60:412–417. doi: 10.1016/S0149-7944(02)00775-4. [DOI] [PubMed] [Google Scholar]
- 339.Tanaka K, Weihrauch D, Ludwig LM, Kersten JR, Pagel PS, Warltier DC. Mitochondrial adenosine triphosphate-regulated potassium channel opening acts as a trigger for isoflurane-induced preconditioning by generating reactive oxygen species. Anesthesiology. 2003;98:935–943. doi: 10.1097/00000542-200304000-00021. [DOI] [PubMed] [Google Scholar]
- 340.Lum H, Roebuck KA. Oxidant stress and endothelial cell dysfunction. Am J Physiol Cell Physiol. 2001;280:C719–C741. doi: 10.1152/ajpcell.2001.280.4.C719. [DOI] [PubMed] [Google Scholar]
- 341.McAmis WC, Schaeffer RC, Jr, Baynes JW, Wolf MB. Menadione causes endothelial barrier failure by a direct effect on intracellular thiols, independent of reactive oxidant production. Biochim Biophys Acta. 2003;1641:43–53. doi: 10.1016/s0167-4889(03)00063-6. [DOI] [PubMed] [Google Scholar]
- 342.McQuaid KE, Keenan AK. Endothelial barrier dysfunction and oxidative stress: roles for nitric oxide? Exp Physiol. 1997;82:369–376. doi: 10.1113/expphysiol.1997.sp004032. [DOI] [PubMed] [Google Scholar]
- 343.Brillault J, Lam TI, Rutkowsky JM, Foroutan S, O'Donnell ME. Hypoxia effects on cell volume and ion uptake of cerebral microvascular endothelial cells. Am J Physiol Cell Physiol. 2008;294:C88–C96. doi: 10.1152/ajpcell.00148.2007. [DOI] [PubMed] [Google Scholar]
- 344.Abe M, O'Connor P, Kaldunski M, Liang M, Roman RJ, Cowley AW., Jr Effect of sodium delivery on superoxide and nitric oxide in the medullary thick ascending limb. Am J Physiol Renal Physiol. 2006;291:F350–F357. doi: 10.1152/ajprenal.00407.2005. [DOI] [PubMed] [Google Scholar]
- 345.Chenais B, Andriollo M, Guiraud P, Belhoussine R, Jeannesson P. Oxidative stress involvement in chemically induced differentiation of K562 cells. Free Radic Biol Med. 2000;28:18–27. doi: 10.1016/s0891-5849(99)00195-1. [DOI] [PubMed] [Google Scholar]
- 346.Richard D, Morjani H, Chenais B. Free radical production and labile iron pool decrease triggered by subtoxic concentration of aclarubicin in human leukemia cell lines. Leuk Res. 2002;26:927–931. doi: 10.1016/s0145-2126(02)00030-9. [DOI] [PubMed] [Google Scholar]
- 347.Sanvicens N, Gomez-Vicente V, Messeguer A, Cotter TG. The radical scavenger CR-6 protects SH-SY5Y neuroblastoma cells from oxidative stress-induced apoptosis: effect on survival pathways. J Neurochem. 2006;98:735–747. doi: 10.1111/j.1471-4159.2006.03914.x. [DOI] [PubMed] [Google Scholar]
- 348.Zhang G, Zhang F, Muh R, Yi F, Chalupsky K, Cai H, Li PL. Autocrine/paracrine pattern of superoxide production through NAD(P)H oxidase in coronary arterial myocytes. Am J Physiol Heart Circ Physiol. 2007;292:H483–H495. doi: 10.1152/ajpheart.00632.2006. [DOI] [PubMed] [Google Scholar]
- 349.Dussmann H, Kogel D, Rehm M, Prehn JH. Mitochondrial membrane permeabilization and superoxide production during apoptosis. A single-cell analysis. J Biol Chem. 2003;278:12645–12649. doi: 10.1074/jbc.M210826200. [DOI] [PubMed] [Google Scholar]
- 350.Roding J, Naujok A, Zimmermann HW. Effects of ethidium bromide, tetramethylethidium bromide and betaine B on the ultrastructure of HeLa cell mitochondria in situ. A comparative binding study. Histochemistry. 1986;85:215–222. doi: 10.1007/BF00494807. [DOI] [PubMed] [Google Scholar]
- 351.Rottenberg H. Membrane potential and surface potential in mitochondria: uptake and binding of lipophilic cations. J Membr Biol. 1984;81:127–138. doi: 10.1007/BF01868977. [DOI] [PubMed] [Google Scholar]
- 352.Hashiguchi K, Zhang-Akiyama QM. Establishment of human cell lines lacking mitochondrial DNA. Methods Mol Biol. 2009;554:383–391. doi: 10.1007/978-1-59745-521-3_23. [DOI] [PubMed] [Google Scholar]
- 353.Budd SL, Castilho RF, Nicholls DG. Mitochondrial membrane potential and hydroethidine-monitored superoxide generation in cultured cerebellar granule cells. FEBS Lett. 1997;415:21–24. doi: 10.1016/s0014-5793(97)01088-0. [DOI] [PubMed] [Google Scholar]
- 354.Verkaart S, Koopman WJ, van Emst-de Vries SE, Nijtmans LG, van den Heuvel LW, Smeitink JA, Willems PH. Superoxide production is inversely related to complex I activity in inherited complex I deficiency. Biochim Biophys Acta. 2007;1772:373–381. doi: 10.1016/j.bbadis.2006.12.009. [DOI] [PubMed] [Google Scholar]
- 355.Koopman WJ, Verkaart S, Visch HJ, van der Westhuizen FH, Murphy MP, van den Heuvel LW, Smeitink JA, Willems PH. Inhibition of complex I of the electron transport chain causes O2-. -mediated mitochondrial outgrowth. Am J Physiol Cell Physiol. 2005;288:C1440–C1450. doi: 10.1152/ajpcell.00607.2004. [DOI] [PubMed] [Google Scholar]
- 356.Veerman EC, Nazmi K, Van't HW, Bolscher JG, Den Hertog AL, Nieuw Amerongen AV. Reactive oxygen species play no role in the candidacidal activity of the salivary antimicrobial peptide histatin 5. Biochem J. 2004;381:447–452. doi: 10.1042/BJ20040208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 357.Kumagai R, Lu X, Kassab GS. Role of glycocalyx in flow-induced production of nitric oxide and reactive oxygen species. Free Radic Biol Med. 2009;47:600–607. doi: 10.1016/j.freeradbiomed.2009.05.034. [DOI] [PMC free article] [PubMed] [Google Scholar]