

Abstract
Malignant hyperthermia (MH) is a pharmacogenetic disorder most often linked to mutations in the type 1 ryanodine receptor (RyR1) or the skeletal L-type Ca2+ channel (CaV1.1). The only effective treatment for an MH crisis is administration of the hydantoin derivative Dantrolene. In addition to reducing voltage induced Ca2+ release from the sarcoplasmic reticulum, Dantrolene was recently found to inhibit L-type currents in developing myotubes by shifting the voltage-dependence of CaV1.1 channel activation to more depolarizing potentials. Thus, the purpose of this study was to obtain information regarding the mechanism of Dantrolene-induced inhibition of CaV1.1. A mechanism involving a general depression of plasma membrane excitability was excluded because the biophysical properties of skeletal muscle Na+ current in normal mouse myotubes were largely unaffected by exposure to Dantrolene. However, a role for RyR1 was evident as Dantrolene failed to alter the amplitude, voltage dependence and inactivation kinetics of L-type currents recorded from dyspedic (RyR1 null) myotubes. Taken together, these results suggest that the mechanism of Dantrolene-induced inhibition of the skeletal muscle L-type Ca2+ current is related to altered communication between CaV1.1 and RyR1.
1. Introduction
In skeletal muscle, depolarization of the transverse tubule network causes conformational rearrangements within the sarcolemmal L-type Ca2+ channel (CaV1.1) that produce a signal which is transmitted to the type 1 ryanodine receptor (RyR1) in the sarcoplasmic reticulum (SR) membrane via a transient protein-protein interaction [1]. This “orthograde” signal gates RyR1, enabling the Ca2+ efflux from the SR into the myoplasm which ultimately initiates contraction. In addition, RyR1 produces a “retrograde” signal that enhances CaV1.1 Po [2, 3] and accelerates CaV1.1 activation kinetics [3–5]. Like orthograde coupling, retrograde coupling is thought to be propagated via protein-protein contracts between RyR1 and CaV1.1 [5–7].
Malignant hyperthermia (MH) is a fulminant pharmacogenetic disorder in which the vast majority of identified causative mutations are found in the genes encoding RyR1 [8, 9] or CaV1.1 [10–13]. MH crises are triggered by heat, depolarizing muscle relaxants, or halogenated anaesthetics [14]. Following exposure to one of these triggers, MH-susceptible individuals enter a potentially lethal hypermetabolic crisis. The only effective treatment for an MH crisis is administration of the hydantoin derivative Dantrolene, which has substantially reduced MH-related mortality since its clinical introduction in the late 1970s [15]. Despite the therapeutic success of Dantrolene, the mechanism(s) by which it ameliorates MH crises is (are) not clear. There is general agreement that one effect of Dantrolene is to stem aberrant Ca2+ efflux from the SR into the myoplasm that occurs during MH crises [15]. Dantrolene and its more soluble analogue azumolene have also been shown to reduce store-operated [16, 17] and voltage-triggered Ca2+ entry [18, 19] into muscle from the extracellular space. The major route of voltage-triggered Ca2+ entry into myotubes is the L-type Ca2+ current conducted by CaV1.1 [19, 20]. In myotubes, Dantrolene reduces such Ca2+ entry by shifting the voltage dependence of CaV1.1 activation to more depolarizing potentials [19]. Despite the aforementioned effects of Dantrolene on L-type current in mammalian muscle, the precise mechanism by which Dantrolene alters CaV1.1 channel activity has not been investigated. In this study, I have sought to determine whether the previously described depolarizing shift was a consequence of a Dantrolene-induced depression in membrane excitability or a modification of bidirectional communication between RyR1 and CaV1.1. In order to investigate the former possibility, the skeletal muscle Na+ current was employed as an assay to gauge membrane excitability. A general depression of membrane excitability appeared an unlikely explanation as Dantrolene had little effect on the biophysical properties of the Na+ current. To probe the latter mechanism, L-type Ca2+ currents were recorded from dyspedic (RyR1 null) myotubes. In these experiments, no Dantrolene-induced effects on the L-type current were observed, indicating a requirement for RyR1 expression in Dantrolene-induced inhibition of CaV1.1.
2. Experimental Procedures
2.1. Myotube Culture
The use of mice in this study was reviewed and approved by the University of Colorado Denver-Anschutz Medical Campus Institutional Animal Care and Use Committee. Primary cultures of normal (+/+ or +/mdg) or dyspedic (RyR1 −/−) myotubes were prepared from newborn mice as described previously [21]. Myoblasts were plated onto 35 mm ECL (#08-110, Millipore, Billerica, MA)-coated, plastic culture dishes (#353801, Falcon, San Jose, CA). Cultures were grown for 6-7 days in a humidified 37°C incubator with 5% CO2 in Dulbecco's Modified Eagle Medium (DMEM; #15-017-CM, Mediatech, Herndon, VA), supplemented with 10% fetal bovine serum/10% horse serum (Hyclone Laboratories, Logan, UT). This medium was then replaced with differentiation medium (DMEM supplemented with 2% horse serum). Myotubes were used in experiments 3–5 days following the switch to differentiation medium.
2.2. Patch-Clamp Recording of Skeletal Muscle Na+ and L-Type Ca2+ Currents
Pipettes were fabricated from borosilicate glass and had resistances of ~2.0 MΩ when filled with a standard internal solution containing (mM): 140 Cs-aspartate, 10 Cs2-EGTA, 5 MgCl2, and 10 HEPES, pH 7.4 with CsOH. In order to record skeletal Na+ currents, the bath solution contained (mM): 140 tetraethylammonium (TEA)-Cl, 5 NaCl, 10 CaCl2, and 10 HEPES, pH 7.4 with TEA-OH. When recording L-type Ca2+ currents, the bath solution contained (mM): 145 TEA-Cl, 10 CaCl2, and 10 HEPES, 0.002 tetrodotoxin; pH 7.4 with TEA-OH. −P/4 and P/4 subtraction were employed to correct for linear current components while recording Na+ and L-type Ca2+ currents, respectively. Electronic compensation was used to reduce the effective series resistance and the time constant for charging the linear cell capacitance. Na+ currents were filtered at 10 kHz and digitized at 50 kHz. L-type Ca2+ currents were filtered at 2 kHz and digitized at 10 kHz. Cell capacitance was determined by integration of the current transient evoked from −80 mV to −70 mV using Clampex 8.0 (Molecular Devices, Foster City, CA). All current-voltage (I-V) curves were fitted using the following equation:
| (1) |
where I is the current for test potential V, V rev is the reversal potential, G max is the maximum inward (either Na+ or Ca2+) conductance, V G is the half-maximal activation potential, and k G is the slope factor. Conductance-voltage (G-V) relationships for either Na+ or L-type Ca2+ currents were derived from the I-V data using:
| (2) |
where the V rev value for Na+ or Ca2+ in each cell was taken from (1). The average conductance values were subsequently fit with the following equation:
| (3) |
where G is the conductance for test potential V, G max is the maximal cation conductance, V 1/2act is the half-maximal activation potential, and k is the slope factor. Steady-state inactivation curves for Na+ currents were fit by the following equation:
| (4) |
where I is the current amplitude for test potential to 0 mV following a 300 ms prepulse to potential V, I max is current amplitude evoked by a test depolarization to 0 mV following a 300 ms prepulse to −110 mV, V 1/2act is the half-maximal inactivation potential, and k is the slope factor.
2.3. Pharmacology
Dantrolene (Sigma no. D9175) was dissolved in dry DMSO to make a 20 mM stock solution, diluted to 10 μM, and sonicated just prior to use. Myotubes were exposed to Dantrolene in the bath solution (~25°) for 10 to 30 minutes. Dantrolene was stored and used in the dark.
2.4. Analysis
Figures were made using the software program SigmaPlot (versions 7.0 and 11.0, SSPS Inc., Chicago, IL). All data are presented as mean ± SEM. Statistical comparisons were by unpaired, two-tailed t-test, with P < 0.05 considered significant.
3. Results
3.1. Dantrolene Does Not Affect the Fast Skeletal Muscle Na+ Current
The whole-cell patch clamp technique [22] was employed to test directly whether Dantrolene affects the fast skeletal muscle Na+ current in cultured myotubes (which is conducted by a combination of Na+ channel isoforms) [23]. With 5 mM external Na+ as the charge carrier, myotubes produced robust, rapidly-activating and -inactivating inward currents (Figure 1(a)). Myotubes exposed to Dantrolene (10 μM) for greater than 10 minutes also produced Na+ current with similar amplitude and kinetics (Figure 1(b)). As shown in Figure 1(c), the I-V relationships obtained in the absence and presence of Dantrolene displayed no significant differences in average peak current density (−9.8 ± 1.3 pA/pF, n = 20 versus −10.4 ± 1.6 pA/pF, n = 11, resp.; P > 0.05). Likewise, fitting of the average conductance values (derived from the I-V data using the reversal potential for each individual cell; see Section 2) revealed little difference in the voltage-dependence of activation (V 1/2act = −9.9 ± 1.8 mV for untreated versus −5.9 ± 3.0 mV for Dantrolene-treated, resp.; P > 0.05 t-test; Figure 1(d)).
Figure 1.
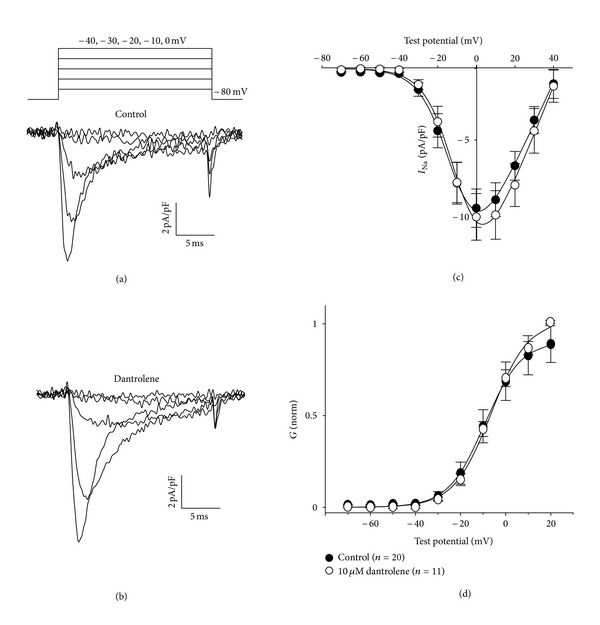
Dantrolene does not affect the I-V relationship in of the fast skeletal muscle Na+ current. Representative recordings of skeletal I Na elicited by 20 ms depolarizations from the steady holding potential of −80 mV to the indicated test potentials (illustrated at top) are shown for untreated control myotubes (a) and myotubes exposed to 10 μM Dantrolene (b). (c) Comparison of peak I-V relationships for control (⚫; n = 20) and Dantrolene-treated (⚪; n = 11) myotubes. Currents were evoked at 0.1 Hz by test potentials ranging from −70 mV through +40 mV in 10 mV increments. Peak current amplitudes were normalized by linear cell capacitance (pA/pF). Smooth I-V curves were fit by (1) (see “Section 2”) with the following respective parameters for control and Dantrolene-treated groups: G max = 305 ± 43 and 355 ± 44 nS/nF; V 1/2 = −9.8 ± 1.7 and −7.4 ± 1.3 mV; k = 6.9 ± 0.3 and 7.2 ± 0.2 mV. (d) comparison of conductance-voltage relationships for control and Dantrolene-treated myotubes. The average normalized conductance values (derived from I-V data using (2); see Section 2) were fit by (3) with the following respective parameters for control and Dantrolene-treated groups: V 1/2 = −9.9 ± 1.8 and −5.9 ± 3.0 mV; k = 6.9 ± 0.3 and 7.9 ± 1.0 mV. Throughout, data are given as mean±SEM, with the numbers in parentheses indicating the number of myotubes tested. For all the data given, the calculated average voltage error was <5 mV.
Next, the voltage protocol illustrated at the inset of Figure 2(a) was employed to determine whether Dantrolene influences steady-state inactivation of the fast Na+ current. Specifically, 300 ms prepulses (ranging from −110 mV to −10 mV) were applied immediately prior to a 20 ms test depolarization to 0 mV. Steady-state inactivation was normalized to the measured current amplitude evoked by the test depolarization to 0 mV following a prepulse to −110 mV. As shown in Figures 2(a)–2(c), steady-state inactivation of the fast Na+ current in the presence of Dantrolene was nearly indistinguishable from that observed in untreated myotubes (V 1/2inact = −41.2 ± 1.1; n = 8 versus −44.6 ± 1.6; n = 11, resp.; P > 0.05). Superimposition of the steady-state inactivation curves with conductance-voltage relationships revealed minimal window current with no obvious differences in between control and Dantrolene-treated groups (Figure 2(c)).
Figure 2.
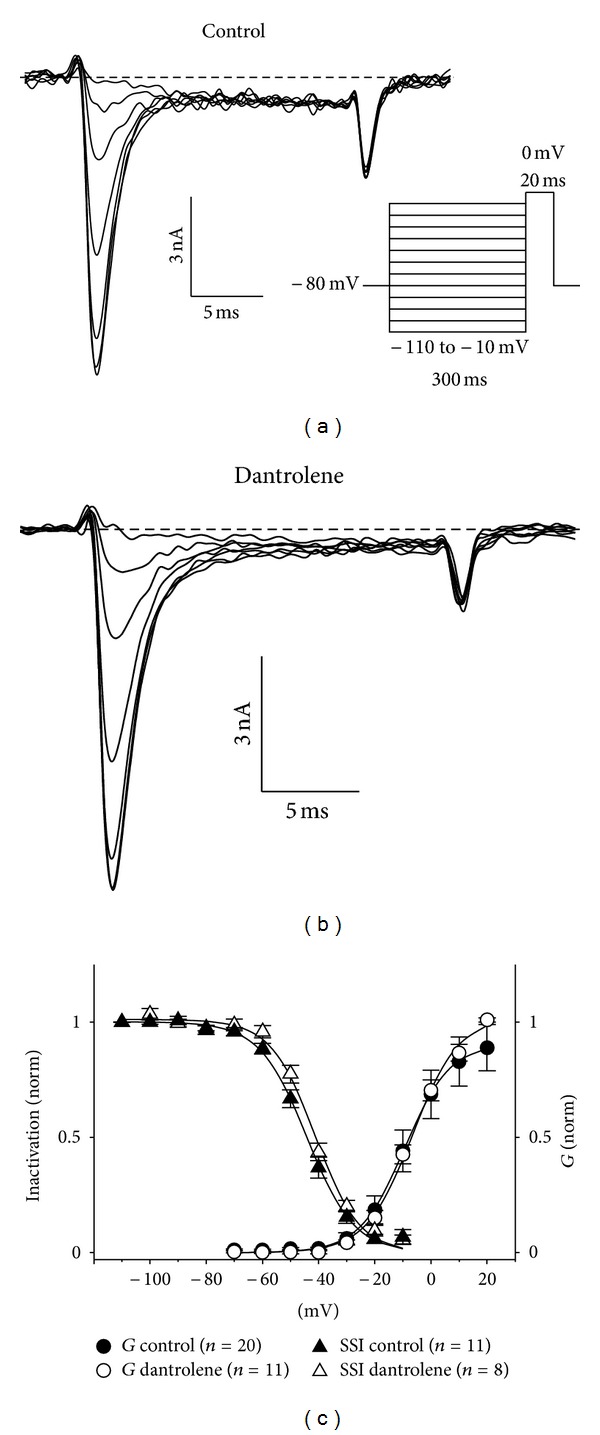
Dantrolene does not affect steady-state inactivation of the fast skeletal muscle Na+ current. Representative recordings of skeletal Na+ current elicited by 20 ms depolarizations to 0 mV immediately following 300 ms prepulses ranging from −110 mV to −10 mV are shown for untreated control myotubes (a) and myotubes exposed to 10 μM Dantrolene (b). The voltage protocol is not drawn to scale. (c) Voltage-dependence of inactivation for control (⚫; n = 11) and Dantrolene-treated (⚪; n = 8) myotubes. Inactivation was fit by (4) (see Section 2) with the following respective parameters for control and Dantrolene treated groups: V 1/2 inact = −44.3 ± 1.6 and −41.2 ± 1.1 mV; k = −8.4 ± 0.9 and −7.7 ± 0.3 mV. Inactivation curves are superimposed with activation curves shown in Figure 1 to illustrate window current.
In the representative dantrolene-treated cells shown in Figures 1 and 2, there appears to be a reduction in tail current amplitude. In both cases, tail current is reflective of the activation phase of L-type Ca2+ current and a lesser contribution of residual Na+ current. Analysis of these tail currents indicated that there is a slight, but not quite significant, difference in the peak amplitude of the tail currents at more depolarized test potentials (−12.8 ± −2.1 pA/pF; n = 10 versus 14.3 ± 1.6 pA/pF; n = 20, resp.; P > 0.05; Figure 3). When a contribution from residual Na+ current is considered, this difference may be more substantial.
Figure 3.
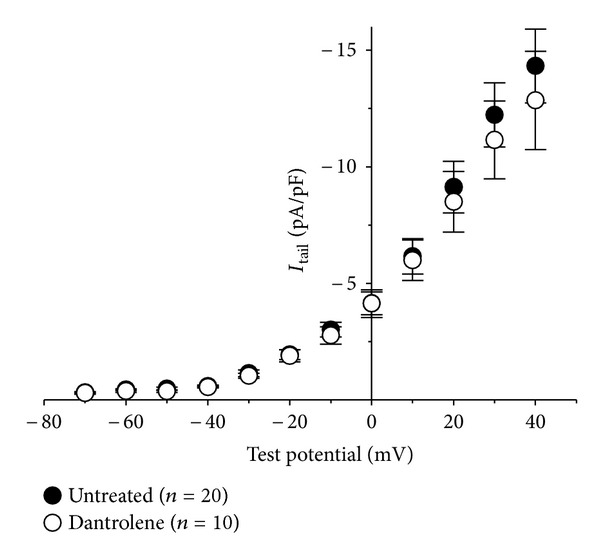
Tail current analysis. (a) Comparison of peak I-V relationships for control (⚫; n = 20) and Dantrolene-treated (⚪; n = 10) myotubes. Tail currents were elicited by repolarization from the indicated test potential to −80 mV. Currents were evoked at 0.1 Hz by test potentials ranging from −70 mV through +40 mV in 10 mV increments. Peak tail current amplitudes were normalized by linear cell capacitance (pA/pF).
The ability of Dantrolene to alter the recovery of the fast Na+ current from inactivation was tested using the voltage protocol illustrated at the top of Figure 4, in which 20 ms reference pulses evoked by depolarizations from −80 mV to 0 mV were followed by test pulses from −80 mV to 0 mV at intervals increasing in 5 ms increments. Recovery from inactivation was assessed as the fraction of the fast Na+ current evoked by the test pulse relative to that evoked by the preceeding reference pulse. In these experiments conducted in wild-type myotubes, the fractional recovery of inactivation was virtually identical for the fast Na+ current in the absence (n = 13; Figures 4(a) and 4(c)) or in the presence of 10 μM Dantrolene (n = 10; Figures 4(b) and 4(c)).
Figure 4.
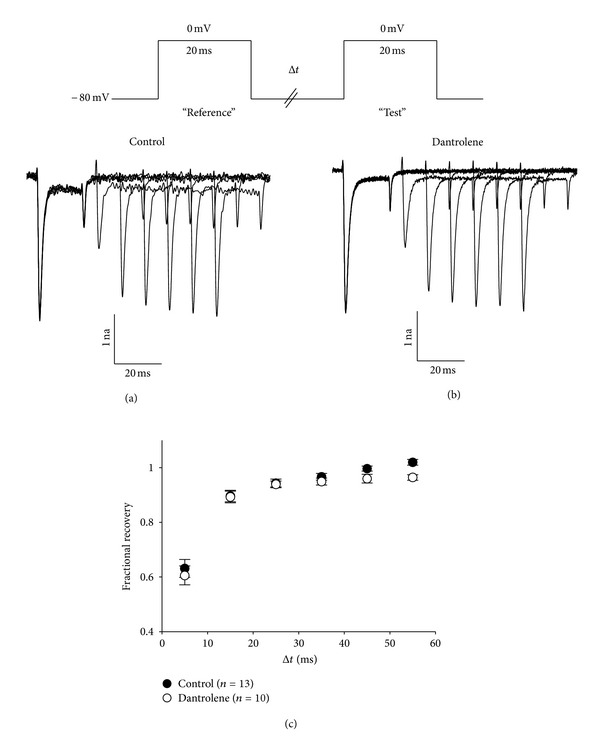
Dantrolene does not affect the recovery of the fast skeletal muscle Na+ current from inactivation. Representative recordings of skeletal Na+ current elicited by two 20 ms depolarizations from −80 mV to 0 mV separated by time intervals ranging from 5 ms to 55 ms in 10 ms increments are shown for untreated control myotubes (a) and myotubes exposed to 10 μM Dantrolene (b). (c) Summary of results.
3.2. Inhibition of L-Type Currents by Dantrolene Requires Expression of RyR1
Dantrolene inhibits voltage dependent Ca2+ entry in normal myotubes by shifting the voltage dependence of CaV1.1 gating to more depolarized potentials [19]. Since RyR1 and CaV1.1 exist in a macromolecular complex supported by protein-protein interactions [24], Dantrolene-induced conformational rearrangements of RyR1 might influence conformational changes of CaV1.1 that are involved in channel gating. To test this idea, L-type currents were recorded from dyspedic (RyR1 null) myotubes. As previously shown for dyspedic myotubes [2–5], L-type currents recorded from these cells were small (−0.74 ± 0.10 pA/pF at +30 mV, n = 8) and had more rapid activation kinetics than L-type currents typically observed in normal myotubes (Figure 5(a)) [3–5]. Exposure to Dantrolene had little effect on the amplitude (−0.79 ± 0.13 pA/pF, n = 14; P > 0.05, unpaired t-test) or activation kinetics of dyspedic L-type currents (Figure 5(b)). Importantly, no depolarizing shift in the I-V relationship was observed (V 1/2 act = 36.6 ± 4.5 mV and 28.1 ± 2.2 mV for control and dantrolene-treated, resp., P > 0.05; Figure 5(c)). However, the reversal potential for L-type currents in dantrolene-treated myotubes displayed a slight hyperpolarizing shift relative to L-type currents in control myotubes (V rev = 59.5 ± 1.7 mV and 64.0 ± 2.0 mV, resp., P > 0.05, unpaired t-test; Figure 5(c)). For this reason, the conductance-voltage relationships for control and Dantrolene-treated dyspedic myotubes conductance relationships were derived using (2) and were found to be similar (Figure 5(d)).
Figure 5.
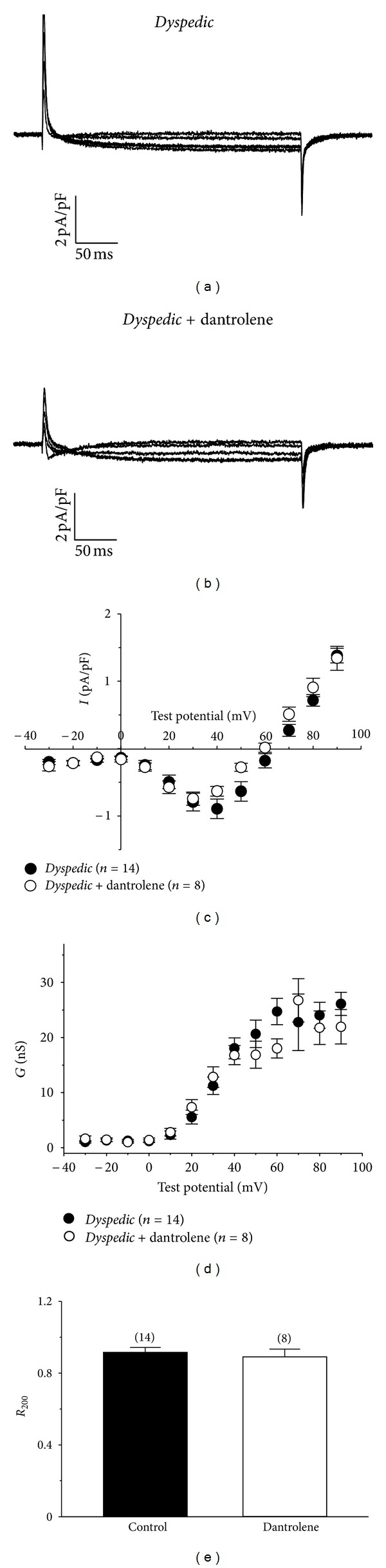
Skeletal L-type Ca2+ currents are little affected by Dantrolene in the absence of RyR1. Representative current families evoked from −80 mV to 0, 10, 20, 30, and 40 mV for dyspedic myotubes treated with DMSO vehicle (a) or 10 μM Dantrolene (b) for >10 minutes at ~25°C. In the case of the latter, the step from −80 mV to 0 mV evoked some T-type current visible at the beginning of the pulse. Peak I-V relationships are shown in (c). Smooth I-V curves were fit by (1) (see “Section 2”) with the following respective parameters for control (n = 8) and Dantrolene-treated (n = 14) groups: G max = 57 ± 6 and 43 ± 5 nS/nF; V 1/2 = 36.6 ± 4.5 and 28.1 ± 2.2 mV; k = 10.9 ± 1.6 and 11.8 ± 2.0 mV. No significant (P > 0.05, unpaired t-test) differences were observed amongst the fit parameters. (d) Comparison of conductance-voltage relationships for control and Dantrolene-treated dyspedic myotubes. The average conductance values (derived from I-V data using (2); see Section 2). (e) Inactivation summary. R 200 = fraction of the peak current remaining at the end of the 200 ms test depolarization to +30 mV.
In an earlier work, Szentesi and colleagues [25] reported that Dantrolene slows inactivation of L-type current in adult rodent fibres. To investigate whether such an effect of Dantrolene on CaV1.1 inactivation requires RyR1, inactivation of the L-type current was quantified in control and Dantrolene-treated dyspedic myotubes as the R 200 value (fraction of the peak current remaining at the end of the 200 ms test depolarization to +30 mV). As summarized in Figure 5(e), the R 200 values for control (0.91 ± 0.03, n = 14) and Dantrolene-treated (0.89 ± 0.04, n = 8) dyspedic myotubes were not significantly different (P > 0.05, unpaired t-test). Taken together, the inability of Dantrolene to modulate the skeletal L-type current in the absence of RyR1 supports the idea that these previously observed effects of Dantrolene on the L-type current [19, 25] in normal muscle resulted from altered conformational coupling between RyR1 and CaV1.1.
4. Discussion
It is widely accepted that Dantrolene inhibits excitation-contraction (EC) coupling without greatly affecting the ability of skeletal muscle fibers to conduct action potentials [26]. However, recent evidence demonstrating that Dantrolene inhibits skeletal muscle L-type Ca2+ current and/or charge movement attributed to CaV1.1 [19, 25, 27] has raised the possibility that Dantrolene may produce inhibition of CaV1.1 by altering the membrane environment. In order to probe depression of plasma membrane excitability as the mechanism of L-type channel inhibition, the effects of Dantrolene on various facets of the skeletal muscle Na+ current were investigated. Thus, exposure to Dantrolene produced no significant effect on the average peak current density, voltage dependence of activation, voltage dependence of inactivation, or recovery from inactivation of the Na+ current in developing myotubes (Figures 1–3). The inability of Dantrolene to modulate these parameters of the Na+ current in this preparation is consistent with the idea that the Dantrolene-induced inhibition of CaV1.1 observed in previous studies [19, 25, 27] most likely was a consequence of Dantrolene interacting with the EC coupling apparatus at plasma membrane-SR junctions, rather than a general nonspecific depression of plasma membrane excitability.
The observation that Dantrolene did not greatly alter the amplitude, voltage dependence, or inactivation kinetics of L-type Ca2+ currents in dyspedic myotubes (Figure 5) lends support to the idea that the inhibition of L-type current by dantrolene is unlikely a consequence of a direct interaction of Dantrolene with CaV1.1 channels. One caveat to this interpretation is that dantrolene may selectively interact with high P0 states of CaV1.1 that only occur with the influence of RyR1 [2, 3]. In either case, the data presented in Figure 5 indicate that expression of RyR1 is necessary for Dantrolene's inhibitory effects on CaV1.1 and that Dantrolene alters conformational coupling between the two channels.
Figure 6(a) shows a simplified model of “normal” bidirectional coupling between CaV1.1 and RyR1 after [2] where the green arrow and the red arrow represent orthograde (i.e., skeletal-type EC coupling) and retrograde (i.e., modulation of L-type current by RyR1) signaling, respectively. The precise mechanism of Dantrolene-induced inhibition of orthograde coupling is area of controversy. A body of evidence points to a mechanism in which Dantrolene interferes with orthograde signaling by inhibiting Ca2+ release from the SR. This view is based largely on the observations that radiolabelled Dantrolene binds to RyR1 directly [28, 29] and inhibits RyR1-mediated Ca2+ efflux from SR vesicles [30, 31]. However, other investigators have found little effect of Dantrolene on the single channel properties of RyR1 in reconstituted lipid bilayers [18, 25].
Figure 6.
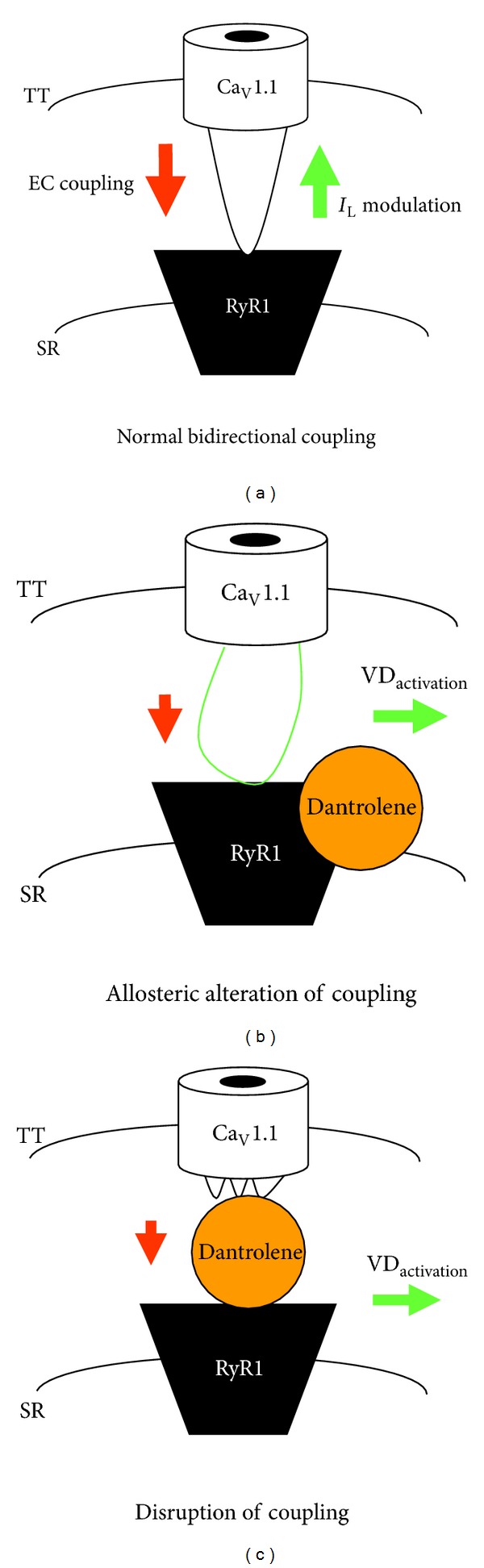
Potential models for Dantrolene-mediated inhibition of L-type Ca2+ current and excitation-contraction coupling in skeletal muscle. (a) A simplified model illustrating bidirectional coupling between RyR1 and CaV1.1, the skeletal muscle L-type Ca2+ channel [2]. The red arrow represents orthograde signal transmitted from CaV1.1 to RyR1 that engages SR Ca2+ release (i.e., EC coupling). The green arrow represents the retrograde communication transmitted from RyR1 to CaV1.1 that enhances channel P0 and modifies L-type current activation kinetics. Panels B and C show two possible mechanisms by which Dantrolene alters CaV1.1 function. (b) Binding of Dantrolene to RyR1 causes conformational changes in RyR1 that reduce SR Ca2+ release (represented by small red arrow) and shifts the activation of L-type currents to more depolarizing test potentials by altering retrograde contacts between RyR1 and CaV1.1 (represented by horizontal green arrow). (c) Alternatively, dantrolene-induced inhibition of SR Ca2+ release (again represented by small red arrow) and alterations in L-type current activation (again represented by horizontal green arrow) may be a consequence of disruption of interactions between RyR1 and CaV1.1 that are critical for bidirectional communication between the two channels.
Although my present observations do not resolve the issue of how Dantrolene inhibits orthograde coupling, they do provide support for the idea that Dantrolene's effect on retrograde signaling stems from a junctional interaction of Dantrolene with the intact (i.e., RyR1-containing) EC coupling macromolecular complex. These findings are incorporated into the two hypothetical models shown in Figures 6(b) and 6(c). In the first model (Figure 6(b)), Dantrolene would alter CaV1.1 gating indirectly by inducing allosteric rearrangements in RyR1. This model is based on the notion that agents that alter the activity of orthograde coupling by altering the functional state of RyR1 may affect retrograde signaling and vice-versa. For example, application of a high concentration of ryanodine (≥200 μM) not only attenuates EC coupling by locking RyR1 in a nonconducting state [32, 33], but also causes hyperpolarizing shifts in skeletal muscle L-type current activation [34, 35] and in charge movement [35]. Similarly, Dantrolene-induced allosteric changes in the myoplasmic region of RyR1 may affect CaV1.1 in such a way that causes a depolarizing shift in L-type current activation.
Another potential mechanism for the inhibition of CaV1.1 gating is that Dantrolene could disrupt the protein-protein interactions between RyR1 and the CaV1.1 heteromultimer that support bidirectional signaling (illustrated in Figure 6(c)). Within the context of this model, Dantrolene not only would impair EC coupling by blocking the transient interaction between the CaV1.1 channel and RyR1 but also would cause inhibition of the L-type current by removing the influence of RyR1 on CaV1.1 gating.
In summary, the anti-MH drug Dantrolene has little effect on the biophysical properties of the Na+ current or L-type Ca2+ current in developing skeletal muscle harvested from normal mice or mice lacking RyR1, respectively. These results indicate (1) that a general depression of plasma membrane excitability seems not to be responsible for inhibition of skeletal L-type Ca2+ current by Dantrolene, and (2) that RyR1 expression is necessary for the effect(s) of Dantrolene on the L-type current. Thus, this study reveals useful information towards the mechanism of Dantrolene's effect on Ca2+ currents mediated by CaV1.1.
Acknowledgments
The author thanks Drs. K. G. Beam, P.D. Allen, and I. N. Pessah for providing the resources needed to perform the study. The author is grateful to Drs. C. A. Proenza and J. D. Ohrtman for constructive criticism regarding the paper and to Ms. O. Moua for expert technical assistance. This work was supported by grants from the National Institutes of Health (AG038778 to R. A. Bannister and AR055104 to K.G.B.).
References
- 1.Beam KG, Bannister RA. Looking for answers to EC coupling’s persistent questions. Journal of General Physiology. 2010;136(1):7–12. doi: 10.1085/jgp.201010461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Nakai J, Dirksen RT, Nguyen HT, Pessah IN, Beam KG, Allen PD. Enhanced dihydropyridine receptor channel activity in the presence of ryanodine receptor. Nature. 1996;380(6569):72–75. doi: 10.1038/380072a0. [DOI] [PubMed] [Google Scholar]
- 3.Avila G, Dirksen RT. Functional impact of the ryanodine receptor on the skeletal muscle L-type Ca2+ channel. Journal of General Physiology. 2000;115(4):467–480. doi: 10.1085/jgp.115.4.467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ahern CA, Sheridan DC, Cheng W, et al. Ca2+ current and charge movements in skeletal myotubes promoted by the β-subunit of the dihydropyridine receptor in the absence of ryanodine receptor type 1. Biophysical Journal. 2003;84(2):942–959. doi: 10.1016/s0006-3495(03)74911-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sheridan DC, Takekura H, Franzini-Armstrong C, Beam KG, Allen PD, Perez CF. Bidirectional signaling between calcium channels of skeletal muscle requires multiple direct and indirect interactions. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(52):19760–19765. doi: 10.1073/pnas.0609473103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Nakai J, Sekiguchi N, Rando TA, Allen PD, Beam KG. Two regions of the ryanodine receptor involved in coupling with L-type Ca2+ channels. The Journal of Biological Chemistry. 1998;273(22):13403–13406. doi: 10.1074/jbc.273.22.13403. [DOI] [PubMed] [Google Scholar]
- 7.Grabner M, Dirksen RT, Suda N, Beam KG. The II-III loop of the skeletal muscle dihydropyridine receptor is responsible for the Bi-directional coupling with the ryanodine receptor. The Journal of Biological Chemistry. 1999;274(31):21913–21919. doi: 10.1074/jbc.274.31.21913. [DOI] [PubMed] [Google Scholar]
- 8.Robinson R, Carpenter D, Shaw MA, Halsall J, Hopkins P. Mutations in RYR1 in malignant hypertheraiia and central core disease. Human Mutation. 2006;27(10):977–989. doi: 10.1002/humu.20356. [DOI] [PubMed] [Google Scholar]
- 9.Lanner JT. Ryanodine receptor physiology and its role in disease. Advances in Experimental Medicine and Biology. 740(1):217–234. doi: 10.1007/978-94-007-2888-2_9. [DOI] [PubMed] [Google Scholar]
- 10.Monnier N, Procaccio V, Stieglitz P, Lunardi J. Malignant-hyperthermia susceptibility is associated with a mutation of the α1-subunit of the human dihydropyridine-sensitive L-type voltage- dependent calcium-channel receptor in skeletal muscle. American Journal of Human Genetics. 1997;60(6):1316–1325. doi: 10.1086/515454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Carpenter D, Ringrose C, Leo V, et al. The role of CACNA1S in predisposition to malignant hyperthermia. BMC Medical Genetics. 2009;10, article 104 doi: 10.1186/1471-2350-10-104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pirone A, Schredelseker J, Tuluc P, et al. Identification and functional characterization of malignant hyperthermia mutation T1354S in the outer pore of the Cavα1S- subunit. American Journal of Physiology. 2010;299(6):C1345–C1354. doi: 10.1152/ajpcell.00008.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Toppin PJ, Chandy TT, Ghanekar A, Kraeva N, Beattie WS, Riazi S. A report of fulminant malignant hyperthermia in a patient with a novel mutation of the CACNA1S gene. Canadian Journal of Anesthesia. 2010;57(7):689–693. doi: 10.1007/s12630-010-9314-4. [DOI] [PubMed] [Google Scholar]
- 14.Hopkins PM. Malignant hyperthermia: pharmacology of triggering. British Journal of Anaesthesia. 2011;107(1):48–56. doi: 10.1093/bja/aer132. [DOI] [PubMed] [Google Scholar]
- 15.Krause T, Gerbershagen MU, Fiege M, Weisshorn R, Wappler F. Dantrolene—a review of its pharmacology, therapeutic use and new developments. Anaesthesia. 2004;59(4):364–373. doi: 10.1111/j.1365-2044.2004.03658.x. [DOI] [PubMed] [Google Scholar]
- 16.Zhao X, Weisleder N, Han X, et al. Azumolene inhibits a component of store-operated calcium entry coupled to the skeletal muscle ryanodine receptor. The Journal of Biological Chemistry. 2006;281(44):33477–33486. doi: 10.1074/jbc.M602306200. [DOI] [PubMed] [Google Scholar]
- 17.Zhao X, Min CK, Ko JK, et al. Increased store-operated Ca2+ entry in skeletal muscle with reduced calsequestrin-1 expression. Biophysical Journal. 2010;99(5):1556–1564. doi: 10.1016/j.bpj.2010.06.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cherednichenko G, Ward CW, Feng W, et al. Enhanced excitation-coupled calcium entry in myotubes expressing malignant hyperthermia mutation R163C is attenuated by dantrolene. Molecular Pharmacology. 2008;73(4):1203–1212. doi: 10.1124/mol.107.043299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bannister RA, Pessah IN, Beam KG. The skeletal L-type Ca2+ current is a major contributor to excitation-coupled Ca2+ entry (ECCE) Journal of General Physiology. 2009;133(1):79–91. doi: 10.1085/jgp.200810105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bannister RA, Beam KG. The cardiac α1C subunit can support excitation-triggered Ca2+ entry in dysgenic and dyspedic myotubes. Channels. 2009;3(4):268–273. doi: 10.4161/chan.3.4.9342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Beam KG, Franzini-Armstrong C. Functional and structural approaches to the study of excitation-contraction coupling. Methods in Cell Biology. 1997;52(1):283–306. doi: 10.1016/s0091-679x(08)60384-2. [DOI] [PubMed] [Google Scholar]
- 22.Hamill OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Archiv. 1981;391(2):85–100. doi: 10.1007/BF00656997. [DOI] [PubMed] [Google Scholar]
- 23.Pappone PA. Voltage-clamp experiments in normal and denervated mammalian skeletal muscle fibres. Journal of Physiology. 1980;306(1):377–410. doi: 10.1113/jphysiol.1980.sp013403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Paolini C, Protasi F, Franzini-Armstrong C. The relative position of RyR feet and DHPR tetrads in skeletal muscle. Journal of Molecular Biology. 2004;342(1):145–153. doi: 10.1016/j.jmb.2004.07.035. [DOI] [PubMed] [Google Scholar]
- 25.Szentesi P, Collet C, Sárközi S, et al. Effects of dantrolene on steps of excitation-contraction coupling in mammalian skeletal muscle fibers. Journal of General Physiology. 2001;118(4):355–375. doi: 10.1085/jgp.118.4.355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ellis KO, Carpenter JF. Studies on the mechanism of action of dantrolene sodium—a skeletal muscle relaxant. Naunyn-Schmiedeberg’s Archives of Pharmacology. 1972;275(1):83–94. doi: 10.1007/BF00505069. [DOI] [PubMed] [Google Scholar]
- 27.Prosser BL, Hernández-Ochoa EO, Zimmer DB, Schneider MF. The Qγ component of intra-membrane charge movement is present in mammalian muscle fibres, but suppressed in the absence of S100A1. Journal of Physiology. 2009;587(18):4523–4541. doi: 10.1113/jphysiol.2009.177238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Palnitkar SS, Mickelson JR, Louis CF, Parness J. Pharmacological distinction between dantrolene and ryanodine binding sites: evidence from normal and malignant hyperthermia-susceptible porcine skeletal muscle. The Biochemical Journal. 1997;326(3):847–852. doi: 10.1042/bj3260847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Parness J, Palnitkar SS. Identification of dantrolene binding sites in porcine skeletal muscle sarcoplasmic reticulum. The Journal of Biological Chemistry. 1995;270(31):18465–18472. doi: 10.1074/jbc.270.31.18465. [DOI] [PubMed] [Google Scholar]
- 30.Fruen BR, Mickelson JR, Louis CF. Dantrolene inhibition of sarcoplasmic reticulum Ca2+ release by direct and specific action at skeletal muscle ryanodine receptors. The Journal of Biological Chemistry. 1997;272(43):26965–26971. doi: 10.1074/jbc.272.43.26965. [DOI] [PubMed] [Google Scholar]
- 31.Ikemoto T, Hosoya T, Aoyama H, Kihara Y, Suzuki M, Endo M. Effects of dantrolene and its derivatives on Ca2+ release from the sarcoplasmic reticulum of mouse skeletal muscle fibres. British Journal of Pharmacology. 2001;134(4):729–736. doi: 10.1038/sj.bjp.0704307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Buck E, Zimányi I, Abramson JJ, Pessah IN. Ryanodine stabilizes multiple conformational states of the skeletal muscle calcium release channel. The Journal of Biological Chemistry. 1992;267(33):23560–23567. [PubMed] [Google Scholar]
- 33.Zimányi I, Buck E, Abramson JJ, Mack MM, Pessah IN. Ryanodine induces persistent inactivation of the Ca2+ release channel from skeletal muscle sarcoplasmic reticulum. Molecular Pharmacology. 1992;42(6):1049–1057. [PubMed] [Google Scholar]
- 34.Balog EM, Gallant EM. Modulation of the sarcolemmal L-type current by alteration in SR Ca2+ release. American Journal of Physiology. 1999;276(1):C128–C135. doi: 10.1152/ajpcell.1999.276.1.C128. [DOI] [PubMed] [Google Scholar]
- 35.Bannister RA, Beam KG. Ryanodine modification of RyR1 retrogradely affects L-type Ca2+ channel gating in skeletal muscle. Journal of Muscle Research and Cell Motility. 2009;30(5-6):217–223. doi: 10.1007/s10974-009-9190-0. [DOI] [PMC free article] [PubMed] [Google Scholar]