

Abstract
Graph theoretical analysis has played a key role in characterizing global features of the topology of complex networks, describing diverse systems such as protein interactions, food webs, social relations and brain connectivity. How system elements communicate with each other depends not only on the structure of the network, but also on the nature of the system's dynamics which are constrained by the amount of knowledge and resources available for communication processes. Complementing widely used measures that capture efficiency under the assumption that communication preferentially follows shortest paths across the network (“routing”), we define analytic measures directed at characterizing network communication when signals flow in a random walk process (“diffusion”). The two dimensions of routing and diffusion efficiency define a morphospace for complex networks, with different network topologies characterized by different combinations of efficiency measures and thus occupying different regions of this space. We explore the relation of network topologies and efficiency measures by examining canonical network models, by evolving networks using a multi-objective optimization strategy, and by investigating real-world network data sets. Within the efficiency morphospace, specific aspects of network topology that differentially favor efficient communication for routing and diffusion processes are identified. Charting regions of the morphospace that are occupied by canonical, evolved or real networks allows inferences about the limits of communication efficiency imposed by connectivity and dynamics, as well as the underlying selection pressures that have shaped network topology.
Introduction
Characterizing the communication efficiency of a complex network should take into account dual sets of constraints, imposed by the topology and by the dynamical process operating on it. In this regard it is crucial to know whether communication is better described by routing (navigation) or by diffusion processes. These two types of dynamics differ radically in terms of whether interactions, matter and energy flows, or communication along paths occurs with partial or full knowledge of the global structure of the network [1], [2]. On the one hand, a routing/navigation process implies that communication flows from a specific source to a specific target along the fastest or most direct route, which implies global knowledge about the network topology. On the other hand, a diffusion process implies that communication occurs in the absence of specific targets, or that, even if targets are specified, a lack of knowledge about global network topology prevents particles or messages from taking shortest paths. To some extent, undirected diffusion or spreading occurs in social networks (innovations [3], rumors [4], contagious diseases [5]), and possibly also in neuronal communication [6].
One of the most widely used efficiency measures is defined as the average of the inverse of shortest path lengths between every pair of nodes in a graph [7], [8], here referred to as routing efficiency Erout. The use of Erout for characterizing network efficiency implies that particles or messages can selectively navigate through shortest paths and that shortening path lengths, for example through the addition of edges, automatically improves the routing efficiency of the system. However, when dynamics are best described by diffusion, shortening the path length, e.g. by adding edges, may decrease the efficiency of communication between pairs of nodes. Another difference between navigation and diffusion dynamics concerns the importance of resources, for example expressed by the number of particles needed to achieve reliable communication. In routing processes, even single particles or messages can reach their destinations along efficient paths, while in diffusion processes the ability to send multiple particles may greatly increase the efficiency of communication. Hence, characterizing the communication efficiency of a system requires considering a combination of three different characteristics: network topology (structure), the ability to find shortest paths among all alternatives (knowledge), and the capacity to increase message traffic (resources).
In this paper we first introduce and define a set of interrelated measures of communication efficiency for systems whose dynamics are based on diffusion processes. These measures capture the probability with which single particles travel through shortest paths, their average propagation velocity across the network, and the degree to which additional resources help the system to approach optimal performance. These measures of diffusion efficiency complement the classical measure of routing efficiency [8]. In the main part of the paper we take routing and diffusion efficiency as the principal axes for defining an “efficiency morphospace” [9] for complex networks, i.e. a space where each combination of routing and diffusion efficiency is associated with characteristic aspects of network topology. We explore this space following three different approaches: (i) by examining a number of canonical and idealized network models; (ii) by employing a multi-objective optimization strategy to evolve network topologies that either facilitate or impede routing and/or diffusion efficiency; and (iii) by investigating routing and diffusion efficiency in a collection of real-world systems, including brain, protein-interaction, genetic regulatory, social, virtual social, transportation and digital circuit networks. We frame our discussion in the context of what our exploration of a morphospace for communication efficiency in complex networks tells us about possible limits and selection pressures on efficient communication imposed by network structure, knowledge and resources.
Materials and Methods
Graph Theory
In this section we briefly define some basic graph theory concepts relevant to the work described in this article (see [7], [10] for detailed reviews on network theory). Let us define a binary undirected graph as 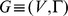 , composed by a set of
, composed by a set of  nodes
nodes 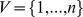 and a set of
and a set of 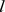 undirected edges
undirected edges  , excluding self-connections. The graph's connection topology is described by a
, excluding self-connections. The graph's connection topology is described by a 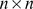 symmetrical adjacency matrix
symmetrical adjacency matrix 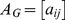 , with
, with  if edge
if edge 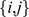 exists, and
exists, and 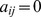 otherwise. The degree of a node
otherwise. The degree of a node  , denoted by
, denoted by 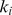 refers to the number of direct neighbors and can be expressed as
refers to the number of direct neighbors and can be expressed as 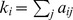 . The average degree of a graph is the mean over all node degrees
. The average degree of a graph is the mean over all node degrees 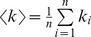 and the graph density is the fraction of edges that are present out of all possible,
and the graph density is the fraction of edges that are present out of all possible, 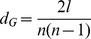 . Assortativity is defined as the Pearson correlation coefficient of all degree-degree pairs of connected nodes [11]. The heterogeneity of the graph's degree distribution can be characterized by its Shannon entropy [12], given as
. Assortativity is defined as the Pearson correlation coefficient of all degree-degree pairs of connected nodes [11]. The heterogeneity of the graph's degree distribution can be characterized by its Shannon entropy [12], given as 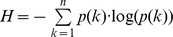 as described in [13], [14]. The graph's community structure can be determined by identifying a partition into non-overlapping modules (using an optimization algorithm such as the Louvain method [15]) that maximizes a modularity metric [16]. All graph algorithms used in this article were implemented in Matlab and are available as part of the Brain Connectivity Toolbox (www.brain-connectivity-toolbox.net) [17].
as described in [13], [14]. The graph's community structure can be determined by identifying a partition into non-overlapping modules (using an optimization algorithm such as the Louvain method [15]) that maximizes a modularity metric [16]. All graph algorithms used in this article were implemented in Matlab and are available as part of the Brain Connectivity Toolbox (www.brain-connectivity-toolbox.net) [17].
Markov Chain Theory
A Markov chain 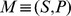 is formed by a set of states
is formed by a set of states 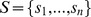 , and a matrix of transition probabilities or transition matrix
, and a matrix of transition probabilities or transition matrix
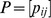 that characterizes the probability of going from one state
that characterizes the probability of going from one state 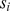 to another state
to another state 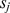 in one step [18]. A connected graph can be expressed as a Markov chain where states
in one step [18]. A connected graph can be expressed as a Markov chain where states 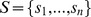 correspond element by element to the set of nodes
correspond element by element to the set of nodes 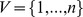 .
.
Diffusion in networks is generally modeled as a random walk process, which in the simplest case involves the use of only local information about connectivity. In a binary network, the probability to go from one state (corresponding to the source node) 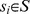 to another state (the target node)
to another state (the target node) 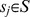 in one step is denoted by
in one step is denoted by 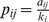 which assumes uniform probability of choosing one of the possible
which assumes uniform probability of choosing one of the possible 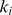 edges.
edges.
The next sections describe a set of measures characterizing communication efficiency. One measure has been introduced previously [8] and addresses communication along shortest paths, hence denoted here as routing efficiency. In addition, we introduce three related measures describing communication associated with diffusion processes, denoted shortest-path probability, diffusion efficiency and resource efficiency. All efficiency measures described in this study assume conservation of particles or walkers. The measures as defined here apply to connected undirected binary graphs, for which a diffusion process or random walk can be modeled by an ergodic (irreducible) Markov process [18]. Ergodicity is ensured by connectedness as it requires that any state can be reached from any other state in a finite number of steps. While most measures can be generalized to connected undirected weighted graphs, for simplicity we limit our analyses in the present paper to connected undirected binary graphs.
Routing Efficiency
The shortest path length between two nodes of an undirected binary graph is defined as the minimum number of edges (and thus steps) that separate two nodes within a graph. The set of shortest path lengths between all node pairs is denoted by the symmetric matrix 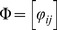 . As defined in [8], assuming parallel communication across the network, a global measure of efficiency (originally termed global efficiency Eglobal and here denoted as routing efficiency Erout), can be computed as
. As defined in [8], assuming parallel communication across the network, a global measure of efficiency (originally termed global efficiency Eglobal and here denoted as routing efficiency Erout), can be computed as
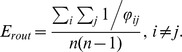 |
This measure assumes that communication operates in the shortest-path-regime.
Shortest-Path Probability
Given a random walk process on a graph, an analytic expression can be derived that gives the probability that a single particle departing from a node  arrives at node
arrives at node 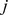 for the first time within exactly
for the first time within exactly 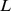 steps [19]. This criterion can be applied for each source-target pair by setting
steps [19]. This criterion can be applied for each source-target pair by setting 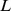 to their shortest-path-length. Let us denote by
to their shortest-path-length. Let us denote by 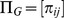 the
the 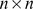 symmetric matrix containing, for each pair of nodes, the probability that a single particle going from node
symmetric matrix containing, for each pair of nodes, the probability that a single particle going from node  to node
to node 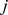 follows the shortest path (or one of them, if there exists more than one). Each entry
follows the shortest path (or one of them, if there exists more than one). Each entry 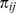 can be computed as
can be computed as
where matrix 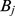 is the transition matrix
is the transition matrix 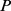 introduced above, but with all zeros in the
introduced above, but with all zeros in the 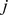 -th column, i.e. with
-th column, i.e. with  acting as an absorbing state [18], [19]. Evaluating shortest-path-lengths ensures that
acting as an absorbing state [18], [19]. Evaluating shortest-path-lengths ensures that 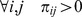 . Hence, considering one particle, the average shortest-path probability of a graph is defined as
. Hence, considering one particle, the average shortest-path probability of a graph is defined as
Diffusion Efficiency
Let us denote by the matrix 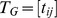 the mean first-passage time of an undirected connected graph
the mean first-passage time of an undirected connected graph 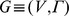 where nodes
where nodes 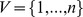 stand for states
stand for states 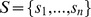 of the Markov chain. The Markov chain associated with an undirected connected graph ensures ergodicity. This property permits to compute the mean first-passage time [18] from the fundamental matrix
of the Markov chain. The Markov chain associated with an undirected connected graph ensures ergodicity. This property permits to compute the mean first-passage time [18] from the fundamental matrix 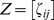 and the fixed row probability vector
and the fixed row probability vector 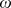 as
as
The probability vector 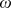 is the left eigenvector associated with the eigenvalue of 1, corresponding to the stationary solution of the underlying Markov process. The fundamental matrix
is the left eigenvector associated with the eigenvalue of 1, corresponding to the stationary solution of the underlying Markov process. The fundamental matrix 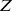 is computed as
is computed as 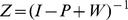 were
were 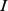 is an
is an 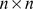 identity matrix,
identity matrix, 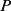 is the transition matrix defined above and
is the transition matrix defined above and 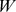 is an
is an 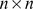 matrix with each column corresponding to the probability vector
matrix with each column corresponding to the probability vector 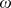 such that
such that 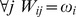 . Analogously to Erout, we express by Ediff the mean value of all inverse entries
. Analogously to Erout, we express by Ediff the mean value of all inverse entries 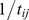 of the
of the 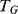 matrix excluding the main diagonal, yielding
matrix excluding the main diagonal, yielding
Resource Efficiency
Let us define 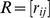 as the amount of resources (i.e. number of particles or messages) required to ensure with probability
as the amount of resources (i.e. number of particles or messages) required to ensure with probability 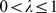 that at least one of them will, after starting at node
that at least one of them will, after starting at node  , arrive at node
, arrive at node 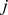 in exactly
in exactly 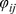 steps (i.e. as fast as possible given the graph structure):
steps (i.e. as fast as possible given the graph structure):
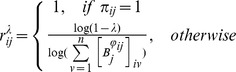 |
The simplest case arises when node 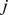 is the only neighbor of node
is the only neighbor of node  (i.e.
(i.e. 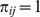 ) and thus just one particle is required for any value of
) and thus just one particle is required for any value of 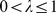 .The term
.The term 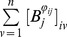 corresponds to
corresponds to 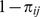 , as explained above. Note that this equation differs from the one proposed in [19] in using the shortest-path-length of each pair instead of a fixed global number of steps for all pairs. Analogously to Erout and Ediff, the efficiency of resources needed for a graph to operate in the shortest-path-regime with a fixed probability
, as explained above. Note that this equation differs from the one proposed in [19] in using the shortest-path-length of each pair instead of a fixed global number of steps for all pairs. Analogously to Erout and Ediff, the efficiency of resources needed for a graph to operate in the shortest-path-regime with a fixed probability 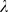 is defined as
is defined as
 |
For the analyses performed in this paper we set  .
.
Scaling of Efficiency Measures
Graph measures are known to be strongly constrained by features such as network size or density [20]. Following an approach to scaling of network metrics applied to small-world attributes such as the clustering coefficient and shortest path length [21], we scale Erout of a given network relative to the average efficiency of one-dimensional lattices and random ensembles (denoted by 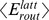 and
and 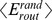 respectively) of the same size and density:
respectively) of the same size and density:
Analogously, scaled versions of 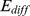 and
and 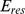 , are obtained and denoted by
, are obtained and denoted by 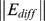 and
and 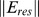 respectively.
respectively.
Efficiency Morphospace
The morphospace is a concept introduced originally in paleontology and evolutionary theory [9] to allow the systematic mapping of biological forms using structural parameters derived from a geometric model. These structural parameters define a space of all possible morphologies which specifies both existent and nonexistent forms. Here, the concept of morphology is extended to include network architectures, and the parameters defining the axes of the morphospace correspond to measures derived from network topology. Specifically, the scaled measures of routing and diffusion efficiency 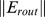 and
and 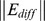 are taken to represent the principal axes of an “efficiency morphospace” within which networks are placed according to the level of communication efficiency they support. Exploring this morphospace allows probing the extent to which the space can be occupied by realizable network topologies, and what characteristic network features are encountered in different regions of the space.
are taken to represent the principal axes of an “efficiency morphospace” within which networks are placed according to the level of communication efficiency they support. Exploring this morphospace allows probing the extent to which the space can be occupied by realizable network topologies, and what characteristic network features are encountered in different regions of the space.
Canonical Network Models
Four canonical network models were used to capture the relationship between density, shortest-path probability and routing efficiency, selected because they capture relevant aspects of network organization encountered in a wide range of real-world systems. Models were Erdös-Rényi random graphs [22] (here implemented with a small variant to fix the overall density), networks generated by preferential attachment [23], one-dimensional lattices [24], and Watts-Strogatz small-world networks [24]. Details on how graphs corresponding to these canonical models were generated can be found in Text S1.
Multi-Objective Optimization
The use of optimization algorithms operating on the efficiency morphospace allowed us to explore the commonalities and differences between 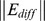 and
and 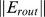 in connected undirected graphs, and the specific topological aspects that lead to such differences. The computational strategy we selected for optimization is an evolutionary algorithm operating on populations of graphs and performing incremental rewiring to change their topology. Population size, network size and density, as well as rewiring rate were selected to allow comprehensive exploration of the morphospace within the limits imposed by computational resources.
in connected undirected graphs, and the specific topological aspects that lead to such differences. The computational strategy we selected for optimization is an evolutionary algorithm operating on populations of graphs and performing incremental rewiring to change their topology. Population size, network size and density, as well as rewiring rate were selected to allow comprehensive exploration of the morphospace within the limits imposed by computational resources.
Simulations were carried out on populations (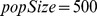 ) of undirected connected graphs of size
) of undirected connected graphs of size 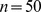 at two different densities determined by an average node degree
at two different densities determined by an average node degree 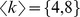 . At the beginning of each experiment, 500 one-dimensional lattices were generated. In order to create some variance in the initial population while preserving connectedness, density and degree-sequence,
. At the beginning of each experiment, 500 one-dimensional lattices were generated. In order to create some variance in the initial population while preserving connectedness, density and degree-sequence, 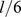 xswap rewiring steps were carried out on each lattice (see Text S1 for details of the randomization algorithm). Following this initialization, multi-objective optimizations [25], [26] were carried out, resulting in the formation of Pareto fronts [25] in the morphospace defined by our two principal efficiency measures,
xswap rewiring steps were carried out on each lattice (see Text S1 for details of the randomization algorithm). Following this initialization, multi-objective optimizations [25], [26] were carried out, resulting in the formation of Pareto fronts [25] in the morphospace defined by our two principal efficiency measures, 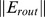 and
and 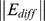 . Four different types of optimizations were implemented and run independently. Each started from the same initial population, but employed a different fitness function to explore all four directions of the two-dimensional morphospace: maximizing
. Four different types of optimizations were implemented and run independently. Each started from the same initial population, but employed a different fitness function to explore all four directions of the two-dimensional morphospace: maximizing 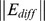 and minimizing
and minimizing 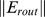 (front 1), maximizing both
(front 1), maximizing both 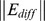 and
and 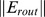 (front 2), minimizing
(front 2), minimizing 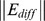 and maximizing
and maximizing 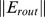 (front 3), and minimizing both
(front 3), and minimizing both 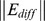 and
and 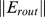 (front 4).
(front 4).
At each epoch, the Pareto front concept was applied to define survival criteria by partitioning the population into non-dominated and dominated solutions [25], which in our case are graphs. Hence a graph is said to be non-dominated (and thus a member of the Pareto front) if and only if no other graph exceeds it in the two gradients along 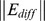 and
and 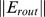 (the sign of each dimension is defined by the front and characterizes whether the objective is maximizing or minimizing each of the axes). Remaining graphs of the population not belonging to the front are said to be dominated. Pareto front members survive intact to the next population (next epoch). Finally, non-Pareto front members became extinct and were replaced in the next epoch by (randomly chosen) Pareto front members which were minimally rewired, with one edge added and one edge removed ensuring connectedness when preserving density and applying xswap when preserving degree sequence (see Text S1 for details). This process was continued over a maximum of 1000 epochs, unless the process was terminated earlier if one of two stopping criteria was reached. The first stopping criterion was based on the “age” of the Pareto front. If the age of Pareto front solutions exceeded, on average, 50 epochs, the optimization driving that front was stopped. The second stopping criterion was based on the balance between dominated and non-dominated graphs. If Pareto front members represented more than 95% of the total population, the evolutionary optimization driving that front was stopped. Two movies showing the advance of the four Pareto fronts, corresponding to the four fitness functions, for the two network densities
(the sign of each dimension is defined by the front and characterizes whether the objective is maximizing or minimizing each of the axes). Remaining graphs of the population not belonging to the front are said to be dominated. Pareto front members survive intact to the next population (next epoch). Finally, non-Pareto front members became extinct and were replaced in the next epoch by (randomly chosen) Pareto front members which were minimally rewired, with one edge added and one edge removed ensuring connectedness when preserving density and applying xswap when preserving degree sequence (see Text S1 for details). This process was continued over a maximum of 1000 epochs, unless the process was terminated earlier if one of two stopping criteria was reached. The first stopping criterion was based on the “age” of the Pareto front. If the age of Pareto front solutions exceeded, on average, 50 epochs, the optimization driving that front was stopped. The second stopping criterion was based on the balance between dominated and non-dominated graphs. If Pareto front members represented more than 95% of the total population, the evolutionary optimization driving that front was stopped. Two movies showing the advance of the four Pareto fronts, corresponding to the four fitness functions, for the two network densities 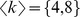 are available (Movie S1; Movie S2).
are available (Movie S1; Movie S2).
Real-World Networks
A total of 23 real-world networks were analyzed (see Text S1 for details and Table S1 for network features). For all networks, the undirected un-weighted version of the giant component was used for all the analyses and for all network and efficiency measures.
Results
Communication Efficiency in Canonical and Idealized Network Models
First we examined how increasing density affects 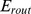 and
and 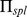 (Figure 1) in four canonical graph models: Erdös-Rényi random graphs, networks generated by preferential attachment, one-dimensional lattices, and Watts-Strogatz small-world networks.
(Figure 1) in four canonical graph models: Erdös-Rényi random graphs, networks generated by preferential attachment, one-dimensional lattices, and Watts-Strogatz small-world networks. 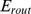 , which represents the efficiency of the shortest-path-regime, increased in all models as the average degree of the network increased due to the addition of edges. In contrast, the probability
, which represents the efficiency of the shortest-path-regime, increased in all models as the average degree of the network increased due to the addition of edges. In contrast, the probability 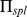 that a single particle travels along shortest paths in a diffusion process was found to be not only small (on the order of 10−2 for networks of 50 nodes) but also tended to decrease as network density increases. Over all network densities and at a given level of
that a single particle travels along shortest paths in a diffusion process was found to be not only small (on the order of 10−2 for networks of 50 nodes) but also tended to decrease as network density increases. Over all network densities and at a given level of 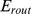 , Erdös-Rényi random graphs afforded the highest
, Erdös-Rényi random graphs afforded the highest 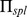 and one-dimensional lattice graphs afforded the lowest
and one-dimensional lattice graphs afforded the lowest 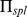 .
.
Figure 1. Relation of routing efficiency and shortest-path-probability for canonical network models.
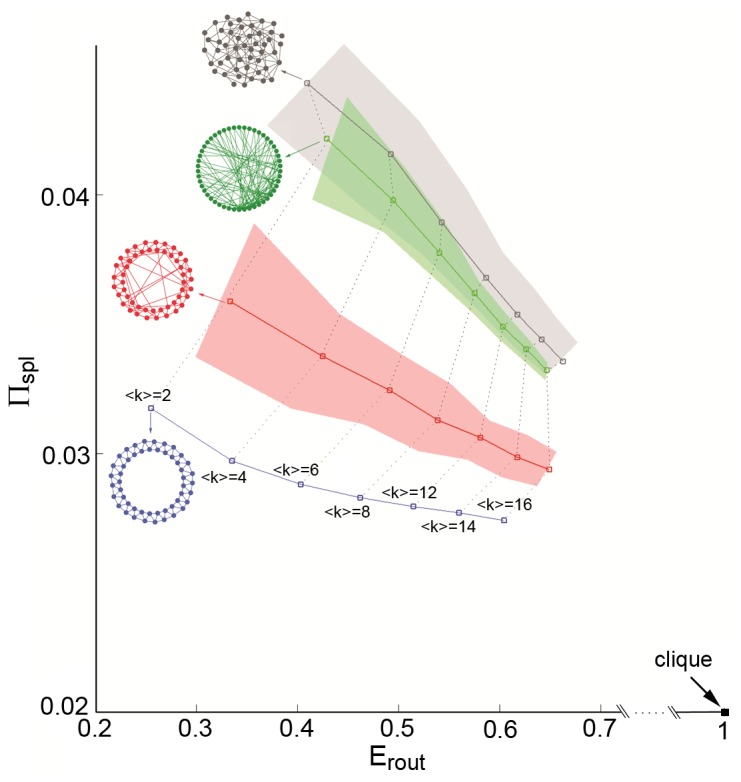
Scatter plot of 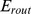 and
and 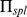 for four canonical network models at different densities: the Erdös-Rényi model (gray), the preferential attachment model (green), the Watts-Strogatz small-world model (red) and the one-dimensional lattice model (blue). The average degree is indicated at the plot line for the lattice model and dotted lines indicate equivalent average degrees for the rest of the models. Network size is fixed at 50 nodes. Each point represents mean values and shaded areas correspond to the 90% confidence interval. To obtain each data point (for each model and each average degree), 100 sample graphs were generated, except for the regular lattice given its deterministic nature. The clique coordinates are given as a reference point with maximum density and thus maximum
for four canonical network models at different densities: the Erdös-Rényi model (gray), the preferential attachment model (green), the Watts-Strogatz small-world model (red) and the one-dimensional lattice model (blue). The average degree is indicated at the plot line for the lattice model and dotted lines indicate equivalent average degrees for the rest of the models. Network size is fixed at 50 nodes. Each point represents mean values and shaded areas correspond to the 90% confidence interval. To obtain each data point (for each model and each average degree), 100 sample graphs were generated, except for the regular lattice given its deterministic nature. The clique coordinates are given as a reference point with maximum density and thus maximum 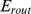 but very low
but very low 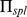 . As the density increases,
. As the density increases, 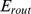 monotonically increases whereas
monotonically increases whereas 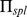 tends to monotonically decrease.
tends to monotonically decrease.
Figure 2 summarizes non-scaled efficiency measures for various idealized network topologies, with 50 nodes and varying edge densities. A clique has the shortest possible path length and hence optimal 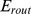 , but also low
, but also low 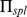 and
and 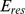 , rendering this topology well suited for routing, but poorly configured for diffusion dynamics approaching the shortest-path-regime, either with a single or with multiple particles. In contrast, chain and ring topologies have long path length and low
, rendering this topology well suited for routing, but poorly configured for diffusion dynamics approaching the shortest-path-regime, either with a single or with multiple particles. In contrast, chain and ring topologies have long path length and low 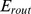 , while also exhibiting high
, while also exhibiting high 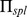 , mainly due to the low network density which limits the total number of non-optimal paths and walks.
, mainly due to the low network density which limits the total number of non-optimal paths and walks.  in chain and ring is low, indicating that a diffusion process on this graph is very inefficient. A regular lattice with
in chain and ring is low, indicating that a diffusion process on this graph is very inefficient. A regular lattice with 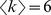 improves with respect to the ring (
improves with respect to the ring (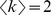 ) in both
) in both 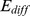 and
and 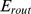 . A star has high
. A star has high 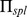 , high
, high 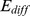 and moderately high
and moderately high 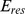 and
and 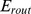 , suggesting that the star topology is well suited for both routing and diffusion processes. Similar characteristics were found in a network containing a rich club [27]. Finally, a bi-modular network, while efficient for routing, is inefficient for diffusion since the presence of modules creates bottlenecks for inter-module communication, resulting in low
, suggesting that the star topology is well suited for both routing and diffusion processes. Similar characteristics were found in a network containing a rich club [27]. Finally, a bi-modular network, while efficient for routing, is inefficient for diffusion since the presence of modules creates bottlenecks for inter-module communication, resulting in low 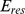 since many particles are needed to overcome these bottlenecks in inter-module communication. Larger networks (
since many particles are needed to overcome these bottlenecks in inter-module communication. Larger networks (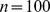 and
and 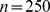 ) showed qualitatively similar results across these different network topologies.
) showed qualitatively similar results across these different network topologies.
Figure 2. Graph and efficiency measures for seven idealized network topologies.

Descriptors shown are: number of nodes  , average degree
, average degree 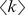 , characteristic path length
, characteristic path length 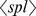 , routing efficiency
, routing efficiency 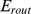 , shortest-path probability
, shortest-path probability 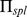 , diffusion efficiency
, diffusion efficiency 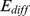 and resources efficiency
and resources efficiency 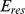 .
.
Evolving Networks in the Efficiency Morphospace
The characterization of these idealized network topologies provided some intuition about which network architectures promote different aspects of communication efficiency. However, the cases listed in Figure 2 differ greatly in density and degree sequence, both known to strongly affect virtually all graph measures. To more objectively characterize the relation of network topologies to measures of network efficiency, we implemented a multi-objective optimization strategy designed to search for network topologies in the two-dimensional efficiency morphospace formed by 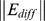 and
and 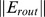 . The algorithm allowed the connection pattern (including the degree sequence and the degree distribution) to evolve through a process of incremental rewiring while preserving connectedness and density (see Text S1 for details of the rewiring methods). Using a multi-objective fitness function, selection pressure was applied to drive networks in four different directions within the scaled efficiency space, resulting in four evolving populations whose leading fronts move towards high
. The algorithm allowed the connection pattern (including the degree sequence and the degree distribution) to evolve through a process of incremental rewiring while preserving connectedness and density (see Text S1 for details of the rewiring methods). Using a multi-objective fitness function, selection pressure was applied to drive networks in four different directions within the scaled efficiency space, resulting in four evolving populations whose leading fronts move towards high 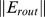 and low
and low 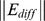 (front 1), high
(front 1), high 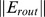 and high
and high 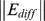 (front 2), low
(front 2), low 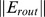 and high
and high 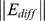 (front 3) and low
(front 3) and low 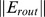 and low
and low 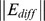 (front 4). Once evolutionary progress slowed, the fronts were considered to have reached an end state (Figure S1). At the end of each run, the surviving networks in each Pareto front can be said to jointly optimize the multiple objectives embodied in the fitness function.
(front 4). Once evolutionary progress slowed, the fronts were considered to have reached an end state (Figure S1). At the end of each run, the surviving networks in each Pareto front can be said to jointly optimize the multiple objectives embodied in the fitness function.
Figure 3 shows a set of four evolutionary simulations, each employing one of the four fitness functions, for networks with 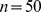 and
and 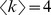 (for
(for 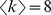 see Figure S2). Representative networks were selected across the entire length of each of the four final Pareto fronts corresponding to the evolved end states and were structurally characterized. Networks in front 2 were found to have evolved towards star-like topologies to the maximum extent permitted by the density imposed, while condensing all the remaining edges into a clique-like dense cluster. Networks in front 4 were found to have evolved towards chains as far as the imposed density permitted, with two dense clusters located at the extreme ends of the chain. Front 1 contained networks with topologies that were intermediate between those encountered in fronts 2 and 4. Along front 1, when moving away from the low-efficiency part of the front, networks gradually dissolved the chain topology into several linked clusters. Moving towards the high-efficiency part of front 1, star-like patterns began to emerge, resembling networks found in front 2. Front 3 exhibited a different transition from chain-like to star-like networks involving intermediate networks characterized by a set of densely interconnected high-degree nodes (akin to “rich club” organization [27]) that link to a number of peripheral low-degree branches. Figure S2 shows representative evolved network topologies for
see Figure S2). Representative networks were selected across the entire length of each of the four final Pareto fronts corresponding to the evolved end states and were structurally characterized. Networks in front 2 were found to have evolved towards star-like topologies to the maximum extent permitted by the density imposed, while condensing all the remaining edges into a clique-like dense cluster. Networks in front 4 were found to have evolved towards chains as far as the imposed density permitted, with two dense clusters located at the extreme ends of the chain. Front 1 contained networks with topologies that were intermediate between those encountered in fronts 2 and 4. Along front 1, when moving away from the low-efficiency part of the front, networks gradually dissolved the chain topology into several linked clusters. Moving towards the high-efficiency part of front 1, star-like patterns began to emerge, resembling networks found in front 2. Front 3 exhibited a different transition from chain-like to star-like networks involving intermediate networks characterized by a set of densely interconnected high-degree nodes (akin to “rich club” organization [27]) that link to a number of peripheral low-degree branches. Figure S2 shows representative evolved network topologies for 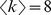 , with all Pareto fronts showing very similar network characteristics to those seen for
, with all Pareto fronts showing very similar network characteristics to those seen for 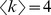 .
.
Figure 3. Multi-objective optimization in the efficiency morphospace.

Results shown are for evolutionary processes driven by network efficiency measures for networks with 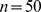 and
and 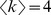 . Blue and red squares indicate the reference points of regular lattices and randomized networks respectively. Green points indicate the initial seed population. Gray circles indicate evolving networks over epochs, with darker shades of gray indicating networks encountered in later epochs. Orange points show Pareto-front (non-dominated) solutions. (a) Snapshots illustrating the expansion of the Pareto fronts at epochs 2, 25, 50, 100, and 200. (b) Final solutions were reached after 517, 704, 977, and 433 epochs for fronts 1, 2, 3, and 4 respectively. Black asterisks denote positions of the example graphs shown in insets. Yellow points show dominated solutions of the final populations. Grey points show coordinates visited during the evolutionary process at different epochs (denoted by the gray-level).
. Blue and red squares indicate the reference points of regular lattices and randomized networks respectively. Green points indicate the initial seed population. Gray circles indicate evolving networks over epochs, with darker shades of gray indicating networks encountered in later epochs. Orange points show Pareto-front (non-dominated) solutions. (a) Snapshots illustrating the expansion of the Pareto fronts at epochs 2, 25, 50, 100, and 200. (b) Final solutions were reached after 517, 704, 977, and 433 epochs for fronts 1, 2, 3, and 4 respectively. Black asterisks denote positions of the example graphs shown in insets. Yellow points show dominated solutions of the final populations. Grey points show coordinates visited during the evolutionary process at different epochs (denoted by the gray-level).
Figure 4 shows graph metrics (the entropy of the degree distribution, assortativity, modularity and normalized resource efficiency) for network topologies encountered throughout the optimization process, mapped along the two dimensions of 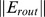 and
and 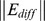 for
for 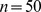 and
and 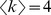 (for
(for 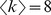 see Figure S3). Degree entropy was observed to be high on fronts 1 and 3, whereas it is lower on front 4, and lowest on front 2, due to the abundance of nodes with either low or high degree. Assortativity and modularity showed two main gradients across the space spanned by
see Figure S3). Degree entropy was observed to be high on fronts 1 and 3, whereas it is lower on front 4, and lowest on front 2, due to the abundance of nodes with either low or high degree. Assortativity and modularity showed two main gradients across the space spanned by 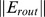 and
and 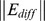 . When moving from front 2 to front 4, networks proceeded from being highly disassortative to highly assortative, and from being moderately modular to highly modular. When moving from front 1 to front 3, networks proceeded from a mixture of non-assortative and disassortative networks, to assortative networks, and from mostly modular networks to less modular networks. Resource efficiency was highest in a region surrounding the random reference point, but lower on all the fronts. With respect to the measure of resource efficiency, lattice-like and star-like topologies behaved similarly when using multiple particles to overcome a lack of structural knowledge, but for different reasons. In lattices, almost any communication between two nodes required a number of particles to ensure using the shortest path with certain probability. In star-like topologies, fast communication between perimeter nodes and central nodes required one or only a few particles, while communication between any two perimeter nodes required a greater number of particles.
. When moving from front 2 to front 4, networks proceeded from being highly disassortative to highly assortative, and from being moderately modular to highly modular. When moving from front 1 to front 3, networks proceeded from a mixture of non-assortative and disassortative networks, to assortative networks, and from mostly modular networks to less modular networks. Resource efficiency was highest in a region surrounding the random reference point, but lower on all the fronts. With respect to the measure of resource efficiency, lattice-like and star-like topologies behaved similarly when using multiple particles to overcome a lack of structural knowledge, but for different reasons. In lattices, almost any communication between two nodes required a number of particles to ensure using the shortest path with certain probability. In star-like topologies, fast communication between perimeter nodes and central nodes required one or only a few particles, while communication between any two perimeter nodes required a greater number of particles.
Figure 4. Graph measures for evolved networks.
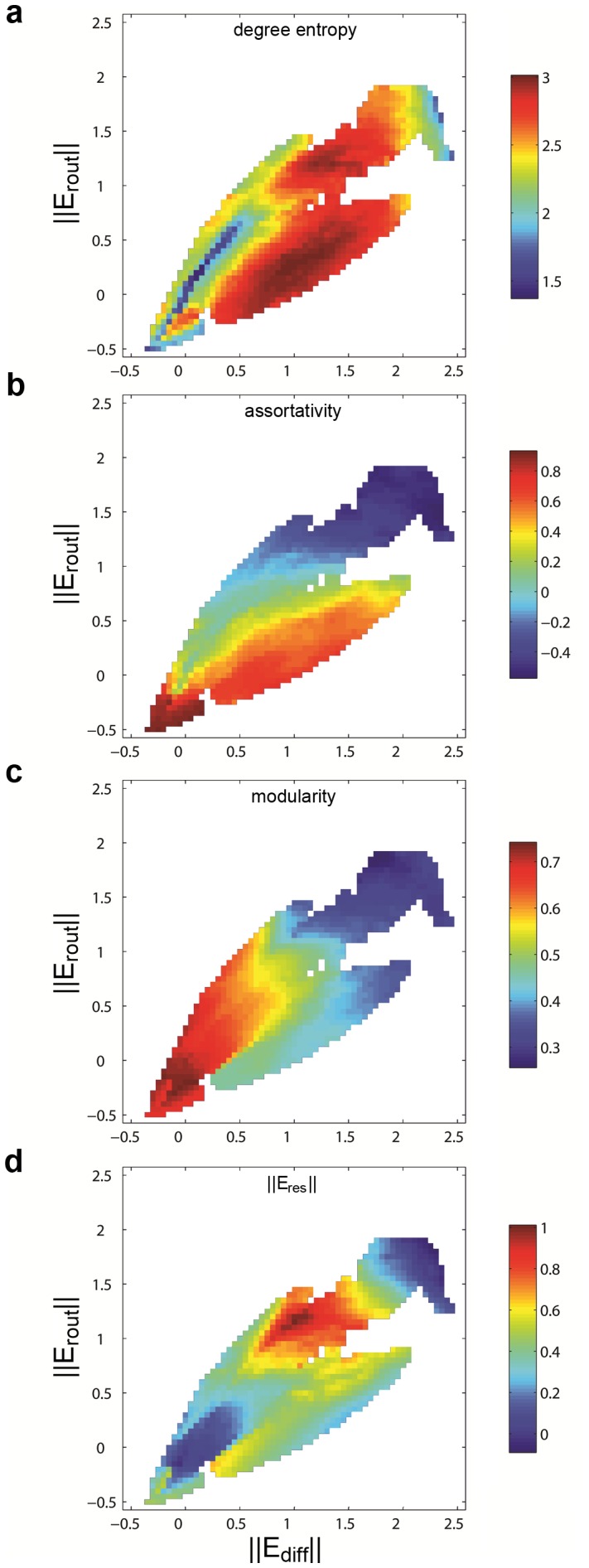
Results shown are for networks with 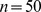 and
and 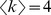 . Heat maps are based on a square grid with cells measuring 0.05 units in each dimension. For each cell, graph measures coming from graphs falling on those coordinates at any epoch of the evolutionary processes (one for each front) were averaged. (a) Degree entropy. (b) Assortativity. (c) Modularity. (d) Scaled resource efficiency
. Heat maps are based on a square grid with cells measuring 0.05 units in each dimension. For each cell, graph measures coming from graphs falling on those coordinates at any epoch of the evolutionary processes (one for each front) were averaged. (a) Degree entropy. (b) Assortativity. (c) Modularity. (d) Scaled resource efficiency 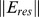 .
.
To further explore the role of the degree distribution in these evolutionary experiments, additional evolutionary optimizations were carried out where all rewiring steps were performed such that the initial uniform degree sequence (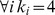 ) was maintained. Preserving the uniformity of the degree sequence was found to greatly restrict the range of
) was maintained. Preserving the uniformity of the degree sequence was found to greatly restrict the range of 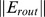 and
and 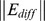 that can be reached by evolving networks (Figure S4), essentially limiting such networks to a narrow region spanning the lattice and random reference points with a very high positive co-variance between
that can be reached by evolving networks (Figure S4), essentially limiting such networks to a narrow region spanning the lattice and random reference points with a very high positive co-variance between 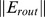 and
and 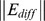 . Thus, the uniformity of the degree sequence (which corresponds to zero degree-sequence entropy) had a strong impact on network communication by contracting the region of the morphospace that is accessible and at the same time creating a strong mutual linear dependency between routing and diffusion efficiency.
. Thus, the uniformity of the degree sequence (which corresponds to zero degree-sequence entropy) had a strong impact on network communication by contracting the region of the morphospace that is accessible and at the same time creating a strong mutual linear dependency between routing and diffusion efficiency.
Real-World Networks in the Efficiency Morphospace
Figure 5 shows the efficiency morphospace represented by 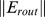 and
and 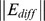 for 23 heterogeneous real-world systems, including brain, protein-interaction, genetic regulatory, social, virtual social, transportation and digital circuit networks (numerical values for network metrics are given in Table S1). Examining Figure 5, three basic observations can be made. First, it appears that most of the morphospace is empty, i.e. most combinations of
for 23 heterogeneous real-world systems, including brain, protein-interaction, genetic regulatory, social, virtual social, transportation and digital circuit networks (numerical values for network metrics are given in Table S1). Examining Figure 5, three basic observations can be made. First, it appears that most of the morphospace is empty, i.e. most combinations of 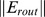 and
and 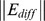 are not actually encountered in real-world networks. Second, networks found within the small region of the morphospace form an elongated cloud of points aligned with the positions of the two reference points for lattice and random topologies and their
are not actually encountered in real-world networks. Second, networks found within the small region of the morphospace form an elongated cloud of points aligned with the positions of the two reference points for lattice and random topologies and their 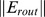 and
and 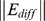 exhibits a strong and significant linear correlation (
exhibits a strong and significant linear correlation (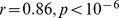 ). Third, most social, virtual social, digital circuit and transportation networks are located close to the random reference point. Protein interaction networks and genetic regulatory networks are the most efficient, exceeding the performance of equivalent random networks both in terms of diffusion and routing. Indeed, a visual inspection of these networks indicates the presence of numerous star-like patterns. Regarding brain networks, we observed a difference in networks recorded at macroscopic (whole-brain regional) and microscopic (single neuron) resolution. Whole-brain human brain networks acquired by diffusion imaging and tractography were placed within a lower efficiency regime compared to the single-neuron network of C. elegans which performed very similar to random networks for both efficiency measures.
). Third, most social, virtual social, digital circuit and transportation networks are located close to the random reference point. Protein interaction networks and genetic regulatory networks are the most efficient, exceeding the performance of equivalent random networks both in terms of diffusion and routing. Indeed, a visual inspection of these networks indicates the presence of numerous star-like patterns. Regarding brain networks, we observed a difference in networks recorded at macroscopic (whole-brain regional) and microscopic (single neuron) resolution. Whole-brain human brain networks acquired by diffusion imaging and tractography were placed within a lower efficiency regime compared to the single-neuron network of C. elegans which performed very similar to random networks for both efficiency measures.
Figure 5. Placing real-world networks in the efficiency morphospace.
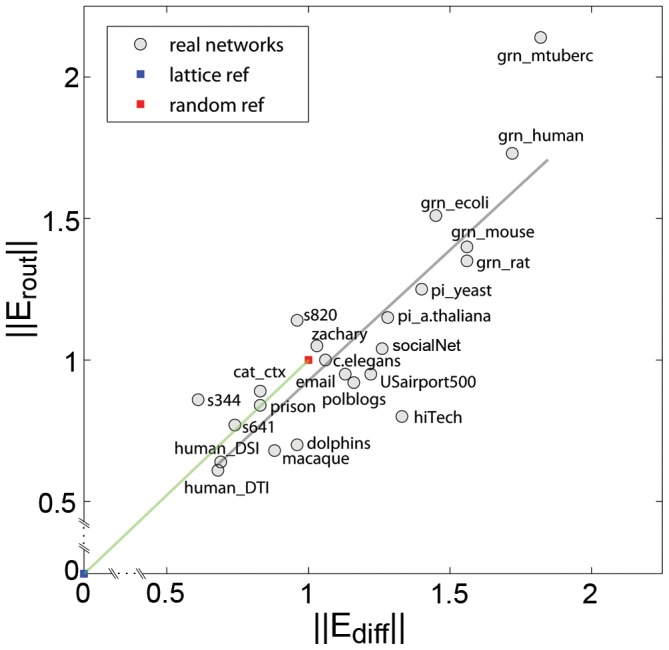
The figure shows a scatter plot of 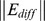 and
and 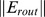 for 23 real-world networks (for description of data sets see Text S1). Blue and red squares indicate lattice and random reference points, respectively, linked by the green reference line. The gray line represents the linear regression across all 23 real-world networks (
for 23 real-world networks (for description of data sets see Text S1). Blue and red squares indicate lattice and random reference points, respectively, linked by the green reference line. The gray line represents the linear regression across all 23 real-world networks (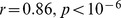 ).
).
Discussion
The efficiency of communication in complex networks is of central interest across many disciplines studying physical, social, technological or biological systems. Here we explore the relation between different measures of communication efficiency based on routing or diffusion processes, and different aspects of network topology. This exploration is carried out within an efficiency morphospace whose two axes are defined by scaled measures of routing and diffusion efficiency. Complex networks are positioned in this space depending on the level of communication efficiency they support. We explore this space by adopting three different approaches. An examination of idealized topologies, evolved architectures and real-world networks reveals characteristic differences in the way different network architectures facilitate or impede communication via routing or diffusion.
Communication efficiency has been characterized in different ways, employing routing [28], diffusion [29], [30], or more complex navigation models that include, for instance, limited information [31], communication based on hierarchical structures [32], [33], [34], selection principles with a trade-off between routing and congestion [35], or walkers capable to benefit from learning [36]. Contrasting different models for communication processes such as routing and diffusion strongly suggests that when considering global measures of network efficiency it is important to recognize that operating in the shortest-path regime requires that system elements can access complete knowledge of the global network structure. This point is particularly salient for models of network communication that are based on diffusion dynamics, traditionally characterized by random walks of single particles across the network [1], [18]. Here system elements do not possess knowledge about the global network topology and instead draw exclusively on local information when moving towards one of the neighbors of the node currently visited. This limited knowledge makes it highly improbable that a diffusion process can proceed along the shortest paths between nodes. As a foundation for exploring the relation of communication efficiency to network topology we introduced a novel set of measures designed to capture the efficiency of communication based on diffusion processes, which complement the classic measure of routing efficiency [8]. Scaling both measures to appropriate random models allowed us to position different network topologies within the space defined by 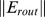 and
and 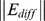 and draw relations between topology and efficiency.
and draw relations between topology and efficiency.
Star-like topologies exhibit high 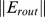 and high
and high 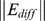 , but their high communication efficiency comes at the expense of low
, but their high communication efficiency comes at the expense of low 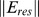 and high vulnerability to targeted attack on central nodes [37]. Modular networks exhibit higher
and high vulnerability to targeted attack on central nodes [37]. Modular networks exhibit higher 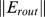 than
than 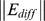 , suggesting a trade-off between shortest paths that permit efficient routing, and high clustering that prevents diffusing messages or random walkers to leave local communities and reach other parts of the network, an attribute that also results in low
, suggesting a trade-off between shortest paths that permit efficient routing, and high clustering that prevents diffusing messages or random walkers to leave local communities and reach other parts of the network, an attribute that also results in low 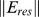 . High
. High 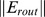 and low
and low 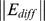 may be desirable in many real systems as the dispersal of noisy perturbations, rumors or contagious diseases remains limited to restricted parts of the network. The inverse pattern, low
may be desirable in many real systems as the dispersal of noisy perturbations, rumors or contagious diseases remains limited to restricted parts of the network. The inverse pattern, low 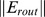 and high
and high 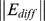 , is encountered in networks with a central and highly connected ‘core’ or ‘rich club of hubs’ linked to low density branches, with improved robustness as indexed by high degree entropy [13]. Finally, long chain structures linking highly connected modules represent network topologies with low
, is encountered in networks with a central and highly connected ‘core’ or ‘rich club of hubs’ linked to low density branches, with improved robustness as indexed by high degree entropy [13]. Finally, long chain structures linking highly connected modules represent network topologies with low 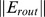 and low
and low 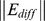 . Such networks, the antithesis of star topologies, minimize communication efficiency regardless of the nature of system dynamics and are rarely (if ever) encountered in real-world systems.
. Such networks, the antithesis of star topologies, minimize communication efficiency regardless of the nature of system dynamics and are rarely (if ever) encountered in real-world systems.
The association of certain network architectures with different levels of communication efficiency for routing and diffusion offers a new perspective on network performance. As in our multi-objective optimization experiments, key characteristics of network architecture encountered in real systems may represent the result of selection pressure on efficient communication given the constraints imposed by system dynamics and the cost of building and running the network's infrastructure [38]. Importantly, the dynamics of many real-world systems likely combine aspects of routing and diffusion. For example, depending on context, network communication in social or biological networks can draw on signaling mechanisms for discovering and navigating specific paths, or on mechanisms that aid undirected dispersion or broadcasting. In some systems, diffusion processes can degrade system performance as they allow the propagation of noisy perturbations, while diffusion may be essential in other systems where network elements have little or no knowledge about the global topology. Thus, depending on the balance and significance of routing and diffusion dynamics, network architectures need to achieve a trade-off regarding these two aspects of performance in communication. The results presented here suggest that different levels of specialization for promoting routing or diffusion can be achieved by implementing more distributed and modular versus more centralized and star-like topologies. Modular networks promote routing at the expense of diffusion, by trapping random walkers within local communities while allowing navigation between modules through inter-modular links. Thus, modular networks may be favored when system performance principally relies on routing rather than diffusion. In contrast, star-like topologies, including those containing a rich club of highly interconnected high-degree nodes, may be viewed as a “magnet” for diffusive signal traffic, attracting and dispersing information among more peripheral parts of the network. Star topologies may have a selective advantage when system dynamics utilizes both routing and diffusion. It is important to note that the propensity for real systems to implement modular or star-like topologies is subject to a number of constraints, not explicitly included in the fitness functions we explored, including structural cost, resistance to missing links, density and spatial constraints.
The apparent antagonism between dynamics dominated by diffusion versus routing can be reconciled by introducing the concept of resource efficiency. Even if the elements of a system cannot access global knowledge about network topology, this lack of knowledge can be partly overcome by multiplying the number of particles or messages (resources) used for communication. As more resources are deployed the probability that at least one particle travels along a shortest path increases. Hence, to achieve targeted (and less noisy) communication the addition of resources (involving an expense of material and/or energy) can compensate for a lack of knowledge about network structure. A different way of quantifying the knowledge needed to achieve shortest-path performance utilizes an information-theoretical approach [39]. Here we propose that the amount of resources needed over the entire graph is itself a characteristic descriptor of the graph's architecture. Network topologies not only promote communication by routing or diffusion processes, they can also afford greater resource efficiency, thus helping to reconcile the difference between routing and diffusion. Resource efficiency refers to an aspect of network cost, since resources (messages, signals) tend to consume energy and increase traffic volume. Interestingly, another aspect of network cost that impacts communication efficiency is network density. If adding edges is cheap, but multiplying resources is expensive, dense networks favoring routing over diffusion offer a better compromise between cost and efficiency. If edges are expensive, but increasing resources is more affordable, star or rich-club topologies become the more economical alternative, which more strongly favors systems with diffusive dynamics.
Our approach towards characterizing communication efficiency in networks can be extended in different directions. First, the measures and approaches introduced here could be fully explored for undirected weighted networks. Second, additional measures for characterizing diffusion in complex networks exist, including for instance the entropy rate of a diffusion process [40] and the multiple-passage time [19]. Third, other variants of random walk models can be explored, including those with degree-biased [40], “greedy routing” [41], [42] and intermediate [43] or learning-based [36] routing/diffusion strategies. Finally, all our aggregate measures can be computed for specific node pairs, allowing for instance the identification of specific sets of nodes and communication paths for which the network architecture favors diffusion and/or routing processes.
Future applications and extensions of the framework for characterizing communication efficiency proposed in this article may offer new insights into how complex networks maximize performance when their elements operate with limited knowledge and resources. Such limits are prominently encountered in, for instance, neuronal networks, where the trade-off between cost and efficiency is a major driving force of brain organization [44].
Supporting Information
Supporting information regarding networks and algorithms.
(DOCX)
Pareto fronts from an evolutionary experiment with
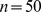 and
and
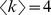 (a,b) and
(a,b) and
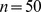 and
and
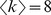 (c,d). Pareto front size refers to the number of networks it contains. The age of each member of the Pareto front is defined as the number of consecutive epochs spent in it. The Pareto front age is the average age of each member. (a,c) Evolution of Pareto front sizes for the different fronts. A large pareto front indicates a front expanding across a wide range of the search space, indicating that the in trying to satisfy the multi-objective function evolving networks cannot find a sharp gradient towards improvement. (b,d) Evolution of the Pareto front age for the different fronts. An old Pareto front indicates that the evolutionary process is unable to move the front towards better solutions in order to satisfy the multi-objective function.
(c,d). Pareto front size refers to the number of networks it contains. The age of each member of the Pareto front is defined as the number of consecutive epochs spent in it. The Pareto front age is the average age of each member. (a,c) Evolution of Pareto front sizes for the different fronts. A large pareto front indicates a front expanding across a wide range of the search space, indicating that the in trying to satisfy the multi-objective function evolving networks cannot find a sharp gradient towards improvement. (b,d) Evolution of the Pareto front age for the different fronts. An old Pareto front indicates that the evolutionary process is unable to move the front towards better solutions in order to satisfy the multi-objective function.
(TIF)
Graph evolution driven by network efficiency measures, for networks with
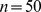 and
and
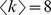 . Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Gray circles indicate evolving networks over epochs, with darker shades of gray indicating networks encountered in later epochs. Orange points show Pareto-front (non-dominated) graphs. (a) Snapshots illustrating the expansion of the Pareto fronts at epochs 2, 25, 50, 100, and 200. (b) Final solutions were reached after 1000, 1000, 1000, and 556 epochs for fronts 1, 2, 3, and 4 respectively. Black points denote positions of the example graphs shown in insets. Yellow points show dominated graphs of the final populations. Grey points show coordinates visited by any graph during the evolutionary process at different epochs (denoted by their grey-level).
. Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Gray circles indicate evolving networks over epochs, with darker shades of gray indicating networks encountered in later epochs. Orange points show Pareto-front (non-dominated) graphs. (a) Snapshots illustrating the expansion of the Pareto fronts at epochs 2, 25, 50, 100, and 200. (b) Final solutions were reached after 1000, 1000, 1000, and 556 epochs for fronts 1, 2, 3, and 4 respectively. Black points denote positions of the example graphs shown in insets. Yellow points show dominated graphs of the final populations. Grey points show coordinates visited by any graph during the evolutionary process at different epochs (denoted by their grey-level).
(TIF)
Graph measures for evolved network topologies with
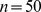 and
and
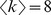 . Heat maps are based on a square grid with cells measuring 0.05 units in each dimension. For each cell, graph measures coming from graphs falling on those coordinates at any time point of the evolutionary processes (one for each front) were averaged. (a) Degree entropy. (b) Assortativity. (c) Modularity. (d) Scaled resource efficiency
. Heat maps are based on a square grid with cells measuring 0.05 units in each dimension. For each cell, graph measures coming from graphs falling on those coordinates at any time point of the evolutionary processes (one for each front) were averaged. (a) Degree entropy. (b) Assortativity. (c) Modularity. (d) Scaled resource efficiency 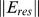 .
.
(TIF)
Graph evolution driven by network efficiency measures, for networks with
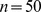 and
and
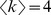 when degree-sequence is preserved. Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Orange points show Pareto-front (non-dominated) solutions. Final solutions were reached after 315, 294, 315, and 144 epochs for fronts 1, 2, 3, and 4, respectively. Yellow points show dominated solutions of the final populations. For comparison, evolving networks and Pareto fronts for networks with
when degree-sequence is preserved. Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Orange points show Pareto-front (non-dominated) solutions. Final solutions were reached after 315, 294, 315, and 144 epochs for fronts 1, 2, 3, and 4, respectively. Yellow points show dominated solutions of the final populations. For comparison, evolving networks and Pareto fronts for networks with 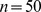 and
and 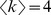 when only density and connectedness are preserved (see Figure 3) are outlined in grey.
when only density and connectedness are preserved (see Figure 3) are outlined in grey.
(TIF)
Graph and normalized efficiency measures for 23 real-world networks including brain, protein interaction, genetic regulatory, social, virtual social, transportation and digital circuit networks. Compare to Figure 5.
(DOCX)
A movie showing the complete evolutionary process towards each front for
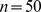 and
and
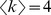 . When a front stalls, it means that its corresponding stop criterion has been reached. Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Gray circles indicate evolving networks over epochs, with darker shades of gray indicating networks encountered in later epochs. Orange points show Pareto-front (non-dominated) graphs.
. When a front stalls, it means that its corresponding stop criterion has been reached. Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Gray circles indicate evolving networks over epochs, with darker shades of gray indicating networks encountered in later epochs. Orange points show Pareto-front (non-dominated) graphs.
(AVI)
A movie showing the complete evolutionary process towards each front for
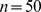 and
and
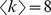 . See Movie S1 for details.
. See Movie S1 for details.
(AVI)
Acknowledgments
The authors appreciate fruitful discussions with Ricard V. Solé and Artemy Kolchinsky.
Funding Statement
This project was funded by the James S. McDonnell Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Noh JD, Rieger H (2004) Random walks on complex networks. Phys Rev Lett 92: 118701. [DOI] [PubMed] [Google Scholar]
- 2. Boguñá M, Krioukov D, Claffy KC (2009) Navigability of complex networks. Nat Phys 5: 74–80. [Google Scholar]
- 3.Gruhl D, Guha R, Liben-Nowell D, Tomkins A (2004) Information diffusion through blogspace. In Proceedings of the 13th international conference on World Wide Web (pp. 491–501). ACM.
- 4. Kostka J, Oswald YA, Wattenhofer R (2008) Word of mouth: Rumor dissemination in social networks. Lect Notes Comput Sci 5058: 185–196. [Google Scholar]
- 5. Pastor-Satorras R, Vespignani A (2001) Epidemic spreading in scale-free networks. Phys Rev Lett 86: 3200–3203. [DOI] [PubMed] [Google Scholar]
- 6. Costa LF, Travieso G (2007) Exploring complex networks through random walks. Phys Rev E 75: 016102. [DOI] [PubMed] [Google Scholar]
- 7. Boccaletti S, Latora V, Moreno Y, Chavez M, Hwang DU (2006) Complex networks: Structure and dynamics. Phys Rep 424: 175–308. [Google Scholar]
- 8. Latora V, Marchiori M (2001) Efficient behavior of small-world networks. Phys Rev Lett 87: 198701. [DOI] [PubMed] [Google Scholar]
- 9.McGhee GR (1999) Theoretical Morphology. The Concept and its Applications. Columbia University Press. New York.
- 10. Newman MEJ (2003) The structure and function of complex networks. SIAM Rev 45: 167–256. [Google Scholar]
- 11. Newman MEJ (2003) Mixing patterns in networks. Phys Rev E 67: 026126. [DOI] [PubMed] [Google Scholar]
- 12.Shannon CE, Weaver W (1949) The mathematical Theory of Information. University of Illinois Press, Urbana.
- 13. Wang B, Tang H, Guo C, Xiu Z (2006) Entropy optimization of scale-free networks' robustness to random failures. Physica A 363: 591–596. [Google Scholar]
- 14. Ferrer i Cancho R, Solé RV (2003) Optimization in complex networks. Lecture Notes Phys 625: 114–126. [Google Scholar]
- 15. Blondel VD, Guillaume JL, Lambiotte R, Lefebvre E (2008) Fast unfolding of communities in large networks. J Stat Mech P10008. [Google Scholar]
- 16. Newman MEJ, Girvan M (2004) Finding and evaluating community structure in networks. Phys Rev E 69: 026113. [DOI] [PubMed] [Google Scholar]
- 17. Rubinov M, Sporns O (2010) Complex network measures of brain connectivity: Uses and interpretations. NeuroImage 52: 1059–1069. [DOI] [PubMed] [Google Scholar]
- 18. Grinstead CM, Snell JL (2003) Introduction to Probability. Amer Mathematical Society [Google Scholar]
- 19. Wang SP, Pei WJ (2008) First passage time of multiple Brownian particles on networks with applications. Physica A 387: 4699–4708. [Google Scholar]
- 20. van Wijk BC, Stam CJ, Daffertshofer A (2010) Comparing brain networks of different size and connectivity density using graph theory. PLOS ONE 5: e13701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Sporns O, Zwi JD (2004) The small world of the cerebral cortex. Neuroinformatics 2: 145–162. [DOI] [PubMed] [Google Scholar]
- 22. Erdős P, Rényi A (1960) On the evolution of random graphs. Magyar Tud Akad Mat Kutató Int Közl 5: 17–61. [Google Scholar]
- 23. Barabasi AL, Albert R (1999) Emergence of scaling in random networks. Science 286: 509–512. [DOI] [PubMed] [Google Scholar]
- 24. Watts DJ, Strogatz SH (1998) Collective dynamics of ‘small-world’ networks. Nature 393: 440–442. [DOI] [PubMed] [Google Scholar]
- 25.Goldberg DE (1989) Genetic algorithms in search, optimization, and machine learning. Addison-Wesley, Reading, MA.
- 26. Santana R, Bielza C, Larrañaga P (2011) Optimizing brain networks topologies using multi-objective evolutionary computation. Neuroinformatics 9: 3–19. [DOI] [PubMed] [Google Scholar]
- 27. Colizza V, Flammini A, Serrano MA, Vespignani A (2006) Detecting rich-club ordering in complex networks. Nat Phys 2: 110–115. [Google Scholar]
- 28. Vragovic EL, Diaz Guilera A (2005) Efficiency of informational transfer in regular and complex networks. Phys Rev E 71: 036122. [DOI] [PubMed] [Google Scholar]
- 29. Fronczak A, Fronczak P (2009) Biased random walks in complex networks: The role of local navigation rules. Phys Rev E 80: 016107. [DOI] [PubMed] [Google Scholar]
- 30. Goñi J, Martincorena I, Corominas-Murtra B, Arrondo G, Ardanza-Trevijano S, et al. (2010) Switcher-random-walks: A cognitive-inspired mechanism for network exploration. Int J Bifurcation Chaos 20: 913–922. [Google Scholar]
- 31. Rosvall M, Minnhagen P, Sneppen K (2005) Navigating networks with limited information. Phys Rev E 71: 066111. [DOI] [PubMed] [Google Scholar]
- 32. Guimerá R, Arenas A, Diaz-Guilera A (2001) Communication and optimal hierarchical networks. Physica A 299: 247–252. [DOI] [PubMed] [Google Scholar]
- 33. Arenas A, Diaz-Guilera A, Guimerá R (2001) Communication in networks with hierarchical branching. Phys Rev Lett 86: 3196–3199. [DOI] [PubMed] [Google Scholar]
- 34. Barthélemy M, Flammini A (2006) Optimal traffic networks. J Stat Mech L07002. [Google Scholar]
- 35. Colizza V, Banavar JR, Maritan A, Rinaldo A (2004) Network structures from selection principles. Phys Rev Lett 92: 198701. [DOI] [PubMed] [Google Scholar]
- 36. Cajueiro DO, Andrade RFS (2009) Learning paths in complex networks. Europhys Lett 87: 58004. [Google Scholar]
- 37. Criado R, Flores J, Hernández-Bermejo B, Pello J, Romance M (2005) Effective measurement of network vulnerability under random and intentional attacks. J Math Model Algorithms 4: 307–316. [Google Scholar]
- 38. Solé RV, Valverde S (2004) Information theory of complex networks: On evolution and architectural constraints. Lect Notes Phys 650: 189–207. [Google Scholar]
- 39. Rosvall M, Trusina A, Minnhagen P, Sneppen K (2005) Networks and cities: An information perspective. Phys Rev Lett 94: 028701. [DOI] [PubMed] [Google Scholar]
- 40. Gomez-Gardenes J, Latora V (2008) Entropy rate of diffusion processes on complex networks. Phys Rev E 78: 065102. [DOI] [PubMed] [Google Scholar]
- 41. Kleinberg JM (2000) Navigation in a small world. Nature 406: 845. [DOI] [PubMed] [Google Scholar]
- 42. Boguñá M, Krioukov D (2009) Navigating ultrasmall worlds in ultrashort time. Phys Rev Lett 102: 058701. [DOI] [PubMed] [Google Scholar]
- 43. Cajueiro DO (2009) Optimal navigation in complex networks. Phys Rev E 79: 046103. [DOI] [PubMed] [Google Scholar]
- 44. Bullmore ET, Sporns O (2012) The economy of brain network organization. Nat Rev Neurosci 13: 336–349. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting information regarding networks and algorithms.
(DOCX)
Pareto fronts from an evolutionary experiment with
and
(a,b) and
and
(c,d). Pareto front size refers to the number of networks it contains. The age of each member of the Pareto front is defined as the number of consecutive epochs spent in it. The Pareto front age is the average age of each member. (a,c) Evolution of Pareto front sizes for the different fronts. A large pareto front indicates a front expanding across a wide range of the search space, indicating that the in trying to satisfy the multi-objective function evolving networks cannot find a sharp gradient towards improvement. (b,d) Evolution of the Pareto front age for the different fronts. An old Pareto front indicates that the evolutionary process is unable to move the front towards better solutions in order to satisfy the multi-objective function.
(TIF)
Graph evolution driven by network efficiency measures, for networks with
and
. Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Gray circles indicate evolving networks over epochs, with darker shades of gray indicating networks encountered in later epochs. Orange points show Pareto-front (non-dominated) graphs. (a) Snapshots illustrating the expansion of the Pareto fronts at epochs 2, 25, 50, 100, and 200. (b) Final solutions were reached after 1000, 1000, 1000, and 556 epochs for fronts 1, 2, 3, and 4 respectively. Black points denote positions of the example graphs shown in insets. Yellow points show dominated graphs of the final populations. Grey points show coordinates visited by any graph during the evolutionary process at different epochs (denoted by their grey-level).
(TIF)
Graph measures for evolved network topologies with
and
. Heat maps are based on a square grid with cells measuring 0.05 units in each dimension. For each cell, graph measures coming from graphs falling on those coordinates at any time point of the evolutionary processes (one for each front) were averaged. (a) Degree entropy. (b) Assortativity. (c) Modularity. (d) Scaled resource efficiency .
(TIF)
Graph evolution driven by network efficiency measures, for networks with
and
when degree-sequence is preserved. Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Orange points show Pareto-front (non-dominated) solutions. Final solutions were reached after 315, 294, 315, and 144 epochs for fronts 1, 2, 3, and 4, respectively. Yellow points show dominated solutions of the final populations. For comparison, evolving networks and Pareto fronts for networks with and when only density and connectedness are preserved (see Figure 3) are outlined in grey.
(TIF)
Graph and normalized efficiency measures for 23 real-world networks including brain, protein interaction, genetic regulatory, social, virtual social, transportation and digital circuit networks. Compare to Figure 5.
(DOCX)
A movie showing the complete evolutionary process towards each front for
and
. When a front stalls, it means that its corresponding stop criterion has been reached. Blue and red squares indicate the reference points of regular lattices and randomized networks, respectively. Green points indicate the initial seed population. Gray circles indicate evolving networks over epochs, with darker shades of gray indicating networks encountered in later epochs. Orange points show Pareto-front (non-dominated) graphs.
(AVI)
A movie showing the complete evolutionary process towards each front for
and
. See Movie S1 for details.
(AVI)