

Abstract
The objective of the present study was to determine the effects of porcine circovirus type 2 (PCV2) and porcine reproductive and respiratory syndrome virus (PRRSV) vaccinations in an experimental PCV2-PRRSV challenge model, based on virological (viremia), immunological (neutralizing antibodies [NAs], gamma interferon-secreting cells [IFN-γ-SCs], and CD4+ CD8+ double-positive cells), and pathological (lesions and antigens in lymph nodes and lungs) evaluations. A total of 72 pigs were randomly divided into 9 groups (8 pigs per group): 5 vaccinated and challenged groups, 3 nonvaccinated and challenged groups, and a negative-control group. Vaccination against PCV2 induced immunological responses (NAs and PCV2-specific IFN-γ-SCs) and reduced PCV2 viremia, PCV2-induced lesions, and PCV2 antigens in the dually infected pigs. However, vaccination against PCV2 did not affect the PRRSV immunological responses (NAs and PRRSV-specific IFN-γ-SCs), PRRSV viremia, PRRSV-induced lesions, or PRRSV antigens in the dually infected pigs. Vaccination against PRRSV did not induce immunological responses (PRRSV-specific IFN-γ-SCs) or reduce PRRSV viremia, PRRSV-induced lesions, or PRRSV antigens in the dually infected pigs. In addition, vaccination against PRRSV increased PCV2 viremia, PCV2-induced lesions, and PCV2 antigens in the dually infected pigs. In summary, vaccination against PCV2 reduced PCV2 viremia, PCV2-induced lesions, and PCV2 antigens in the dually infected pigs. However, vaccination against PRRSV increased PCV2 viremia, PCV2-induced lesions, and PCV2 antigens in the dually infected pigs. Therefore, the PCV2 vaccine decreased the potentiation of PCV2-induced lesions by PRRSV in dually infected pigs. In contrast, the PRRSV vaccine alone did not decrease the potentiation of PCV2-induced lesions by PRRSV in dually infected pigs.
INTRODUCTION
Porcine circovirus type 2 (PCV2), which is a single-stranded circular DNA virus within the family Circoviridae, is now recognized as the primary etiological agent of porcine circovirus-associated diseases (PCVAD) such as postweaning multisystemic wasting syndrome (PMWS), porcine respiratory disease complex (PRDC), and other clinical diseases (1, 2). Porcine reproductive and respiratory syndrome virus (PRRSV), which is the causative agent of PRRS, is a single-stranded, positive-sense, enveloped RNA virus in the family Arteriviridae and the order Nidovirales (3). PRRSV infection in swine is characterized by late-term abortion in gilts and sows and severe respiratory diseases in neonatal and nursing pigs (4). PRDC is a serious problem in the pig industry. PRRSV and PCV2 are the most commonly associated primary pathogens in PRDC. Epidemiological analyses have revealed that coinfection with PCV2 and PRRSV is most commonly observed in field cases (5–7).
Several studies have previously demonstrated the relationship between PCV2 and PRRSV. PCV2 does not affect PRRSV replication or lesions, but PRRSV increases PCV2 DNA loads in the sera of coinfected pigs (8, 9) and increases the levels of PCV2 antigens in tissues (10), which results in more severe PCV2-associated lesions (11). Based on these results, one possible way to minimize the effects of the PRRSV-associated enhancement of the replication of PCV2 and the induction of PMWS may be the use of a PRRSV-based vaccine in preweaned pigs. However, there are no reports in the literature describing the effects of PCV2 and PRRSV challenges on pigs that have been immunized with either PCV2 or PRRSV vaccines. In the absence of such a study, the PCV2 vaccine-PCV2-PRRSV and PRRSV vaccine-PCV2-PRRSV interactions have not been elucidated completely. Therefore, the objective of the present study was to determine the effects of PCV2 and PRRSV vaccinations in an experimental PCV2-PRRSV challenge model, based on virological (viremia), immunological (neutralizing antibodies [NAs], gamma interferon-secreting cells [IFN-γ-SCs], and CD4+ CD8+ double-positive cells), and pathological (lesions and antigens in lymph nodes and lungs) evaluations.
MATERIALS AND METHODS
Commercial vaccine.
The inactivated chimeric PCV1-2 vaccine (Fostera PCV vaccine; Pfizer Animal Health, New York, NY) and modified live PRRS vaccine (Ingelvac PRRS MLV; Boehringer Ingelheim Animal Health, St. Joseph, MO) were used in this study. The inactivated chimeric PCV1-2 vaccine contains the genomic backbone of the nonpathogenic PCV1 with the PCV2 ORF2 capsid gene in place of the PCV1 capsid gene (12). The modified live PRRS vaccine (Ingelvac PRRS MLV) originated from the American isolate ATCC VR-2332 and was attenuated by serial passages in cell culture. The vaccine contained at least 1 × 104.9 50% tissue culture infective doses (TCID50) in 2 ml. Forty pigs were vaccinated with 2.0-ml doses of either the PCV2 or PRRSV vaccine or both intramuscularly at 3 weeks of age (Table 1). All of the vaccines that were used in this study were administered according to the manufacturer's instructions (1 dose, intramuscular route).
Table 1.
Study design with vaccination and challenge statuses for PCV2 and PRRSVa
| Group no. | Vaccination (−28 dpc) |
Challenge (0 dpc) |
||
|---|---|---|---|---|
| PCV2 | PRRSV | PCV2 | PRRSV | |
| 1 | + | − | + | − |
| 2 | + | − | + | + |
| 3 | − | + | − | + |
| 4 | − | + | + | + |
| 5 | + | + | + | + |
| 6 | − | − | + | − |
| 7 | − | − | − | + |
| 8 | − | − | + | + |
| 9 | − | − | − | − |
There were eight animals in each group, and necropsy was performed at 21 dpc in all cases.
Animals and housing.
A total of 72 colostrum-fed, cross-bred, conventional piglets were weaned and purchased at 18 days of age from a PRRSV-free commercial farm. They were all negative for PCV2, PRRSV, swine influenza virus, and Mycoplasma hyopneumoniae, according to routine serological testing. PCV2 and PRRSV were not detected in serum samples by real-time PCR (13, 14). Individual piglets from 7 days of age were uniquely identified by their ear notches.
All of the pigs were housed in an environmentally controlled building with pens over completely slatted floors throughout the experiment. To avoid environmental contamination, the building was completely emptied, cleaned three times with hot (>95°C) water, and disinfected with a 2% potassium peroxymonosulfate- and sodium chloride-based product (Virkon S; Antec International, Sudbury, Suffolk, United Kingdom) for 3 days. The building was emptied for an additional 21 days before the pigs were introduced, and each pig was housed separately within the facility.
Experimental design.
A total of 72 pigs were randomly divided into 9 groups (8 pigs per group): 5 vaccinated and challenged (VC) groups (1, 2, 3, 4, and 5), 3 nonvaccinated and challenged (NVC) groups (6, 7, and 8), and a negative-control group (9).
At 21 days of age (−28 days postchallenge [dpc]), pigs in groups 1 and 2 were immunized with the PCV2 vaccine, pigs in groups 3 and 4 were immunized with the PRRSV vaccine, and pigs in group 5 were immunized with both the PCV2 and PRRSV vaccines. At 49 days of age (0 dpc), pigs in the VC and NVC groups were intratracheally administered an intranasal 3-ml dose of PCV2b (strain SNUVR000463; 5th passage) containing 1.2 × 105 TCID50/ml and/or a 3-ml dose of PRRSV (strain SNUVR090851; North American genotype; 5th passage) containing 1 × 105 TCID50/ml. Pigs in groups 1 and 6 were challenged with PCV2. Pigs in groups 3 and 7 were challenged with PRRSV. Pigs in groups 2, 4, 5, and 8 were challenged with both PCV2 and PRRSV (Table 1).
Serology.
Blood samples from each pig were collected by jugular venipuncture at −28, 0, 10, and 21 dpc, and the sera were stored at −20°C. The serum samples were tested using a commercial PRRSV enzyme-linked immunosorbent assay (ELISA) (HerdCheck PRRS 2XR; Idexx Laboratories Inc., Westbrook, ME) and a PCV2 ELISA (Synbiotics, Lyon, France). A serum virus neutralization (SVN) test for PCV2 was performed as previously described (15). NA titers were expressed as the reciprocal of the highest serum dilution that completely blocked the infection in PK15 cells compared with the virus control. An SVN test for PRRSV was also performed as previously described (16).
Quantification of PCV2 DNA in blood.
DNA was extracted from the serum samples obtained at −28, 0, 10, and 21 dpc by use of a QIAamp DNA minikit (Qiagen Ltd., Crawley, United Kingdom). The DNA extracts were used to quantify the PCV2 genomic DNA copy numbers by real-time PCR as previously described (13).
Quantification of PRRSV RNA in blood.
RNA was extracted from the serum samples obtained at −28, 0, 10, and 21 dpc for all of the pigs that were used in this study, as previously described (14, 17). Real-time PCRs for the PRRSV and vaccine strains were used to quantify the PRRSV genomic cDNA copy numbers following RNA extraction of the serum samples, which was performed as previously described (14, 17).
Preparation of PCV2 and PRRSV antigens.
The same PCV2 strain (or PRRSV strain) that was used for the challenge in the pigs was propagated in PCV-free PK15 cells (or MARC-145 cells for the PRRSV strain) to a titer of 104 TCID50/ml and then treated with two freeze-thaw cycles. The PCV2 antigen was prepared by concentrating the virus that was present in the cell culture by ultracentrifugation at 100,000 × g at 4°C for 3 h. The virus pellet was resuspended in phosphate-buffered saline (PBS). The concentrated PCV2 (or PRRSV) was inactivated by exposure to an 8-W germicidal UV lamp at a distance of 15 cm for 1 h. Inactivation was confirmed by the absence of the virus antigen from the PK15 cells (or MARC-145 cells for the PRRSV strain) as determined by an immunoperoxidase assay, as previously described (18, 19).
ELISPOT assay.
The numbers of PCV2- and PRRSV-specific IFN-γ-SCs were determined for peripheral blood mononuclear cells (PBMCs) obtained at −28, 0, 10, and 21 dpc as previously described (20). Briefly, 100 μl containing 2 × 106 PBMCs in RPMI 1640 medium that was supplemented with 10% fetal bovine serum (HyClone Laboratories, Inc., SelectScience, Bath, United Kingdom) was seeded into plates that were precoated overnight with anti-porcine IFN-γ monoclonal antibody (5 μg/ml) (Mabtech, Mariemont, OH) and incubated with 100 μl of PCV2 antigen (20 μg/ml), PRRSV antigen (20 μg/ml), and phytohemagglutinin (10 μg/ml) (Roche Diagnostics GmbH, Mannheim, Germany) as positive controls or with PBS as a negative control for 40 h at 37°C in a 5% humidified CO2 atmosphere. The wells were then washed five times with PBS (200 μl per well). Thereafter, the procedure was conducted according to the manufacturer's instructions, using a commercial enzyme-linked immunosorbent spot (ELISPOT) assay kit (Mabtech, Mariemont, OH). The spots on the membranes were read by an automated ELISPOT reader (AID ELISpot reader; AID GmbH, Strassberg, Germany). The results were expressed as numbers of IFN-γ-SCs per million PBMCs.
Flow cytometry.
Phenotypic analyses of the CD4+ CD8+ double-positive cells in PBMCs were performed using flow cytometry with two monoclonal antibodies, i.e., CD4a–R-phycoerythrin (CD4a–R-PE) and CD8a-fluorescein isothiocyanate (CD8a-FITC) (SouthernBiotech, Birmingham, AL), and a FACSCalibur flow cytometer (Becton, Dickinson, San Jose, CA) as previously described (21).
Immunohistochemistry.
Superficial inguinal lymph nodes that had previously shown consistent and intense labeling for PCV2 and typical granulomatous lesions (22) were collected at necropsy for immunohistochemical analyses. A rabbit polyclonal anti-PCV2 antibody (23) (1:200 in PBS containing 0.1% Tween 20) and the SR30 monoclonal antibody (Rural Technologies Inc., Brookings, SD) (1:10,000 in PBS containing 0.1% Tween 20) were used as the primary antibodies. Immunohistochemical analyses for PCV2 and PRRSV were performed as previously described (24, 25).
Morphometric analyses.
For morphometric analyses of the microscopic pulmonary and lymph lesion scores, lung sections were examined blindly and assigned estimated scores ranging from 1 (mild interstitial pneumonia) to 4 (severe interstitial pneumonia), as previously described (26). Superficial inguinal lymph node sections were also examined blindly, and their scores ranged from 0 (normal, i.e., no lymphoid depletion or granulomatous replacement) to 5 (severe lymphoid depletion and granulomatous replacement), as previously described (27).
For immunohistochemistry morphometric analyses, 3 sections were cut from each of three blocks of tissue from one entire pulmonary lobe and from the superficial inguinal lymph nodes of each pig. The slides were analyzed using the NIH Image J 1.43m program (http://rsb.info.nih.gov/ij) to obtain the quantitative data. For each slide, 10 fields were randomly selected, and the number of positive cells per unit area (0.95 mm2 for the pulmonary lobe and 0.25 mm2 for the lymph node) was determined as previously described (22, 28). The mean values were also calculated.
Statistical analyses.
Summary statistics were calculated for all of the groups to assess the overall quality of the data, including normality. For single comparisons, analysis of variance (ANOVA) with a post hoc Tukey's test was used to compare the primary variables (immunohistochemical scores) among groups. The continuous data for PCV2 and PRRSV serology and PCV2 DNA and PRRSV cDNA quantifications were analyzed using an ANOVA for each time point. When one-way ANOVA revealed a significance level with a P value of <0.05, Tukey's honestly significant difference test was used to determine the significance of individual between-group differences. Discrete data (histopathologic lung and lymphoid lesion scores) were analyzed by the chi-square and Fisher's exact tests. P values of <0.05 were considered to be significant.
RESULTS
Serology of PCV2.
At challenge, all pigs vaccinated against PCV2 were seropositive, and all nonvaccinated pigs were negative by ELISA and SVN tests. Pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1), pigs which received the PCV2 vaccine followed by dual challenge (group 2), and pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5) had significantly higher anti-PCV2 IgG antibody levels (P < 0.001) and NA titers (P < 0.01) (Fig. 1A) than those of pigs which received the PRRSV vaccine followed by dual challenge (group 4), pigs which were challenged with PCV2 only (group 6), and pigs which were challenged with both PCV2 and PRRSV (group 8) at 10 and 21 dpc. Pigs in group 6 had significantly higher anti-PCV2 IgG antibody levels (P < 0.01) than those of pigs in groups 4 and 8 at 21 dpc.
Fig 1.
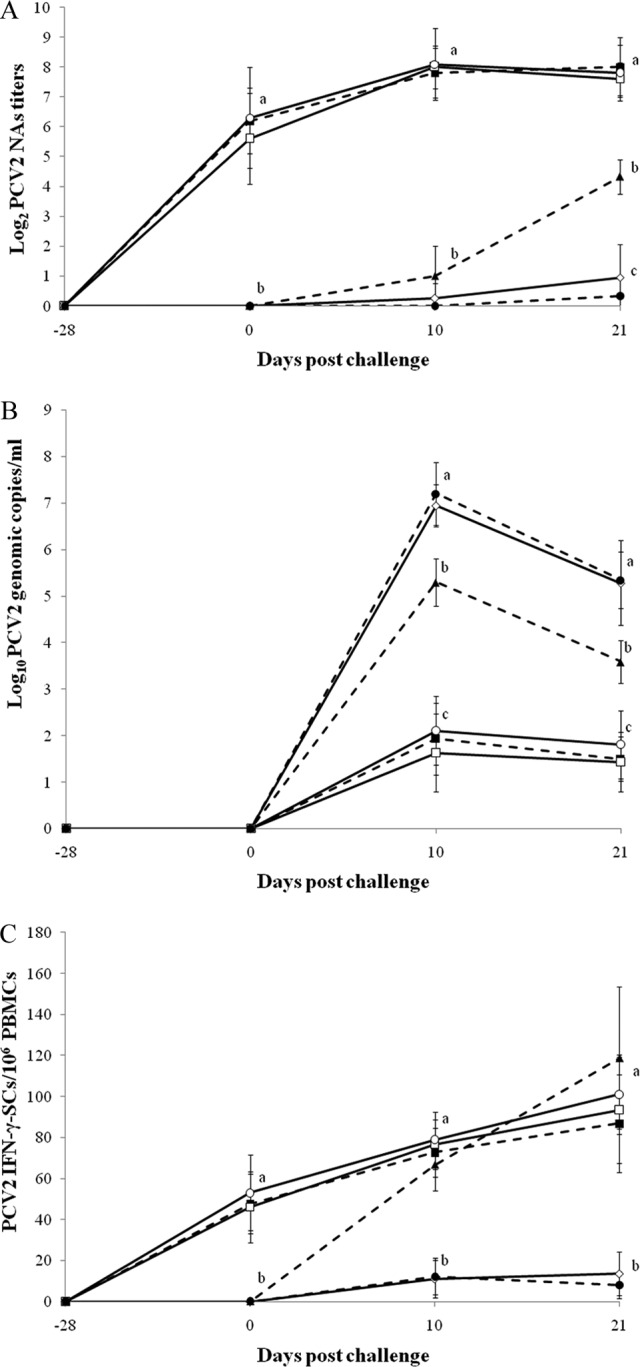
(A) Mean serum NA titers against PCV2. (B) Mean genomic copy numbers of PCV2 DNA in serum samples. (C) Mean numbers of PCV2-specific IFN-γ-SCs in PBMCs. ■, pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1); □, pigs which received the PCV2 vaccine followed by dual challenge (group 2); ♢, pigs which received the PRRSV vaccine followed by dual challenge (group 4); ○, pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5); ▲, pigs which were challenged with PCV2 (group 6); ●, pigs which were challenged with PCV2 and PRRSV (group 8). Different letters (a, b, and c) indicate significant (P < 0.05) differences between groups.
Serology of PRRSV.
At challenge, all pigs vaccinated against PRRSV were seropositive, and all nonvaccinated pigs were negative by ELISA. Pigs which received the PRRSV vaccine followed by PRRSV challenge only (group 3), pigs which received the PRRSV vaccine followed by dual challenge (group 4), and pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5) had significantly higher anti-PRRSV IgG antibody levels (P < 0.001) than pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which were challenged with PRRSV only (group 7), and pigs which were challenged with both PCV2 and PRRSV (group 8) at 10 dpc. Pigs which received PRRSV followed by PRRSV challenge only (group 3) and pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5) had significantly higher anti-PRRSV IgG antibody levels (P < 0.05) than pigs in the other 4 groups (2, 4, 7, and 8) at 21 dpc (Fig. 2A). Low PRRSV NA titers (<2 log2) were detected in all 8 groups throughout the experiment.
Fig 2.
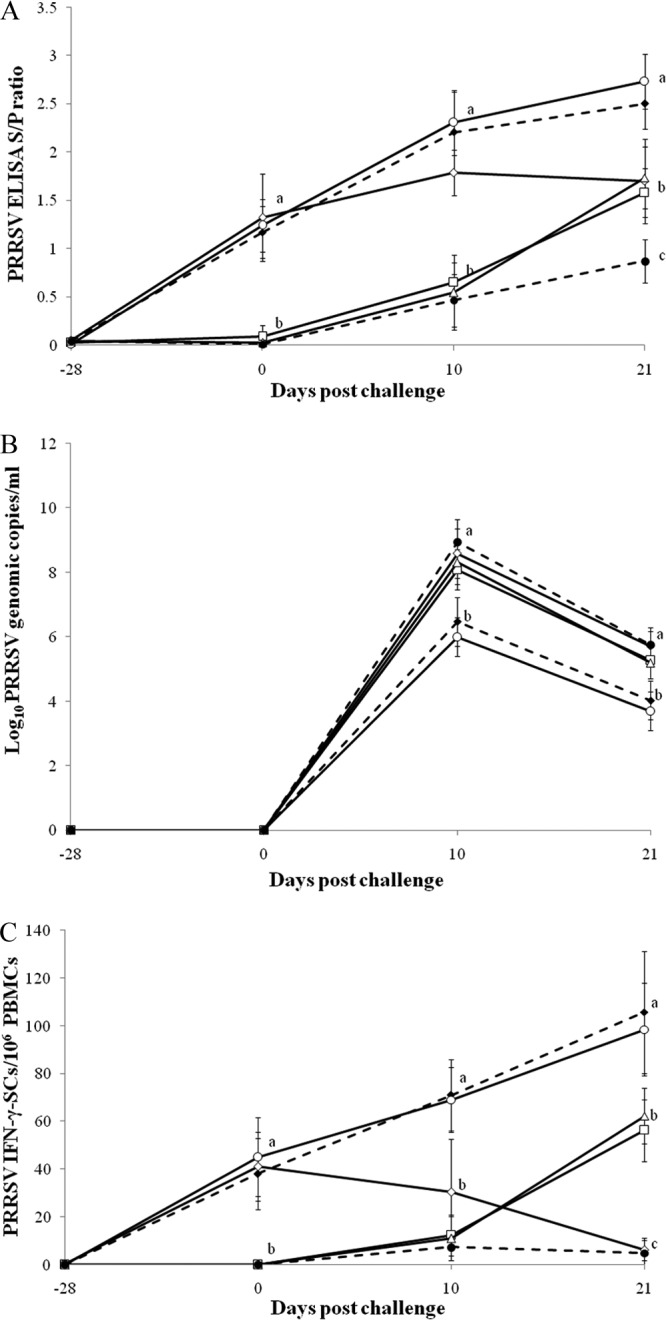
(A) Mean anti-PRRSV IgG antibody levels in serum samples. (B) Mean genomic copy numbers of PRRSV DNA in serum samples. (C) Mean numbers of PRRSV-specific IFN-γ-SCs in PBMCs. □, pigs which received the PCV2 vaccine followed by dual challenge (group 2); ◆, pigs which received the PRRSV vaccine followed by PRRSV challenge (group 3); ♢, pigs which received the PRRSV vaccine followed by dual challenge (group 4); ○, pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5); △, pigs which were challenged with PRRSV (group 7); ●, pigs which were challenged with PCV2 and PRRSV (group 8). Different letters (a, b, and c) indicate significant (P < 0.05) differences between groups. S/P, sample to positive ratio of ≥0.4 is considered to be positive.
Quantification of PCV2 DNA in blood.
At challenge, no genomic copies of PCV2 were detected in any of the serum samples from all 8 groups. Pigs which received the PRRSV vaccine followed by dual challenge (group 4) and pigs which were challenged with both PRRSV and PCV2 (group 8) had significantly more genomic copies of PCV2 in the serum than pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1), pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5), and pigs which were challenged with PCV2 only (group 6) at 10 and 21 dpc (P < 0.05). Pigs which were challenged with PCV2 only (group 6) had significantly more genomic copies of PCV2 in the serum than pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1), pigs which received the PCV2 vaccine followed by dual challenge (group 2), and pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5) at 10 and 21 dpc (P < 0.05) (Fig. 1B). No genomic copies of PCV2 were detected in any of the serum samples from nonchallenged pigs (groups 3, 7, and 9) throughout the experiment.
Quantification of PRRSV cDNA in blood.
At challenge, no genomic copies of PRRSV were detected in any of the serum samples from all 8 groups. Pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received the PRRSV vaccine followed by dual challenge (group 4), pigs which were challenged with PRRSV only (group 7), and pigs which were challenged with both PCV2 and PRRSV (group 8) had significantly more genomic copies of PCV2 in the serum than pigs which received the PRRSV vaccine followed by PRRSV infection only (group 3) and pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5) at 10 and 21 dpc (P < 0.05) (Fig. 2B). No genomic copies of PRRSV were detected in any of the serum samples from nonchallenged pigs (groups 1, 6, and 9) throughout the experiment.
PCV2-specific IFN-γ-SCs.
No PCV2-specific IFN-γ-SCs were detected in PBMCs at −28 dpc for any of the pigs. PCV2-specific IFN-γ-SCs were detected in PBMCs from the PCV2-vaccinated pigs (groups 1, 2, and 5) at 0 dpc. Pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1), pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5), and pigs which were challenged with PCV2 only (group 6) had significantly more PCV2-specific IFN-γ-SCs than pigs which received the PRRSV vaccine followed by dual challenge (group 4) and pigs which were challenged with both PCV2 and PRRSV (group 8) at 10 and 21 dpc (P < 0.01) (Fig. 1C).
PRRSV-specific IFN-γ-SCs.
No PRRSV-specific IFN-γ-SCs were detected in PBMCs at −28 dpc for any of the pigs. PRRSV-specific IFN-γ-SCs were detected in PBMCs from the PRRSV-vaccinated pigs (groups 3, 4, and 5) at 0 dpc. Pigs which received the PRRSV vaccine followed by PRRSV infection only (group 3) or followed by dual challenge (group 5) had significantly more PRRSV-specific IFN-γ-SCs than pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received the PRRSV vaccine followed by dual challenge (group 4), pigs which were challenged with PRRSV only (group 7), and pigs which were challenged with both PCV2 and PRRSV (group 8) at 10 and 21 dpc (P < 0.01). Pigs which received the PCV2 vaccine followed by dual challenge (group 2) and pigs which were challenged with PRRSV only (group 7) had significantly more PRRSV-specific IFN-γ-SCs than the pigs in 2 groups (groups 4 and 8) at 21 dpc (P < 0.01) (Fig. 2C).
Identification of CD4+ CD8+ double-positive cells.
Pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1), pigs which received the PCV2 vaccine followed by dual challenge (group 2), and pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5) had significantly larger proportions of CD4+ CD8+ double-positive cells than pigs which received the PRRSV vaccine followed by dual challenge (group 4), pigs which were challenged with both PCV2 and PRRSV (group 8), and negative-control pigs (group 9) at 21 dpc (P < 0.05). Negative-control pigs had significantly larger proportions of CD4+ CD8+ double-positive cells than pigs which received the PRRSV vaccine followed by dual challenge (group 4) and pigs which were challenged with both PCV2 and PRRSV (group 8) at 21 dpc (P < 0.05) (Fig. 3A).
Fig 3.

Analyses of CD4+ CD8+ double-positive cells (A) and CD4+ cells (B) in PBMCs from the different groups. From left to right, the bars indicate the following groups: pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1), pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received the PRRSV vaccine followed by PRRSV challenge (group 3), pigs which received the PRRSV vaccine followed by dual challenge (group 4), pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5), pigs which were challenged with PCV2 (group 6), pigs which were challenged with PRRSV (group 7), pigs which were challenged with PCV2 and PRRSV (group 8), and negative-control pigs (group 9). Different letters (a, b, and c) indicate significant (P < 0.05) differences between groups.
Pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1), pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received the PRRSV vaccine followed by PRRSV infection only (group 3), pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5), and negative-control pigs (group 9) had significantly larger proportions of CD4+ cells than pigs which received the PRRSV vaccine followed by dual challenge (group 4) and pigs which were challenged with both PCV2 and PRRSV (group 8) at 21 dpc (P < 0.03) (Fig. 3B).
Histopathologic lesion scores.
The typical granulomatous inflammatory reaction and lymphoid depletion that are associated with PCV2 infection in pigs, which are consistent with the histopathologic lesions in PCVAD, were observed in the lymph nodes from pigs which received the PRRSV vaccine followed by dual challenge (group 4) (Fig. 4A) and pigs which were challenged with both PCV2 and PRRSV (group 8). Pigs in these groups (4 and 8) had significantly higher lymphoid lesion scores than pigs in the other 6 groups (1, 2, 3, 5, 6 [Fig. 4B], and 7) (P < 0.03). Pigs which were challenged with PCV2 only (group 6) had significantly higher lymphoid lesion scores than pigs in the other 5 groups (1, 2, 3, 5, and 7) (P < 0.03). No histopathologic lymph node lesions were observed in the negative-control pigs (group 9).
Fig 4.

Immunohistochemistry for detection of PCV2 antigens in lymph nodes. (A) PCV2 antigens (arrows) were detected at 21 dpc in granulomatous inflammations of lymph nodes from pigs which received the PRRSV vaccine followed by dual challenge (group 4). (B) PCV2 antigens (arrow) were occasionally detected at 21 dpc in macrophages of lymph nodes from pigs which were challenged with PCV2 (group 6).
The pulmonary histopathologic lesions were characterized by alveolar proteinaceous and karyorrhectic debris that was interspersed with macrophages. The alveolar septa in the pigs which were challenged with both PCV2 and PRRSV (group 8) were thickened with increased numbers of interstitial macrophages and lymphocytes. Pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received the PRRSV vaccine followed by dual challenge (group 4) (Fig. 5A), pigs which were challenged with PRRSV only (group 7), and pigs which were challenged with both PCV2 and PRRSV (group 8) had significantly higher pulmonary lesion scores than pigs in the other 4 groups (1, 3 [Fig. 5B], 5, and 6) (P < 0.05) at 21 dpc (Table 2). The lungs of the negative-control pigs (group 9) were normal.
Fig 5.

Immunohistochemistry for detection of PRRSV antigens in the lung. (A) PRRSV antigens (arrows) were detected at 21 dpc in macrophages in severely thickened alveolar septa of lungs from pigs which received the PRRSV vaccine followed by dual challenge (group 4). (B) PRRSV antigens (arrow) were occasionally detected at 21 dpc in macrophages in mildly thickened alveolar septa of lungs from pigs which received the PRRSV vaccine followed by PRRSV challenge (group 3).
Table 2.
Histopathologic lesions and antigen levels for PCV2 and PRRSV in different groups at 21 dpca
| Group no. | Histopathology score |
PCV2 antigen level |
PRRSV antigen level |
|||
|---|---|---|---|---|---|---|
| Lung | Lymph node | Lung | Lymph node | Lung | Lymph node | |
| 1 | 0.5 ± 0.53b | 0.37 ± 0.51c | 7 ± 5.09b | 17 ± 6.05c | ||
| 2 | 2.12 ± 0.64a | 0.62 ± 0.74c | 6.5 ± 5.44b | 20 ± 6.87c | 26.5 ± 5.44b | 4.5 ± 2.03 |
| 3 | 0.87 ± 0.64b | 0.5 ± 0.53c | 14.75 ± 4.03c | 5.5 ± 1.29 | ||
| 4 | 2.75 ± 0.7a | 3.87 ± 0.83a | 44 ± 5.29a | 61 ± 8.2a | 40.5 ± 3.69a | 6.5 ± 2.08 |
| 5 | 0.4 ± 0.23b | 0.45 ± 0.42c | 8 ± 6.91b | 22 ± 10.28c | 16.5 ± 4.38c | 4 ± 2.63 |
| 6 | 0.75 ± 0.7b | 2.12 ± 0.64b | 13.5 ± 5b | 43.5 ± 8.69b | ||
| 7 | 2.25 ± 0.7a | 0.5 ± 0.53c | 27.75 ± 3.77b | 8.5 ± 2.64 | ||
| 8 | 2.87 ± 0.64a | 4.25 ± 0.7a | 53 ± 4.24a | 67.5 ± 8.38a | 41.75 ± 5.9a | 7.5 ± 2.64 |
| 9 | 0.25 ± 0.46b | 0.25 ± 0.46c | ||||
Data are means ± standard deviations. The antigen level in the lung means the number of positive cells per unit area (0.95 mm2). The antigen level in the lymph node means the number of positive cells per unit area (0.25 mm2). Different letters (a, b, and c) indicate significant (P < 0.05) differences between groups.
Immunohistochemical scores.
Pigs which received the PRRSV vaccine followed by dual challenge (group 4) (Fig. 4A) and pigs which were challenged with both PCV2 and PRRSV (group 8) had significantly more PCV2-positive cells per unit area in the lymph node and lung than pigs which received the PCV2 vaccine followed by PCV2 challenge only (group 1), pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5), and pigs which were challenged with PCV2 only (group 6) (Fig. 4B) (P < 0.05). Pigs in group 6 had significantly more PCV2-positive cells per unit area in the lymph node and lung than pigs in groups 1, 2, and 5 (P < 0.05).
Pigs which received the PRRSV vaccine followed by dual challenge (group 4) and pigs which were challenged with both PCV2 and PRRSV (group 8) had significantly more PRRSV-positive cells per unit area in the lung than pigs which received the PCV2 vaccine followed by dual challenge (group 2), pigs which received the PRRSV vaccine followed by PRRSV challenge only (group 3) (Fig. 5B), pigs which received both the PCV2 and PRRSV vaccines followed by dual challenge (group 5), and pigs which were challenged with PRRSV only (group 7) (P < 0.02).
DISCUSSION
The goal of the present study was to determine the effects of PCV2 and PRRSV vaccinations in pigs that were challenged with PCV2, PRRSV, or both. Our study focused on the PRRSV vaccine-PCV2-PRRSV and PCV2 vaccine-PCV2-PRRSV interactions. With the single-challenge model, the PCV2 and PRRSV vaccines reduced PCV2 and PRRSV viremia, respectively, in pigs. However, it was surprising that the reduction of those viruses in the blood by each of the vaccines was different in the dual-challenge model than in the single-challenge model. Theoretically, one way to minimize the effect of the PRRSV-associated enhancement of the replication of PCV2 and the induction of PMWS may be the PRRSV-based vaccination of preweaned pigs in PCV2-PRRSV-coinfected herds. Interestingly, pigs which received the PRRSV vaccine followed by dual challenge showed significantly increased PCV2 viremia, PCV2-induced lesions, and PCV2 antigens compared to pigs challenged with PCV2 only but did not significantly differ from pigs challenged with both PCV2 and PRRSV. Hence, the PRRSV vaccine-PCV2-PRRSV model suggests that the PRRSV vaccine and/or PRRSV itself could potentially result in the enhancement of PCV2 replication rather than in its control.
Pigs have a prominent CD4+ CD8+ double-positive cell population (reportedly up to 60% of total lymphocytes) in the peripheral blood (29). These CD4+ CD8+ double-positive cells are memory and effector cells and have the capacity to produce large amounts of IFN-γ. Hence, these double-positive cells might play a role in protective immunity against viral diseases (30). Although depletion of CD4+ CD8+ double-positive cells by PCV2 was not demonstrated directly, enhancement of PCV2 replication may impair the immune system. Once the replication of PCV2 reached high levels, the numbers of CD4+ CD8+ double-positive cells and PRRSV-specific IFN-γ-SCs were significantly reduced at 10 dpc in pigs which received the PRRSV vaccine followed by dual challenge. Decreased numbers of CD4+ CD8+ double-positive cells and PRRSV-specific IFN-γ-SCs are significant because PRRSV-specific IFN-γ-SCs are predominantly CD4+ CD8+ double-positive cells (31) and play key roles in protective cell-mediated immunity against PRRSV infection (16, 32).
In the present study, pigs which received the PRRSV vaccine followed by dual challenge showed significantly increased PRRSV viremia, PRRSV-associated lesions, and PRRSV antigens compared to pigs challenged with PRRSV only. The impairment of the protective cell-mediated immunity against PRRSV by PCV2 in the PRRSV vaccine-PCV2-PRRSV model may have allowed for increases in PRRSV viremia, PRRSV-associated lesions, and PRRSV antigens rather than their reduction following the PRRSV vaccine. Because these 3 parameters were significantly reduced in pigs which received the PRRSV vaccine followed by PRRSV challenge only compared with pigs challenged with PRRSV only, our results ruled out its possible limited efficacy against a heterologous virus. Regarding the efficacy, it is clear that the modified live PRRS vaccine offers protection against reinfection with homologous and, in some cases, heterologous viruses (33).
Regardless of the PRRSV challenge, PCV2 vaccination induces high levels of NAs and PCV2-specific IFN-γ-SCs, which are important protective immune parameters for the control of PCV2 infection (31, 34–36). The induction of protective immunity significantly reduced PCV2 viremia, PCV2-induced lesions, and PCV2 antigens in both pigs which received the PCV2 vaccine followed by PCV2 challenge only and pigs challenged with both PCV2 and PRRSV compared with pigs challenged with PCV2 only. Our results suggest that PCV2 vaccination is able to reduce the levels of PCV2 viremia in pigs regardless of the PRRSV infection status, as reported in a previous study (9). Our results may explain why the PCV2 vaccine is effective under field conditions where PCV2 and PRRSV are widespread in the pig population, and accordingly, it is likely that most herds in which PCV2 vaccines are being used are infected with PRRSV (37). In contrast, the levels of PRRSV viremia were significantly higher in pigs which received the PRRSV vaccine followed by dual challenge than in pigs which were challenged with PRRSV only. These results suggest that the reduction of PRRSV viremia by the PRRSV vaccine is affected by the pig's PCV2 infection status. Our results agree with those of previous similar studies, in which PRRSV infection at the time of PCV2 vaccination had no impact on the efficacy of the vaccine but PCV2 infection at the time of PRRSV vaccination did affect the efficacy (38, 39). Our results are clinically meaningful, indicating that swine practitioners should check PCV2 infection statuses in pigs before PRRSV vaccines are used to control PRRSV infection in swine herds. Therefore, the control of PCV2 infections by the PCV2 vaccine may positively affect the full induction of immunity by the PRRSV vaccine in herds that are suffering from a coinfection with PCV2 and PRRSV.
In summary, vaccination against PCV2 reduced PCV2 viremia, PCV2-induced lesions, and PCV2 antigens in dually infected pigs. However, vaccination against PRRSV increased PCV2 viremia, PCV2-induced lesions, and PCV2 antigens in dually infected pigs. Therefore, the PCV2 vaccine decreased the potentiation of PCV2-induced lesions by PRRSV in dually infected pigs. In contrast, the PRRSV vaccine alone did not decrease the potentiation of PCV2-induced lesions by PRRSV in dually infected pigs.
ACKNOWLEDGMENTS
This research was supported by contract research funds from the Research Institute for Veterinary Science (RIVS) of the College of Veterinary Medicine and by the Brain Korea 21 Program for Veterinary Science in the Republic of Korea.
Footnotes
Published ahead of print 9 January 2013
REFERENCES
- 1.Chae C. 2004. Postweaning multisystemic wasting syndrome: a review of aetiology, diagnosis and pathology. Vet. J. 168:41–49 [DOI] [PubMed] [Google Scholar]
- 2.Chae C. 2005. A review of porcine circovirus 2-associated syndromes and diseases. Vet. J. 169:326–336 [DOI] [PubMed] [Google Scholar]
- 3.Cavanagh D. 1997. Nidovirales: a new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 142:629–633 [PubMed] [Google Scholar]
- 4.Zimmerman JJ, Benfield DA, Dee SA, Murtaugh MP, Stadejek T, Stevenson GW, Torremorell M. 2012. Porcine reproductive and respiratory syndrome virus (porcine arterivirus), p 461–486. In Zimmerman JJ, Karriker LA, Ramirez A, Schwartz KJ, Stevenson GW. (ed), Diseases of swine, 10th ed Wiley-Blackwell, West Sussex, United Kingdom [Google Scholar]
- 5.Kim J, Chung HK, Chae C. 2003. Association of porcine circovirus 2 with porcine respiratory disease complex. Vet. J. 166:251–256 [DOI] [PubMed] [Google Scholar]
- 6.Kim J, Chung HK, Jung T, Cho WS, Choi C, Chae C. 2002. Postweaning multisystemic wasting syndrome of pigs in Korea: prevalence, microscopic lesions and coexisting microorganisms. J. Vet. Med. Sci. 64:57–62 [DOI] [PubMed] [Google Scholar]
- 7.Pallarés FJ, Halbur PG, Opriessnig T, Sorden SD, Villar D, Janke BH, Yaeger MJ, Larson DJ, Schwartz KJ, Yoon KJ, Hoffman LJ. 2002. Porcine circovirus type 2 (PCV-2) coinfections in US field cases of postweaning multisystemic wasting syndrome (PMWS). J. Vet. Diagn. Invest. 14:515–519 [DOI] [PubMed] [Google Scholar]
- 8.Rovira A, Balasch M, Segalés J, García L, Plana-Durán J, Rosell C, Ellerbrok H, Mankertz A, Domingo M. 2002. Experimental inoculation of conventional pigs with porcine reproductive and respiratory syndrome virus and porcine circovirus 2. J. Virol. 76:3232–3239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Opriessnig T, Madson DM, Prickett JR, Kuhar D, Lunney JK, Elsener J, Halbur PG. 2008. Effect of porcine circovirus type 2 (PCV2) vaccination on porcine reproductive and respiratory syndrome virus (PRRSV) and PCV2 coinfection. Vet. Microbiol. 131:103–114 [DOI] [PubMed] [Google Scholar]
- 10.Allan GM, McNeilly F, Ellis J, Krakowka S, Meehan B, McNair I, Walker I, Kennedy S. 2000. Experimental infection of colostrum deprived piglets with porcine circovirus 2 (PCV2) and porcine reproductive and respiratory syndrome virus (PRRSV) potentiates PCV2 replication. Arch. Virol. 145:2421–2429 [DOI] [PubMed] [Google Scholar]
- 11.Harms PA, Sorden SD, Halbur PG, Bolin SR, Larger KM, Morozov I, Paul PS. 2001. Experimental reproduction of severe disease in CD/CD pigs concurrently infected with type 2 porcine circovirus and porcine reproductive and respiratory syndrome virus. Vet. Pathol. 38:528–539 [DOI] [PubMed] [Google Scholar]
- 12.Fenaux M, Opriessnig T, Halbur PG, Elvinger F, Meng XJ. 2004. A chimeric porcine circovirus (PCV) with the immunogenic capsid gene of the pathogenic PCV type 2 (PCV2) cloned into the genomic backbone of the nonpathogenic PCV1 induces protective immunity against PCV2 infection in pigs. J. Virol. 78:6297–6303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gagnon CA, del Castillo JR, Music N, Fontaine G, Harel J, Tremblay D. 2008. Development and use of a multiplex real-time quantitative polymerase chain reaction assay for detection and differentiation of Porcine circovirus-2 genotypes 2a and 2b in an epidemiological survey. J. Vet. Diagn. Invest. 20:545–558 [DOI] [PubMed] [Google Scholar]
- 14.Wasilk A, Callahan JD, Christopher-Hennings J, Gay TA, Fang Y, Dammen M, Reos ME, Torremorell M, Polson D, Mellencamp M, Nelson E, Nelson WM. 2004. Detection of U.S., Lelystad, and European-like porcine reproductive and respiratory syndrome viruses and relative quantitation in boar semen and serum samples by real-time PCR. J. Clin. Microbiol. 42:4453–4461 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pogranichnyy RM, Yoon KJ, Harms PA, Swenson SL, Zimmerman JJ, Sorden SD. 2000. Characterization of immune response of young pigs to porcine circovirus type 2 infection. Viral Immunol. 13:143–153 [DOI] [PubMed] [Google Scholar]
- 16.Zuckermann FA, Garcia EA, Luque ID, Christopher-Hennings J, Doster A, Brito M, Osorio F. 2007. Assessment of the efficacy of commercial porcine reproductive and respiratory syndrome virus (PRRSV) vaccines based on measurement of serologic response, frequency of gamma-IFN-producing cells and virological parameters of protection upon challenge. Vet. Microbiol. 123:69–85 [DOI] [PubMed] [Google Scholar]
- 17.Han K, Seo HW, Shin JH, Oh Y, Kang I, Park C, Chae C. 2011. Effect of the modified live porcine reproductive and respiratory syndrome virus (PRRSV) vaccine on European and North American PRRSV shedding in semen from infected boars. Clin. Vaccine Immunol. 18:1600–1607 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bautista EM, Molitor TW. 1997. Cell-mediated immunity to porcine reproductive and respiratory syndrome virus in swine. Viral Immunol. 10:83–94 [DOI] [PubMed] [Google Scholar]
- 19.Rodríguez-Arrioja GM, Segalés J, Balasch M, Rosell C, Quintant J, Folch JM, Plana-Durán J, Mankertz A, Domingo M. 2000. Serum antibodies to porcine circovirus type 1 and type 2 in pigs with and without PMWS. Vet. Rec. 146:762–764 [DOI] [PubMed] [Google Scholar]
- 20.Diaz I, Mateu E. 2005. Use of ELISPOT and ELISA to evaluate IFN-γ, IL-10 and IL-4 responses in conventional pigs. Vet. Immunol. Immunopathol. 106:107–112 [DOI] [PubMed] [Google Scholar]
- 21.Oh Y, Seo HW, Han K, Park C, Chae C. 2012. Protective effect of the maternally derived porcine circovirus type 2 (PCV2)-specific cellular immune response in piglets by dam vaccination against PCV2 challenge. J. Gen. Virol. 93:1556–1562 [DOI] [PubMed] [Google Scholar]
- 22.Kim J, Choi C, Chae C. 2003. Pathogenesis of postweaning multisystemic wasting syndrome reproduced by co-infection with Korean isolates of porcine circovirus 2 and porcine parvovirus. J. Comp. Pathol. 128:52–59 [DOI] [PubMed] [Google Scholar]
- 23.West KH, Bystrom JM, Wojnarowicz C, Shantz N, Jacobson M, Allan GM, Haines DM, Clark EG, Krakowka S, McNeilly F, Konoby C, Martin K, Ellis JA. 1999. Myocarditis and abortion associated with intrauterine infection of sows with porcine circovirus 2. J. Vet. Diagn. Invest. 11:530–532 [DOI] [PubMed] [Google Scholar]
- 24.Han K, Seo HW, Oh Y, Kang I, Park C, Kang SH, Lee BH, Kwon B, Chae C. 2012. Evaluation of monoclonal antibody-based immunohistochemistry for the detection of European and North American porcine reproductive and respiratory syndrome virus and a comparison with in situ hybridization and reverse transcription polymerase chain reaction. J. Vet. Diagn. Invest. 24:719–724 [DOI] [PubMed] [Google Scholar]
- 25.Kim J, Chae C. 2004. A comparison of virus isolation, polymerase chain reaction, immunohistochemistry, and in situ hybridization for the detection of porcine circovirus 2 and porcine parvovirus in experimentally and naturally coinfected pigs. J. Vet. Diagn. Invest. 16:45–50 [DOI] [PubMed] [Google Scholar]
- 26.Halbur PG, Paul PS, Frey ML, Landgraf J, Eernisse K, Meng XJ, Lum MA, Andrews JJ, Rathje JA. 1995. Comparison of the pathogenicity of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet. Pathol. 32:648–660 [DOI] [PubMed] [Google Scholar]
- 27.Kim J, Chae C. 2004. Expression of monocyte chemoattractant protein-1 and macrophage inflammatory protein-1 in porcine circovirus 2-induced granulomatous inflammation. J. Comp. Pathol. 131:121–126 [DOI] [PubMed] [Google Scholar]
- 28.Halbur PG, Paul PS, Frey ML, Landgraf J, Eernisse K, Meng XJ, Andrews JJ, Lum MA, Rathje JA. 1996. Comparison of the antigen distribution of two US porcine reproductive and respiratory syndrome virus isolates with that of the Lelystad virus. Vet. Pathol. 33:159–170 [DOI] [PubMed] [Google Scholar]
- 29.Saalmuller A, Redehasse MJ, Buhring Jonjic H-JS, Koszinowski UH. 1987. Simultaneous expression of CD4 and CD8 antigens by a substantial proportion of resting porcine T lymphocytes. Eur. J. Immunol. 17:1297–1301 [DOI] [PubMed] [Google Scholar]
- 30.Rodriques-Carreno MP, López-Fuertes L, Revilla C, Ezquerra A, Alonso F, Domínguez J. 2002. Phenotypic characterization of porcine IFN-γ-producing lymphocytes by flow cytometry. J. Immunol. Methods 259:171–179 [DOI] [PubMed] [Google Scholar]
- 31.Mateu E, Diaz I. 2008. The challenge of PRRS immunology. Vet. J. 177:345–351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Meier WA, Galeota J, Osorio FA, Husmann RJ, Schnitzlein WM, Zuckermann FA. 2003. Gradual development of the interferon-γ response of swine to porcine reproductive and respiratory syndrome virus infection or vaccination. Virology 309:18–31 [DOI] [PubMed] [Google Scholar]
- 33.Lunney JK, Benfield DA, Rowland RRR. 2010. Porcine reproductive and respiratory syndrome virus: an update on an emerging and re-emerging viral disease of swine. Virus Res. 154:1–6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chae C. 2012. Commercial porcine circovirus type 2 vaccine: efficacy and clinical application. Vet. J. 194:151–157 [DOI] [PubMed] [Google Scholar]
- 35.Fort M, Sibila M, Perez-Martin E, Nofrarias M, Mateu E, Segalés J. 2009. One dose of a porcine circovirus 2 (PCV2) sub-unit vaccine administered to 3-week-old conventional piglets elicits cell-mediated immunity and significantly reduces PCV2 viremia in an experimental model. Vaccine 27:4031–4037 [DOI] [PubMed] [Google Scholar]
- 36.Opriessnig T, Patterson AR, Madson DM, Pal N, Halbur PG. 2009. Comparison of efficacy of commercial one dose and two dose PCV2 vaccines using a mixed PRRSV-PCV2-SIV clinical infection model 2-3-months post vaccination. Vaccine 27:1002–1007 [DOI] [PubMed] [Google Scholar]
- 37.Fachinger V, Bischoff R, Jedidia SB, Saalmuller A, Elbers K. 2008. The effect of vaccination against porcine circovirus type 2 in pigs suffering from porcine respiratory disease complex. Vaccine 26:1488–1499 [DOI] [PubMed] [Google Scholar]
- 38.Opriessnig T, McKeown NE, Harmon KL, Meng XJ, Halbur PG. 2006. Porcine circovirus type 2 infection decreases the efficacy of a modified live porcine reproductive and respiratory syndrome virus vaccine. Clin. Vaccine Immunol. 13:923–929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sinha A, Shen HG, Schalk S, Beach NM, Huang YW, Halbur PG, Meng XJ, Opriessnig T. 2010. Porcine reproductive and respiratory syndrome virus infection at the time of porcine circovirus type 2 vaccination has no impact on vaccine efficacy. Clin. Vaccine Immunol. 17:1940–1945 [DOI] [PMC free article] [PubMed] [Google Scholar]