

Abstract
In this work we describe a series of simple protocols using planaria as an animal model in toxicology and behavioral pharmacology. These procedures have proven useful to provide significant research experience to undergraduate students, including coauthorship in peer-reviewed publications. The methods described in this work have proven useful to allow students to visualize concepts related to concentration-effect curves for toxicity and behavioral experiments, without the need to consider factors that must be taken into account when working with vertebrate animals.
Keywords: planaria, motility, behavior, toxicology, pharmacology, DMSO, caffeine
Planarian flatworms (Figure 1) are one of the best-characterized animal models in developmental biology and regeneration research (Newmark and Sánchez Alvarado, 2002), and are currently being rediscovered as a very useful animal model in neuropharmacology (Buttarelli et al., 2008). Planarians are the first example of organisms displaying cephalization, including a primitive “brain,” with many features common to vertebrate nervous systems, such as multipolar neurons and dendritic spines (Sarnat and Netsky, 1985, 2002). In fact, the planarian nervous system is more similar to vertebrate nervous systems than to invertebrate nervous systems (like insects, for example), in terms of cell morphology and physiology (Sarnat and Netsky, 1985, 2002). Additionally, nearly every neurotransmitter found in mammals is present in planarians (Buttarelli et al., 2000, 2008; Ribeiro et al., 2005). Planarians are also extremely useful to study nervous system development and regeneration (Cebrià, 2007; Agata and Umesono, 2008).
Figure 1.

Planarian worm (Dugesia sp.). Courtesy of Dr. Richard Woodruff, Department of Biology, West Chester University.
Working with this organism has several distinct advantages. They are commercially available at very low cost and are very easily kept in the laboratory. No special equipment such as incubators, etc., is necessary. Additionally, these worms display specific behavioral responses to psychoactive substances, such as cocaine, cannabinoids, amphetamines and opiates (Buttarelli et al., 2002; Kusayama and Watanabe, 2000; Pagán et al., 2008; Palladini et al., 1996; Raffa and Desai, 2005). Planarians have also been used in chemical and behavioral toxicology studies (Best and Morita, 1991; Grebe and Schaeffer 1991; Kapu and Schaeffer, 1991; Li, 2008) and as tools for the identification of chemical teratogens (Sabourin et al., 1985; Schaeffer, 1993). Many of these substances are hydrophobic, needing solubility-aiding agents for them to be dissolved in aqueous solutions. One example of such substances is dimethylsulfoxide (DMSO, Balakin et al., 2006).
In vertebrates, DMSO can induce analgesic- and anesthetic-like effects (Evans et al., 1993; Castroman and Ness, 2002). DMSO display toxic and behavioral effects in planarian worms as well (Pagán et al., 2006). The characterization of DMSO effects by itself is an essential control in experiments using this substance as a solubility-enhancer.
Caffeine is a well-known adenosine receptor antagonist, which is widely used to increase alertness in humans, and displays many other physiological effects (Yun et al., 2007). In planaria, low doses of caffeine during short periods increases oxygen consumption, which is consistent with its metabolic effects in vertebrates (Hinrichs, 1924).
Here we describe a set of simple procedures using the planarian worm as an animal model in toxicology and behavioral pharmacology, using DMSO and caffeine as our experimental compounds. The methods described here are useful to visualize concentration-effect curves for toxicity/lethality and behavioral observations without the usual considerations needed when working with vertebrate animals. Additionally, the protocols outlined here can be carried out at very low cost, using common laboratory materials and supplies. Planarians are commercially available through various companies or can be directly harvested from ponds.
The application of these methods in our laboratory has provided significant research experience to undergraduate students; these experiences have resulted in student presentations at internal and national scientific meetings, as well as coauthorship in peer-reviewed research publications (Pagán et al., 2006, 2008; Rowlands and Pagán, 2008).
MATERIALS AND METHODS
General laboratory materials and reagents were purchased from Fisher Scientific (Suwanee, GA) or Sigma- Aldrich (St. Louis, MO). In this work, we will use dimethylsulfoxide (DMSO) and caffeine as experimental compounds. The data was analyzed using the PRISM software package (GraphPad, San Diego, CA). The methods describe here are modifications of published procedures (Raffa et al., 2001, as modified in Pagán et al., 2006, 2008). The planarians used in this work were purchased form Ward’s (Rochester, NY), but can be obtained through a variety of biological suppliers.
TOXICOLOGY - MATERIALS
Planarian worms (1–2 cm long; Dugesia sp.)
Polypropylene dishes (100 × 15 mm) – 4 compartments
Small paint brush
Dissecting microscope
Artificial pond water (APW, NaCl, 6 mM; NaHCO3, 0.1 mM; CaCl2, 0.6 mM; pH 6.9).
Experimental compounds in APW, as indicated
TOXICOLOGY - METHODS
Label the lid of a four-compartment dish (Figure 2A) with the appropriate solutions that it will contain. Always use one of the compartments as a control (APW only).
Using a small paintbrush, transfer (gently) 6–7 planarian worms to each compartment.
After transferring the worms, add 4 mL of the appropriate solution (APW or the different DMSO concentration).
Using the dissecting microscope, observe the worms for a period of 30 minutes, recording your observations (% of live worms) in Table 1. Death in planarians is a rather unambiguous process; the worms begin to disintegrate, usually head first (Behre, 1918; Pagán et al., 2006).
Plot the data recorded in Table 1 as % of live wormsvs.DMSO % at the different time periods.
- Fit these data points to an empirical Hill equation (Equation 1) in the form:
(Equation 1)
where F is the fraction of control for each DMSO concentration, n is the Hill coefficient, C is the experimental compound concentration and E50 is the compound concentration that induces 50 % of the observed effect. An empirical equation such as the one shown above is the best initial treatment of the data, since we have no information about the possible mechanism of action of the experimental compounds in planaria. The Hill Coefficient can indicate cooperativity between two or more binding sites. Generally, n’s significantly higher or lower than unity indicate positive or negative cooperativity, respectively, provided that the putative binding sites differ on their affinity for a specific ligand. Two different binding sites with similar affinities for the ligand would be indistinguishable with this equation. Even in binding studies, however, the Hill equation provides little or no information about the specific mechanism (Weiss, 1997). This is primarily a behavioral work; in a whole organism there will be multiple binding sites, which will interact to induce specific behaviors. If desired, the data can be fit to other equations according to specific experimental designs.
Figure 2.
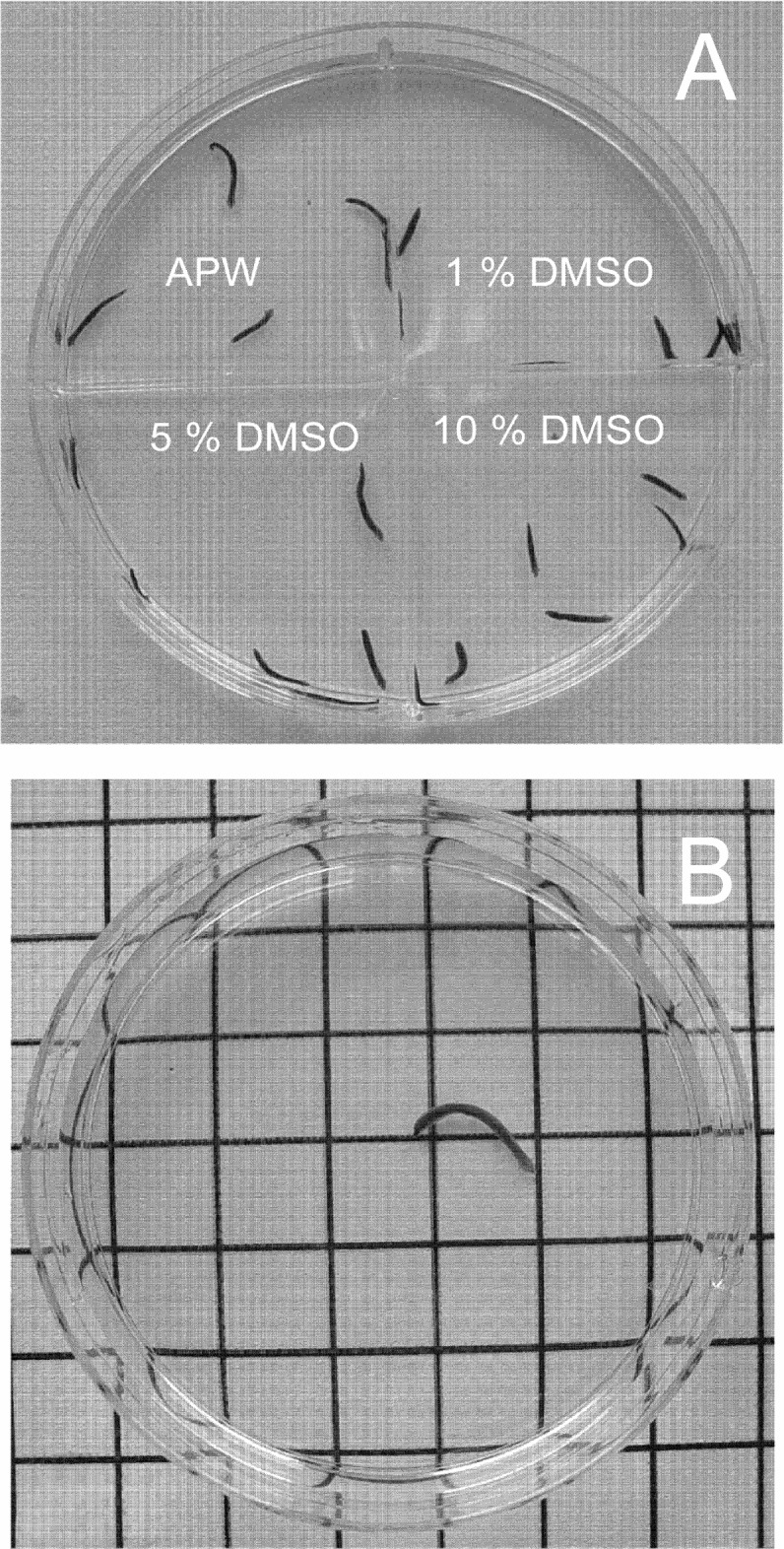
A. Toxicity experiments setup. B. Motility experiments setup.
Table 1.
Planaria survival in the presence of DMSO at the indicated incubation times. For examples of the data plot from this table please see Figure 3A.
| Incubation time (minutes) | APW Fraction alive | DMSO 0.1 % Fraction alive | DMSO 1 % Fraction alive | Etc. |
|---|---|---|---|---|
| 0 | ||||
| 3 | ||||
| 5 | ||||
| 10 | ||||
| 15 | ||||
| 20 | ||||
| 30 |
BEHAVIOR - MATERIALS
Planarian worms (1–2 cm long; Dugesia sp.)
6 cm polypropylene dishes on a 1 cm2 gridline (Figure 2B)
Small paint brush
Artificial pond water (APW, NaCl, 6 mM; NaHCO3, 0.1 mM; CaCl2, 0.6 mM; pH 6.9).
Experimental compounds in APW, as indicated
BEHAVIOR – METHODS
Using a small paintbrush, gently transfer one planarian worm (about 1–2 cm long) to one 6 cm plastic dish and set it on a piece of graph paper with grid lines placed at 1 cm intervals (Figure 4).
Add 5 mL of APW or the experimental solutions.
As the worm glides around, count how many times it crosses or recrosses a line in one minute over a period of eight minutes. One cross is defined as the point where the worm’s head touches a gridline.
Record your observations as outlined in Table 2.
Plot your cumulative crosses as a function of time. Fit your data to a linear regression model.
Normalize the experimental slopes obtained from the fit above to control slopes.
Plot your normalized data as the fraction of controlvs.the experimental compound concentration and fit it to Equation 1.
Figure 4.

A. Representative linear plots showing planarian motility decrease by two DMSO concentrations. The lines were generated by fitting the data to a linear equation. B. The experimental slopes were normalized to control slopes (APW) and graphed as a function of the DMSO concentration. The line was generated by fitting the data to Equation 1 (see methods). Each curve represents a minimum of three experiments. Error bars represent the standard error of the mean.
Table 2.
Gridlines crossed per minute and cumulative crosses (See Methods). For examples of the data plot from this table please see Figures 4A and 5A.
| Time (minutes) | Crosses | Cumulative Crosses |
|---|---|---|
| 1 | ||
| 2 | ||
| 3 | ||
| 4 | ||
| 5 | ||
| 6 | ||
| 7 | ||
| 8 |
RESULTS and DISCUSSION
Figure 3A shows a representative graph of the effect of DMSO on planarian survival at different time periods. The data demonstrates that DMSO lethality is dependent on both the DMSO concentration and the time of exposure (Figure 3B). These results are consistent with published reports (Pagán, et al., 2006). Figure 4A shows representative plots of cumulative crosses as a function of time under control conditions and at two DMSO concentrations, indicating that the DMSO-induced motility decrease is concentration-dependent. Figure 4B shows a concentration-response curve for this motility decrease. The E50 value was close to 1 %, in agreement with previous reports (Pagán, et al., 2006). Figure 5 shows the caffeine-induced, concentration-dependent increase in planarian motility. This behavioral effect is consistent with known stimulant effects of caffeine in vertebrates (Nehlig et al., 1992).
Figure 3.

A. DMSO toxicity to planarians as a function of concentration at the following time periods: 1 minute (closed circles), 8 minutes (open circles), 15 minutes (closed squares) and 30 minutes (open squares). The lines were generated by fitting the data to Equation 1 (see methods). Inset: DMSO structure. B. The E50 values are inversely proportional to the time of exposure. Each curve represents a minimum of three experiments. Error bars represent the standard error of the mean.
Figure 5.

A. Linear plots showing examples of caffeine-induced planarian motility increase. The lines were generated by fitting the data to a linear equation. Inset: caffeine structure B. The experimental slopes were normalized to control slopes (APW) and graphed as shown, indicating the caffeine-induced increase in planarian motility. Each line/bar represents a minimum of two experiments. Error bars represent the standard error of the mean.
Planarian worms are being rediscovered as useful organisms in neurobiology and pharmacology. The experimental designs outlined here can be easily adapted to a wide variety of experimental compounds. The similarity of the planarian nervous system to vertebrate nervous systems (Sarnat and Netsky, 1985, 2002; Buttarelli et al., 2008) allows for the observation of behavioral responses in planarians comparable to vertebrate behavioral responses.
What will the students learn using these protocols? In many cases, undergraduate practical experience in behavioral pharmacology and toxicology is limited, since experiments using vertebrates tend to be more expensive in terms of animal housing, materials, disposal, etc. Additionally, vertebrate experimental protocols require more preparation in terms of permissions and ethical concerns. For example, it is questionable and impractical to sacrifice hundreds of vertebrate animals in experimental procedures showing lethality of toxic compounds solely for demonstration or training purposes. Planarians can be used to construct lethality curves (Figure 3A), as well as other types of data analyses (Figures 3, 4 and 5). The main advantage of using planarians as opposed to other invertebrates was discussed in the introduction of this paper.
If desired, assessment of the learning outcomes can be incorporated in the design of the laboratory work. Overall, the presented experiments show the potential of planarian worms as a low-cost experimental organism in neuroscience education in general and in pharmacology and toxicology education in particular.
Acknowledgments
Dr. Oné R. Pagán wishes to dedicate this paper to the memory of his father, Mr. Onésimo Pagán, who passed away on July 8, 2009. He was a wonderful dad. The authors wish to thank Dr. Maureen Knabb, Department of Biology, West Chester University (WCU), for careful reading of this manuscript and for useful suggestions. The financial support from the Department of Biology, the College of Arts & Sciences and the Office of Sponsored Research, WCU, in the form of departmental funds, two CASSDA Awards and a Faculty Development Grant (To O.R.P.) is gratefully acknowledged. Additional financial support was provided by the SOMAS program (Support for Mentors and their Students in the Neurosciences, NSF-DUE-0426266, to O.R.P. via Dr. Julio Ramirez, Davidson College). Ms. Coudron and Ms. Kaneria are WCU Psychology/Biology and Biology Majors respectively.
REFERENCES
- Agata K, Umesono Y. Brain regeneration from pluripotent stem cells in planarian. Philos Trans R Soc Lond B Biol Sci. 2008;363:2071–2078. doi: 10.1098/rstb.2008.2260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balakin KV, Saychuk NP, Tetko IV. In silico approaches to prediction of aqueous and DMSO solubility of drug-like compounds: trends, problems and solutions. Curr Med Chem. 2006;13:223–241. doi: 10.2174/092986706775197917. [DOI] [PubMed] [Google Scholar]
- Behre EH. An Experimental Study of Acclimation to Temperature in Planaria dorotocephala. Biol Bull. 1918;35:277–317. [Google Scholar]
- Best JB, Morita M. Toxicology of planarians. Hydrobiologia. 1991;227:375–383. [Google Scholar]
- Buttarelli FR, Pellicano C, Pontieri FE. Neuropharmacology and behavior in planarians: translations to mammals. Comp Biochem Physiol C Toxicol Pharmacol. 2008;147:399–408. doi: 10.1016/j.cbpc.2008.01.009. [DOI] [PubMed] [Google Scholar]
- Buttarelli FR, Pontieri FE, Margotta V, Palladini G. Cannabinoid-induced stimulation of motor activity in planaria through an opioid receptor-mediated mechanism. Prog Neuropsychopharmacol Biol Psychiatry. 2002;26:65–68. doi: 10.1016/s0278-5846(01)00230-5. [DOI] [PubMed] [Google Scholar]
- Buttarelli FR, Pontieri FE, Margotta V, Palladini G. Acetylcholine/dopamine interaction in planaria. Comp Biochem Physiol C Toxical Pharmacol. 2000;125:225–231. doi: 10.1016/s0742-8413(99)00111-5. [DOI] [PubMed] [Google Scholar]
- Castroman PJ, Ness TJ. Spinal neurophysiologic correlates of the analgesic actions of intravesical dimethyl sulfoxide and capsaicin in the rat. J Pain. 2002;3:394–400. doi: 10.1054/jpai.2002.126789. [DOI] [PubMed] [Google Scholar]
- Cebrià F. Regenerating the central nervous system: how easy for planarians! Dev Genes Evol. 2007;217:733–748. doi: 10.1007/s00427-007-0188-6. [DOI] [PubMed] [Google Scholar]
- Evans MS, Reid KH, Sharp JB. Dimethylsulfoxide (DMSO) blocks conduction in peripheral nerve C fibers: a possible mechanism of analgesia. Neurosci Lett. 1993;150:145–148. doi: 10.1016/0304-3940(93)90522-m. [DOI] [PubMed] [Google Scholar]
- Grebe E, Schaeffer DJ. Planarians in toxicology, standardization of a rapid neurobehavioral toxicity test using phenol in a crossover study. Bull Environ Contam Toxicol. 1991;46:866–870. doi: 10.1007/BF01689731. [DOI] [PubMed] [Google Scholar]
- Hinrichs MA. A study of the physiological effects of caffein upon Planaria dorotocephala. J Exp Zool. 1924;40:271–300. [Google Scholar]
- Kapu MM, Schaeffer DJ. Planarians in toxicology. Responses of asexual Dugesia dorotocephala to selected metals. Bull Environ Contam Toxicol. 1991;47:302–307. doi: 10.1007/BF01688656. [DOI] [PubMed] [Google Scholar]
- Kusayama T, Watanabe S. Reinforcing effects of methamphetamine in planarians. Neuroreport. 2000;11:2511–2513. doi: 10.1097/00001756-200008030-00033. [DOI] [PubMed] [Google Scholar]
- Li MH. Effects of nonionic and ionic surfactants on survival, oxidative stress, and cholinesterase activity of planarian. Chemosphere. 2008;70:1796–1803. doi: 10.1016/j.chemosphere.2007.08.032. [DOI] [PubMed] [Google Scholar]
- Nehlig A, Daval JL, Debry G. Caffeine and the central nervous system: mechanisms of action, biochemical, metabolic and psychostimulant effects. Brain Res Brain Res Rev. 1992;17:139–170. doi: 10.1016/0165-0173(92)90012-b. [DOI] [PubMed] [Google Scholar]
- Newmark PA, Sánchez Alvarado A. Not your father’s planarian: a classic model enters the era of functional genomics. Nat Rev Genet. 2002;3:210–219. doi: 10.1038/nrg759. [DOI] [PubMed] [Google Scholar]
- Pagán OR, Rowlands AL, Azam M, Urban KR, Bidja AH, Roy DM, Feeney RB, Afshari LK. Reversal of cocaine-induced planarian behavior by parthenolide and related sesquiterpene lactones. Pharmacol Biochem Behav. 2008;89:160–170. doi: 10.1016/j.pbb.2007.12.008. [DOI] [PubMed] [Google Scholar]
- Pagán OR, Rowlands AL, Urban KR. Toxicity and behavioral effects of dimethylsulfoxide in planaria. Neurosci Lett. 2006;407:274–278. doi: 10.1016/j.neulet.2006.08.073. [DOI] [PubMed] [Google Scholar]
- Palladini G, Ruggeri S, Stocchi F, De Pandis MF, Venturini G, Margotta V. A pharmacological study of cocaine activity in planaria. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. 1996;115:41–45. doi: 10.1016/s0742-8413(96)00053-9. [DOI] [PubMed] [Google Scholar]
- Raffa RB, Desai P. Description and quantification of cocaine withdrawal signs in Planaria. Brain Res. 2005;1032:200–202. doi: 10.1016/j.brainres.2004.10.052. [DOI] [PubMed] [Google Scholar]
- Raffa RB, Holland LJ, Schulingkamp RJ. Quantitative assessment of dopamine D2 antagonist activity using invertebrate (Planaria) locomotion as a functional endpoint. J Pharmacol Toxicol Methods. 2001;45:223–226. doi: 10.1016/s1056-8719(01)00152-6. [DOI] [PubMed] [Google Scholar]
- Ribeiro P, El-Shehabi F, Patocka N. Classical transmitters and their receptors in flatworms. Parasitology. 2005;131(Suppl):S19–40. doi: 10.1017/S0031182005008565. [DOI] [PubMed] [Google Scholar]
- Rowlands AL, Pagán OR. Parthenolide prevents the expression of cocaine-induced withdrawal behavior in planarians. Eur J Pharmacol. 2008;583:170–172. doi: 10.1016/j.ejphar.2008.01.012. [DOI] [PubMed] [Google Scholar]
- Sabourin TD, Faulk RT, Goss LB. The efficacy of three non-mammalian test systems in the identification of chemical teratogens. J Appl Toxicol. 1985;5:227–233. doi: 10.1002/jat.2550050404. [DOI] [PubMed] [Google Scholar]
- Sarnat HB, Netsky MG. The brain of the planarian as the ancestor of the human brain. Can J Neurol Sci. 1985;12:296–302. doi: 10.1017/s031716710003537x. [DOI] [PubMed] [Google Scholar]
- Sarnat HB, Netsky MG. When does a ganglion become a brain? Evolutionary origin of the central nervous system. Semin Pediatr Neurol. 2002;9:240–253. doi: 10.1053/spen.2002.32502. [DOI] [PubMed] [Google Scholar]
- Schaeffer DJ. Planarians as a model system for in vivo teratogenesis studies. Qual Assur. 1993;2:265–318. [PubMed] [Google Scholar]
- Weiss JN. The Hill equation revisited: uses and misuses. FASEB J. 1997;11:835–841. [PubMed] [Google Scholar]
- Yun AJ, Doux JD, Daniel SM. Brewing controversies: Darwinian perspective on the adaptive and maladaptive effects of caffeine and ethanol as dietary autonomic modulators. Med Hypotheses. 2007;68:31–36. doi: 10.1016/j.mehy.2006.01.068. [DOI] [PubMed] [Google Scholar]