

Abstract
CCK2 receptor antagonists potentiate pain relief by MOP receptor agonists. In an attempt to enhance this effect, we prepared bivalent ligands incorporating CCK2 receptor antagonist and MOP receptor agonist pharmacophores.9 Ligands with 16- to 22-atom spacers could simultaneously bind both receptors but provided no advantage in activity over individual ligands. We now examine the effect of these ligands on receptor internalization as a mechanism of receptor regulation. We prepared CHO cell lines expressing nonfluorescent halves (YN and YC) of yellow fluorescent protein attached to each receptor. Spatial approximation of constructs was needed to yield fluorescence. Monovalent MOP agonist 1 signaled normally and internalized the MOP receptor. Monovalent CCK2 antagonist 2 did not stimulate receptor internalization. In the dual receptor-bearing cells, bivalent ligands 3a–c capable of simultaneously binding both receptors resulted in cell surface fluorescence and internalization of the fluorescent complex in a time- and temperature-dependent manner. Bivalent ligand 4 with spacer too short to occupy both receptors simultaneously yielded no signal. Receptor tethering with appropriate bivalent ligands can down-regulate signaling by moving a nonactivated receptor into the endocytic pathway.
Introduction
Approaches to developing more effective, biologically active ligands have included the combination of two pharmacophores within a ligand that bind to distinct recognition sites within target receptors.1–3 This approach can also be directed toward recognition sites within distinct proteins that are associated with each other within a complex. In this regard, there has been substantial interest in the combination of μ-opioid (MOP) receptor agonists and type 2 cholecystokinin (CCK2a) receptor antagonists for more effective control of pain.3–5 This has been achieved by administering two distinct drugs, as well as by the development of peptide and non-peptidyl bivalent ligands that can bind to both MOP and CCK2 receptors, having agonist activity at the former and antagonist activity at the latter.2,6–9
In a recent report, we developed a series of such bivalent ligands having the MOP receptor agonist pharmacophore separated from the CCK2 receptor antagonist pharmacophore by flexible spacers of different lengths.9 Three members of this series (3a–c) with spacers of 16–22 atoms were shown to be able to simultaneously bind to both receptors, while an analogous bivalent ligand 4 with a nine-atom spacer could only bind to each receptor univalently.9 This dependence of spacer length on the simultaneous binding of both pharmacophores supported the interpretation of induced association of these two receptors by a single bivalent ligand. These results provided an opportunity to explore how a ligand that is capable of tethering two distinct receptors would regulate trafficking of those receptors.
We now demonstrate with a CHO cell line that coexpresses both MOP and CCK2 receptors that, while the monovalent MOP agonist 1 stimulates its receptor internalization and the monovalent CCK2 antagonist 2 binds without stimulating the internalization of its receptor, the tethering bivalent ligands 3a–c induce internalization of both receptors via endocytosis. The present study establishes the ability of a bivalent ligand that is capable of simultaneously binding to two receptors to induce the internalization of one of those receptors that would otherwise remain on the cell surface. This could have important pharmacological implications to induce the down-regulation of nonactivated cell surface receptors in other similar systems in which reduced biological responsiveness could be advantageous.
Results
Bimolecular Fluorescence Complementation
We previously utilized bioluminescence resonance energy transfer studies (BRET) to demonstrate that MOP receptors and CCK2 receptors can each form constitutive homo-oligomers but that they do not spontaneously form hetero-oligomers when they are coexpressed.9 We also demonstrated that association of these receptors could be induced by the use of bivalent ligands for each receptor having a suitable length spacer between their respective pharmacophores.9 Figure 2 confirms these previous observations, now using the technique of bimolecular fluorescence complementation. In this experimental strategy, the nonfluorescent halves of a fluorophore like yellow fluorescent protein (YFP) can associate when they are in appropriate proximity to each other and have appropriate relative orientation to form an intact fluorescent indicator (YFP). This provides a fluorescence signal only when the two tagged molecules are physically associated with each other. For this reason, this technique provides a clear advantage for the study of receptor trafficking, in which a fluorescent signal indicates evidence that the specific bimolecular complex is intact in the cellular compartment in which the fluorescence is observed.
Figure 2.

Bimolecular fluorescence complementation. Shown are representative epifluorescence images of COS cells expressing the indicated receptor constructs. Microscopic images show the effective physical complementation of YN-tagged receptor and YC-tagged receptor to yield fluorescent YFP. These observations support the previous reports of constitutive homo-oligomerization of the CCK2 and MOP receptors.9 No YFP fluorescence complementation was observed when the complementarily tagged CCK2 and MOP receptors were coexpressed, again supporting the previous BRET study showing absence of hetero-oligomerization.9 Scale bar: 25 μm.
Indeed, no YFP fluorescence was observed when the individual YN- or YC-tagged CCK2 receptor molecules or the individual YN- or YC-tagged MOP receptor molecules were expressed on the surface of COS cells. When the same receptors were tagged with intact YFP, they produced a strong surface fluorescent signal. In an important series of controls based on the established homo-oligomerization of each of these receptors, surface YFP fluorescence was induced in COS cells coexpressing YN- and YC-tagged CCK2 receptors or coexpressing YN- and YC-tagged MOP receptors. Consistent with the known absence of hetero-oligomeric complexes that might include both of these receptors, no YFP fluorescence was observed when the complementary YN-tagged CCK2 receptor and YC-tagged MOP receptor constructs were coexpressed.
Cells Coexpressing CCK2 and MOP Receptors
A cell line stably expressing the complementarily tagged receptors would be quite useful for studying the possible internalization of the intact complex containing both the CCK2 receptor and the MOP receptor that might be established after treatment with the bivalent ligands. Such a cell line was therefore established in the Chinese hamster ovary (CHO) cell background. The CHO-CCK2-YN/MOP-YC cells stably expressing both YN-tagged CCK2 receptor and YC-tagged MOP receptor were characterized to ensure surface expression of functional receptors. CCK radioligand binding studies established that the CCK2 receptors were expressed on the cell surface in a form capable of binding peptide ligand. The benzodiazepine antagonist L-365,260 (compound 5)9 was able to compete for radioiodinated CCK binding in a concentration-dependent manner (Figure 3A). This was similar to the ability of this antagonist to compete for CCK radioligand binding to wild type CCK2 receptor-bearing CHO cells. [D-Ala2,NMe-Phe4, Gly-ol]enkephalin (DAMGO) (compound 6) stimulation also established the expression of the MOP receptors in a functional form on the cell surface. Here, 6 was able to inhibit the forskolin-stimulated cAMP responses in a concentration-dependent manner (Figure 3B). This, too, was fully analogous to the responses in MOP receptor-bearing CHO cells.
Figure 3.
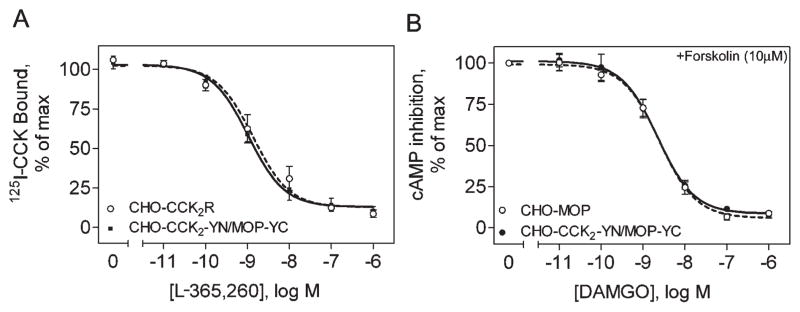
Receptor expression on CHO-CCK2-YN/MOP-YC cells. Shown are concentration–response curves in intact cells, which represent compound 5 competing for CCK radioligand binding (A) and compound 6 inhibiting forskolin-stimulated cAMP responses (B). Data are expressed as the mean ± SEM of values from three to four independent experiments performed in duplicate.
Internalization of CCK and MOP Receptors by Specific Ligands
We investigated the ligand-dependent internalization of MOP and CCK2 receptors in CHO cells stably expressing either YFP-tagged MOP receptors or YFP-tagged CCK2 receptors. Figure 4 shows that monovalent MOP agonist (compound 6) was able to stimulate internalization of the MOP receptor in a time- and temperature-dependent manner. After 30 min, the predominant portion of MOP receptors was in endocytic compartments. In contrast, occupation of the CCK2 receptor with its antagonist, compound 5, did not stimulate its internalization. Analogous observations were made with the monovalent ligands including a spacer with a MOP receptor agonist (compound 1) or a spacer with a CCK2 receptor antagonist (compound 2).
Figure 4.

Ligand-dependent internalization of MOP receptors and CCK2 receptors on CHO cell lines. Shown are representative epifluorescence microscopic images illustrating time-dependent internalization of YFP-tagged MOP receptor exposed to compound 6 (A) and absence of internalization of the YFP-tagged CCK2 receptor exposed to compound 5 (B). In contrast, CCK stimulation of the CCK2 receptor is known to stimulate receptor internalization.19 Also shown are similar images from these cells stimulated with the monovalent MOP receptor agonist (compound 1) (C) and monovalent CCK2 receptor antagonist (compound 2) (D). Scale bar: 25 μm.
Analogous studies were performed with the CHO-CCK2-YN/MOP-YC cell line coexpressing both CCK2 and MOP receptors with the complementary YN and YC tags, respectively (Figure 5). Typical YFP fluorescence was observed in these cells when they were exposed to bivalent ligands binding both receptors simultaneously. This was only true of those ligands having adequately long spacers allowing the ligand to bind to both receptors simultaneously (spacer lengths of 16, 18, and 22 carbon atoms, representing compounds 3a, 3b, and 3c, respectively) (Figure 5).9 The bivalent ligand with a nine-atom spacer (compound 4) that is too short to allow tethering of the two receptors did not induce formation of intact, fluorescent YFP in these cells. Similarly, the bivalent peptidyl ligand capable of binding to these two receptors, Tyr-D-Ala-Gly-D-Trp-NMe-Nle-Asp-Phe-NH2 (compound 7),6 did not induce the formation of intact YFP.
Figure 5.

Bivalent ligand-induced fluorescence complementation in CHO-CCK2-YN/MOP-YC cells. Shown are representative phase contrast and epifluorescence microscopic images of CHO-CCK2-YN/MOP-YC cells exposed to various ligands for 90 min at 4 °C. Compounds 1 and 2, representing monovalent ligands, were unable to form intact fluorescent YFP. Compounds 3a, 3b, and 3c, representing bivalent ligands with adequate length spacers to bind to both receptors simultaneously, were able to form intact fluorescent YFP on the cell surface under these conditions. Compound 4, with the short nine-atom spacer, and the bivalent peptidyl ligand (Tyr-D-Ala-Gly-D-Trp-NMe-Nle-Asp-Phe-NH2, compound 7) were not able to form fluorescent YFP. Scale bar: 25 μm.
Of note, the bivalent ligands that were able to simultaneously bind to both receptors (compounds 3a, 3b, and 3c) also stimulated the internalization of the complex containing both the MOP receptor and the CCK2 receptor into endocytic compartments in the CHO-CCK2-YN/MOP-YC cells, as demonstrated by the presence of reconstituted YFP fluorescence in these compartments (Figure 6). The stimulation of this internalization was observed to be time- and temperature-dependent. No such internalization was observed after exposure of these cells to the bivalent ligand with spacer that is too short to simultaneously bind to both receptors (compound 4) or to the monovalent ligands (compounds 1 and 2).
Figure 6.

Time-dependent internalization of CCK2 and MOP receptors in CHO-CCK2-YN/MOP-YC cells. Bivalent ligands, compounds 3a, 3b, and 3c, that were able to elicit fluorescent YFP by complementation of the tags on the CHO-CCK2-YN/MOP-YC cells also stimulated the time-dependent internalization of these complexes. Scale bar: 25 μm.
Discussion
Agonist receptor ligands often down-regulate their receptor targets by internalization into the cell, leading to desensitization of the signaling mechanism. This can represent a problem for use of such drugs in long-term treatment. In the use of opioid agonists for pain management, this type of receptor regulation can be challenging. One of the therapeutic strategies to potentiate MOP agonist activity has been the addition of a CCK2 receptor antagonist, although the molecular and cellular basis for this is not yet clear. In a recent report, we developed a series of bivalent ligands that incorporate a spacer to link a MOP agonist with a CCK2 receptor antagonist.9 Those ligands included spacers of adequate length to be able to simultaneously bind to both receptors, inducing association of these two receptors. Although these bivalent ligands were not any more effective in producing antinocioception than the combination of the two monovalent ligands, the impact of such agents on receptor internalization has not previously been studied.
Receptor antagonists, unlike agonist ligands, often do not elicit any regulatory response to their binding to their receptors. There are some examples of antagonists stimulating receptor internalization,10,11 but these tend to represent relatively rare exceptions. Similarly, some ligands previously believed to represent antagonists have actually turned out to be biased agonists, eliciting non-G protein-mediated signaling events, often mediated by arrestin-like protein interactions with their receptors.12,13 In the current report, it is well demonstrated that the CCK2 receptor antagonist component of the bivalent agents studied does not stimulate receptor internalization, and it was previously shown to not elicit any signaling response in receptor-bearing cells.9
As is often the case, the MOP agonist pharmacophore incorporated into the bivalent ligand stimulated the internalization of its receptor in a time- and temperature-dependent manner. The pattern of internalization was quite typical of receptor-mediated endocytosis that is so common after agonist stimulation of G-protein-coupled receptors.14
Perhaps the most interesting feature of this work is what the effect might be on the internalization of both the MOP receptor and the CCK2 receptor after their occupation with the bivalent ligands. The bivalent ligand with the short nine-atom spacer that does not allow simultaneous binding to both receptors stimulated internalization of the MOP receptor but not internalization of the CCK2 receptor. The bivalent peptide ligand that was previously reported (7)6 had a similar effect on these receptors. Without existing precedent, it was equally plausible that simultaneous occupation of both the MOP and the CCK2 receptors with a tethering bivalent ligand could interfere with the internalization of the MOP receptor by being retained on the cell surface or could stimulate internalization of the CCK2 receptor. Indeed, the latter turned out to represent the dominant action. All three bivalent ligands that were capable of bridging the MOP and CCK2 receptors had the effect of stimulating the internalization of both MOP and CCK2 receptors in a time- and temperature-dependent manner.
Since receptor internalization induced by the bivalent ligands prevailed over the tendency for the second receptor to remain on the cell surface, this effect might have the expected impact of down-regulating the CCK2 receptor and the biological effects of subsequent stimulation with CCK2 receptor agonists. This could also theoretically result in making such bivalent ligands less effective than two monovalent ligands. The other interesting extension of these observations is the possibility of targeted “dragging” of a receptor from the cell surface without the presence of an agonist for that receptor in order to induce its down-regulation. This might represent a novel strategy to regulate a receptor-mediated process.
Experimental Section
Materials
Synthetic cholecystokinin octapeptide (CCK-8; CCK-26–33) was purchased from Peninsula Laboratories (Belmont, CA). Monovalent agonist ligand for the μ-opioid (MOP) receptor (compound 1 in Figure 1), monovalent antagonist ligand for the CCK2 receptor (compound 2 in Figure 1), and analogous bivalent ligands for both receptors (compounds 3a, 3b, 3c, and 4 in Figure 1) were prepared as described previously.9 [D-Ala2,NMe-Phe4,Gly-ol]enkephalin (compound 6) was purchased from Bachem (Torrance, CA). The previously described bivalent peptidyl ligand of the MOP receptor and the CCK2 receptor, Tyr-D-Ala-Gly-D-Trp-NMe-Nle-Asp-Phe-NH2 (compound 7),6 was synthesized in our laboratory and was purified to homogeneity by reversed-phase high performance liquid chromatography (HPLC). The CCK-like radioligand, 125I-D-Tyr-Gly-[(Nle28,31)CCK-26–33], was prepared in our laboratory, as described previously.15 Forskolin, isobutyl-1-methylxanthine (IBMX), and DEAE-dextran were purchased from Sigma Chemical Co. (St. Louis, MO). Dulbecco’s modified Eagle’s medium (DMEM), Ham’s F-12 medium, and lipofectamine-plus reagent were obtained from Invitrogen (Carlsbad, CA). The cAMP assay kit using LANCE technology and white optiplates were obtained from PerkinElmer Life Sciences (Wellesley, MA). Fetal clone II culture medium supplement was obtained from Hyclone Laboratories (Logan, UT). All other reagents were analytical grade.
Figure 1.

Structures of monovalent and bivalent ligands. Shown are the chemical structures of the monovalent ligands with MOP (1) or CCK2 (2) receptor pharmacophores and bivalent ligands (3a–c, 4) containing both pharmacophores.
Receptor Constructs
Receptor constructs representing YFP-(1–158) (YN) attached in frame to the carboxyl terminus of the CCK2 receptor or YFP(159–238) (YC) attached in frame to the carboxyl terminus of the MOP receptor were prepared by inserting the relevant fragments in front of the TGA stop codons. Constructs were subcloned into pcDNA 3.0 vector (Invitrogen, Carlsbad, CA) for G-418 resistance and pcDNA 3.0/Zeo+ vector (Invitrogen, Carlsbad, CA) for zeocin resistance. All sequences were confirmed by direct DNA sequencing.
Cell Culture
African green monkey kidney (COS-1) cells cultured in DMEM supplemented with 5% fetal clone II in the presence of 5% CO2 on tissue culture plasticware in a humidified incubator were used for transient receptor expression. For this, cells were plated in 10 cm tissue culture dishes at a density of 0.5 million cells/dish 24 h prior to transfection. The cells were transfected with 3 μg of DNA using the DEAE-dextran method16 48–72 h prior to use.
Stable receptor-bearing cell lines were prepared in CHO cells. Cells stably coexpressing YN-tagged CCK2 receptor and YC-tagged MOP receptor were created for the internalization studies using sequential transfections. Parental CHO cells that do not endogenously express either CCK2 or MOP receptors were used to make the cell lines. Approximately 1 × 106 CHO cells were seeded in 10 cm tissue culture dishes in Ham’s F-12 medium and were transfected with 5 μg of the YN-tagged CCK2 receptor construct using the lipofectamine-plus reagent, as per instructions of the manufacturer. Forty-eight hours later, the receptor-expressing cells underwent selection with zeocin (1 mg/mL) and were cloned by limiting dilution. Selected CCK2 receptor-bearing cells were used for subsequent transfection with the YC-tagged MOP receptor construct. These cells (CHO-CCK2-YN/MOP-YC) were selected with G-418 (1 mg/mL) and were also cloned by limiting dilution. Expression of both receptors was confirmed by radioligand binding and agonist-stimulated biological activity.
Isolation of Cell Membranes
Cell membranes were isolated from confluent CHO cell lines, as described previously.17 Cells were harvested mechanically using a cell scraper and were suspended in phosphate-buffered saline (PBS) (pH 7.4), and the receptor-bearing membrane fraction was isolated using differential sucrose density centrifugation.18 Membranes were suspended in Krebs–Ringer HEPES (KRH) medium containing 25 mM HEPES, pH 7.4, 104 mM NaCl, 5 mM KCl, 2 mM CaCl2,1 mM KH2PO4, 1.2 mM MgSO4, 0.01% soybean trypsin inhibitor, and 1 mM phenylmethylsulfonyl fluoride and were stored at −80 °C until use.
Radioligand Binding Assays
Receptor binding characterization was performed with intact cells or cell membranes, as previously described.18 Cells or membrane suspensions were incubated with 1–2 pM radioligand (125I-D-Tyr-Gly-[(Nle28,31)CCK-26–33], specific radioactivity 2000 Ci/mmol) in the absence or presence of nonradioactive ligand for 1 h at room temperature in KRH medium, pH 7.4, containing 0.2% bovine serum albumin and 0.01% soybean trypsin inhibitor. Receptor-bound radioligand was separated from free radioligand by centrifugation and washing with ice-cold buffer. Receptor-bound radioactivity was quantified using a γ spectrometer. Nonsaturable binding, determined in the presence of 1 μM CCK, represented less than 15% of total binding. Data were analyzed and plotted using the nonlinear least-squares curve-fitting routine in Prism (GraphPad 4.0, San Diego, CA).
Biological Activity Assays
Approximately 8000 cells were plated in each well of a 96-well tissue culture plate for cAMP assays, performed as previously described.16 Cells were exposed to various concentrations of compound 6 in the presence of forskolin (10 μM). Stimulation of the Gi-coupled MOP receptor inhibited the forskolin-stimulated adenylate cyclase-induced cAMP responses. Competition-binding assays to quantify cAMP were performed according to manufacturer’s instructions using white optiplates, LANCE kits, and the 2103 Envision plate reader from PerkinElmer (Wellesley, MA).
Receptor Internalization Assays
Morphological approaches were utilized to evaluate receptor localization and internalization of YFP-tagged receptors or the bimolecular fluorescence complementation of YN- and YC-tagged constructs. Cells were grown on coverslips and were washed twice with PBS containing 0.08 mM CaCl2 and 0.1 mM MgCl2. The cells were then incubated with 100 nM ligand (compounds 1, 2, 3a, 3b, 3c, and 4) at 4 °C for 90 min. After incubation, the cells were washed with ice-cold PBS and the occupied receptors were then allowed to internalize in the presence of prewarmed 37 °C PBS for various periods of time. At each time point, the cells were fixed with 2% paraformaldehyde, washed twice with PBS, and then mounted on slides. Fluorescence was observed using an Axiovert 200 M inverted epifluorescence microscope (Carl Zeiss, Thornwood, NY) with fixed YFP filter set (excitation, 500/20 nm; dichroic mirror, Q515 lp; and emission, 535/30 nm) (Chroma Technology Corp., Brattleboro, VT). Images were collected using an ORCA-12ER CCD camera (Hamamatsu, Bridgewater, NJ) with automated QED-InVivo 2.03 image acquisition software (Media Cybernetics Inc., Silver Spring, MD). Final morphologic figures were assembled using Photo-shop 7.0 (Adobe Systems, Mountain View, CA).
Acknowledgments
The authors thank Mary-Lou Augustine and Alicja M. Ball for their excellent technical assistance. This work was supported by grants from the National Institutes of Health (Grant DK32878 to L.J.M. and Grant DA01533 to P.S.P.) and the Fiterman Foundation (L.J.M.)
Footnotes
Abbreviations: BRET, bioluminescence resonance energy transfer studies; CCK, cholecystokinin; CCK2, type 2 cholecystokinin; CHO, Chinese hamster ovary; DAMGO, [D-Ala2,NMe-Phe4,Gly-ol]enkephalin; DMEM, Dulbecco’s modified Eagle’s medium; HPLC, high performance liquid chromatography; KRH, Krebs–Ringer HEPES medium; PBS, phosphate buffered saline; YFP, yellow fluorescent protein; YN, YFP(1–158); YC, YFP(159–238).
References
- 1.Daniels DJ, Kulkarni A, Xie Z, Bhushan RG, Portoghese PS. A bivalent ligand (KDAN-18) containing delta-antagonist and kappa-agonist pharmacophores bridges delta2 and kappa1 opioid receptor phenotypes. J Med Chem. 2005;48:1713–1716. doi: 10.1021/jm034234f. [DOI] [PubMed] [Google Scholar]
- 2.Erez M, Takemori AE, Portoghese PS. Narcotic antagonistic potency of bivalent ligands which contain beta-naltrexamine. Evidence for bridging between proximal recognition sites. J Med Chem. 1982;25:847–849. doi: 10.1021/jm00349a016. [DOI] [PubMed] [Google Scholar]
- 3.Hruby VJ, Agnes RS, Davis P, Ma SW, Lee YS, Vanderah TW, Lai J, Porreca F. Design of novel peptide ligands which have opioid agonist activity and CCK antagonist activity for the treatment of pain. Life Sci. 2003;73:699–704. doi: 10.1016/s0024-3205(03)00390-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Faris PL, Komisaruk BR, Watkins LR, Mayer DJ. Evidence for the neuropeptide cholecystokinin as an antagonist of opiate analgesia. Science. 1983;219:310–312. doi: 10.1126/science.6294831. [DOI] [PubMed] [Google Scholar]
- 5.Lu L, Huang M, Liu Z, Ma L. Cholecystokinin-B receptor antagonists attenuate morphine dependence and withdrawal in rats. NeuroReport. 2000;11:829–832. doi: 10.1097/00001756-200003200-00034. [DOI] [PubMed] [Google Scholar]
- 6.Agnes RS, Lee YS, Davis P, Ma SW, Badghisi H, Porreca F, Lai J, Hruby VJ. Structure–activity relationships of bifunctional peptides based on overlapping pharmacophores at opioid and cholecystokinin receptors. J Med Chem. 2006;49:2868–2875. doi: 10.1021/jm050921q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lee YS, Agnes RS, Badghisi H, Davis P, Ma SW, Lai J, Porreca F, Hruby VJ. Design and synthesis of novel hydrazide-linked bifunctional peptides as delta/mu opioid receptor agonists and CCK-1/CCK-2 receptor antagonists. J Med Chem. 2006;49:1773–1780. doi: 10.1021/jm05085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lee YS, Agnes RS, Davis P, Ma SW, Badghisi H, Lai J, Porreca F, Hruby VJ. Partial retro-inverso, retro, and inverso modifications of hydrazide linked bifunctional peptides for opioid and cholecystokinin (CCK) receptors. J Med Chem. 2007;50:165–168. doi: 10.1021/jm061268p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zheng Y, Akgun E, Harikumar KG, Hopson J, Powers MD, Lunzer MM, Miller LJ, Portoghese PS. Induced association of mu opioid (MOP) and type 2 cholecystokinin (CCK2) receptors by novel bivalent ligands. J Med Chem. 2009;52:247–258. doi: 10.1021/jm800174p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Akgun E, Korner M, Gao F, Harikumar KG, Waser B, Reubi JC, Portoghese PS, Miller LJ. Synthesis and in vitro characterization of radioiodinatable benzodiazepines selective for type 1 and type 2 cholecystokinin receptors. J Med Chem. 2009;52:2138–2147. doi: 10.1021/jm801439x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Roettger BF, Ghanekar D, Rao R, Toledo C, Yingling J, Pinon D, Miller LJ. Antagonist-stimulated internalization of the G protein-coupled cholecystokinin receptor. Mol Pharmacol. 1997;51:357–362. [PubMed] [Google Scholar]
- 12.Rajagopal K, Whalen EJ, Violin JD, Stiber JA, Rosenberg PB, Premont RT, Coffman TM, Rockman HA, Lefkowitz RJ. Beta-arrestin2-mediated inotropic effects of the angiotensin II type 1A receptor in isolated cardiac myocytes. Proc Natl Acad Sci US A. 2006;103:16284–16289. doi: 10.1073/pnas.0607583103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Violin JD, Lefkowitz RJ. Beta-arrestin-biased ligands at seven-transmembranereceptors. Trends Pharmacol Sci. 2007;28:416–422. doi: 10.1016/j.tips.2007.06.006. [DOI] [PubMed] [Google Scholar]
- 14.Roettger BF, Rentsch RU, Pinon D, Holicky E, Hadac E, Larkin JM, Miller LJ. Dual pathways of internalization of the cholecystokinin receptor. J Cell Biol. 1995;128:1029–1041. doi: 10.1083/jcb.128.6.1029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Powers SP, Pinon DI, Miller LJ. Use of N,O-bis-Fmoc-D-Tyr-ONSu for introduction of an oxidative iodination site into cholecystokinin family peptides. Int J Pept Protein Res. 1988;31:429–434. doi: 10.1111/j.1399-3011.1988.tb00899.x. [DOI] [PubMed] [Google Scholar]
- 16.Harikumar KG, Pinon DI, Miller LJ. Transmembrane segment IV contributes a functionally important interface for oligomerization of the class II G protein-coupled secretin receptor. J Biol Chem. 2007;282:30363–30372. doi: 10.1074/jbc.M702325200. [DOI] [PubMed] [Google Scholar]
- 17.Hadac EM, Ghanekar DV, Holicky EL, Pinon DI, Dougherty RW, Miller LJ. Relationship between native and recombinant cholecystokinin receptors: role of differential glycosylation. Pancreas. 1996;13:130–139. doi: 10.1097/00006676-199608000-00003. [DOI] [PubMed] [Google Scholar]
- 18.Harikumar KG, Pinon DI, Wessels WS, Prendergast FG, Miller LJ. Environment and mobility of a series of fluorescent reporters at the amino terminus of structurally related peptide agonists and antagonists bound to the cholecystokinin receptor. J Biol Chem. 2002;277:18552–18560. doi: 10.1074/jbc.M201164200. [DOI] [PubMed] [Google Scholar]
- 19.Cheng ZJ, Harikumar KG, Ding WQ, Holicky EL, Miller LJ. Analysis of the cellular and molecular mechanisms of trophic action of a misspliced form of the type B cholecystokinin receptor present in colon and pancreatic cancer. Cancer Lett. 2005;222:95–105. doi: 10.1016/j.canlet.2004.09.008. [DOI] [PubMed] [Google Scholar]