

Abstract
We examined the interactions between visual recognition memory, working memory, and categorization by examining 6-month-old infants’ (N = 168) memory for individual items in a categorized list (e.g., images of dogs or cats). In Experiments 1 and 2, infants were familiarized with 6 different cats or dogs, presented one at a time on a series of 15-s familiarization trials. When the test occurred immediately after the sixth familiarization trial (Experiment 1), infants showed strong novelty preference for items presented on the fourth or fifth familiarization trial, but not for the items presented on the first three trials or on the sixth trial. When a brief (15-s) retention delay occurred between the end of the sixth trial and the test trials (Experiment 2), memory for the sixth item was enhanced, memory for the fourth item was impaired, and memory for the fifth was unchanged relative to when no retention delay was included. Experiment 3 confirmed that infants can form a memory for the first item presented. These results reveal how factors such as interference and time to consolidate influence infants’ visual recognition memory as they categorize a series of items.
Infants’ visual recognition memory has been studied for decades (see Fagan, 1990; and Rose, Feldman, & Jankowski, 2004 for reviews). The vast majority of these studies have used tasks in which infants first are familiarized with one or more items on a series of trials and then their preference for a novel item over the now-familiar item is assessed. Variations of this procedure have revealed that the strength of infants’ preference for novelty in this task depends on the infants’ age, amount of study time, stimuli used, and other factors (Fagan, 1990). Such findings have lead to conclusions about how memory in this context is influenced by speed of processing, attention, and other factors (see Rose, et al., 2004 for a review).
A variation of this task has been used to examine categorization in infants. Rather than familiarizing infants with a single item, studies aimed at uncovering categorization processes use a procedure in which several items from within a category are presented during familiarization (see Oakes, 2008 for a review), and then infants are shown novel items from the now-familiar category and from a contrasting category. Categorization is inferred by their pattern of novelty preference. Studies using this approach have revealed that young infants are sensitive to adult-defined categories such as cat and dog (Furrer & Younger, 2005; Oakes & Ribar, 2005; Quinn, Eimas, & Rosenkrantz, 1993) and form prototypes of items presented during familiarization (Younger, 1985). Moreover, this response to the novel category has been observed in studies using familiarization even when infants’ looking time did not decrease over familiarization with multiple items from within a category (Furrer & Younger, 2005; Quinn, 2004; Quinn & Eimas, 1996).
Despite the fact that studies of memory and categorization in infancy use nearly identical procedures, little work has examined how memory and categorization together influence infants’ responding in such tasks. For example, when infants who were familiarized with one face fail to prefer a novel face, does this mean that they do not remember the now familiar face, recognize and attend to the fact that both images are faces, or some combination? Similarly, when infants who have been familiarization with a series of cats look equally at a novel cat and dog, does this mean they do not recognize a narrow category that includes cats but not dogs or that they recognize that both novel instances are new? Although a few studies have examined infants’ memory for the individual items in these categorization tasks (Kovack-Lesh, Horst, & Oakes, 2008; Quinn, Yahr, Kuhn, Slater, & Pascalis, 2002), because we cannot tell infants that the task is a categorization or memory task it is difficult to know how their responding during test reflects one process or the other, or a combination of the two. The goal of the present investigation was to probe this relation by systematically examining memory processes as infants categorize.
Although studied separately—and generally by different researchers—work in the adult categorization and memory literatures have examined, to some extent, these kinds of interactions. For example, there is a significant controversy in the categorization literature about the role of memory processes during categorization (Ashby & O’Brien, 2005; Knowlton & Squire, 1993; Zaki & Nosofsky, 2001). A highly influential class of theories, known as exemplar theories, has argued that categorization is achieved through the representation of individual exemplars (Nosofsky & Johansen, 2000). According to this view, forming memory representations for instances and abstract representations of the commonalities among those instances is intertwined. Indeed, computational modeling accounts (Love & Gureckis, 2007), and fMRI studies (Nomura et al., 2007) have shown involvement of medial temporal brain structures—regions involved in recognition memory—in some aspects of categorization. However, patients with damage in these regions have preserved categorization abilities and impaired recognition memory for the items that were categorized (Knowlton & Squire, 1993; but see Palmeri & Flanery, 1999 for an alternative account of these findings), bolstering the claim that recognition memory and categorization are subserved by different neural systems. This controversy reflects, in part, debates about multiple memory systems, and whether categorization is related to one or many of these separate memory systems (see Ashby & O’Brien, 2005 for a review). Regardless of which view is correct, it is clear that researchers attempting to understand categorization in adults have considered how memory and categorization interact.
Researchers focused on memory processes in adults have been less explicit in addressing this question, but some findings suggest that the commonalities across to-be-remembered items—an essential component of categorization—may be important in memory processes. For example, learning a categorized list of items seems to encourage processing of item similarities, and not item-specific information (Hunt & Lamb, 2001). In addition, people remember items in categorized lists when the list structure highlights categorical relatedness better than when it does not (Howe, 2004; Medin & Bettger, 1994). Finally, when learning a categorized list, adults have more false memory for frequent category members than for infrequent category members (Dewhurst, 2001), suggesting that even when the task was to remember the individual words, categorical information was represented and/or activated. The point is that for adults, the relation between the to-be-remembered items contributes to encoding and recalling of items.
To further our understanding of the interactions between categorization and memory processes, we examined infants’ visual recognition memory for individual items following familiarization with multiple items from within a category (i.e., dogs or cats). Memory for a singly presented item has been widely studied in 5- to 7-month-old infants (Fagan, 1990; Rose, et al., 2004), but few studies have examined infants’ memory for multiple items presented on successive trials. Some studies have examined infants’ memory for the individual items in a categorized sequence of familiarization trials (Kovack-Lesh, et al., 2008; Quinn, et al., 2002), but the few studies aimed at uncovering the processes of visual recognition memory in this context have used sets of unrelated items presented on a sequence of trials (Cornell & Bergstrom, 1983; Rose, Feldman, & Jankowski, 2001). Because a large number of studies have shown that across the first year infants familiarized with images of dogs and cats during test exhibit preferences for the novel category (Furrer & Younger, 2005; Kovack-Lesh, et al., 2008; Quinn, 2004; Quinn & Eimas, 1996), we assume that such familiarization will induce categorization. The question we asked here is given that infants are categorizing, and recognizing and encoding the commonalities among the items, what processes are engaged in their encoding of and retaining the details of the individual items?
Specifically, we asked how infants’ memory for individual items was influenced by position in the study “list” (i.e., which familiarization trial), proactive and retroactive interference, and time between study and test. By comparing infants’ memory for items occurring relatively early and relatively late during familiarization, we could probe for the primacy and recency effects characteristic of adults’ performance on memory tasks (Hay, Smyth, Hitch, & Horton, 2007; Howard, Venkatadass, Norman, & Kahana, 2007; Neath, 1993b; Neath & Crowder, 1996; Tan & Ward, 2000). Infants may encode the details of the first items encountered, and fail to encode the details of later presented items (perhaps focusing on representing the commonalities among the items), and show a primacy effect (Murdock, 1961). Alternatively, infants may attend to and encode the details of each of the items encountered, and once their memory capacity has been reached, the activities involved in encoding items encountered later items may “push out” the memories for the earlier encoded items, producing a recency effect (see Surprenant & Neath, 2009 for a review of the recency effect). Infants’ memory does seem to be subject to position effects. Two studies found evidence of a recency effect when 6- to 7-month-old infants were familiarized with a series of 3 or 4 unrelated visual stimuli (Cornell & Bergstrom, 1983; Rose, et al., 2001). Cornell and Bergstrom also found that infants had superior recognition for the first items presented. These studies examined position effects under different conditions than they are often observed in adult subjects, however. Cornell and Bergstrom repeatedly presented the 3-item “list” (sequence of pictures) to infants; therefore the “first” item was presented on the first trial, fourth trial, seventh trial, and tenth trial. Rose et al. (2001) tested infants’ memory for all the items in the list; infants’ memory for the third item, therefore, was tested after their memory for the first and second item had been tested. Gulya, Sweeney, and Rovee-Collier (2001) used a very different procedure and found that when infants were trained to kick their leg to make different mobiles move (thus the “lists” were a series of mobiles presented in sequence on three daily training sessions), 3- and 6-month-old infants’ memory for the mobiles showed position effects after a 1-week delay. The point is that, like adults, infants’ memory for particular items in a study list is influenced by item position in the list.
Such position effects presumably reflect retroactive interference, or the interfering effects of encoding and storing new items on the maintenance of memories of previously encountered items, and proactive interference, or the interfering effects of having learned items encountered earlier on the encoding and storage of items encountered later (Roediger, Weinstein, & Agarwal, 2010). When learning multiple lists, adults’ memory for the first lists learned becomes increasingly worse as other lists are presented before memory for the first list is tested (Postman & Keppel, 1967). Apparently, the activity involved in learning subsequent information interferes with memory for the information learned first. Other work has shown that subjects’ memory for information encountered on early trials can hinder their performance on subsequent trials (Makovski & Jiang, 2008), illustrating that the storage of old information can inhibit the processing of and memory for newly encountered information. The evidence on the effects of interference on infants’ memory is mixed (see Cohen, DeLoache, & Pearl, 1977; Fagan, 1977; McCall, Kennedy, & Dodds, 1977). When required to remember one or two items, infants only experience interference when they fully process interfering information. Because no studies have examined the effects of interference when infants are presented with multiple items in succession—a context in which infants may or may not have the resources, time, or motivation to fully process each item presented—it is unclear how retroactive and proactive interference will influence infants’ memory for serially presented items. Uncovering such effects was another goal of the present study.
It is important to note that the effects of position and interference are confounded with differences in the passage of time between study and test. For example, when more information is encountered between study and test (thus providing more opportunities for interference), there may also have been more time between study and test. Memory involves processes that occur over time, even after the encoding episode is complete (McGaugh, 1966), and people’s memory for events and stimuli vary depending on the delay between encoding and recall (Knoedler, Hellwig, & Neath, 1999; Posner & Konick, 1966). Even infants’ memory changes over time. The main measure of memory in infants, their preference for a novel stimulus as compared to a previously seen one, varies as a function of time between study and test. When familiarization is relatively short, infants prefer the familiar item, and their preference for the novel item becomes stronger with increases in the familiarization period. Although such novelty preferences can be maintained for seconds, minutes, hours, and even days after the encoding period (see Fagan, 1990, for a review), when there is a very long delay between study and test, infants show a null preference, and a preference for the familiar stimulus may re-emerge when the delay is very long (Bahrick & Pickens, 1995; Courage & Howe, 2001).
There is some controversy about how memories change over time. Traditionally, the primary effect of time on memory was thought to be forgetting, because of decay or loss from storage, or because memories have become inaccessible to retrieval (see Roediger, et al., 2010, for a review). Even in infancy, apparently forgotten memories can be reactivated with a reminder (Bahrick, Hernandez-Reif, & Pickens, 1997), suggesting that memories have not decayed, but have become inaccessible to retrieval processes. Moreover, in adults, forgetting appears to be a function, in part, of the activities of processing, encoding, and storing information encountered after the memories occur (Nairne, 2002). Regardless of the precise mechanisms, infants and adults do “forget” learned information over time.
Memories also can become stronger over time as consolidation and storage processes that occur after encoding that render memories more stable (Alvarez & Cavanagh, 2004; Dudai, 2004), perhaps due to neural organization and reorganization (Dudai, 2004; Hardt, Einarsson, & Nader, 2009). In adults, memory is weaker if interfering information occurs too soon after the study period than if it is presented after a delay, suggesting that consolidation of the to-be-remembered information during the period immediately after study is important for subsequent memory of that information (Dewar, Garcia, Cowan, & Della Sala, 2009). Such processes seem to operate in infancy. Catherwood et al. (1996) found that when infants were exposed to visual stimuli for a few seconds on a series of familiarization trials, the amount of time between stimulus onsets—regardless of whether or not during that time the to-be-remembered-items were visible or masked—determined memory performance. Catherwood et al. concluded that although some features were perceived and rapidly encoded, other features required processing beyond the exposure time for memories to become stable enough to allow infants to show a systematic response during test. Thus, time to fully process the stimuli—regardless of whether those stimuli are present—may be essential for evidence of visual recognition memory in infancy. We addressed this possibility in the present investigation.
We examined 6-month-old infants’ visual recognition memory for categorized items (i.e., pictures of cats or dogs) presented over a series of trials. By comparing infants’ memories for items presented in different positions in the sequence (e.g., the first presented versus the fourth presented versus the last presented item), we begin to understand how infants’ memory changed over time and is influenced by encoding, maintaining, storing, etc. of other presented items. To preview our results, Experiment 1 reveals better memory for items presented in some positions in the “list,” although the results do not fit typical primacy and recency effects. Experiment 2 shows that including a short retention delay (15 s) between the last presented item and the test of memory causes infants to show robust memory for items in a different set of serial positions than did the infants in Experiment 1 who did not experience the retention delay. Experiment 3 confirms that infants can remember the first item presented, at least when it is not followed by a series of other similar items. Together, these experiments show how infants’ memory changes over time.
Experiment 1
Method
Participants
The final sample consisted of 72 healthy, full-term 6-month-old infants (M = 197.95 weeks, SD = 7.93; 38 girls and 34 boys; 37 infants had a cat or dog at home), with no known history of neurological or vision problems. Twelve additional infants were tested but were excluded because they became too fussy to complete the session or they refused to look at the computer monitors. Sixty-five infants were tested at The University of Iowa and seven infants were tested at UC Davis. Sixty-six infants were reported to be White, four infants were mixed race, and the race of two infants was not reported; one of the infants was reported to be Hispanic. All mothers had completed high school; all but four mothers had some college, and 51 mothers had completed at least a bachelor’s degree. Thus, as a whole, infants in the sample had generally well-educated parents, and infants tested in the two locations did not differ in SES.
Infant names were obtained from the State Department of Health or purchased from a professional list broker in California and from county birth records in Iowa. All parents were sent informational letters and then were contacted by phone and/or e-mail. Infants were given a small toy or t-shirt in appreciation for their participation.
Apparatus
The stimuli were presented on two 17-in (43.2 cm) CRT monitors (the same monitors were used at both locations) center-to-center distance between the two monitors was 52 cm. A box, with a light that could blink accompanied by a beeping sound, was placed directly between the two monitors. A Macintosh G5 computer was used to present the stimuli and time infant looking time using specially designed software (Cohen, Atkinson, & Chaput, 2000–2002). A low-light security camera was placed beneath the center of the two monitors to record infant behavior on a VCR or DVR.
The equipment sat on a table. A black curtain obstructed the infants’ view of the equipment. Holes in the curtain revealed the two monitors, the box with the attention-getter, and the low-light camera.
Stimuli
The stimuli were scanned images of dogs and cats, taken from books on dog and cat breeds (see Figure 1). The images were cropped and removed from any background using Adobe Photoshop. They were all approximately the same brightness and were approximately 19 by 15 cm (30° by 21° visual angle at a viewing distance of 40 cm). Twelve images of dogs and 12 images of cats were used. The animals were of different breeds, coloring, etc. Animals were all standing and facing the right side of the monitor. Half the dogs and half the cats had their faces turned so you could see their full face; the other dogs and cats had their faces in profile.
Figure 1.

Procedure
Infants were seated on a parent’s lap, approximately 40 cm from the two monitors. Parents wore opaque glasses to occlude their view of the stimuli, thus ensuring that parental behavior did not bias infants’ responding. Each trial began with the blinking, beeping light. Once infants fixated on this light, an experimenter, seated out of sight, initiated the trial by pressing a key on the computer that simultaneously ended the blinking light and presented the stimuli for that trial. The experimenter pressed one key when he or she judged that the infant was looking at the left screen and a different key when he or she judged that the infant was looking at the right screen. A second coder recorded looking times from the recordings of 15 infants’ sessions. Reliability between the on-line and off-line coding was good; average correlation between the looking times recorded on each trial by the two coders was r = .96, and the average mean difference in looking per trial was .40 s.
Infants first received six 15-s familiarization trials; on each trial, two identical images of a different dog or cat were presented (half of the infants in each group were familiarized with cats and half were familiarized with dogs). Thus on each familiarization trial, infants could learn about and form a memory for a single animal. Across familiarization infants saw six different dogs or six different cats.
To test memory for the 6 familiarization items, immediately following familiarization we presented each infant with two 10-s test trials on which one of the familiar items was paired with a new item from the same category (e.g., a new cat if the infant had been familiarized with cats). The average inter-trial interval between the last familiarization trial and the first test trial across all conditions was M = 2.54 s (SD = 2.03). For each infant, the same items were presented on the two test trials; the left-right position was counterbalanced across trials, thus minimizing the effect of strong side biases. Infants were randomly assigned to one of six groups (n = 12 per group) depending on which of the familiarization items was presented as the familiar test item. By examining which group of infants showed a significant novelty preference (i.e., looking longer at the new item more than 50% of the time), we could establish whether infants remembered items presented in some serial positions than in others.
To ensure that none of our effects could be explained by preferences for particular stimuli, each infant in each condition received a different pair of stimuli, and each stimulus was the novel stimulus in many different conditions. Moreover, each stimulus was presented approximately the same number of times as the familiar stimulus as it was the novel stimulus. Thus, we used as many different pairs of stimuli as possible as tests. Many stimuli were novel for more than one group of infants. This aspect of our design ensures that our results are generalizable and not specific to particular stimuli, and by creating many different pairs of novel and familiar stimuli the effects of preferences for one particular item over another particular item are minimized.1
Results
Initial analyses confirmed that there were no effects on infants’ responding in any of the experiments due to infant gender, previous pet experience2, or the particular category presented during familiarization. Therefore, all of the reported analyses collapsed across these variables. We analyzed infants’ responding during familiarization and test separately.
Responding during familiarization
The mean looking time during each familiarization trial for infants tested on the items in each serial position is presented in Table 1. These data illustrate several important facts about infants’ responding. First, across the six groups, there was little variation in how long infants looked at the item that was eventually presented during test (note that during familiarization, neither the infants nor the experimenter could be aware of which item was to be presented during test). We confirmed this observation by entering infants’ looking during the trial in which the to-be-familiar item was presented (the first familiarization trial for infants who would be tested on their memory for the first item, the second familiarization trial for infants who would be tested on their memory for the second item, and so on) into an Analysis of Variance (ANOVA) with group (i.e., the particular item presented as the familiar during test) as the between factor. This analysis revealed no significant effect of group, p = .94, indicating that infants looked similarly at the item that was used as a familiarization item, regardless of when it was presented during familiarization. Thus, if there are differences in infants’ responding to a novel item when it is compared to one of these familiar items, it cannot be attributed to differences in infants’ study time to the relevant item.
Table 1.
Mean looking per trial (in s, SD in parentheses) for infants in each experiment grouped by the particular item used as the familiar item.
| Group | N | Trial 1 | Trial 2 | Trial 3 | Trial 4 | Trial 5 | Trial 6 |
|---|---|---|---|---|---|---|---|
| Experiment 1 | 72 | 8.38 (3.24) | 7.81 (3.19) | 8.34 (2.76) | 7.28 (3.06) | 7.60 (3.33) | 7.82 (2.91) |
| One | 12 | 8.44 (2.97) | 6.63 (2.96) | 7.68 (2.76) | 6.91 (3.53) | 6.53 (4.19) | 7.43 (3.05) |
| Two | 12 | 9.44 (3.00) | 8.33 (2.60) | 8.46 (2.66) | 7.38 (2.45) | 8.78 (2.69) | 7.33 (2.51) |
| Three | 12 | 7.93 (3.73) | 8.43 (3.51) | 8.10 (3.45) | 7.40 (3.02) | 8.13 (3.36) | 7.55 (2.61) |
| Four | 12 | 8.45 (2.95) | 8.65 (2.65) | 9.47 (3.47) | 7.74 (3.51) | 7.98 (3.06) | 8.52 (3.31) |
| Five | 12 | 8.48 (3.30) | 6.86 (3.53) | 7.78 (3.77) | 6.34 (2.48) | 7.37 (3.25) | 7.56 (3.26) |
| Six | 12 | 7.56 (3.76) | 8.23 (3.76) | 8.58 (2.60) | 7.93 (3.56) | 6.83 (3.39) | 8.49 (2.98) |
| Experiment 2 | 72 | 8.83 (2.83) | 8.47 (2.58) | 8.00 (3.19) | 7.67 (3.25) | 7.64 (3.18) | 7.67 (3.60) |
| Four | 24 | 9.08 (2.46) | 7.90 (2.85) | 7.38 (3.31) | 7.17 (3.14) | 7.65 (3.37) | 6.72 (3.40) |
| Five | 24 | 8.53 (3.08) | 9.32 (2.47) | 9.35 (3.04) | 8.53 (3.57) | 8.55 (3.21) | 9.38 (3.7) |
| Six | 24 | 8.89 (3.00) | 8.20 (2.28) | 7.28 (2.91) | 7.30 (2.97) | 6.70 (2.77) | 6.90 (3.19) |
| Experiment 3 | |||||||
| One | 24 | 8.53 (3.11) | |||||
Second, although infants’ looking varied somewhat from trial to trial, looking did not systematically decrease across trials and the groups did not differ in their pattern of looking. Infants’ looking on each familiarization trial was entered into an ANOVA with trial (1 through 6) as the within-subjects factor and group (the item presented during test as the familiar) as the between subject factor. This analysis revealed a marginally significant effect of trial, F (5, 330) = 2.01, p = .08, ηp2 = .03, but no significant effect of group or group by trial interaction, ps > .94. The linear, quadratic, and cubic trends were not significant, ps ≥ .16, suggesting that the pattern across the trials was nonsystematic. Indeed, we compared the individual means (averaged across all six groups) using two-tailed t-tests and evaluating significance using the False Discovery Rate (FDR) (Benjamini & Hochberg, 1995) for multiple comparisons. These comparisons revealed that the only significant difference was that infants looked significantly less on trial 4 than trial 3, t(71) = 2.96, p = .004, d = .35, and the difference in their looking between trials 1 and 4 was marginally significant, t(71) = 2.61, p = .01, d = .31. None of the other comparisons were significant (trial 1 versus trial 5, p = .09, trial 3 versus trial 5, p = .10; all other p-values ≥ .12). Trial-by-trial comparisons should be considered with caution, however, as infants’ looking on any given trial can be influenced by a number of factors in addition to their processing of the stimuli (Cohen & Menten, 1981).
The lack of a systematic, robust decrease in looking across familiarization is not surprising. Several previous studies in which infants were familiarized with multiple images of dogs and cats have shown failures to decrease looking in a nearly identical procedure, and yet had robust novelty preferences when tested on their category formation (Furrer & Younger, 2005; Quinn, 2004; Quinn & Eimas, 1996). One benefit of the procedure used here is that novelty preference is not dependent on habituation of looking to a familiar stimulus, and requiring habituation may actually underestimate infants’ memory abilities (Fagan, 1990).
Moreover, the lack of a decrease is potentially important because any differences we observe will not be the result of differences in level of interest at the end of the session, fatigue, or any other factor unrelated to the hypotheses being tested here. Because infants in this experiment studied the items presented on each trial for approximately the same amount of time, there is no reason to predict that there are differences in how well infants have encoded the items presented on the 6 familiarization trials.
Responding to the test trials
The key analyses were those conducted on infants’ novelty preference during test. We calculated a novelty preference score by dividing infants’ looking during test to the novel item by their total looking duration to the novel and familiar items during test. The mean novelty preference scores for each group (and the scores for each individual infant) are presented in Figure 2. Chance level, reflecting equal looking at the two images, is .50. As a group, infants failed to exhibit a robust and consistent novelty preference when their memory for the first, second, or third item presented during familiarization was tested (the mean responding is represented by the line intersecting the bar for each group). Infants in this experiment did exhibit a robust and consistent novelty preference when their memory for fourth or fifth item presented during familiarization was tested. Interestingly, the group of infants tested on the sixth item failed to exhibit a robust or consistent novelty preference.
Figure 2.
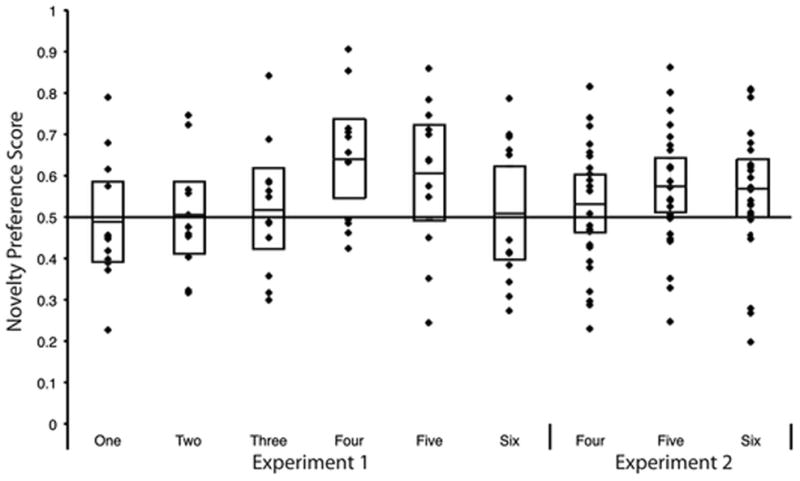
We confirmed these observations by comparing the novelty preference score for each group of infants to chance using two-tailed t-tests. Infants tested on the fourth familiarization item preferred the novel item more than would be expected by chance, t(11) = 3.25, p = .008, d = .94, and infants tested on the fifth familiarization item had a novelty preference score that was marginally significantly different from chance, t(11) = 2.06, p = .06, d = .60. Infants tested on the first, second, third, and sixth familiarization items did not have scores that differed significantly from chance, all ts < 1.0. Thus, these data suggest that infants remembered the fourth and fifth items in the list, but they did not show evidence of having remembered the first through third or the sixth items in the list.
We conducted an ANOVA on the novelty preference scores with group as the between subject factor. The main effect of group approached significance, F(5, 71) = 1.95, p = .10, ηp2 = .13. As is evident from Figure 2, infants’ preference for novelty was complexly related to the serial position of the familiar item. Consistent with the results comparing infants’ responding to chance, comparison of the individual groups of infants, determining significance using FDR (Benjamini & Hochberg, 1995), revealed that infants tested with the fourth familiarization item had a novelty preference score that was significantly greater than the scores of infants tested with the first familiarization item, t(22) = 2.47, p = .02, d = 1.00, or second familiarization item, t(22) = 2.44, p = .02, d = 1.00, and marginally greater than infants tested on the third familiarization item, t(22) = 2.04, p = .05, d = .83, or sixth familiarization item, t(22) = 1.96, p = .06, d = .80. None of the other comparisons were significant, although the novelty preference for infants tested on the fifth familiarization item was marginally greater than the novelty preference for infants tested with the first and second items, ps ≤ .10, effect sizes .71 and .68 respectively.
Figure 2 also shows each individual infant’s novelty preference score. No standard exists for determining when infants have exhibited a novelty preference, null preference, or familiarity preference. Inspection of the figure reveals that infants tested with the fourth and fifth item tended to have scores above .50 (67 and 75 percent of the infants in these groups had scores over .5), whereas a smaller proportion of infants tested on the other items had scores above .50. Moreover, the distribution of the scores differed for the groups. It is particularly striking that the scores for groups of infants tested on the first three items tended to cluster around .50, with few infants showing extreme familiarity or novelty preference scores. The scores for the group of infants tested with the sixth item, in contrast, who also had a mean novelty preference score that was near .50, was bimodal. About half the infants had relatively strong novelty preferences, and the other infants had modest to relatively strong familiarity preferences; few infants had scores near .50. Inspection of these scores both confirms the impression from the analysis of the mean novelty preference scores that only infants tested with the fourth and fifth item showed a clear novelty preference, and also revealed that the null preferences exhibited by infants tested with the first through third item reflected a different distribution of individual scores than for the infants tested with the sixth item.
Discussion
These results provide an initial understanding into infants’ memory for the individual items presented in a categorized list. Although we did not observe clear primacy or recency effects as observed by others (Cornell & Bergstrom, 1983; Rose, et al., 2001), our results do suggest that infants’ visual recognition memory in this context is influenced by position of the items during study. We found that infants showed the strongest memory for the items in the middle of the list, and no clear memory for the last item presented. It is possible that our results differed from previously reported results due to procedural or stimulus differences. This issue will be revisited in the General Discussion.
This pattern may reflect the effects of interference as infants maintained and continued processing items previously encountered when new items are encountered. That is, if forming a stable, enduring representation of the individual items required more processing time than was available during the 15-s trials, infants would still be processing this information after the trial ended, presumably engaging working memory processes. Moreover, because in our procedure inter-trial-intervals were extremely brief (2 to 3 s), this post-exposure processing would have occurred during the next trial, when a new item was available. Thus, infants may have experienced a conflict for processing resources. Their continued processing of the previously encountered item may have interfered with their processing of newly encountered items, particularly if they were simultaneously categorizing and forming memories of those individual items. In addition, encoding and storing newly encountered items may have interfered with maintenance of previously processed items. That is, infants’ responding may have reflected, in part, the combined effects of proactive and retroactive interference on infants’ memory.
The evidence for retroactive interference is that infants did remember items relatively late in the list, but not those encountered first. Encoding and maintaining memory for the fourth and fifth item in the list—and even the activities surrounding encoding of the sixth item—may have interfered with infants’ maintenance or recall of the first three items encountered. Proactive interference is suggested by the fact that infants did not show robust memory for sixth item. That is, infants’ encoding, consolidation, and retention of the items encountered on the immediately preceding trials may have interfered with their processing of that sixth item. In other words, processing items may both have “pushed out” memories of previously encountered items and at the same time delayed the processing of subsequently encountered items. As a result, infants may have the strongest memories for (or the least ambiguous responding to) items that have effectively interfered both with memories for previously encountered and with forming memories of later encountered items—in this study, items 4 and 5.
This conclusion is based on several assumptions. First, infants’ memory capacity in this context must be limited. Indeed, previous work has established that infants’ limited memory capacity in this context (Rose, et al., 2001), as well as limited capacity of visual short-term or working memory systems that may be used when the items are not visible (Kaldy & Leslie, 2005; Ross-Sheehy, Oakes, & Luck, 2003). As a result of these capacity limitations, we should observe that infants robustly remember only 1 or 2 items in this task. A second assumption is that forming strong memories for some items may have interfered with the maintenance of the previously stored items. That is, after items are no longer visible, infants must store them in short-term or working memory; when new items are encountered and capacity is exceeded, the new representations “push out” the old representations. In this case, infants should not show memory for the items encountered before the remembered items. If we test a condition in which infants show robust memory for items 5 and 6, therefore, retroactive interference should result in poor memory for item 4.
A third assumption is that storage processes take more than 15 s (the duration of the trial) to complete—particularly when infants are engaged in the simultaneous tasks of storing previously encountered items, encoding newly encountered items, comparing items across trials, extracting commonalities among items, and other processes related to categorization. Thus, after the 15-s trial ends, infants will continue to process information presumably stored in short-term or working memory. Because in the present context the inter-trial interval is relatively short, during every trial starting with trial 2, infants will still be engaged in storing items from previous trial(s), and those processes may interfere with infants’ ability to store the now visible item. Infants may only be able to engage encoding, storage, and/or consolidation processes with respect to a new item only after some aspect of the storage of the previously encountered item is complete. Thus, for item 6, the test with the familiar and novel item occurred while at least some of the infants were still engaged in processing the item presented on that trial; those infants showed a familiarity preference, as if they had not fully processed that item. In this case, providing infants with more time to process the last item should facilitate their memory for that item. If providing infants with additional time does not influence their memory for that item, we will have evidence that the failure of infants to remember the sixth item in Experiment 1 was not the result of proactive interference.
It is worth noting that in the previous demonstrations of recency effects in infants’ visual recognition memory, test for the last encountered item (i.e., the most recent item) did not occur immediately after the first presentation with that item—it occurred after the list was presented several times (Cornell & Bergstrom, 1983) or after infants’ memory for the earlier encountered items had been tested (Rose, et al., 2001). Such procedures may have effectively released infants from proactive interference, allowing them to show strong memory for the last encountered item. In addition, in these previous studies, the to-be-remembered items were unrelated. Thus, it is also possible that the effect of proactive interference we observed was due to infants’ simultaneously categorizing and forming memories for items.
Variations of this task have revealed robust novelty preferences on immediate retention tests (e.g., Fagan, 1977; Rose, 1981; Rose, Gottfried, Mello-Carmina, & Bridger, 1982). Several differences between the present context and these previous studies may have resulted in better retention on immediate tests in the previous tasks than in the present task. For example, studies differ in the nature of the stimuli. The earlier studies tended to use photographs of human faces or abstract geometric patterns; it is possible that stimuli that are more or less familiar, include more or less detail, or are more or less compelling, differently induce memory processes. In addition, we asked infants to remember 6 different items and previous studies asked infants to remember only a single item. Immediate retention may have been impaired in our study because of the effects of retroactive and proactive interference just described, which only can operate when infants are processing multiple stimuli.
The timing of the immediate test also differed across studies. Most previous studies of infant memory used an apparatus in which the stimuli were presented on cards that were physically placed in an apparatus that was wheeled or rotated in the infants’ view; the delay this procedure creates is rarely reported, but it is sometimes reported to be between 3 to 10 s (Cornell & Bergstrom, 1983; Fagan, 1990; Rose, 1981; Rose, Jankowski, & Feldman, 2002, 2008). This contrasts with our inter-trial-interval of about 2.5 s. When infants have several seconds of unfilled time to engage in post-encoding processes before the memory test, as they did in many previous studies, they may show more robust evidence of memory in “immediate” tests. Indeed, Morgan and Hayne (2006) also found a lack of a novelty preference on immediate tests following short familiarization periods in a visual recognition memory task using computer-controlled stimulus presentation which likely also involved relatively short inter-trial-intervals.
The individual infants’ scores depicted in Figure 2 supports the hypothesis that infants’ failure to show robust memory for item 6 is the result of different processes than is their failure to show robust memory for items 1–3. Specifically, for items 1–3 most individual infants showed a novelty preference near .50, whereas for item 6 some infants showed a relatively strong novelty preference and other infants showed a relatively strong familiarity preference (note the differences in the spread of scores in these different groups). Familiarity preferences are typically observed when familiarization is relatively short, and are often taken to mean that infants have not fully processed the stimulus (Roder, Bushnell, & Sasseville, 2000; Rose, et al., 1982). Infants generally show more robust and stable memories when given longer time to process the stimuli, and in the present case infants may have shown a more consistent response indicating memory for the sixth item if they had a longer period of time to process that item, even in the absence of the item (such as was the case for the fourth and fifth items). In other words, the results reported in Experiment 1 may show the unfolding of storage and retention processes in infants over periods of 15 to 45 seconds. Specifically, these results may reflect the fact that memories become more stable over time (McGaugh, 1999, 2000), presumably due to post-encoding processes such as consolidation.
Experiment 2 tested the specific hypothesis that the delay between familiarization and test contributed to infants’ ambiguous response when tested with the sixth familiarization item. We replicated three conditions of Experiment 1 with a minor modification: we included an unfilled retention delay between the sixth familiarization trial and the test. This unfilled retention delay would provide infants extra time to continue processing the last item in the absence of interference from processing an additional item. Thus, if infants’ failure in Experiment 1 to show consistent robust memory for the last item reflected insufficient time to engage in post-encoding processes, the inclusion of this delay should enhance their memory for that item. In addition, if infants do show memory for the last item in Experiment 2, their memory for previously encountered items will provide insight into capacity limits and the role of retroactive interference. If infants can maintain more than 2 items long enough to detect the novel item during test, then they may show novelty preference when tested with items 4, 5, and 6. However, if their capacity is limited to 2 items, and if the activities involved in perceiving and encoding item number 6 causes interference with their memory for previously encountered items, infants’ memory for the fourth item will be impaired by the inclusion of this retention delay. Infants’ memory for the fifth item should be unaffected.
Experiment 2
Method
Participants
The final sample consisted of 72 healthy, full-term 6-month-old infants (M = 191.72 days, SD = 9.03; 40 boys, 44 had pets at home). Eleven infants were tested at University of Iowa and 61 infants were tested at UC Davis; none had participated in Experiment 1. Infants were recruited as described in Experiment 1. Forty-five infants were White and not Hispanic, 5 were Asian, 2 were Native Hawaiian or Pacific Islander, 1 infant was African American, 8 were of mixed race, 10 infants (who were either White or did not have race reported) were Hispanic, and 1 did not have race reported. One mother had completed eighth grade; all the other mothers had completed high school and all but 2 had at least some college.
An additional 16 infants were tested (14 at UCDavis), but were excluded from the final analysis because they fussed and/or refused to look at the monitors (n = 10), experimenter error (n = 3), having novelty preference scores more than 2.25 SD different from the overall mean (n = 2), or the parent interfered (n = 1).
Stimuli, apparatus, and procedure
The stimuli, apparatus, and procedure were the same as in Experiment 1 except that following the 6 familiarization trials infants received a single 15-s trial with a blank screen. From the infants’ perspective, all that happened during this period was the attention-getter stopped blinking for 15 s—there was no change in the stimulus monitors. Thus, any signal that this was a delay period was weak. This unfilled delay between the last familiarization and test allowed additional time for storage and consolidation of the last item without introducing a new item that would cause additional retroactive interference. This 15-s delay plus the inter-trial intervals before and after the delay resulted in a retention period of 18.48 s on average (SD = 1.99) between the end of the sixth familiarization trial and start of the first test trial. The familiar test was the sixth item for 24 infants, the fifth item for 24 infants, and the fourth item for 24 infants.
Reliability was assessed for 20 infants’ sessions. Agreement for the duration of looking on each trial between the original on-line observer and a second observer recording looking time off-line from recordings of the infants’ session was high; average r = .97, and average difference between the coders for each trial was .42 s.
Results
Responding during familiarization
Infants’ looking during each familiarization trial is presented in Table 1. The looking is similar to that exhibited by infants in Experiment 1. An ANOVA conducted on infants’ looking during each trial revealed a significant effect of trial, F(5, 345) = 2.99, p = .01, ηp2 = .04. The linear trend was highly significant, F(1, 69) = 8.74, p = .004, ηp2 = .11. Comparisons of the individual means with two-tailed t-tests, determining significance using FDR (Benjamini & Hochberg, 1995), revealed that infants looked longer on trial 1 than on trial 4, t(71) = 2.65, p = .01, d = .31, trial 5, t(71) = 2.60, p = .01, d = .31 and trial 6, t(71) = 2.41, p = .02, d = .28. Infants looked marginally longer on trial 2 than on trials 4, t(71) = 2.27, p = .03, d = .27, and 5, t(71) = 2.14, p = .04, d = .25. None of the other comparisons were significant.3
Interestingly, infants in this sample did show a decrease in looking from the first to the last trial, creating the potential problems of interpretation described in Experiment 1. However, although infants did decrease significantly their looking from trial 1 to trial 6, the magnitude of the decrease was modest (overall approximately a 13% decrease in looking, as compared the standard 50% decrease in looking required in habitation tasks, Oakes, 2010). Thus, this decrease, although greater decrease than in Experiment 1 (which was approximately 7% decrease), does not seem to reflect a dramatic decrease in processing time as would a 50% decrement in looking. It is nonetheless possible that general habituation resulted in infants in Experiment 2 engaging in less processing of the items encountered late in the series, and their novelty preferences may be less robust, consistent with findings that differences in familiarization time are related to infants’ relative preference for the novel or familiar item (Roder, et al., 2000; Rose, et al., 1982). Because the quantitative aspects of this relation have not been precisely worked out, it is not clear whether or how much weaker the novelty preference might be in Experiment 2 relative to Experiment 1.
This logic may particularly apply to the sixth item, as in this sample, infants’ looking to the sixth item was relatively low—in fact, infants who were eventually tested with the sixth item, looked less on that familiarization trial than any group of infants in Experiment 1. This low level of looking might lead to the prediction that infants would show poor memory for the last item. But, comparison of the two groups of infants eventually tested on the sixth item showed that the difference in looking on the sixth familiarization trial in Experiments 1 and 2 was not significant, t(34) = 1.44, p = .16, d= .51. And, the inclusion of short retention period following the sixth familiarization trial provided the infants in Experiment 2 additional time to continue the storage processes for that item, even if they studied it for a shorter duration. Indeed, Catherwood and her colleagues (1996) found that infants’ memory was influenced by total time to store and consolidate the item; not just the amount of time stimuli were visible for study. Thus, infants’ novelty preference for this sixth item will presumably reflect infants total amount of processing, and not solely the duration of looking when the item was visible.
Responding during test
The primary analysis was whether infants responded to the novel item more than would be expected by chance. The novelty preference for each group in this experiment is on the far right of Figure 2. The results for infants tested with the fourth and sixth items are strikingly different from the corresponding groups from Experiment 1; in contrast to the first experiment, infants in Experiment 2 tested with the sixth familiarization item showed a significant novelty preference, t(23) = 2.10, p = .05, d = .43, and infants in Experiment 2 tested with the fourth familiarization items, failed to significantly prefer the novel item, t(23) = .97, p = .34, d = .20. Infants tested with the fifth item responded essentially the same as did infants in Experiment 1; they had a significant preference for the novel item over that familiar item, t(23) = 2.43, p = .02, d = .50.
To more fully determine the effect of the delay on infants’ responding, we conducted additional analyses comparing responding by these groups of infants, as well as comparing infants in Experiments 1 and 2. First, to confirm our impression that the delay influenced infants’ responding to the fourth and sixth item, we entered into an ANOVA the novelty preference scores for these two groups of infants and the infants in Experiment 1 tested with the fourth familiarization item and those tested with the sixth familiarization item with Experiment (1 versus 2) and Serial Position (4 versus 6) as between subjects factors. This analysis revealed the expected significant Experiment by Serial Position interaction, F (1, 68) = 4.22, p = .04, ηp2 = .06, indicating that the delay had a different influence on infants’ memory for the items in each serial position. Note that both groups tested on the sixth item had no intervening items between study and test, and both groups tested on the fourth item had two intervening items; these groups primarily differed in the inclusion of a delay between the sixth item and test. Clearly, this delay—and the storage and consolidation processing of the sixth item during that delay as well as any resulting retroactive interference effects of that processing—contributed to the strength of infants’ novelty preference when tested on either the fourth or sixth familiarization item. Direct comparisons of infants’ responding in Experiments 1 and 2 when tested on the fifth item revealed no significant difference in their Novelty Preference score, t(34) = .52, p = .61, d = .18, suggesting that including the delay did not negatively influence infants’ memory for that item.
The question we next asked was how is infants’ memory influenced by delay alone. That is, the design of these two experiments afforded us the opportunity to compare responding by infants whose memory test occurred after approximately the same temporal delay, but that differed in terms of the number of intervening items. Infants in Experiment 1 tested with the third item did not differ from infants in Experiment 2 tested on the fourth item, t(34) = .21, p = .83, d = .06; infants in Experiment 1 tested with the fourth item did not differ from infants in Experiment 2 who were tested on the fifth item, t(34) = 1.18, p = .24, d = .42; and infants in Experiment 1 tested with the fifth item did not differ from infants in Experiment 2 tested on the sixth item, t(34) = .61, p = .54, d = .22. Thus, infants’ novelty preference was similar when the time between study and test was the same regardless of the number of intervening items.
Discussion
Experiment 2 showed that infants’ responding when tested on their memory for the fourth, fifth, or sixth items presented during familiarization was influenced by the inclusion of a retention delay of approximately 18 s between the sixth item and the test trials (15 s delay period, plus inter-trial-intervals). In Experiment 2, infants showed better memory for the sixth item and worse memory for the fourth item than did infants in Experiment 1. Infants’ memory for the fifth item was uninfluenced by the inclusion of a delay. The combined pattern of results from Experiments 1 and 2 suggests that storage and consolidation processes play an important role in infants’ visual recognition memory.
Experiment 3
There is still one basic assumption that we have not tested: that infants actually encoded, consolidated, and stored the first items encountered, and those items were “forgotten” as new items were encoded, consolidated and stored. That is, we assume that infants’ failure in Experiment 1 to show a novelty preference when tested with the first 3 items reflects the processes of retroactive interference.
However, it is possible that infants failed to show a novelty preference for those first items because they never encoded them. That is, infants may not encode, consolidate and store the first items they encounter in this task because they have not yet adopted that strategy. Because we cannot instruct infants about our goals or their task, that infants simply may have failed to engage in memory processes on the first few trials.
We think this explanation unlikely for several reasons. First, we observed that infants did show memory for item number 4 in Experiment 1 but not in Experiment 2, suggesting the effects of retroactive interference in Experiment 2 when the final item in the list was more fully processed. Second, many studies have shown robust memory for a singly presented item by 5- to 7-month-old infants (see Fagan, 1990; Rose, et al., 2004 for reviews), suggesting that infants do begin to engage memory processes from the start of this type of procedure.
Nevertheless, it is possible that infants in our task with these stimuli failed to engage those processes at the start of the task. Thus, to confirm that infants do in fact begin to encode and store items from the start of the session, in Experiment 3 we used the procedure in Experiment 2 (e.g., familiarization followed by a brief delay period followed by a test), but assessed infants’ novelty preference when only a single item was presented on one trial during familiarization. Note that in this experiment, because infants were presented with only a single item during familiarization, we are assessing infants’ visual recognition memory in a context that likely did not engage categorization. Thus, in this experiment, infants did not need to form a robust and stable memory for the item while at the same time making comparisons among items and representing commonalities. However, this was also true when infants in Experiments 1 and 2 encountered the first item in the sequence—it was not until they had encountered and begun to process the second item that the processes of comparison and extracting commonalities would be induced. Thus, Experiment 3 mimics the conditions under which infants would have encoded and retained information about the first item in the familiarization series of Experiments 1 and 2.
Method
Participants
The final sample was 24 6-month-old infants (M age = 192.38 days, SD = 9.61; 10 boys) who had not participated in Experiments 1 or 2, recruited as described earlier. Nine infants were tested at University of Iowa, the remaining infants were tested at UC Davis. Twenty infants were White and not Hispanic, 2 infants were mixed race, and 2 infants did not have race reported (one was Hispanic). All mothers had graduated high school and 19 had earned a bachelor’s degree or higher. An additional 7 infants (4 at UC Davis) were tested but excluded from the final analyses because of fussiness or refusing to look at the stimuli (n = 2) or experimenter error (n = 5).
Stimuli, apparatus, procedure
All aspects of the stimuli, apparatus, and procedure were identical to that described earlier, except infants received only one familiarization trial. Following familiarization, there was a short retention delay (M = 18.32 s, SD = 2.08), as in Experiment 2, and then two test trials in which the now familiar item was paired with a novel item. Reliability assessed on 7 infants was high; average correlation between the two coders for the duration of looking was .92, with a mean difference in looking recorded of .55 s.
Results and Discussion
Infants’ looking during their single familiarization trial is presented in Table 1. Infants’ looking on this trial was similar to the looking during the first trial by infants in the other experiments. Evaluation of infants’ novelty preference scores during test revealed that the mean novelty preference score in this experiment, M = .57, SD = .14, was significantly greater than chance, t(23) = 2.22, p = .04, d = .45. In addition, their score was marginally greater than the infants tested with the first item in Experiment 1, t(34) = 1.47, p = .07, d = .52 (this comparison should be considered with caution given the large number of differences between the familiarization experience between the two groups of infants). Thus, when familiarized with a single item, infants show a preference for novelty during a test that occurs approximately 18 s after the end of the study period. Thus, the failure of infants in Experiment 1 to recognize the first items in the list likely does reflect the effects of the interference effects of encoding and retaining multiple additional items while simultaneously categorizing those items; the failure of infants in Experiment 1 to recognize the first item in the series does not appear to be due to infants simply not encoding the first items encountered.
General Discussion
These results provide understanding into infants’ memory for individual images of cats and dogs presented over a series of trials. Importantly, we provide evidence not only that infants can remember these individual items, but we also provide insight into the processes of infants’ visual recognition memory, particularly when memory and categorization processes are both engaged. These contributions will be discussed in detail in the following sections.
Implications for memory processes in infancy
Our results show that infants can encode and remember multiple individual items in a list, presumably while simultaneously categorizing those items. Thus, these results add to our understanding of how visual recognition memory, working memory, and categorization interact. We found that memory processes observed in adulthood are evident in infancy. For example, like adults, infants’ memory for individual items in a list is influenced by the serial position of those items in the list. However, we did not observe straightforward primacy and recency effects, as are often found in adults’ recall and recognition (Roediger, et al., 2010). Moreover, the position effects were different from the few other studies reported in the literature examining such effects in infants’ visual recognition memory (Cornell & Bergstrom, 1983; Rose, et al., 2001). In some ways, the fact that studies employing different experimental procedures and stimuli revealed different position effects is not surprising given that even for adults serial position effects are a function of factors such as the timing of item presentation and characteristics of the to-be-remembered items (Neath, 1993a). For example, the different position effects we observed in our experiments may be due to differences in temporal distinctiveness, which contributes to serial position effects in adults (Neath & Crowder, 1996). The temporal gap in Experiment 2 may have made the sixth item temporally distinct, allowing infants to remember that item in Experiment 2 but not Experiment 1. However, the overall pattern of results are not easily explainable by temporal distinctiveness: infants in Experiment 1 remembered the fourth and fifth item, and infants in Experiment 2 remembered the fifth item, and none of these items were temporally distinct.
Our results also indicate that infants’ memory in this context is influenced by the activities of maintaining previously learned information can interfere with the learning of new information (proactive interference) and the activity of learning new information can interfere with maintaining previously learned information (retroactive interference), as has been observed in adults (see Roediger, et al., 2010). Retroactive interference explains why infants were unable to maintain a memory for the first encountered items as they perceived, encoded, and stored the last three items in the list. In Experiment 2, infants’ formation of memories for the fifth and sixth item may have similarly interfered with their memory for the fourth item. This analysis depends on infants having actually formed a memory for the first several items presented. Experiment 3 showed that infants formed memories for at least the first item presented, and the results of Experiment 1 suggest that when that item was followed by a series of other items, they “forgot” that first item as they encoded and remembered the later encountered items and categorized all the items.
The fact that infants in Experiments 1 and 2 responded differently when tested on the sixth item suggests that their memory is also influenced by proactive interference. In Experiment 1, infants showed ambiguous (at best) memory for the last item, perhaps due to proactive interference—that is, the activities involved in maintaining representations of the fourth and fifth items may have caused confusion with the last item, resulting in ambiguous responding. Indeed, the fact that infants better remembered the last item when a brief post-stimulus delay was included (Experiment 2) suggests that infants’ memory in this task is determined by the amount of time needed to fully process each individual item. Specifically, consolidation and storage of the individual items apparently continued after the encoding period (i.e., the 15-s trial) had ended. When infants encountered a new item (e.g., item number 6), the continued processing of the previous item (in this case number 5) interfered with the processing of the current item.
The point is memory processes that occur over time—both during encoding and after the item is no longer available—contribute to the stability of memory, and how robustly infants prefer a novel stimulus. In Experiment 1, the brief time between exposure to the sixth item and the test of infants’ memory did not allow them to fully consolidate and store that item. In Experiment 2, increasing the time between exposure to the sixth item and the memory test with that item was sufficient for more complete processing, This general reasoning is consistent with the finding that adults’ memories become stronger and more resilient over time (McGaugh, 2000). Note that the incomplete memory processes with respect to item 6 in Experiment 1 did not interfere with the maintenance of infants’ memory for the fourth and fifth items, but the complete memory processes with respect to that item in Experiment 2 did interfere with the memory for the fourth item. Thus, infants’ memory in this task seems to be a function of both consolidation and interference. Indeed, in general, retroactive interference has been thought to result from interfering with the process of consolidating memories (McGaugh, 1999). In the present results, retroactive interference appears to have occurred when infants had formed a stable memory for items later in the list—that is, items for which they showed a robust novelty preference.
This interactive effect of consolidation and retroactive interference on infants’ memory is consistent with the extant literature on the effect of interference on infants’ memory. Three studies published in 1977 suggested that intervening distracting stimuli interfered with infants’ memory of previously studied items only if infants encoded and processed the distractors (Cohen, et al., 1977; Fagan, 1977; McCall, et al., 1977). Little or no interference was observed for infants who did not seem to learn those distracting stimuli. Thus, intervening stimuli can influence infants’ memory for previously studied stimuli if infants encode and retain those intervening stimuli in memory. The present results are consistent with this finding: when infants had weak memory of item 6, it did not interfere with the memory for item 4 (Experiment 1), but when they fully processed item 6, this processing interfered with the memory for item 4 (Experiment 2).
Understanding visual recognition memory in the context of categorization
These experiments also add to our understanding of infants’ visual recognition memory because they uncover memory processes in the context of categorization. That is, many previous studies have examined infants’ memory for a single item presented during familiarization (see Fagan, 1990; and Rose, et al., 2004 for reviews). Here, we examined visual recognition memory for a series of categorized images. That is, previous studies examined provided a more “pure” test of visual recognition memory, much like the present Experiment 3. In previous studies, and Experiment 3, in which infants were familiarized with only a single image, the task induced visual recognition and working memory processes, but not categorization processes. We used a task, in contrast, that has been shown to induce categorization in infants between 4 and 6 months of age (Kovack-Lesh, et al., 2008; Quinn, et al., 2002). Not only did infants in our task need to encode, consolidate, and maintain multiple individual items, they did so while comparing those items and detecting and representing commonalities among the items. The few other studies that have examined infants’ visual recognition memory for a series of items involved unrelated items (Cornell & Bergstrom, 1983; Rose, et al., 2001). These studies, therefore, induced processing of multiple items, but not categorization. Without a direct comparison it is impossible to know how the category relatedness of the to-be-remembered items contributes to infants’ performance, but it is likely that the infants in our study were engaged not only in memory processes but also categorization processes. As a result, encoding and storing a stable representation of any of the items in the list may have taken more time that if the infants had not been categorizing. Moreover, interference may have resulted from either categorization or memory processes, or a combination of the two processes. Future studies may examine how lists of categorized and non-categorized lists influence infants’ novelty preference, examining, for example, whether like adults, infants better remember individual items when the items can be categorized (Howe, 2004; Medin & Bettger, 1994).
We focus here on the fact that we have studied infants’ memory in a categorization task, whereas previous studies did not. However, it is important to point out that there are many differences between studies that may contribute to differences in results. For example studies differ in the stimuli, the way the stimuli are presented, and the delay between study and test. Any of these differences may contribute to differences in findings. Consider, for example, the fact that infants typically have been reported to show better memory for immediate tests than for delayed tests (Fagan, 1990), and we observed in contrast infants show best memory when there was a brief delay between encoding and test, even if that delay was filled with the presentation of another item. However, these different studies had different definitions of “immediate” tests. Due to technical differences in the procedures used, in our study the delay on the immediate test was likely shorter than in previous studies (i.e., here less than 3 s, in studies using cards positioned in a Fagan testing chamber the delay may be between 3 and 10 s). Given that Experiment 2 revealed that even small differences in timing can yield different results, it would not be surprising if features of the task that caused greater inter-trial-intervals or slightly longer delays between study and test did contribute to the pattern of responding observed.
However, the fact that apparently minor technical differences can yield different results should not be taken to mean that no study can provide insight into general memory processes. Rather, by systematically studying of the effects of such differences we gain insight into those memory processes. Indeed, the literature yields several generalizations about infants’ memory. For example, across a wide range of studies, using a variety of stimuli and different means of presenting those stimuli, infants in the first year show evidence of forming memories of items that they visually inspect for 15 to 30 s, whether they are familiarized with only a single item (Fagan, 1990; Rose, et al., 2004) or with multiple items (Kovack-Lesh, et al., 2008; Quinn, et al., 2002; Rose, et al., 2001). Second, infants’ memory processes occur over time—novelty preferences are more robust when infants have longer study times (Rose, et al., 1982), and the present work suggests that those processes need not occur in the presence of the to-be-remembered item (see also Catherwood, et al., 1996). Finally, visual recognition memory processes do not occur in isolation, but rather can be engaged as infants use other cognitive processes, such as categorization (Kovack-Lesh, et al., 2008; Quinn, et al., 2002). Thus, studies like the one presented here are an important step in solving the “Humpty Dumpty” problem in the study of infant cognition (Oakes, 2009), and help us understand how cognitive processes work together.
Conclusions
In summary, the present results provide additional understanding into infants’ the processes of visual recognition memory. In particular, we show how the processes of remembering individual items and categorizing related items influences infants’ memory for individual items. The results showed that, as in adults, these processes in infancy are intertwined, and not only does exposure to intervening items interfere with infants’ maintenance of memories of previously encountered items, but interference depends crucially on how much the interfering item has been proceed. One of the difficulties of studying infant cognition is that infant responding seems to depend crucially on features of the stimuli, how the stimuli are presented, and procedural differences. The present results provide an important step in understanding how contextual variations influence the processes of visual recognition memory in infancy.
Acknowledgments
This research and preparation of this manuscript were made possible by NIH grants HD49840 and HD56018 awarded to LMO. Some of these results were reported at the biennial meeting of the Society for Research in Child Development, March 2007, Boston, and the biennial meeting of the Cognitive Development Society, October 2007, Santa Fe, NM. We thank Shaena McGivern, Lisa Christoffer, Emily Spring, and the undergraduate students in the Infant Cognition Laboratories at the University of Iowa and the University of California, Davis, for their help with this project.
Footnotes
It is always possible that we would have obtained different results if we had used different stimuli. However, we used the same stimuli and test pairs across conditions and across the three experiments so we are confident that any differences across experiments we observe are due to differences in the variables of interest, and not due to differences in stimuli.
This lack of an effect of pet experience on infants’ responding may seem surprising given recent results reported by Kovack-Lesh and colleagues (2008), but differences in the age of the infants, stimulus presentation format, and the number of different stimuli shown each infant may contribute to the different findings with respect to pet ownership.
Although the linear trend was not significant in Experiment 1, but it was in Experiment 2, inspection of the means provided in Table 1 shows that responding over familiarization in the two experiments was very similar. Likely as a result of random fluctuation, infants in Experiment 1 had relatively low looking on trial 2, making the overall trend look different from that in Experiment 2. But the patterns are more similar than different.
References
- Alvarez GA, Cavanagh P. The capacity of visual short-term memory is set by total information load and by number of objects. Psychological Science. 2004;15:106–111. doi: 10.1111/j.0963-7214.2004.01502006.x. [DOI] [PubMed] [Google Scholar]
- Ashby FG, O’Brien JB. Category learning and multiple memory systems. Trends in Cognitive Sciences. 2005;9:83–89. doi: 10.1016/j.tics.2004.12.003. [DOI] [PubMed] [Google Scholar]
- Bahrick L, Hernandez-Reif M, Pickens J. The effect of retrieval cues on visual preferences and memory in infancy: Evidence for a four-phase attention function. Journal of Experimental Child Psychology. 1997;67:1–20. doi: 10.1006/jecp.1997.2399. [DOI] [PubMed] [Google Scholar]
- Bahrick L, Pickens JN. Infant memory for object motion across a period of three months: Implications for a four-phase attention function. Journal of Experimental Child Psychology Special Issue: Early memory. 1995;59:343–371. doi: 10.1006/jecp.1995.1017. [DOI] [PubMed] [Google Scholar]
- Benjamini Y, Hochberg Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. Journal of the Royal Statistical Society Series B (Methodological) 1995;57:289–300. [Google Scholar]
- Catherwood D, Skoien P, Green V, Holt C. Assessing the primary moments of infant encoding of compound visual stimuli. Infant Behavior & Development. 1996;19:1–11. [Google Scholar]
- Cohen LB, Atkinson DJ, Chaput HH. Habit 2000: A new program for obtaining and organizing data in infant perception and cognition studies. (Version 1.0) [Computer Software] Austin TX: The University of Texas; 2000–2002. [Google Scholar]
- Cohen LB, DeLoache JS, Pearl RA. An examination of interference effects in infants’ memory for faces. Child Development. 1977;48:88–96. [PubMed] [Google Scholar]
- Cohen LB, Menten TG. The rise and fall of infant habituation. Infant Behavior & Development. 1981;4 [Google Scholar]
- Cornell EH, Bergstrom LI. Serial-position effects in infants’ recognition memory. Memory & Cognition. 1983;11:494–499. doi: 10.3758/bf03196986. [DOI] [PubMed] [Google Scholar]
- Courage ML, Howe ML. Long-term retention in 3.5-month-olds: Familiarization time and individual differences in attentional style. Journal of Experimental Child Psychology. 2001;79:271–293. doi: 10.1006/jecp.2000.2606. [DOI] [PubMed] [Google Scholar]
- Dewar M, Garcia YF, Cowan N, Della Sala S. Delaying interference enhances memory consolidation in amnesic patients. Neuropsychology. 2009;23:627–634. doi: 10.1037/a0015568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dewhurst S. Category Repetition and False Recognition: Effects of Instance Frequency and Category Size* 1. Journal Of Memory And Language. 2001;44:153–167. [Google Scholar]
- Dudai Y. The neurobiology of consolidations, or, how stable is the engram. Annual Review of Psychology. 2004;55:51–86. doi: 10.1146/annurev.psych.55.090902.142050. [DOI] [PubMed] [Google Scholar]
- Fagan JF. Infant recognition memory: Studies in forgetting. Child Development. 1977;48:68–78. doi: 10.1111/j.1467-8624.1977.tb04244.x. [DOI] [PubMed] [Google Scholar]
- Fagan JF. The paired-comparison paradigm and infant intelligence. Annals of the New York Academy of Sciences. 1990;608:337–364. doi: 10.1111/j.1749-6632.1990.tb48902.x. [DOI] [PubMed] [Google Scholar]
- Furrer SD, Younger BA. Beyond the distributional input? A developmental investigation of asymmetry in infants’ categorization of cats and dogs. Developmental Science. 2005;8:544–550. doi: 10.1111/j.1467-7687.2005.00446.x. [DOI] [PubMed] [Google Scholar]
- Gulya M, Galluccio L, Wilk A, Rovee-Collier C. Infants’ long-term memory for a serial list: Recognition and reactivation. Developmental Psychobiology. 2001;38:174–185. [PubMed] [Google Scholar]
- Hardt O, Einarsson Eñ, Nader K. A bridge over troubled water: Reconsolidation as a link between cognitive and neuroscientific memory research traditions. Annual Review of Psychology. 2009;61:141–167. doi: 10.1146/annurev.psych.093008.100455. [DOI] [PubMed] [Google Scholar]
- Hay DC, Smyth MM, Hitch GJ, Horton NJ. Serial position effects in short-term visual memory: A SIMPLE explanation? Memory & Cognition. 2007;35:176–190. doi: 10.3758/bf03195953. [DOI] [PubMed] [Google Scholar]
- Howard MW, Venkatadass V, Norman KA, Kahana MJ. Associative processes in immediate recency. Memory & Cognition. 2007;35:1700–1711. doi: 10.3758/bf03193503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howe ML. The role of conceptual recoding in reducing children's retroactive interference. Developmental Psychology. 2004;40:131–139. doi: 10.1037/0012-1649.40.2.131. [DOI] [PubMed] [Google Scholar]
- Hunt RR, Lamb CA. What Causes the Isolation Effect? Learning Memory. 2001;27:1359–1366. [PubMed] [Google Scholar]
- Kaldy Z, Leslie AM. A memory span of one? Object identification in 6.5-month-old infants. Cognition. 2005;97:153–177. doi: 10.1016/j.cognition.2004.09.009. [DOI] [PubMed] [Google Scholar]
- Knoedler AJ, Hellwig KA, Neath I. The shift from recency to primacy with increasing delay. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1999;25:474–487. [Google Scholar]
- Knowlton BJ, Squire LR. The learning of categories: Parallel brain systems for item memory and category knowledge. Science. 1993;262:1747–1749. doi: 10.1126/science.8259522. [DOI] [PubMed] [Google Scholar]
- Kovack-Lesh KA, Horst JS, Oakes LM. The cat is out of the bag: The joint influence of previous experience and looking behavior on infant categorization. Infancy. 2008;13:285–307. [Google Scholar]
- Love BC, Gureckis TM. Models in search of a brain. Cognitive, Affective, and Behavioral Neuroscience. 2007;7:90–108. doi: 10.3758/cabn.7.2.90. [DOI] [PubMed] [Google Scholar]
- Makovski T, Jiang YV. Proactive interference from items previously stored in visual working memory. Mem Cognit. 2008;36:43–52. doi: 10.3758/mc.36.1.43. [DOI] [PubMed] [Google Scholar]
- McCall RB, Kennedy CB, Dodds C. The interfering effect of distracting stimuli on the infant’s memory. Child Development. 1977;48:79–87. [PubMed] [Google Scholar]
- McGaugh JL. Time-dependent processes in memory storage. Science. 1966;153:1351–1358. doi: 10.1126/science.153.3742.1351. [DOI] [PubMed] [Google Scholar]
- McGaugh JL. The perseveration-consolidation hypothesis: Mueller and Pilzecker, 1900. Brain Research Bulletin Special Issue: Highlights of twentieth century neuroscience. 1999;50:445–446. doi: 10.1016/s0361-9230(99)00126-4. [DOI] [PubMed] [Google Scholar]
- McGaugh JL. Memory: A century of consolidation. Science. 2000;287:248–251. doi: 10.1126/science.287.5451.248. [DOI] [PubMed] [Google Scholar]
- Medin D, Bettger J. Presentation order and recognition of categorically related examples. Psychonomic Bulletin & Review. 1994;1:250–254. doi: 10.3758/BF03200776. [DOI] [PubMed] [Google Scholar]
- Morgan K, Hayne H. The effect of encoding time on retention by infants and young children. Infant Behavior and Development. 2006;29:599–602. doi: 10.1016/j.infbeh.2006.07.009. [DOI] [PubMed] [Google Scholar]
- Murdock BB. The retention of individual items. Journal of Experimental Psychology. 1961;62:618–625. doi: 10.1037/h0043657. [DOI] [PubMed] [Google Scholar]
- Nairne J. Remembering over the short-term: The case against the standard model. Annual Review of Psychology. 2002:53–82. doi: 10.1146/annurev.psych.53.100901.135131. [DOI] [PubMed] [Google Scholar]
- Neath I. Contextual and distinctive processes and the serial position function. Journal of Memory and Language. 1993a;32:820–840. [Google Scholar]
- Neath I. Distinctiveness and serial position effects in recognition. Memory & Cognition. 1993b;21:689–698. doi: 10.3758/bf03197199. [DOI] [PubMed] [Google Scholar]
- Neath I, Crowder RG. Distinctiveness and very short-term serial position effects. Memory. 1996;4:225–242. doi: 10.1080/09658211.1996.9753032. [DOI] [PubMed] [Google Scholar]
- Nomura EM, Maddox WT, Filoteo JV, Ing AD, Gitelman DR, Parrish TB, Reber PJ. Neural correlates of rule-based and information-integration visual category learning. Cerebral cortex (New York, NY : 1991) 2007;17:37–43. doi: 10.1093/cercor/bhj122. [DOI] [PubMed] [Google Scholar]
- Nosofsky RM, Johansen MK. Exemplar-based accounts of "multiple-system" phenomena in perceptual categorization. Psychonomic Bulletin & Review. 2000;7:375–402. [PubMed] [Google Scholar]
- Oakes LM. Concepts and categorization skills. In: Haith MM, Benson J, editors. Encyclopedia of infant and early child development. Vol. 1. San Diego: Academic Press; 2008. pp. 249–259. [Google Scholar]
- Oakes LM. The "Humpty Dumpty Problem" in the Study of Early Cognitive Development: Putting the Infant Back Together Again. Perspectives on Psychological Science. 2009;4:352–358. doi: 10.1111/j.1745-6924.2009.01137.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oakes LM. Using Habituation of Looking Time to Assess Mental Processes in Infancy. Journal of Cognition & Development. 2010;11:255–268. doi: 10.1080/15248371003699977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oakes LM, Ribar RJ. A comparison of infants’ categorization in paired and successive presentation familiarization tasks. Infancy. 2005;7:85–98. doi: 10.1207/s15327078in0701_7. [DOI] [PubMed] [Google Scholar]
- Palmeri TJ, Flanery MA. Learning about Categories in the Absence of Training: Profound Amnesia and the Relationship between Perceptual Categorization and Recognition Memory. Psychological Science. 1999;10:526–530. [Google Scholar]
- Posner MI, Konick AF. On the role of interference in short-term retention. J Exp Psychol. 1966;72:221–231. doi: 10.1037/h0023458. [DOI] [PubMed] [Google Scholar]
- Postman L, Keppel G. Retroactive inhibition in free recall. J Exp Psychol. 1967;74:203–211. doi: 10.1037/h0024592. [DOI] [PubMed] [Google Scholar]
- Quinn PC. Development of Subordinate-Level Categorization in 3- to 7-Month-Old Infants. Child Development. 2004;75:886–899. doi: 10.1111/j.1467-8624.2004.00712.x. [DOI] [PubMed] [Google Scholar]
- Quinn PC, Eimas PD. Young infants’ use of facial information in the categorical differentiation of natural animal species: The effect of inversion. Infant Behavior and Development. 1996;19:381–384. [Google Scholar]
- Quinn PC, Eimas PD, Rosenkrantz SL. Evidence for repesentations of perceptually similar categories by 3-month-old and 4-month-old infants. Perception. 1993;22:463–475. doi: 10.1068/p220463. [DOI] [PubMed] [Google Scholar]
- Quinn PC, Yahr J, Kuhn A, Slater AM, Pascalis O. Representation of the gender of human faces by infants: A preference for female. Perception. 2002;31:1109–1121. doi: 10.1068/p3331. [DOI] [PubMed] [Google Scholar]
- Roder BJ, Bushnell EW, Sasseville AM. Infants’ preferences for familiarity and novelty during the course of visual processing. Infancy. 2000;1:491–507. doi: 10.1207/S15327078IN0104_9. [DOI] [PubMed] [Google Scholar]
- Roediger HL, Weinstein Y, Agarwal PK. Forgetting: Preliminary considerations. In: Della Sala S, editor. Forgetting. New York: Psychology Press; 2010. pp. 1–22. [Google Scholar]
- Rose SA. Developmental changes in infants’ retention of visual stimuli. Child Development. 1981;52:227–233. [PubMed] [Google Scholar]
- Rose SA, Feldman JF, Jankowski JJ. Visual short-term memory in the first year of life: Capacity and recency effects. Developmental Psychology. 2001;37:539–549. doi: 10.1037//0012-1649.37.4.539. [DOI] [PubMed] [Google Scholar]
- Rose SA, Feldman JF, Jankowski JJ. Infant visual recognition memory. Developmental Review. 2004;24:74–100. doi: 10.1037/0012-1649.39.3.563. [DOI] [PubMed] [Google Scholar]
- Rose SA, Gottfried AW, Mello-Carmina P, Bridger WH. Familiarity and novelty preferences in infant recognition memory: Implications for information processing. Developmental Psychology. 1982;18:704–713. [Google Scholar]
- Rose SA, Jankowski JJ, Feldman JF. Speed of processing and face recognition at 7 and 12 months. Infancy. 2002;3:435–455. [Google Scholar]
- Rose SA, Jankowski JJ, Feldman JF. The inversion effect in infancy: The role of internal and external features. Infant Behavior & Development. 2008;31:470–480. doi: 10.1016/j.infbeh.2007.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross-Sheehy S, Oakes LM, Luck SJ. The development of visual short-term memory capacity in infants. Child Development. 2003;74:1807–1822. doi: 10.1046/j.1467-8624.2003.00639.x. [DOI] [PubMed] [Google Scholar]
- Surprenant AM, Neath I. The 9 lives of short-term memory. In: Thorn A, Page M, editors. Interactions between short-term and long-term memory in the verbal domain. Hove, UK: Psychology Press; 2009. pp. 16–43. [Google Scholar]
- Tan L, Ward G. A recency-based account of the primacy effect in free recall. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2000;26:1589–1625. doi: 10.1037//0278-7393.26.6.1589. [DOI] [PubMed] [Google Scholar]
- Younger BA. The segregation of items into categories by 10-month-old infants. Child Development. 1985;56:1574–1583. [PubMed] [Google Scholar]
- Zaki SR, Nosofsky RM. A single-system interpretation of dissociations between recognition and categorization in a task involving object-like stimuli. Cognitive, Affective, and Behavioral Neuroscience. 2001;1:344–359. doi: 10.3758/cabn.1.4.344. [DOI] [PubMed] [Google Scholar]