

Abstract
BACKGROUND
Longer periods of recovery reduce the likelihood of relapse, which may be due to a reduced ability of various stimuli to occasion alcohol or drug seeking. However, this hypothesis remains largely uninvestigated.
METHODS
Here we assessed the ability of intermediate stimuli to occasion responding for ethanol in rats trained to discriminate an 8kHz tone signaling a food fixed-ratio (FR) of 5 and an ethanol FR5, from a 16kHz tone signaling a food FR150 and ethanol FR5. In the presence of the 8kHz tone responding for food predominates, and in the presence of the 16 kHz tone, responding for ethanol predominates.
RESULTS
In the context of alternation between these conditions, varying the tone from 8 to 16kHz produces a graded increase in ethanol (versus food) responding, consistent with a stimulus generalization function. A recent history of responding under food-predominant choice conditions, either during the test session or in the four sessions that precede it shifts the generalization function downwards. Extending this history to nine sessions shifts the curve further downwards. The stimulus generalization function was similar in a separate group, trained with different relative ratios for food and ethanol, but with similar behavioral allocation under each discriminative stimulus. Finally, withholding access to food and ethanol for 4 or 16 sessions did not affect the stimulus generalization gradient.
CONCLUSION
These results suggest that longer histories of reinforced alternative behavior might reduce the likelihood of relapse by decreasing the control exerted over alcohol- or drug-seeking by stimuli similar to those that previously occasioned alcohol- or drug-seeking.
Keywords: addiction, stimulus control, choice, discrimination
1. INTRODUCTION
Alcoholism and other addictions are often characterized as a loss of control over drinking or drug use. This can be interpreted in two different ways (Keller, 1972). Loss of control can describe an inability to refrain from initiating a drinking bout when confronted with stimuli that have come to occasion drinking. The sight of a preferred bar or socializing with a particular group of friends can come to occasion initiation of drinking. It follows, then, that there is likely a range of stimuli or situations that might occasion initiation of a drinking bout, and over time the range of such stimuli might broaden beyond those that initially occasioned drinking (Schuster, 1986). Loss of control can also mean that once drinking starts, it does not stop until intoxication is so severe drinking cannot continue.
Recovery can be viewed as the converse of loss of control, so drinking may be moderate after initiation of a drinking bout. Likewise, a person in recovery may be able to abstain even in the presence of stimuli that had previously occasioned drinking (Bickel and Kelley, 1997). We postulate that as alternative behavior supplants drinking, the range of stimuli that occasion drinking diminishes. This may, in part, explain why for a variety of addictions including alcoholism (Gilpin et al., 1997; Gossop et al., 1990; Hunt et al., 1971; McKay et al., 2001) the probability of relapse diminishes as recovery lengthens.
Presumably, as stimuli that had occasioned heavy drinking lose their effectiveness, other, similar stimuli also lose their ability to occasion drinking. For example, early in recovery, walking by a bar may occasion a relapse to drinking, even if it is not the same bar where most of the problematic drinking had occurred. This is called stimulus generalization. Over time, the ability of such similar stimuli to occasion drinking may decline so that walking by a bar is no longer likely to precipitate relapse. Although this has been posited as a crucial mechanism in successful recovery, there is little data regarding this notion (Bickel and Kelley, 1997; Schuster, 1986). Procedures to study stimulus generalization are well-established in experimental psychology (Honig and Urcuioli, 1981). Subjects are trained to make different responses in the presence of distinct stimuli, A and B. Once this behavior is established, responding is assessed in the presence of varying stimuli that are intermediate between A and B. Typically, intermediate stimuli more similar to A result in responses that were trained in the presence of A, while those more similar to B result in responses that were trained in the presence of B. Experimental manipulations can then be performed and their effects on the stimulus generalization gradient observed.
Despite the conceptualization of addiction as a broadening of the stimuli that occasion drug use (Bickel and Kelley, 1997; Schuster, 1986), stimulus generalization has not been used to study stimulus control over drug-reinforced behavior. Further, stimulus generalization has not been used in situations where a choice between two different commodities (such as food and drug) is made.
Here, we provide concurrently available food and ethanol and, by changing the response requirement for food, produce food-predominant or ethanol-predominant responding. We then examine stimulus generalization in a situation where one stimulus occasions food-predominant responding and another occasions ethanol-predominant responding. With this baseline, we examine if the history of responding in only the food-predominant situation can decrease the ability of a range of stimuli to occasion responding for ethanol. First, we determine stimulus generalization functions in test sessions in which alternating or only food-predominant conditions are presented during reinforced components. We then test whether a history of four or nine preceding sessions in which only the food-predominant response conditions were presented affects the stimulus generalization function. We further determine whether changing the relative response requirements for food versus ethanol affect stimulus generalization. Finally, we examine whether a history of not responding for ethanol (or food, by removing the rats from the operant apparatus for the equivalent of 16 sessions) has a similar impact on the stimulus generalization function. Our choice of four, nine, and sixteen sessions was based on earlier studies using a similar procedure. In these studies, four and sixteen preceding sessions in which only food-predominant conditions are presented increased the amount of responding for food when rats were re-exposed to a stimulus signaling ethanol-predominant responding (Ginsburg and Lamb, submitted).
If longer periods of recovery reduce the risk of relapse by decreasing the range of stimuli that result in problematic use, we might expect that longer periods in which responding for ethanol does not occur will decrease the effectiveness of intermediate stimuli to occasion responding for ethanol. If this is due to an increase in the frequency of alternative behavior in the same context in which drug-use had occurred, a history of responding predominately for food should shift the stimulus generalization function. If, how ever, this is simply due to time spent not responding for ethanol, a similar shift should be seen after a period in which rats are prevented from responding for ethanol.
2. METHODS AND MATERIALS
2.1 General Materials and Methods
2.1.1 Subjects
Male, adult, singly housed Lewis rats (Harlan, Frederick, MD) served as subjects. All experiments included data from n=5 rats, except as indicated in Experiments 3 and 6 (Sections 2.4 and 2.7). Separate groups of rats were used for Experiments 1–4 and Experiments 5 and 6. Rats used in experiments 1–4 had previously been involved in studies related to resumption of responding for ethanol following extended periods of food -predominant responding under the same procedure (Ginsburg and Lamb, submitted). Upon arrival, rats weighed 260–265g and spent one week habituating to our facilities with food and water provided ad libitum. Subsequently, water was always available in the rats’ cages, however food was restricted to maintain body weights of 280–320 g for the remainder of the study (approximately 12–15g/day).
2.1.2 Apparatus
Training and testing occurred in a commercially available apparatus (Standard Rat Chamber, Med Associates, St. Albans, VT). On one wall of the chamber, two response levers were arranged horizontally, one on each side of the wall. Equidistant between the levers was a receptacle that provided access to 45 mg food pellets (Bio-Serve, Frenchtown, NJ) via a pellet dispenser and to a solution via a 0.1-ml dipper. Chambers also had a speaker connected to a tone generator (ANL- 926, Med Associates, St. Albans, VT) which produced pure tones that served as stimuli. Stimuli presentation and reinforcement delivery as well as data collection were accomplished by custom software written using a commercially available programming language (Med-PC, Med Associates, St. Albans, VT). Computer-generated pink noise was broadcast in the procedure room to mask ambient noise. Ethanol (190 proof) was obtained from Decon Labs, Inc. (King of Prussia, PA) and mixed with tap water to obtain a 10% (w/v) solution. Solutions were made fresh daily, and allowed to reach room temperature before being presented to the rats.
2.1.3 Training
Sessions were conducted on weekdays and were 30-min in duration. Rats were first trained to respond on the left lever for 8% sucrose solution in the presence of a 16 kHz, 80 dB tone. Upon completion of the response requirement, a dipper containing 0.1 ml of solution was raised and the 16 kHz tone was replaced with white noise at 80 dB. The dipper remained accessible for 30-s, at which point it returned to the inaccessible position, and white noise was replaced with the 16kHz tone. During this 30-s period, responses had no programmed consequences. During this portion of training, responses on the right lever had no programmed consequences. Once rats earned over 80 sucrose deliveries in a 30-min session (typically 2–7 sessions), the fixed-ratio (FR) was increased over a few sessions until rats were required to respond five times for a sucrose delivery. Subsequently, ethanol was introduced into the solution at 10% w/v, then sucrose was removed from the solution over the next 10–25 sessions so rats responded for 10% (w/v) ethanol solution in tap water.
Responding for food was trained in a subsequent 30-min session during presentation of a 8 kHz tone at 80dB. Under this stimulus condition, responses on the right lever resulted in the delivery of a 45 mg food pellet, and the tone changed to 0.1 kHz. Over the next several sessions, the FR was increased to 25, then rats were introduced to the multiple concurrent schedule.
Rats were trained to respond under a multiple concurrent schedule of food and ethanol reinforcement. In this schedule, components alternated randomly between a 16kHz tone and associated contingencies (food FR150, ethanol FR5 for experiments 1–4 and food FR75, ethanol FR5 for experiments 5 and 6) and an 8kHz tone and associated contingencies (food FR5, ethanol FR5 for experiments 1–4 and food FR25, ethanol FR5 for experiments 5 and 6). The order of components was randomized within each block of two components. Delivery of reinforcement was accompanied by a change in stimuli present in the chamber for 30-sec, which was followed by the next component. Sessions were 30-min in length.
2.1.4 Stimulus Generalization Test
Stimulus generalization functions were determined within a single session. These sessions consisted of a multiple schedule in which components included probe trials as well as reinforced components. During probe trials, a tone (6, 8, 10, 12, 14, 16, or 18 kHz) was presented. The first five responses on either lever ended the trial. The tone was silenced, and a 30-s timeout in which responses had no programmed consequences ensued. The next trial began after this timeout. Probe trials were interspersed with reinforced components in which either an 8 or 16 kHz tone was presented and the appropriate contingencies were active. Presentation of either a probe trial or reinforced component followed random selection without replacement from a block of 8 possibilities where 2 of the 8 possibilities were probe trials and the rest were reinforced components. Thus, during the test sessions, contingent reinforcement was not delivered in 1/4 of the trials. If a probe trial was selected, the tone presented was randomly selected without replacement from a list of the possible tones, so that each tone was presented before the list was refreshed. This allowed rats to respond in the presence of each probe tone at least two times per session in almost every test session.
2.1.5 Analysis
Ethanol responses expressed as a percentage of total responses during each probe trial was calculated and averaged for each subject and served as the measure for each comparison. Comparisons were made using a repeated measures analysis of variance (ANOVA). First and second baseline function determinations were compared with tone and determination order as factors. The impact of a preceding history of food-predominant responding for 4 or 9 sessions were also compared with the baseline function using a repeated measures ANOVA with tone and preceding session conditions (alternating, four or nine food -predominant condition only sessions) as factors. Another repeated -measures ANOVA was performed to compare stimulus generalization functions obtained during test sessions in which reinforced components alternated or only consisted of food-predominant response conditions. The effect of conditions present during reinforced components of the test session (alternating versus food-predominant) was assessed with a repeated-measures ANOVA following matched histories of preceding sessions with tone and test session conditions as factors. Stimulus generalization functions generated in rats trained with food fixed ratios of 5 and 150 were compared against the function generated in a separate group of rats trained with corresponding food fixed-ratios of 25 and 75 (ethanol fixed-ratios were 5 in every case) using a mixed ANOVA (tone as within-subject, and group as between-subjects). Finally, the stimulus generalization function generated in the group with food FRs of 25 and 150 after four preceding sessions in which conditions alternated was compared with the function generated in the same group following suspended training for the equivalent of 4 or 16 sessions using a repeated-measures ANOVA. For all comparisons, F-values where p<0.05 were considered significant.
Because ANOVA revealed significant differences among stimulus generalization functions due to our experimental manipulations, a non-linear logistic regression was used to determine the parameters defining the baseline function. This allowed comparison of the form of the function between the baseline and other conditions. The data were fit to the function: using a non-linear least squares regression with a Gauss-Newton algorithm and starting values of Asym=100, T50=12 kHz, and k=1. In this sigmoidal function, Asym describes the upper asymptote, T50 describes the inflection point of the sigmoidal curve (the tone value where the portions of the curve above and below are symmetrical), and k is a scaling factor that describes the general shape of the function. The lower asymptote is assumed to be 0. Changes in Asym reflect shifts in the upper asymptote. The T50 represents the tone where percent ethanol responding equals 1/2 Asym. Changes in T50 reflect leftward or rightward shifts in the function. Ninety-five percent confidence limits for each of these parameters were determined for the first stimulus generalization function (baseline) and parameters derived for the other conditions and second baseline function compared against these limits. Parameter values for conditions that fell outside of these confidence limits were considered significantly different from the baseline function.
All analyses were performed using the R statistical program (R Development Core Team, 2011). ANOVA was performed using the aov function. Non-linear regression was performed using the nls function from the lme4 package. Briefly, the deriv function was used to compute the derivatives of the equation shown above. This result was then used to model the non-linear function and calculate parameter estimates and confidence intervals for the control stimulus generalization function.
2.2 Experiment 1: Stability of the stimulus generalization function over time
Stimulus generalization during two test sessions in which reinforced components alternated between food FR150 and food FR5 conditions were determined after four consecutive sessions in which conditions also alternated. The two test sessions were separated by approximately 9 months and all experimental manipulations in this group. As described in Results (section 3.2), these determinations did not differ, and the first determination was used as the basis for subsequent comparisons.
2.3 Experiment 2: Impact of presenting only food FR5, ethanol FR5 conditions during determination of the stimulus generalization function
The first stimulus generalization function determination described above was compared with a stimulus generalization function generated in a test session in which only 8 kHz tone, food FR5, and ethanol FR5 conditions were presented during reinforced components. These test sessions were preceded by at least four consecutive sessions under the multiple concurrent schedule.
2.4 Experiment 3: Stimulus generalization after four or nine consecutive sessions of exposure to only food FR5 conditions
Stimulus generalization functions generated during test sessions where reinforced component conditions alternated after 4 or 9 preceding sessions in which only 8kHz tone, food FR5, ethanol FR5 conditions were compared with the baseline stimulus generalization function from Experiment 1. Data for the 9 preceding session condition was collected in n=4 rats.
2.5 Experiment 4: The impact of presenting only food-predominant response conditions during determination of the stimulus generalization function after nine preceding sessions of responding under only food-predominant response conditions
We next assessed whether conditions present during reinforced components of the test session could influenced the stimulus generalization function generated after nine preceding sessions of food-predominate response conditions. To do this, we compared the function generated after the nine preceding session condition in Experiment 3 with a function generated after the same history, but where only food -predominate response conditions were presented during reinforced components of the test session.
2.6 Experiment 5: The impact of different relative response requirements for food versus ethanol on stimulus generalization
In order to assess whether the relative response requirements affected the stimulus generalization function, a separate group of rats was trained in the same manner as the first group, but in these rats, response requirements for food associated with 8 and 16 kHz tones FR25 and 75, respectively. The stimulus generalization function was determined in a test session in which reinforced components alternated between food FR25 and FR75 conditions after four preceding sessions in which components also alternated. This function was compared with the first stimulus generalization function determined in rats responding under food FR5 and FR25 contingencies (Experiment 1).
2.7 Experiment 6: Stimulus generalization during a test session in which reinforced components alternated between food FR25 and food FR75 after training was suspended for the equivalent of 4 or 16 sessions
In order to determine if time not responding for ethanol affected the stimulus generalization function, functions were determined during a test session in which reinforced components alternated between food FR25 and food FR75 conditions after the equivalent of 4 or 16 sessions where training was suspended. During these periods, rats remained in their home cages, and were never placed in the operant apparatus. Stimulus generalization functions after suspended training were compared to the function generated in Experiment 5. Data for the 4 session suspended training condition was collected in n=4 rats.
2.8 Ethanol consumption
Ethanol consumption was determined after a session in which rats responded under the multiple concurrent schedule. Using a method described by Javors et al., (2005) breath ethanol levels were determined immediately after the session ended. From the breath ethanol levels, blood ethanol levels can be estimated. After the session ended, rats were placed in a modified commercially available restraint such that their head was inside a heated chamber. Expired air was collected in the chamber for 30-sec, then analyzed for ethanol content with gas chromatography.
During these sessions, rats earned 28.2 ± 2.7 ethanol deliveries. This represents an earned dose of approximately 0.94 g/kg. Immediately after theses sessions the estimated BAC for these rats was 0.07 ± 0.02 g/dl. This represents consumption of a pharmacologically active amount of ethanol.
3. RESULTS
3.1 Performance under the multiple concurrent schedule
Between experimental manipulations, rats responded under the multiple concurrent schedule. In these sessions, stimuli and associated conditions alternated between the 8 kHz tone and the 16 kHz tone. In the presence of the 8 kHz tone, 3.7% ± 2.1 (mean ± S.E.M.) of the total responses were on the ethanol lever. In the presence of the 16 kHz tone, 82.0% ± 3.2 of the total responses made were on the ethanol lever. In these sessions, rats earned 29.9 ± 1.3 food pellets and 31.4 ± 2.0 ethanol deliveries. This equates to an earned dose of approximately 1.0 g/kg per 30-min session.
3.2 Experiment 1: The stimulus generalization function is stable over time
Stimulus generalization functions observed under the same conditions were stable over time. Despite being separated by over nine months and various intervening experimental manipulations, there was no effect of determination order (first versus second, F[1, 56] = 0.4, p>0.05) nor was there an interaction between order and tone (F[6, 56] = 1.0, p>0.05) on the stimulus generalization function (see Tables 1 and 2), though an effect of tone was present (F[6, 56]=19.7, p<0.001). Thus, the stimulus generalization function appears stable over time and various experimental manipulations.
Table 1.
Percent Ethanol Responses During Probe Trials on Two Different Occasions in Each Subject.
| Rat | 1 | 2 | 3 | 4 | 5 | Average | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Determination | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | 1 (s.d.) | 2 (s.d.) | |
| Tone (kHz) | 6 | 0 | 0 | 0 | 0 | 28 | 0 | 0 | 6 | 0 | 0 | 6 (12) | 1 (3) |
| 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 (0) | 1 (3) | |
| 10 | 0 | 42 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 (0) | 8 (19) | |
| 12 | 28 | 36 | 100 | 0 | 15 | 67 | 42 | 86 | 0 | 0 | 37 (35) | 38 (39) | |
| 14 | 50 | 100 | 83 | 100 | 56 | 21 | 92 | 67 | 42 | 0 | 64 (20) | 58 (46) | |
| 16 | 0 | 92 | 100 | 83 | 67 | 94 | 36 | 89 | 81 | 100 | 57 (35) | 92 (6) | |
| 18 | 92 | 69 | 63 | 71 | 67 | 50 | 71 | 67 | 86 | 83 | 76 (11) | 68 (12) | |
Ethanol responding as a percentage of total responses was assessed in a test session where reinforced components alternated between 8 kHz and 16 kHz. See text for further details
Table 2.
Estimated Parameters for Fitted Models for Stimulus Generalization Functions Observed Under Different Experimental Situations
| Row | Determination | Preceding sessions | Test session | Asymptote | T50 (kHz) | Scale | |||
|---|---|---|---|---|---|---|---|---|---|
| Mean | [95% C.I.] | Mean | [95% C.I.] | Mean | [95% C.I.] | ||||
| 1 | 1 | Alternating (4d) | Alternating | 79 | [63–100] | 12.3 | [11.0–13.9] | 0.9 | [0.4–N.D.] |
| 2 | 2 | Alternating (4d) | Alternating | 66 | 11.9 | 2.4 | |||
| 3 | Alternating (4d) | Food-preferred | 47 | 12.2 | 0.6 | ||||
| 4 | Food-preferred (4d) | Alternating | 40 | 12.1 | 0.6 | ||||
| 5 | Food-preferred (9d) | Alternating | 23 | 9.8 | 1.2 | ||||
| 6 | Food-preferred (9d) | Food-preferred | 12 | 13.4 | 0.7 | ||||
Confidence intervals were established for the model fitted to the first determination in which conditions alternated between ethanol- and food-preferred conditions during the test session and in preceding sessions. Parameters for other situations were considered significantly different (p<0.05) if the estimate was not within the interval for the first model.
95% C.I.–95% confidence interval
N.D. - parameter was not determined.
3.3 Experiment 2: Presenting only food-predominant conditions during reinforced components in the test session shifts the stimulus generalization function downwards
During test sessions when only food FR5, ethanol FR5 contingencies and the 8 kHz stimulus were presented during reinforced components, the stimulus generalization function is shifted downwards. The stimulus generalization function generated during a test session in which only the 8kHz tone, food FR5, and ethanol FR5 conditions were presented during reinforced components was compared with the function generated during a test session in which reinforced components alternated between 8kHz and 16 kHz tones and associated contingencies. Significant effects of test condition (F[1, 49] = 5.1, p<0.05) and tone (F[6, 49]=10.6, p<0.001) were present, but there was no interaction (F[6, 49]=0.8, p>0.05) when these two functions were compared. This shift is shown in Figure 1 and Table 2 (row 1 versus row 3) where the asymptote was lower for the stimulus generalization function generated during a test session in which only food FR5 conditions (versus alternating FR5 and FR150) were presented during reinforced components. Thus, reinforcement contingencies present during the stimulus generalization test session can influence the stimulus generalization function.
Figure 1.

Effect of conditions during the test session on the stimulus generalization function. Stimulus generalization functions generated during a test session in which conditions during reinforced components alternated (open circles, solid line) between an 8kHz tone signaling food FR5 and ethanol FR5 (food-predominant) and a 16kHz tone signaling food FR150 and ethanol FR5 (ethanol-predominant) or were only food-predominant conditions (solid triangles, dashed line). Test sessions were conducted after four preceding sessions in which conditions alternated. Points show the mean percent ethanol responding (left axis) and food responding (right axis) for 5 rats during presentation of each tone indicated on the abscissa. Bars represent the standard error of the mean, and lines represent the sigmoidal function. Some points have been nudged along the abscissa for clarity.
3.4 Experiment 3: A history of responding under food-predominant conditions shifts the stimulus generalization function downwards
We next assessed whether providing a history of 4 or 9 preceding sessions in which only 8 kHz, food FR5, ethanol FR5 conditions were presented could produce shifts in the stimulus generalization function. Reinforced components during the test sessions alternated in these studies. A significant main effect of history(F[2, 72]=5.9, p<0.01) and tone (F[6, 72]=8.9, p<0.001) was present, but there was no interaction present (F[12, 72] = 1.7, p>0.05). As shown in Figure 2, 4 or 9 preceding sessions in which only food FR5, ethanol FR5 conditions were presented resulted in a downwards shift of the stimulus generalization function. This shift was time-dependent, as can be seen in Table 2 (row 1 versus rows 4 and 5). These shifts were due to lower asymptotes of the curve without changes in the values of k following 4 or 9 preceding sessions. The inflection point (T50) was shifted significantly leftward after nine (but not four) preceding sessions of only food FR5 conditions. Thus, a history of responding under the stimulus and contingency conditions that result in food-predominant responding can shift the stimulus generalization function by decreasing responding for ethanol, even in the presence of the training stimulus (16 kHz) which had been associated with over 80% ethanol-lever responding under baseline conditions.
Figure 2.

The effect of a history of responding under only the food -predominant condition on the stimulus generalization function. All stimulus generalization functions were generated during a test session with alternating (between food-predominant and ethanol-predominant conditions) reinforced components. The function generated following four preceding sessions of alternating conditions is reproduced from Figure 1 (open circles, solid lines). The function determined after four preceding sessions of responding only under food -predominant conditions is shown by the upright triangles and dashed line, and after nine preceding sessions of responding under only food -predominant conditions by the downward triangles and dotted line. Points represent the mean ± S.E.M. Lines represent the sigmoidal function fitted to each data set. All points represent data from five rats except for the downwards triangle which represents four rats. Some points have been nudged along the abscissa for clarity.
3.5 Experiment 4: Presenting only food FR5 conditions during reinforced components in the test session shifts the function generated after nine preceding sessions of food FR5 conditions further downwards
Extending a history of 9 preceding sessions of exposure to only food-predominant response conditions into the test session further shifted the stimulus generalization gradient. This result is shown in Figure 3. An ANOVA comparing functions generated after nine sessions of only food-predominant conditions during test sessions with alternating or only food -predominant conditions during reinforced components revealed effects of tone (F[6, 49]=2.6, p<0.05) and test condition (F[1, 49]=7.3, p<0.01), but not an interaction (F[6, 49]=2.1, p>0.05). Thus, reinforcement contingencies present during the stimulus generalization test session can enhance the effect of history on the stimulus generalization function.
Figure 3.
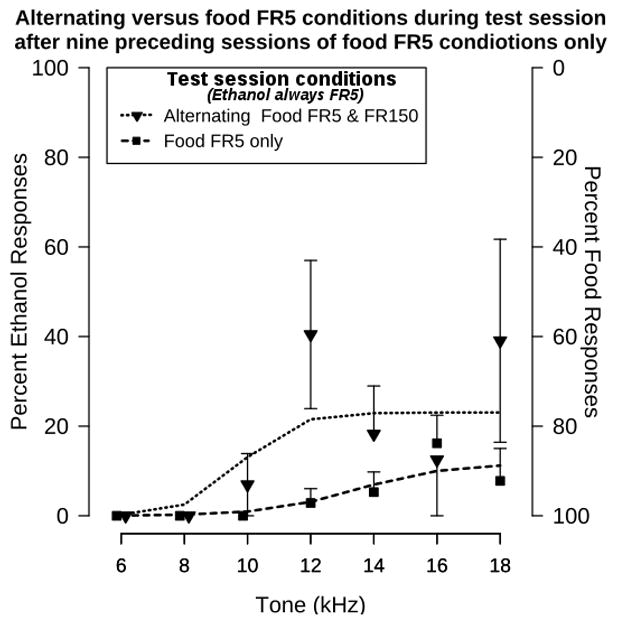
Effect of conditions during the test session on stimulus generalization functions generated following 9 preceding session of responding under only food -predominant conditions. The function generated during a test session with alternating (between food-predominant and ethanol-predominant conditions) reinforced components is represented by the downward facing triangles and dotted line, redrawn from Figure 2. The function generated during a test session in which only food predominant conditions were present during reinforced components is represented by the squares and dashed line. Points represent the mean ± S.E.M for 5 rats. Lines represent the sigmoidal function fitted to each data set. Some points have been nudged along the abscissa for clarity.
3.6 Experiment 5: Effect of different food contingencies on stimulus generalization function
To determine whether different relative contingencies for food versus ethanol affect the stimulus generalization function, a second group of rats was trained to respond under the same procedure, except that food contingencies were FR25 and FR75 instead of FR5 and FR150. Changing the contingencies for food did not greatly affect the response allocation under either stimulus condition. During the four sessions that preceded the test session, during components where the 8 kHz tone was presented and contingencies were food FR25 and ethanol FR5, responding was primarily on the food -associated lever (median=3.6% ethanol-lever responses, IQR=1.6–9.1%). During components where the 16 kHz tone was presented and contingencies were food FR75 and ethanol FR5, responding was predominately on the ethanol-associated lever (median=82.4%, IQR=26.9–93.6%). These results did not differ from those obtained in rats trained under food FR5 and FR150 contingencies. In those rats, during components where the 8 kHz tone was presented and contingencies were food FR5 and ethanol FR5 in the four sessions that preceded the test session, responding was primarily on the food-associated lever (median=1.2% ethanol-lever responses, IQR=1.2–5.8%). During components where the 16 kHz tone was presented and contingencies were food FR150 and ethanol FR5, responding was predominately on the ethanol-associated lever (median=85.4%, IQR=67.4–90.8%). Thus, the stimuli and contingencies controlled behavior similarly to the extent that responding for food predominated when response requirements were similar and responding for ethanol predominated when the response requirement for food was relatively high.
Likewise, as shown in Figure 4, the stimulus generalization function generated in the group of rats trained with food FR25 and FR75 contingencies did not differ from that generated in the group trained to respond under the food FR5 and FR150 contingencies. This was confirmed by an ANOVA in which a significant effect of tone was present (F[6,53]=12.8, p<0.05), but not of group (F[1, 53]=3.4, p>0.05). Thus, the stimulus generalization function appears to depend more on the response allocation than on the relative response requirements that result in the allocation.
Figure 4.
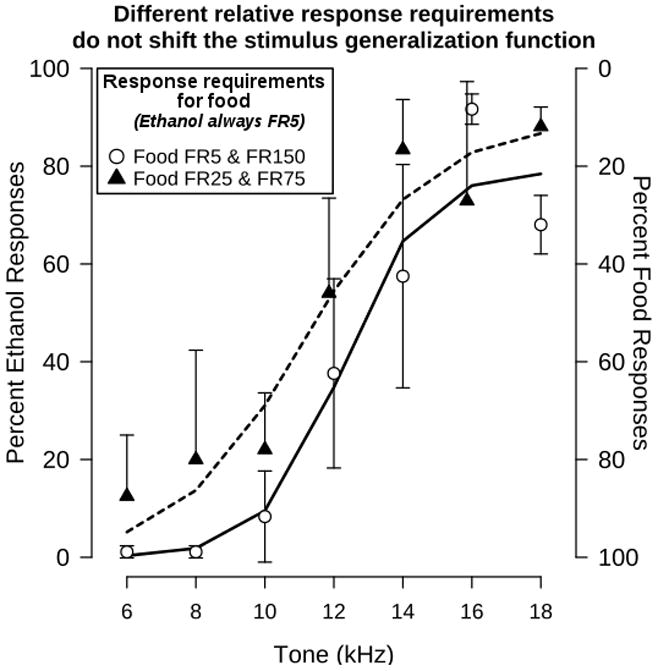
Effect of different relative response requirements for food and ethanol on the stimulus generalization function. The stimulus generalization function generated in rats responding under food FR150/ethanol FR5 and food FR5/ethanol FR5 during a test session with alternating reinforced components following four sessions of alternating contingency conditions is reproduced from Figure 1 (open circles, solid lines). This function is compared with the function generated in a separate group of 5 rats responding under food FR75/ethanol FR5 and food FR25/ethanol FR5 contingencies (triangles, dashed line). Points represent the mean ± S.E.M for 5 rats. Lines represent the sigmoidal function fitted to each data set. Some points have been nudged along the abscissa for clarity.
3.7 Experiment 6: Effect of suspended training on stimulus generalization function
In order to assess whether the changes in stimulus generalization observed in Experiment 1 could be due simply to not being exposed to the 16 kHz tone and not responding for ethanol, stimulus generalization functions were determined after the equivalent of 4 or 16 sessions of suspended training. During this time, rats remained in their home cages. As shown in Figure 5, suspending training did not affect the stimulus generalization function. This was confirmed with ANOVA where a main effect of tone was present (F[6, 154]=15.5, p<0.05), but not of suspended versus ongoing training (F[2, 154]=0.4, p>0.05). There was also no interaction between tone and training condition (F[12, 154] = 0.9, p>0.05). Thus, suspending training did not change the stimulus generalization function.
Figure 5.

Effect of suspending training on the stimulus generalization function. The stimulus generalization function generated following four sessions of alternating contingency conditions in rats responding under food FR75/ethanol FR5 and food FR150/ethanol FR5 conditions is reproduced from Figure 4 (open triangles, dashed line). This function is compared with the function generated following suspended training for the equivalent of 4 or 16 sessions (filled triangles, dashed and dotted lines). Points represent the mean ± S.E.M for 5 rats. Lines represent the fitted model for each function. Some points have been nudged along the abscissa for clarity.
4. DISCUSSION
Recent history can affect how likely an animal is to respond for ethanol in the presence of both trained and various untrained stimuli in a choice between food and ethanol. This finding has implications for the treatment of alcoholism, and extends the stimulus generalization literature. When rats are trained to discriminate between tones signaling contingencies that result either in ethanol-predominant or food-predominant responding, varying the tone presented between the two trained stimuli results in a graded change in responding for ethanol versus food. A recent history of responding under only food -predominant conditions shifts the stimulus generalization function for responding for ethanol downwards such that longer periods of exposure to only the food -predominant response conditions (8kHz, food FR5, ethanol FR5) shift the upper asymptote of the function down. Establishing the generalization function during a test session in which only food-predominant conditions are present during reinforced components enhances this downward shift. These shifts result from a change in the asymptote of the generalization function, and not to a change in its position along the abscissa (T50) or its form (k). Thus, the likelihood of responding for ethanol in the presence of stimuli similar to those previously occasioning ethanol-predominant choice diminishes with repeated exposure to conditions that occasion a different behavior in the same context. These results are relevant to processes that may occur in recovery, and may provide a basis for improving treatment outcomes.
Stimulus control has been posited as important in the development and maintenance of addiction, as well as in the development and maintenance of recovery. Repeated exposure to stimuli associated with drug use has been proposed to broaden the stimulus generalization gradient, increasing the likelihood of subsequent drug-seeking in a variety of environments (Schuster, 1986). Conversely, the potential importance of diminution of the stimulus generalization gradient during recovery such that drug use generalizes to fewer and fewer stimuli has also been suggested (Bickel and Kelley, 1997; Schuster, 1986). Unfortunately, this area has been largely unexplored. Here, we use a choice procedure and explore generalization of a stimulus that occasions alcohol-predominant responding under conditions that are similar to those seen during recovery.
During recovery, drug use may be reduced by a variety of contingencies, including reinforcement of alternative behavior (Higgins, 1997; Iguchi et al., 1997). A preclinical model which shares this feature of recovery could provide insight into the mechanisms responsible for the reduced likelihood of relapse over longer periods of recovery. Moreover, a preclinical model must capture this effect to allow its study. Unfortunately, the most commonly used animal model of relapse does not. Rather, longer periods of not responding for drug or alcohol increase reinstated responding upon re-exposure to stimuli that had previously occasioned drug- or alcohol-seeking after it has been reduced by withdrawing access (e.g. Grimm et al., 2003). Here we present a preclinical model which reduces alcohol-seeking by providing an alternative behavior, and in which the allocation of responding for ethanol versus food shifts toward responding for food in the presence of a stimulus that had been associated with ethanol-predominant responding with longer periods of reduced alcohol use.
In this procedure, rats respond predominantly for food when exposed to an 8 kHz tone associated with food FR5 and ethanol FR5, and predominantly for ethanol when exposed to a 16 kHz tone associated with a food FR150 and ethanol FR5. Following a series of multiple component sessions in which conditions alternate, increasing the tone from 8 kHz to 16 kHz results in a graded increase in the proportion of responses on the ethanol versus food lever.
This pattern of responding is similar to that reported by Hanson (1959) using food reinforced behavior. In that study, pigeons were trained to discriminate between two wavelengths of light. One signaled the availability of food (S+), while the other signaled extinction (S−). As the stimulus was varied from S− to S+, the probability of responses for food increased. However there are some important procedural differences between this kind of discrimination training and that used in the present study. First, in the present study, rats’ response allocation was guided by the contingencies in place for food and ethanol rather than their availability. Thus, the gradient seen in the present work was formed by a discrimination between a stimulus signaling contingencies that resulted in ethanol-predominant responding versus a stimulus that signaled food-predominant responding, even while ethanol remained available at the same FR. Second, in the study by Hanson, generalization gradients were determined under extinction conditions, while in the present study functions were also determined under extinction, but in the context of reinforced responding in the presence of the trained discriminative stimuli. Results from this study show that conditions present during the test session can influence the generalization gradient (e.g., the decrease in the asymptote during a test session with only food-predominant response conditions during reinforced components compared to alternating components after the same preceding four session history).
Reduction of the upper asymptote of the stimulus generalization function following increasing periods of exposure to only the stimulus and contingencies associated with food -predominant choice appears on its face similar to results reported by Thomas and King (1959). In that study, the stimulus generalization function about a stimulus associated with food availability was shifted downward by increasing the body weights of pigeons. This would imply that, in our experiment, the motivation to respond for ethanol decreased in response to exposure to conditions that resulted in reduced ethanol intake, or that motivation for food increased. However, alternative explanations are also possible.
The present results can also be interpreted in the context of studies performed by Jenkins and Harrison (1960). This study, together with others that followed provide evidence that the control by one set of stimuli is diminished by another if the former set no longer provides additional information (Honig and Urcuioli, 1981). In the study by Jenkins and Harrison, one group of rats were exposed to either a 450 Hz or 2500 Hz tone which signaled contingent food availability, or no tone which signaled no food availability and exhibited a stimulus generalization function with peaks corresponding to the two training tone frequencies. However, another group of rats exposed to the same 450 and 2500 Hz tones signaling contingent food availability, but not exposed to the silent condition signaling food non-availability, exhibited a flat stimulus generalization function (Jenkins and Harrison, 1960). These results were interpreted as a shift in attention from the tone to other stimuli present, such as the lighting conditions or the test chamber itself in the group not exposed to the silent non-reinforced condition, as the tones provided no additional information about the contingencies in place. However, in the group exposed to the extinction condition, the tones provided important information about the availability of food, and thus were attended to by these subjects (Honig and Urcuioli, 1981).
One possible explanation for the loss of stimulus control by the tones over responding is a transition from goal-directed to habitual behavior where responding occurs on the food lever regardless of the tone presented (Everitt et al., 2001; Robinson and Berridge, 1993). In the situation where only food-predominate response conditions are presented, the 8 kHz stimulus tone provides no additional information; contingencies remain fixed and responding proceeds on the food lever throughout the session. In this situation, the context of the experimental chamber becomes the most salient stimulus, as it predicts the availability of reinforcement and the associated contingencies. This may lead to habitual responding for food that is less influenced by the tone present in the chamber. The flattening of the stimulus generalization function may reflect this transition.
A converse conceptualization has been proposed as a mechanism for the development of substance abuse: during repeated, frequent use, control of drug use by stimuli or contingencies weakens and drug use becomes habitual (Everitt et al., 2001). It might then follow that a transition from substance abuse could occur if some alternative behavior that is incompatible with substance use becomes habitual. If an alternative to drinking becomes habitual, stimuli may lose their ability to occasion drinking. This would seem to provide protection when an individual re-encounters cues that might precipitate relapse.
Interpreted in this context, the present results suggest that longer periods of exposure to conditions where alternative behavior is reinforced can diminish the ability of stimuli to occasion alcohol or drug use by reducing the likelihood of attending to these stimuli. This would be consistent with the finding that heavy drinkers exhibit greater attentional bias to alcohol-related stimuli than social drinkers and that attentional bias to alcohol-related stimuli predicts relapse (Garland et al., 2012; Townshend and Duka, 2001), and may explain why longer periods of recovery are correlated with reduced propensity to relapse in several studies (Gilpin et al., 1997; Gossop et al., 1990; McKay et al., 2001). As more general aspects of the environment come to be associated with alternative behavior, stimuli associated with alcohol or drug use are attended to less, and their ability to occasion alcohol or drug seeking diminishes. Treatments that hasten this attenuation of the ability of stimuli to occasion alcohol or drug use would be potentially useful adjuncts for alcoholism or addiction therapies.
The present experiments demonstrate that the flattening of the generalization function due to a history of food-predominant responding is blunted when reinforced components in the test session alternate between food- and ethanol-predominant conditions. In other words, responding for ethanol in the presence of untrained stimuli is more likely when the behavior still occurs in the general context. There are a several important implications of this finding. First, as previously noted, conditions present during the test session clearly influence stimulus generalization functions. Second, the assessment of craving or other responses thought to be related to relapse may be influenced by the testing environment. This is consistent with a report by Childress et al. (1986) in which methadone-maintained patients reported their craving after viewing a video containing drug-related content. Reported craving was greatest when patients viewed the video in their home (presumably an environment where drug use commonly occurred), less in a community clinic (more distant from the site of most of their drug seeking and use, but still in the neighborhood), and was further reduced when presented in a laboratory (a setting in which drug seeking and consumption had presumably never occurred). Finally, this result could reveal a risk for controlled drinking by those in recovery. Controlled drinking may prevent loss of control once a drinking bout is initiated, due to repeated experience drinking moderately during recovery. However, regular controlled drinking likely results in a broad range of stimuli still occasioning drinking. There is a growing consensus that controlled drinking may pose an acceptable risk in recovery from moderate, but not severe alcoholism (Anonymous, 2009; Dawson et al., 2007; Sobell and Sobell, 1995). This may be related to these different aspects of loss of control (initiation versus amount of drinking after initiation).
In conclusion, recent experience with contingencies occasioning an alternative to ethanol-seeking reduces the control that ethanol-related stimuli exert over ethanol-seeking. This is consistent with longer periods of recovery being associated with reduced rates of relapse and offers a potential mechanism for this phenomenon. These results are also consistent with data from our laboratory in which longer periods of reduced responding for ethanol under the same procedure results in a reduced propensity to respond for ethanol upon re-exposure to the stimulus that had previously occasioned ethanol-predominant responding (Ginsburg and Lamb, submitted). Additionally, stimulus generalization functions are influenced by conditions present during the test session. This seems consistent with humans recovering from opiate addiction who report greater subjective craving after viewing drug-related images in their home environment versus a clinic or laboratory. Reducing the ability of stimuli to occasion alcohol or drug use is likely to be critical in the maintenance of recovery. Finding treatments that enhance this decay in stimulus generalization may be helpful to those wishing to maintain recovery.
Acknowledgments
Role of Funding Source: “Nothing Declared”
The authors wish to thank Olivia Dominguez for her technical assistance. This work was funded by Public Health Service grant AA016987 and the Alcoholism Breakthrough Endowed Research Grant provided by a private donor to the institution. The authors also with to thank Dr. Charles France for valuable discussion related to data presentation. Finally, the authors wish to thank Dr. Charles Schuster for his intellectual contributions to the field that form the basis for these investigations.
Footnotes
Conflict of Interest: “No Conflict Declared”
Contributors: R.J. Lamb
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Anonymous. Alcohol abstinence vs. moderation Degree of dependence predicts which strategy works best. Harv Ment Health Lett. 2009;25:6. [PubMed] [Google Scholar]
- Bickel WK, Kelley T. Stimulus control processes in drug taking: implications for treatment. In: Baer DM, Pinkston EM, editors. Environment and Behavior. Westview Press; Boulder: 1997. pp. 185–193. [Google Scholar]
- Childress AR, McLellan AT, O’Brien CP. Conditioned responses in a methadone population. A comparison of laboratory, clinic, and natural settings. J Subst Abuse Treat. 1986;3:173–179. doi: 10.1016/0740-5472(86)90018-8. [DOI] [PubMed] [Google Scholar]
- Dawson DA, Goldstein RB, Grant BF. Rates and correlates of relapse among individuals in remission from DSM-IV alcohol dependence: a 3-year follow-up. Alcohol Clin Exp Res. 2007;31:2036–2045. doi: 10.1111/j.1530-0277.2007.00536.x. [DOI] [PubMed] [Google Scholar]
- Everitt BJ, Dickinson A, Robbins TW. The neuropsychological basis of addictive behaviour. Brain Res Rev. 2001;36:129–138. doi: 10.1016/s0165-0173(01)00088-1. [DOI] [PubMed] [Google Scholar]
- Garland EL, Franken IHA, Howard MO. Cue-elicited heart rate variability and attentional bias predict alcohol relapse following treatment. Psychopharmacology. 2012;222:17–26. doi: 10.1007/s00213-011-2618-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilpin EA, Pierce JP, Farkas AJ. Duration of smoking abstinence and success in quitting. J Natl Cancer Inst. 1997;89:572–576. doi: 10.1093/jnci/89.8.572. [DOI] [PubMed] [Google Scholar]
- Gossop M, Green L, Phillips G, Bradley B. Factors predicting outcome among opiate addicts after treatment. Br J Clin Psychol. 1990;29(Pt 2):209–216. doi: 10.1111/j.2044-8260.1990.tb00871.x. [DOI] [PubMed] [Google Scholar]
- Grimm JW, Lu L, Hayashi T, Hope BT, Su TP, Shaham Y. Time-dependent increases in brain-derived neurotrophic factor protein levels within the mesolimbic dopamine system after withdrawal from cocaine: implications for incubation of cocaine craving. J Neurosci. 2003;23:742–747. doi: 10.1523/JNEUROSCI.23-03-00742.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanson HM. Effects of discrimination training on stimulus generalization. J Exp Psychol. 1959;58:321–334. doi: 10.1037/h0042606. [DOI] [PubMed] [Google Scholar]
- Higgins ST. The influence of alternative reinforcers on cocaine use and abuse: a brief review. Pharmacol Biochem Behav. 1997;57:419–427. doi: 10.1016/s0091-3057(96)00446-7. [DOI] [PubMed] [Google Scholar]
- Honig WK, Urcuioli PJ. The legacy of Guttman and Kalish (1956): twenty-five years of research on stimulus generalization. J Exp Anal Behav. 1981;36:405–445. doi: 10.1901/jeab.1981.36-405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt WA, Barnett LW, Branch LG. Relapse rates in addiction programs. J Clin Psychol. 1971;27:455–456. doi: 10.1002/1097-4679(197110)27:4<455::aid-jclp2270270412>3.0.co;2-r. [DOI] [PubMed] [Google Scholar]
- Iguchi MY, Belding MA, Morral AR, Lamb RJ, Husband SD. Reinforcing operants other than abstinence in drug abuse treatment: an effective alternative for reducing drug use. J Consult Clin Psychol. 1997;65:421–428. doi: 10.1037//0022-006x.65.3.421. [DOI] [PubMed] [Google Scholar]
- Javors MA, Ginsburg BC, Friesenhahn G, Delallo L, Lamb RJ. Rat breathalyzer. Alcohol Clin Exp Res. 2005;29:1853–1857. doi: 10.1097/01.alc.0000183228.07510.a2. [DOI] [PubMed] [Google Scholar]
- Jenkins HM, Harrison RH. Effect of discrimination training on auditory generalization. J Exp Psychol. 1960;59:246–253. doi: 10.1037/h0041661. [DOI] [PubMed] [Google Scholar]
- Keller M. On the loss-of-control phenomenon in alcoholism. Br J Addict Alcohol Other Drugs. 1972;67:153–166. doi: 10.1111/j.1360-0443.1972.tb01188.x. [DOI] [PubMed] [Google Scholar]
- McKay JR, Merikle E, Mulvaney FD, Weiss RV, Koppenhaver JM. Factors accounting for cocaine use two years following initiation of continuing care. Addiction. 2001;96:213–225. doi: 10.1046/j.1360-0443.2001.9622134.x. [DOI] [PubMed] [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing R. Foundation for Statistical Computing; Vienna, Austria: 2011. [Google Scholar]
- Robinson TE, Berridge KC. The neural basis of drug craving: an incentive-sensitization theory of addiction. Brain Res Rev. 1993;18:247–291. doi: 10.1016/0165-0173(93)90013-p. [DOI] [PubMed] [Google Scholar]
- Schuster CR. Implications of laboratory research for the treatment of drug dependence. In: Stolerman IP, Goldberg SR, editors. Behavioral Analysis of Drug Dependence. Academic Press Inc; London: 1986. pp. 357–386. [Google Scholar]
- Sobell MB, Sobell LC. Controlled drinking after 25 years: how important was the great debate? Addiction. 1995;90:1149–1153. discussion 1157–1177. [PubMed] [Google Scholar]
- Thomas DR, King RA. Stimulus generalization as a function of level of motivation. J Exp Psychol. 1959;57:323–328. doi: 10.1037/h0042183. [DOI] [PubMed] [Google Scholar]
- Townshend JM, Duka T. Attentional bias associated with alcohol cues: differences between heavy and occasional social drinkers. Psychopharmacology. 2001;157:67–74. doi: 10.1007/s002130100764. [DOI] [PubMed] [Google Scholar]