

Abstract
Biologically active macrocycles containing a cyclic imine were isolated for the first time from aquaculture sites in Nova Scotia, Canada, during the 1990s. These compounds display a “fast-acting” toxicity in the traditional mouse bioassay for lipophilic marine toxins. Our work aimed at developing receptor-based detection method for spirolides using a microsphere/flow cytometry Luminex system. For the assay two alternatives were considered as binding proteins, the Torpedo marmorata nicotinic acetylcholine receptor (nAChR) and the Lymnaea stagnalis acetylcholine binding protein (Ls-AChBP). A receptor-based inhibition assay was developed using the immobilization of nAChR or Ls-AChBP on the surface of carboxylated microspheres and the competition of cyclic imines with biotin-α-bungarotoxin (α-BTX) for binding to these proteins. The amount of biotin-α-BTX bound to the surface of the microspheres was quantified using phycoerythrin (PE)-labeled streptavidin and the fluorescence was analyzed in a Luminex 200 system. AChBP and nAChR bound to 13-desmethyl spirolide C efficiently; however the cross-reactivity profile of the nAChR for spirolides and gymnodimine more closely matched the relative toxic potencies reported for these toxins. The nAChR was selected for further assay development. A simple sample preparation protocol consisting of an extraction with acetone yielded a final extract with no matrix interference on the nAChR/microsphere-based assay for mussels, scallops and clams. This cyclic imine detection method allowed the detection of 13-desmethyl spirolide C in the range of 10–6000 μg/kg of shellfish meat, displaying a higher sensitivity and wider dynamic range than other receptor-based assays previously published. This microsphere-based assay provides a rapid, sensitive and easily performed screening method that could be multiplexed for the simultaneous detection of several marine toxins.
Keywords: microsphere-based assay, marine phycotoxins, spirolide, gymnodimine, nicotinic acetylcholine receptor, acetylcholine binding protein, α-bungarotoxin
INTRODUCTION
Shellfish constitute a worldwide food resource that may be contaminated with toxins produced by dinoflagellates, representing a potential threat to human health and having an important economic impact on the aquaculture industry. Spirolides and gymnodimines (Figure 1) are marine phycotoxins that belong to the group of macrocyclic imines 1,2 due to the presence of a cyclic imine moiety in their macrocyclic chemical structure. Other marine toxins of the cyclic imine structural group are known as pinnatoxins, pteriatoxins, prorocentrolides and spiro-prorocentrimine 3–7.
Figure 1.

Chemical structures of spirolides and gymnodimines
The spirolides were discovered during routine monitoring of the presence of the lipophilic diarrhetic shellfish poisoning (DSP) toxins in aquaculture sites along the South Eastern coast of Nova Scotia, Canada, in the early 1990s 8. The marine dinoflagellates Alexandrium ostenfeldii and Alexandrium peruvianum have been identified as the spirolide-producing microorganisms 9–11. Gymnodimines are produced by the planktonic dinoflagellate Karenia selliformis and were first isolated from New Zealand oysters in 1994 12,13. Although, spirolides and gymnodimines have not been linked to human intoxication so far 14, these compounds displayed “fast-acting” toxicity in the traditional bioassay 14,15.
The mouse bioassay (MBA) has been the most commonly used method to detect marine toxins 14, and it has been a basis for protecting human health for decades 16. However, such techniques have several drawbacks because of ethical concerns, sensitivity and specificity 17. In the last few years several techniques using mass spectrometry (MS) have also been developed for the detection of macrocyclic imines 1,18–24. However these MS-based methods will only detect known compounds for which standards are available being blind to the presence of new toxic analogues or groups. Furthermore the use of MS-based techniques for routine detection of toxins requires expensive instrumentation and highly trained laboratory personnel. The evidence that spirolides and gymnodimine target muscular and neuronal nAChR subtypes with high affinity 25,26 has prompted the development of alternative techniques using a receptor-based approach for the detection of spirolides and gymnodimines by fluorescence polarization and chemiluminescence 27–29.
In recent years multiplexed binding assays using microspheres have become a widely used tool for clinical diagnostics and research 30. The Luminex technology is based on the use of polystyrene beads internally dyed with two spectrally distinct fluorochromes, resulting in 100 different microsphere populations. The surface carboxyl groups of the microspheres can be used to couple a binding molecule specific for the interaction with an analyte of interest. The binding molecule immobilized on the surface of a microsphere class confers specificity to each microsphere population for a certain analyte, and the possibility of multidetection of several analytes by combining different microsphere classes in the same sample. The discrimination of microsphere classes and quantification of binding signal take place in a Luminex analyzer by detecting two different fluorescence signals per microsphere.
Macrocyclic imines, as mentioned above, have been shown to interact with nAChRs and to block their activity 25,26. Recently, a soluble acetylcholine binding protein (AChBP) from the water snail Lymnaea stagnalis, has also been demonstrated to bind these toxins 25. Actually, soluble AChBPs have been used as surrogates of nAChRs to study the interaction of different molecules with nAChRs by crystallographic and solution-based techniques due to the similarity of their ACh binding sites 25,31–33. The aim of this study was to develop a microsphere-based assay for the detection of cyclic imines using binding proteins/receptors in a Luminex system, and to characterize its advantages versus previously published receptor-based methods in terms of sensitivity, flexibility, simplicity and assay duration.
MATERIALS AND METHODS
Materials
Biotin-α-bungarotoxin (α-bungarotoxin peptide sequence: IVCHTTATSPISAVTC PPGENLCYRKMWCDAFCSSRGKVVELGCAATCPSKKPYEEVTCCSTDKCNPHP KQRPG, Molecular formula: C338H529N97O105S11) was obtained from Molecular Probes (Eugene, OR). 13-desmethyl spirolide C, 13, 19-didesmethyl spirolide C, 20-methyl spirolide G, okadaic acid and yessotoxin standard solutions were purchased from Laboratorio Cifga (Lugo, Spain). Gymnodimine, saxitoxin, azaspirazid-1 and domoic acid standard solutions were obtained from the Institute for Marine Biosciences, National Research Council (Halifax, Canada). Bovine serum albumin (BSA), Tween-20, and N-hydroxysulfosuccinimide (NHS) were obtained from Sigma-Aldrich (Madrid, Spain) and 1-Ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride (EDC) was obtained from Pierce (Rockford, IL, USA). Acetone, absolute ethanol, sodium chloride, and sodium phosphates were obtained from Panreac (Barcelona, Spain). The nAChR-rich membranes from Torpedo marmorata were purified as previously described 27. Toxin-free scallops (Pecten maximus), clams (Ruditapes philippinarum) and mussels (Mytilus galloprovincialis) were purchased from the market (Lugo, Spain). The acetylcholine binding protein from Lymnaea stagnalis (Ls-AChBP), the acetylcholine binding protein from Aplysia californica (Ac-AChBP) and the Y55W mutant of Ac-AChBP were purified and characterized as described previously 32,34. Calibration and performance verification kits for Luminex 200, carboxylated microspheres (LC10027-01 and LC10004-01) and sheath fluid were from Luminex Corporation (Austin, Texas). PhycoLink Streptavidin-R-Phycoerythrin (PE) was purchased from Prozyme (Hayward, CA, USA). Amicon Ultra-4, PLGC Ultracel-PL membrane, 10-kDa-molecular-weight-cutoff (MWCO) filters, 33 mm Millex filters with a 0.22 μm pore size and Multiscreen 96-well filter plates (Durapore membrane) were obtained from Millipore (Madrid, Spain). Phosphate-buffered saline solution (PBS) was 130 mM NaCl, 1.5 mM NaH2PO4, 8.5 mM Na2HPO4, pH 7.4. PBS-BT was PBS supplemented with 0.1% w/v BSA and 0.1% v/v Tween-20, and PBS-x2BT was PBS supplemented with 0.2% w/v BSA and 0.2% v/v Tween-20. All solutions were filtered through a 0.22 μm pore size filter before use.
Coupling of binding proteins to carboxylated microspheres
Binding proteins were immobilized on the surface of microspheres following the protocol recommended by Luminex for protein coupling to carboxylated microspheres. The immobilization protocol was performed with LC10027-01 or LC10004-01 microspheres (4 × 105 – 1 x 106) in 1.5 mL microtubes. The carboxyl groups were activated by a mixture of 40 μL of EDC/NHS 1:1 (both reagents at concentrations of 50 mg/mL in water) and 160 μL of the activation buffer (0.1 M NaH2PO4, pH 6.2). After 20 min of incubation, the microspheres were centrifuged and the supernatant was carefully discarded. The beads were then washed three times with 500 μL of PBS and 200 μL of binding protein solution were added to pre-activated microspheres and allowed to react for 2 h. Protein dilutions used for the immobilization were 1:40 for nAChR membrane stock (protein concentration of 1.5 mg/mL), 1:200 for Ls-AChBP stock (protein concentration of 6.4 mg/mL), 1:10 and 1:100 for Ac-AChBP and Y55W Ac-AChBP stocks (protein concentrations of 6.5 mg/mL and 7 mg/mL respectively). The protein solutions were sonicated for 10 s before being added to the microsphere suspension. Finally, the microspheres were washed with PBS-BT and stored in PBS-BT at 4 °C in the dark. Removal of reagents and washes were performed by centrifugation at 8000 xg for 2 min at room temperature, and all incubations were performed with constant shaking (200 rpm) at room temperature in the dark.
Microsphere-based assay protocol for detecting cyclic imines
A microsphere-based assay was developed using the competition between α-BTX, which binds to the ACh binding site of the nAChR, and cyclic imines for the interaction with binding proteins attached to the microsphere surface. The assay was performed in a well of an opaque, 1.2 μm filter plate. For each well, 25 WL of the calibration solution or sample were added to 4 × 103 binding protein-coated microspheres (final volume 50 WL) and incubated for 30 min. Then, 50 WL of 50 nM biotin-labeled α-BTX were added to the same well and incubated for 30 min. The buffer was removed by filtration and the microspheres were washed three times with PBS-BT. A volume of 100 WL of 4 μg/mL PE-labeled streptavidin was added to the microspheres for 30 min. Finally, the microspheres were washed and suspended in 100 WL of PBS-BT by shaking immediately before detection. The reagents used in the assay were diluted to the required concentration in PBS-BT. Incubations were performed with constant shaking (700 rpm) at room temperature in the dark. All washes consisted of addition and subsequent removal of 200 WL of PBS-BT. For the removal of reagent and washing solutions, a vacuum manifold was used without exceeding 5 mm Hg of vacuum pressure.
Quantification of the fluorescence signal
PE fluorescence bound to the microsphere surface was measured with a Luminex 200™ analyzer (LuminexCorp, Austin, Texas). The Luminex system uses a 635 nm laser to discriminate between different microsphere classes based on their internal fluorescence. A second 532 nm laser is used to quantify PE-labeled streptavidin fluorescence bound to the microsphere surface. Default values of 7500–13500 were used for doublet discriminator gating of microspheres. The acquisition volume was 75 μL and minimum bead count was 100. Biotin-α-BTX binding response was calculated as follows: , where % R is the percentage of α-BTX binding response, Si is the signal obtained for a given condition, Smax the signal obtained in control wells containing protein-bound microspheres and biotin-labeled α-BTX, and Smin the signal obtained in control wells containing protein-bound microspheres and no biotin-labeled α-BTX.
Shellfish extraction procedure
Shellfish meat (whole body of mussel or clam, and muscle plus gonad of scallop) was homogenized with a blender, divided into aliquots and stored at −20 °C until use. The extraction procedure consisted of the addition of 4 mL of acetone to 1 g of shellfish homogenate. The mixture was vortexed for 10 s and roller mixed for 15 min at room temperature. After centrifugation at 3500 xg for 10 min at 4 °C, the supernatant was saved and the pellet was re-extracted twice with acetone, as described. The combined supernatants were evaporated and the resulting residue was dissolved in 20 mL ethanol:PBS (4:6 v/v) and filtered through a 10 kDa MWCO filter (Millipore). Finally, an equal volume of PBS-2xBT was added. The mussel, clam and scallop homogenates used for this study were previously analyzed by LC-MS/MS for the presence of 13-desmethyl spirolide C, 13, 19-didesmethyl spirolide C, 20-methyl spirolide C and gymnodimine as described previously 28, to be sure that the shellfish samples did not contain these compounds at levels at least 10 times lower than the LoD of the Luminex assay.
Safety
Cyclic imines are fast-acting toxins with very high acute intraperitoneal toxicity in mice. Although no human intoxication has been reported, standard solutions should be handled with gloves and eye protection should be worn at all times. Appropriate disposal methods should also be utilized.
Data analysis
The results were expressed as mean ± SEM (standard error of the mean) and in every experiment each condition was tested in duplicate. The calibration curves were fitted using a four-parameter logistic equation obtained by a nonlinear regression fitting procedure (GraphPad Prism 5.0). The four-parameter fit equation was: , where Min is the signal in the absence of ligand, Max is the maximum binding signal at saturating ligand, logEC50 is the concentration when the response is halfway between Min and Max and nH is the Hill Slope. Statistical analysis for multiple comparisons was performed using ANOVA (p < 0.05). The limit of detection (LoD) was calculated by subtracting 3 times the standard deviation (SD) of raw binding signals of the highest point of the calibration curve from the maximum binding signal and interpolating this value in the calibration curve.
RESULTS
Selection of binding proteins for the microsphere-based assay
The feasibility of using alternative binding proteins in a microsphere-based assay for the detection of cyclic imines using α-BTX competition was explored in order to select optimal binding characteristics. The proteins tested were nAChRs from the Torpedo marmorata electric organ, AChBP from the freshwater snail Lymnaea stagnalis (Ls-AChBP), AChBP from the saltwater mollusk Aplysia californica (Ac-AChBP) and the Y55W mutant of Ac-AChBP. Ls-AChBP, nAChR, Ac-AChBP and Y55W A-AChBP were immobilized on the microsphere surface and binding of 13-desmethyl spirolide C was evaluated by inhibition of α-BTX binding to the protein-coated surface of the microspheres. The Ac-AChBP and Y55W-coated microspheres did not show a sufficient α-BTX binding signal under these conditions to perform a competition assay (data not shown), probably due to a much lower affinity for α-BTX than other AChBPs 32,34, and therefore they were not suitable for this assay design. In contrast, Ls-AChBP- and nAChR-coated microspheres showed excellent α-BTX binding signals, with maximum binding signal to background ratios around 2000 and 500 respectively, under these assay conditions. Ls-AChBP and nAChR-immobilized microspheres were then used to obtain 13-desmethyl spirolide C calibration curves. The solvent of the 13-desmethyl spirolide C standard stock solution, methanol-TFA (Trifluoroacetic Acid), was evaporated, since methanol has been previously shown to interfere with binding properties of nAChR-rich membrane fragments 27, and 13-desmethyl spirolide C was dissolved in PBS-BT containing 20% ethanol (PBS-BT/20% EtOH). Serial dilutions of 13-desmethyl spirolide C at concentrations of 0.01, 0.1, 1, 10, 100 and 500 nM were prepared in PBS-BT/20% EtOH (all assay wells contained the same final concentration of ethanol in order to ensure cyclic imine toxin solubility) and assayed using the nAChR and Ls-AChBP-based assays. 13-desmethyl spirolide C inhibited binding of α-BTX to both proteins with similar IC50 values (Figure 2). However, the dynamic range (IC20- IC80) for 13-desmethyl spirolide C was considerably wider for the nAChR assay than for the Ls-AChBP assay (Figure 2B and 2D).
Figure 2.

Cross-reactivity profiles for the detection of 13-desmethyl spirolide C, 13, 19-didesmethyl spirolide C, 20-methyl spirolide G, and gymnodimine with the nAChR- and Ls-AChBP-based inhibition assays. The data are expressed as percentage of biotin-α-BTX maximum binding response. The calibration curves were obtained with a four-parameter fit. (A) Calibration curves for the spirolides and gymnodimine with the nAChR inhibition assay. The four curves belong to non-matched experiments (mean ± SEM; 13-desmethyl spirolide C, n = 6; 13, 19-didesmethyl spirolide C, n = 3; 20-methyl spirolide G, n = 3 and gymnodimine, n = 3). (B) IC50, dynamic range and cross- reactivity (%) of the nAChR-microsphere-based assay were obtained from calibration curves in (A). (C) Calibration curves for the spirolides and gymnodimine with the Ls-AChBP inhibition assay. The four curves belong to non-matched experiments (mean ± SEM; 13-desmethyl spirolide C, n = 9; 13, 19-didesmethyl spirolide C, n = 3; 20-methyl spirolide G, n = 4 and gymnodimine, n =2). (D) IC50, dynamic range and cross- reactivity (%) of the Ls-AChBP-microsphere based assay from calibration curves in (C).
Cross reactivity profile of the nAChR and L-AChBP assays
Cross-reactivities of the nAChR and L-AChBP assays were evaluated by comparing the inhibition of α-BTX binding by several members of the macrocyclic imine toxins. Calibration curves of 13-desmethyl spirolide C, 13, 19-didesmethyl spirolide C, 20-methyl spirolide G and gymnodimine were obtained using nAChR or Ls-AChBP-based competition assays in buffer (PBS-BT/20% EtOH). The concentrations of the calibration solutions for the four toxins ranged from 0.01 to 500 nM. All the phycotoxins tested inhibited binding of α-BTX to the surface of the two classes of protein-coated microspheres (Figure 2A and 2C). The IC50 of the calibration curves was used to evaluate the cross-reactivity of the assays for these compounds (Figure 2B and 2D). The cross-reactivity of every toxin tested was expressed related to 13-desmethyl spirolide C reactivity and calculated as follows: % Cross-reactivity (% CR) = (IC50 of 13-desmethyl spirolide C / IC50 of cyclic imine) x 100. The results showed that the cross-reactivity profile of cyclic imine binding to nAChR- and Ls-AChBP-coated microspheres differed considerably. The most remarkable difference was the efficient binding of gymnodimine relative to spirolides to Ls-AChBP-coated microspheres (Figure 2C and 2D), while gymnodimine binding in the nAChR assay was weaker than spirolide binding (Figure 2A and 2B). After the evaluation of the assay characteristics, the nAChR-based assay was selected for further development based on its dynamic range and cyclic imine cross-reactivity profile. The interference of representative toxins of the regulated toxin groups with the nAChR-based assay was tested using 10 μM saxitoxin, 10 μM okadaic acid, 10 μM domoic acid, 5 μM yessotoxin and 1 μM azaspirazid-1, and no effects on maximum and minimum binding signals were evidenced.
Additionally, the Luminex system offers the possibility of multi-detection, and therefore nAChR- and Ls-AChBP-coated microspheres were combined in the same well to explore the performance of both assays simultaneously, in order to provide a higher sensitivity for gymnodimine if desirable. The combination of nAChR-coated microspheres (LC-10027-01) and Ls-AChBP-coated microspheres (LC-10004-01) in the same well yielded calibration curves, similar to the individual assays (data not shown).
Effect of shellfish matrixes on the nAChR/microsphere assay
The compatibility of this nAChR/microsphere-based assay with shellfish extracts was assessed using mussels, scallops and clams. In order to evaluate the matrix effect on spirolide detection, calibration curves of 13-desmethyl spirolide C, and maximum and minimum binding signals of the assay in buffer and in shellfish extracts were compared. The extracts were prepared as described in the Methods section, starting with an acetone extraction followed by evaporation and reconstitution in ethanol/buffer and filtration. The presence of 20 % ethanol in the final extract did not affect the assay, since this concentration of ethanol was present in all assay conditions tested in this study, as reported in the previous sections. For 13-desmethyl spirolide C calibration curves, the calibration solutions were prepared as previously described in buffer or shellfish extract, and assayed using the nAChR/microsphere-based assay. The calibration curves obtained in clam, mussel and scallop extracts and in buffer were virtually identical (Figure 3A). The dynamic ranges, estimated LoDs and IC50 values calculated from the calibration curves were very similar in buffer and shellfish extract (Table 1). Maximum and minimum binding signals of the nAChR/microsphere-based assay performed with mussel, scallop or clam extracts, did not differ significantly from those obtained with buffer (Figure 3B). The 13-desmethyl spirolide C IC50 values obtained in mussel, scallop and clam extracts would correspond to levels of 451.3 ± 42.4, 344.2 ± 60.3, and 411.6 ± 54.8 Wg/kg of shellfish meat, assuming 100% recoveries. During the optimization of the extraction procedure, methanol was also tested as extraction solvent instead of acetone, and the interference of the different shellfish extracts (mussels, scallops or clams) obtained with this alternative extraction method was also very low in preliminary experiments (data not shown). However, the acetone extraction protocol was selected for future studies in order to shorten the duration of the assay, due to its rapid evaporation.
Figure 3.
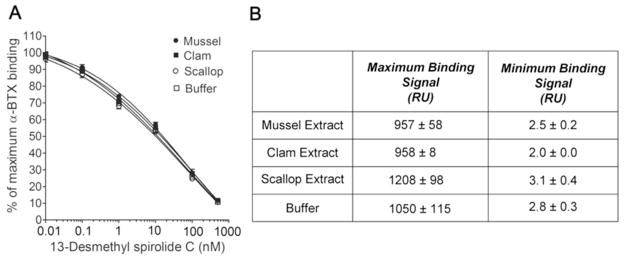
Effect of mussel, clam and scallop matrices on the detection of 13-desmethyl spirolide C using the nAChR/microsphere-based assay. (A) Calibration curves of 13-desmethyl spirolide C in mussel, clam and scallop extracts and in buffer using the nAChR inhibition assay. The data are expressed as percentage of biotin-α-BTX maximum binding response. The calibration curves were obtained with a four-parameter fit. The four curves belong to non-matched experiments (mean ± SEM; mussel, n = 6; clam, n = 2; scallop, n =5 and buffer, n = 8). (B) Maximum and minimum binding signals in shellfish extracts and buffer from the experiments in (A).
Table 1.
IC50, dynamic range and estimated LoD of the nAChR/microsphere-based assay for 13-desmethyl spirolide C detection in shellfish extracts and buffer. The IC50, dynamic range and LoDs correspond to the experiments that generated figure 3A (mean ± SEM; mussel, n = 6; clam, n = 2; scallop, n =5; and buffer, n =8).
| Mussel | Clam | Scallop | Buffer | |
|---|---|---|---|---|
| IC50 (nM) IC50 (ng/mL) |
16.3 ± 1.5 11.3 ± 1.1 |
14.9 ± 1.9 10.3 ± 1.4 |
12.4 ± 2.2 8.6 ± 1.5 |
10.8 ± 0.9 7.4 ± 0.7 |
| IC20 (nM) IC20(ng/mL) |
0.6 ± 0.2 0.4 ± 0.1 |
0.5 ± 0.01 0.3 ± 0.01 |
0.5 ± 0.2 0.4 ± 0.2 |
0.4 ± 0.07 0.2 ± 0.04 |
| IC80 (nM) IC80(ng/mL) |
232 ± 11 160 ± 8 |
231 ± 23 160 ± 6. |
201 ± 9 139 ± 6.5 |
1998 ± 8.0 137 ± 5 |
| LoD (nM) LoD(ng/mL) |
0.1 ± 0.05 0.08 ± 0.03 |
0.1 ± 0.08 0.07 ± 0.05 |
0.05 ± 0.02 0.04 ± 0.02 |
0.1 ± 0.03 0.08 ± 0.02 |
Recovery of 13-desmethyl spirolide C with the nAChR/microsphere-based assay
The recovery of 13-desmethyl spirolide C using a simple acetone extraction protocol combined with detection by the nAChR/microsphere-based assay was evaluated in samples of different shellfish species. Blank samples of mussel, scallop and clam were spiked with 13-desmethyl spirolide C. Six 1 g aliquots of shellfish homogenate (6 per species) were contaminated with 280 ng (280 Wg/kg) of 13-desmethyl spirolide C, extracted and analyzed with the nAChR-based assay within the same day. The content of toxin was calculated using calibration curves in extract. Recoveries are shown as a percentage: % Recovery = 100 x (measured content / fortification level). The amount of 13-desmethyl spirolide C detected with the assay was 209.5 ± 4.0, 204.1 ± 0.7 and 220.3 ± 13.3 Wg/kg in mussels, clams and scallops resulting in recovery efficiencies of 74.8 ± 1.4 %, 72.9 ± 0.3 % and 78.7 ± 4.8 % respectively (mean ± SEM of 6 replicates).
Repeatability of the spirolide-detection assay
The intra-assay and inter-assay repeatabilities of our method were determined for the detection of 13-desmethyl spirolide C in spiked shellfish samples. Intra-assay repeatability was evaluated for mussel, scallop and clam and inter-assay repeatability was evaluated only for mussel. Aliquots of mussel, scallop or clam homogenate (6 x 1 g) were spiked with 280 ng of 13-desmethyl spirolide C, extracted and assayed. The procedure was repeated in two different days for mussels. Then the extracts were analyzed with the nAChR assay and the amount of 13-desmethyl spirolide C was quantified using a calibration curve in shellfish extract. The coefficients of variation (CV) within the same run (intra-assay) were 11.3 %, 20.5 % and 8.5 % for mussel, scallop and clam respectively, and between runs (inter-assay) repeatability for mussels was 6.7 %.
Stability of 13-desmethyl spirolide C in buffer, shellfish extracts and mussel homogenate
The stability of 13-desmethyl spirolide C was evaluated in buffer and shellfish extracts stored at 4–8 ºC or -20 ºC for 2 or 4 weeks. The 13-desmethyl spirolide C was diluted in buffer (PBS-BT/20% EtOH) or extract (mussel and scallop) at a concentration of 7 ng/mL, divided in 30 aliquots and stored at 4–8 ºC (12 aliquots) or -20 ºC (12 aliquots). The 6 remaining aliquots were analyzed on the same day. Half of the aliquots stored at 4–8 ºC or -20 ºC were further analyzed after two weeks, and the other half after 4 weeks of storage. At the indicated times, the amount of 13-desmethyl spirolide C was detected with the nAChR/microsphere-based assay using freshly prepared calibration curves in buffer, or shellfish extract (mussel or scallop) as required. These samples showed no significant degradation of 13-desmethyl spirolide C after 4 weeks of storage at −20 ºC or 4 ºC (Table 2).
Table 2.
Stability of 13-desmethyl spirolide C in buffer and shellfish extracts (mussel and scallop) for 4 weeks at different storage conditions. The stability was expressed as a percentage of toxin detected in samples after storage versus freshly analyzed samples. Aliquots of buffer (PBS-BT/20% EtOH) and shellfish extracts were spiked with 7 ng/ml of 13-desmethyl spirolide C, and analyzed fresh or stored over four weeks at 4 °C or -20 °C. After this time period the stored samples were assayed using freshly prepared calibrants (mean ± SEM, n= 6).
| % Recovery | Buffer | Mussel Extract | Scallop Extract |
|---|---|---|---|
| 4–8 °C | 109 ± 6 | 135 ± 23 | 92.0 ± 9.2 |
| −20 °C | 98.1 ± 6.3 | 121 ± 16 | 85.5 ± 5.5 |
The stability of 13-desmethyl spirolide C in mussel homogenate was also analyzed. For this purpose three aliquots of 1 g of mussel homogenate were spiked with 13-desmethyl spirolide C, one aliquot was analyzed fresh and the other two were stored at −20 °C for 1 and 2 weeks. After these time periods the mussel homogenate was thawed and extracted to measure the amount of toxin using the nAChR/microsphere-based assay. The calibration curve was freshly prepared in mussel extract. After two weeks of storage in the freezer 105.7 ± 5.3 % of 13-desmethyl spirolide C was recovered.
DISCUSSION
In this study a solid-phase receptor-based assay was developed and pre-validated for the detection of cyclic imines using the xMap Luminex technology. This assay is based on the competition between cyclic imines and α-BTX for the interaction with a binding protein immobilized on the surface of microspheres. Two different binding proteins known to interact with spirolides, the Torpedo marmorata nAChR and the Lymnaea stagnalis AChBP 2,25 were considered suitable for assay development. While both proteins bound efficiently to 13-desmethyl spirolide C, the nAChR provided a much wider dynamic range for this toxin and a cross-reactivity profile that matched better the relative toxicity of 13-desmethyl spirolide C and gymnodimine. The broader dynamic range of the nAChR assay is probably a consequence of the non-identical binding sites at the αγ and αδ subunit interfaces of the Torpedo marmorata electric organ nAChR (heteropentamer α2β d) for these competing ligands, similar to what has been previously reported for other muscle nAChR antagonists with apparent Hill coefficients less than 1.0 35–38. The AChBP, instead, is a homomeric pentamer with identical subunit interfaces and therefore binding sites 32. Although, AChBP characteristics provide some advantages such as sharp concentration dependence for accurate measurements of affinity and a greater homogeneity and stability of the binding protein preparation, the wide dynamic range of the nAChR assay is a valuable feature for screening multiple toxins.
Regarding cross-reactivity, in our hands the Ls-AChBP assay is 14-fold more efficient for gymnodimine than 13-desmethyl spirolide C. On the contrary, the nAChR assay detects 13-desmethyl spirolide C 24 times more efficiently than gymnodimine. Actually, since AChBP is considered a structural homologue of the extracellular domain of the α7 nAChR subtype 39, the higher sensitivity of our AChBP assay for gymnodimine is consistent with previous reports of higher affinity of gymnodimine for the chimeric protein a7-5HT3 than for the T. marmorata nAChR 26. The LD50 of gymnodimine and 13-desmethyl spirolide C fed to fasted mice has been reported as >7500 μg/kg and 500 μg/kg respectively 40,41, which means that gymnodimine would be >15 times less toxic than the spirolide by this route of administration. The cross-reactivity data for 13, 19-didesmethyl spirolide C should be considered as preliminary estimations, since to date there are no certified standard solutions available for this compound worldwide. A certified standard solution of 20-methyl spirolide G has been recently released to the market and has been used for cross-reactivity profiling in this study. The 95% cross-reactivity obtained for 20-methyl spirolide G agrees with previous toxicity studies performed in mice with 13-desmethyl spirolide C and 20-methyl spirolide G, with very similar oral LD50 values of 160 Wg/kg by gavage and 500 Wg/kg when administered with food for both toxins 41. However, some of these toxicity studies were performed before certified standards of these toxins were produced and therefore, the correlation between assay cross-reactivity and relative toxicity of these compounds should be reviewed when more toxicological data and standard solutions are available, So far, our results clearly demonstrate that the nAChR assay can detect the three spirolides and gymnodimine with an acceptable correlation with toxicity data currently available. Therefore the nAChR was chosen as the binding protein for subsequent assay development and pre-validation taking into consideration assay cross-reactivity and dynamic range.
Additionally, in the Luminex system the different cross-reactivities of nAChR and L-AChBP assays can be used to generate a multiplexed assay with additional higher sensitivity towards gymnodimine than the single plex nAChR assay. The multiplexed alternative does not seem necessary at the moment since macrocyclic imines have not been regulated yet. However, some investigators in the marine toxin field regard the spirolides as potentially more toxic, while gymnodimine is generally considered a lesser threat. In case the presence of spirolides in seafood, and not the gymnodimines, become regulated, a multiplexed assay would be useful to discriminate between them.
The characterization of the nAChR/Luminex microsphere assay performance was conducted following the recommendations for method validation of the European Decision 2002/657/EC 42 as far as the availability of toxins and blank shellfish samples would allow. The nAChR/Luminex microsphere-based assay provides higher sensitivity for the 13-desmethyl spirolide C (IC50 = 10 nM) than previously published competition assays based also on interactions with the nAChR, such as the chemiluminescence assay (IC50 = 35 nM) and the fluorescence polarization assay (IC50 = 100 nM) 27,28. Another characteristic of the Luminex assay is a wider dynamic range than other receptor-based assays 27–29. The increased sensitivity of the Luminex-based method for spirolide detection has allowed an important improvement versus other receptor-based competition assays for cyclic imines previously published: the elimination of matrix interferences by higher dilutions, allowing the use of a simple and fast extraction method. The extraction method optimized for this assay consists of an extraction with acetone, reconstitution in ethanol/PBS and filtration, which implies the elimination of several solvent partitions needed to clean extracts for other assays 27,28. The new extraction method is much shorter and less laborious, permiting the simultaneous extraction of a higher number of samples. The recovery rates of the new method also appear improved, probably due to the reduction of the number of extraction steps.
The nAChR microsphere-based assay in combination with this sample preparation protocol allows the detection of the 13-desmethyl spirolide C in three different shellfish extracts, mussels, clams and scallops, in the range of 10–6000 μg/kg of shellfish meat, a wider detection range than the previous chemiluminescent assay (40–1000 μg/kg) 28, and a LoD of 3 μg/kg. LoD has been used to report sensitivity instead of CCα and CCβ due to the impossibility to obtain enough blank shellfish matrixes and purified toxin to perform the experiments required for CCα and CCβ determination. The assay performance for the detection of 13-desmethyl spirolide C in spiked shellfish samples was deemed adequate in terms of recovery, with values around 75 %, and repeatability, with intra and inter-assay CVs lower than 12 %, except for scallop matrix. Although the toxicity of spirolides to humans is still unknown, and no regulatory limits have been defined for these toxins, a safe level of spirolide content in shellfish has been estimated by some investigators as 400 μg/kg 43, a value within the range of our assay. The stability of spirolides in buffer and shellfish extracts as well as in mussel homogenates stored at −20 °C or 4 °C during four weeks makes it feasible to include in a single assay samples received in the lab at different times.
The nAChR/Luminex -based assay can be used as a semi-quantitative method to screen shellfish samples for the presence of cyclic imines, yielding an overall estimation of cyclic imine-related sample toxicity based on binding to the nAChR. Moreover, representative toxins of other groups, saxitoxin, okadaic acid, yessotoxin, domoic acid and azaspiracid-1 do not interfere with the Luminex nAChR-based assay. In previous studies, nAChR based assays have not shown any interferences with saxitoxin, okadaic acid, brevetoxin-2, yessotoxin and azaspiracid-2 27,28.
The Luminex system has been developed as a multiplex detection technology that would allow simultaneous detection of several classes of analytes in the same sample. This cyclic imine-detection assay could be included in the future in a multi-detection assay for several groups of marine toxins. In this regard, the compatibility with methanol extraction in this nAChR assay is useful information, since acetone is not a commonly used solvent to extract other marine toxin groups. However, recoveries of cyclic imines with methanol extraction would have to be tested in the Luminex assay and the multiplexed assay optimized due to the possibility of unexpected interactions appearing when combining several competition assays 44.
In summary, this nAChR/Luminex-based assay provides a suitable system to screen shellfish samples for the presence of cyclic imines. This new method is capable of detecting concentrations of 13-desmethyl spirolide C above 3 Wg/kg. This offers a screening method that is more sensitive, faster and easier to perform than previously published receptor-based assays. Moreover, this technology affords the possibility of including spirolide detection in future toxin multidetection assays.
Acknowledgments
This work was funded with the following FEDER cofunded-grants from: Ministerio de Ciencia y Tecnología, Spain: SAF2009-12581 (subprograma NEF), AGL2009 13581-CO2-01, TRA2009-0189, AGL2010-17875, Xunta de Galicia, Spain: GRC 2010/10, and PGDIT 07MMA006261PR, PGIDIT (INCITE) 09MMA003261PR, PGDIT (INCITE) 09261080PR, 2009/XA044, and 10PXIB261254 PR, EU VIIth Frame Program: 211326 – CP (CONffIDENCE), 265896 BAMMBO, 265409 WAQUA, and 262649 BEADS, 312184 PharmaSea, RO-1 GM18360-40 National Institutes of Health, U.S.A.,.and the Atlantic Area Programme (Interreg IVB Trans-national): 2009-1/117 Pharmatlantic.
Abbreviations
- AChBP
acetylcholine binding protein
- Ac-AChBP
acetylcholine binding protein from Aplysia californica
- BSA
bovine serum albumin
- α-BTX
α-bungarotoxin
- CR
cross-reactivity
- CV
coefficient of variation
- DSP
diarrheal shellfish poisoning
- EDC
1-Ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride
- i.p
intraperitoneal
- Ls-AChBP
acetylcholine binding protein from Lymnaea stagnalis
- LC-MS
liquid chromatography-mass spectrometry
- LoD
limit of detection
- MS
mass spectrometry
- MBA
mouse bioassay
- MWCO
molecular-weight-cutoff
- nAChR
nicotinic acetylcholine receptor
- NHS
N-hydroxysulfosuccinimide
- PBS
phosphate-buffered saline
- PE
phycoerythrin
- SD
standard deviation
- SEM
standard error of the mean
- TFA
trifluoroacetic acid
References
- 1.Biré R, Krys S, Frémy JM, Dragacci S, Stirling D, Kharrat R. J Nat Toxins. 2002;11:269–275. [PubMed] [Google Scholar]
- 2.Molgó J, Girard E, Benoit E. In: Phycotoxins: Chemistry and Biochemistry. Botana LM, editor. Blackwell Ames; 2007. pp. 319–335. [Google Scholar]
- 3.Takada N, Umemura N, Suenaga K, Chou T, Nagatsu A, Haino T, Yamada K, Uemura D. Tetrahedron Lett. 2001;42:3491–3494. [Google Scholar]
- 4.Uemura D, Chou T, Haino T, Nagatsu A, Fukuzawa S, Zheng SZ, Chen HS. J Am Chem Soc. 1995;117:1155–1156. [Google Scholar]
- 5.Takada N, Umemura N, Suenaga K, Uemura D. Tetrahedron Letters. 2001;42:3495–3497. [Google Scholar]
- 6.Torigoe K, Murata M, Yasumoto T. J Am Chem Soc. 1988;110:7876–7877. [Google Scholar]
- 7.Lu CK, Lee GH, Huang R, Chou HN. Tetrahedron Lett. 2001;42:1713–1716. [Google Scholar]
- 8.Hu T, Curtis JM, Oshima Y, Quilliam MA, Walter JA, Watson-Wright WM, Wright JLC. J Chem Soc, Chem Commun. 1995;20:2159–2161. [Google Scholar]
- 9.Cembella AD, Lewis NI, Quilliam MA. Nat Toxins. 1999;7:197–206. doi: 10.1002/1522-7189(200009/10)7:5<197::aid-nt62>3.0.co;2-h. [DOI] [PubMed] [Google Scholar]
- 10.Cembella AD, Lewis NI, Quilliam MA. Phycologia. 2000;39:67–74. [Google Scholar]
- 11.Touzet N, Franco JM, Raine R. Appl Environ Microbiol. 2008;7:782–797. doi: 10.1128/AEM.02161-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Haywood AJ, Steidinger KA, Truby EW, Bergquist PR, Bergquist PL, Adamson J, Mackenzie L. J Phycol. 2004;40:165–179. [Google Scholar]
- 13.Seki T, Satake M, Mackenzie L, Kaspar HF, Yasumoto T. Tetrahedron Lett. 1995;36:7093–7096. [Google Scholar]
- 14.Richard D, Arsenault E, Cembella A, Quilliam M. In: Harmful Algal Blooms 2000: Proceedings of the Ninth International Conference on Harmful Algal Blooms. Hallegraeff GM, Blackburn SI, Bolch CJ, Lewis RJ, editors. Intergovernmental Oceanographic Commission of UNESCO; Paris: 2001. pp. 383–390. [Google Scholar]
- 15.Gill S, Murphy M, Clausen J, Richard D, Quilliam M, MacKinnon S, LaBlanc P, Mueller R, Pulido O. Neurotoxicology. 2003;24:593–604. doi: 10.1016/S0161-813X(03)00014-7. [DOI] [PubMed] [Google Scholar]
- 16.Botana LM. In: Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection. 2. Botana LM, editor. CRC Press; Boca Raton, FL: 2008. pp. 149–161. [Google Scholar]
- 17.Holland PT. In: Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection. 2. Botana LM, editor. CRC Press; Boca Raton, FL: 2008. pp. 21–51. [Google Scholar]
- 18.Aasen J, MacKinnon SL, LeBlanc P, Walter JA, Hovgaard P, Aune T, Quilliam MA. Chem Res Toxicol. 2005;18:509–15. doi: 10.1021/tx049706n. [DOI] [PubMed] [Google Scholar]
- 19.Ciminiello P, Dell'Aversano C, Fattorusso E, Magno S, Tartaglione L, Cangini M, Pompei M, Guerrini F, Boni L, Pistocchi R. Toxicon. 2006;47:597–604. doi: 10.1016/j.toxicon.2006.02.003. [DOI] [PubMed] [Google Scholar]
- 20.Fux E, McMillan D, Biré R, Hess P. J Chromatogr A. 2007;1157:273–280. doi: 10.1016/j.chroma.2007.05.016. [DOI] [PubMed] [Google Scholar]
- 21.Hu T, Burton IW, Cembella AD, Curtis JM, Quilliam MA, Walter JA, Wright JL. J Nat Prod. 2001;64:308–12. doi: 10.1021/np000416q. [DOI] [PubMed] [Google Scholar]
- 22.Sleno L, Volmer DA. Anal Chem. 2005;77:1509–17. doi: 10.1021/ac0486600. [DOI] [PubMed] [Google Scholar]
- 23.Stirling DJ. N Z J of Mar and Freshwater Res. 2001;35:851–857. [Google Scholar]
- 24.Villar Gonzalez A, Rodriguez-Velasco ML, Ben-Gigirey B, Botana LM. Toxicon. 2006;48:1068–74. doi: 10.1016/j.toxicon.2006.09.001. [DOI] [PubMed] [Google Scholar]
- 25.Bourne Y, Radic Z, Aráoz R, Talley TT, Benoit E, Servent D, Taylor P, Molgó J, Marchot P. Proc Natl Acad Sci U S A. 2010;107:6076–81. doi: 10.1073/pnas.0912372107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kharrat R, Servent D, Girard E, Ouanounou G, Amar M, Marrouchi R, Benoit E, Molgó J. J Neurochem. 2008;107:952–63. doi: 10.1111/j.1471-4159.2008.05677.x. [DOI] [PubMed] [Google Scholar]
- 27.Vilariño N, Fonfría ES, Molgó J, Aráoz R, Botana LM. Anal Chem. 2009;81:2708–14. doi: 10.1021/ac900144r. [DOI] [PubMed] [Google Scholar]
- 28.Rodríguez LP, Vilariño N, Molgó J, Aráoz R, Antelo A, Vieytes MR, Botana LM. Anal Chem. 2011;83:5857–63. doi: 10.1021/ac200423s. [DOI] [PubMed] [Google Scholar]
- 29.Otero P, Alfonso A, Alfonso C, Aráoz R, Molgó J, Vieytes MR, Botana LM. Anal Chim Acta. 2011;701:200–208. doi: 10.1016/j.aca.2011.05.034. [DOI] [PubMed] [Google Scholar]
- 30.Kellar KL, Iannone MA. Exp Hematol. 2002;30:1227–1237. doi: 10.1016/s0301-472x(02)00922-0. [DOI] [PubMed] [Google Scholar]
- 31.Brejc K, Van Dijk WJ, Klaassen RV, Schuurmans M, Van der Oost J, Smit AB, Sixma TK. Nature. 2001;411:269–276. doi: 10.1038/35077011. [DOI] [PubMed] [Google Scholar]
- 32.Hansen SB, Radif Z, Talley TT, Molles BE, Deerinck T, Tsigelny I, Taylor P. J Biol Chem. 2002;277:41299–41302. doi: 10.1074/jbc.C200462200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rucktooa P, Smit AB, Sixma TK. Biochem Pharmacol. 2009;78:777–787. doi: 10.1016/j.bcp.2009.06.098. [DOI] [PubMed] [Google Scholar]
- 34.Hansen SB, Talley TT, Radic Z, Taylor P. J Biol Chem. 2004;279:24197–24202. doi: 10.1074/jbc.M402452200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sine SM, Taylor P. J Biol Chem. 1979;254:3315–3325. [PubMed] [Google Scholar]
- 36.Sine SM, Taylor P. J Biol Chem. 1980;255:10144–10156. [PubMed] [Google Scholar]
- 37.Sine SM, Taylor P. J Biol Chem. 1981;256:6692–6699. [PubMed] [Google Scholar]
- 38.Weiland G, Taylor P. Mol Pharmacol. 1979;15:197–212. [PubMed] [Google Scholar]
- 39.Ulens C, Akdemir A, Jongejan A, van Elk R, Bertrand S, Perrakis A, Leurs R, Smit AB, Sixma TK, Bertrand D, de Esch IJP. J Med Chem. 2009;52:2372–2383. doi: 10.1021/jm801400g. [DOI] [PubMed] [Google Scholar]
- 40.Munday R, Towers NR, Mackenzie L, Beuzenberg V, Holland PT, Miles CO. Toxicon. 2004;44:173–178. doi: 10.1016/j.toxicon.2004.05.017. [DOI] [PubMed] [Google Scholar]
- 41.Munday R, Quilliam MA, LeBlanc P, Lewis N, Gallant P, Sperker SA, Ewart HS, MacKinnon SL. Toxins. 2012;4:1–14. doi: 10.3390/toxins4010001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Off EC. J Eur communities. 2002;L221:8. [Google Scholar]
- 43.Munday R. In: Seafood and freshwater toxins: Pharmacology, Physiology and Detection. 2. Botana LM, editor. CRC Press; Boca Raton, FL: 2008. pp. 581–594. [Google Scholar]
- 44.Peters J, Bienenmann-Ploum M, de Rijk T, Haasnoot W. Mycotoxin Res. 2011;27:63–72. doi: 10.1007/s12550-010-0077-0. [DOI] [PMC free article] [PubMed] [Google Scholar]