

Abstract
Dengue viruses 1–4 (DENV1-4) rely heavily on the host cell machinery to complete their life cycle, while at the same time evade the host response that could restrict their replication efficiency. These requirements may account for much of the broad gene-level changes to the host transcriptome upon DENV infection. However, host gene function is also regulated through transcriptional start site (TSS) selection and post-transcriptional modification to the RNA that give rise to multiple gene isoforms. The roles these processes play in the host response to dengue infection have not been explored. In the present study, we utilized RNA sequencing (RNAseq) to identify novel transcript variations in response to infection with both a pathogenic strain of DENV1 and its attenuated derivative. RNAseq provides the information necessary to distinguish the various isoforms produced from a single gene and their splice variants. Our data indicate that there is an extensive amount of previously uncharacterized TSS and post-transcriptional modifications to host RNA over a wide range of pathways and host functions in response to DENV infection. Many of the differentially expressed genes identified in this study have previously been shown to be required for flavivirus propagation and/or interact with DENV gene products. We also show here that the human transcriptome response to an infection by wild-type DENV or its attenuated derivative differs significantly. This differential response to wild-type and attenuated DENV infection suggests that alternative processing events may be part of a previously uncharacterized innate immune response to viral infection that is in large part evaded by wild-type DENV.
Author Summary
Dengue is the most common insect-borne viral disease globally. The continued absence of an effective therapy stems from an incomplete understanding of disease pathogenesis, of which the host response to infection is thought to play a central role. While previous studies have described the changes in total gene expression with dengue virus infection, they have not been able to provide any information on the subtle variations of the host RNA. These variations lead to the production of gene isoforms that can have a profound effect on gene function. In the current study, we have used the newly developed technique of RNA sequencing to more accurately interrogate the variations in the host RNA after infection with a wild-type dengue virus or its attenuated derivative. Findings from this study show that there is an extensive amount of previously uncharacterized variation in host RNA response to dengue infection. The response to infection with the wild-type dengue also differs significantly from infection with the vaccine strain. This suggests that variations in the host RNA comprise a part of the host response to viral infection that is in large part evaded by wild-type dengue viruses.
Introduction
Dengue viruses 1–4 (DENV1-4) are the world's most prevalent arthropod-borne viruses [1]. DENVs are responsible for an estimated 50–100 million cases of debilitating or life-threatening infection every year and an estimated 2.5 billion people in over 100 endemic countries are at risk of infection [1], [2]. The economic impact of DENVs has been estimated to be as high, if not higher than other major global health menaces such as malaria, tuberculosis, hepatitis, bacterial meningitis and others [3]–[8]. Despite the considerable health and economic impact, there are as yet no licensed vaccines or antiviral drugs to combat DENVs and an incomplete understanding of the biology of DENV infection has hampered progress on both of these fronts.
Given the limited coding capacity of their ∼11 kb RNA genome, DENVs must parasitize the host cell machinery to complete their life cycle. At the same time, these viruses must effectively evade or suppress the host responses that act to restrict their replication [9]–[11]. This interplay between host and virus and the effect it has on host gene expression has been described previously [12]–[29]. Largely uncharacterized, however, is whether the transcriptional start site (TSS) and post-transcriptional variations of host RNA, leading to the production of different gene isoforms, may play a role in DENV infection. Differential RNA processing is known to be a major factor underlying cellular and functional complexity [30], [31]. In order to interrogate TSS and post-transcriptional RNA variations across the entire genome in response to DENV infection, we harnessed the power of RNA sequencing (RNAseq). RNAseq is a recently developed approach to transcriptome profiling that permits a precise quantification of RNA levels and their alternatively processed variants by means of high throughput, massively parallel sequencing and subsequent mapping of the resultant short sequence fragments onto a reference genome [32], [33].
We utilized two strains of DENV1 in our RNAseq study to identify strain-specific TSS and post-transcriptional variations in response to infection. The first strain, DENV1-16007, was isolated from the serum of a patient in Thailand in 1964. The second strain of DENV1 used in this study is an attenuated derivative of DENV1-16007. This attenuated virus, DENV1-PDK13 was passaged 13 times in primary dog kidney cells and was shown to be immunogenic but minimally reactogenic in human volunteers [34], [35]. Our RNAseq data indicate that significant differences exist between these two strains of DENV1, not only at the transcript level but also at the level of alternative splicing. Similar trends were observed in RNAseq of an additional two low-passaged DENV1 clinical isolates. These findings suggest that subversion of the host response includes TSS and post-transcriptional modification and is part of the mechanism of virulence. These findings also suggest that variations in the viral genome can have a profound effect in modifying host response to infection.
Materials and Methods
Cells and virus stock
HuH7, C6/36 and BHK-21 cells were purchased from the American Type Culture Collection (ATCC) and cultured according to ATCC recommendation. DENV1 strains 16007 and PDK13 were obtained from the Division of Vector-borne Diseases, Centers for Disease Control and Prevention. Sequence analysis in our laboratory indicates that these strains match the published sequences for these viruses (GenBank accession numbers AF180817.1 and AF180818.1, respectively). These viruses were amplified three times in C6/36 cells prior to use in the current study. Additionally, the DENV1 clinical isolates EDEN3300 and SL107, which were obtained from previously reported studies [36], [37] and passaged in C6/36 cells for <5 times were also included. The supernatant of all virus cultures were harvested six days post infection, clarified by centrifugation at 450× g for 10 min at 4°C, filtered and concentrated by centrifugation at 30,000× g for 3 hrs at 4°C. Virus pellets were re-suspended in DMEM medium with 2% FBS (Invitrogen) and stored at −80°C until use. Infectious titer was determined by plaque assay as described previously [38].
Virus infection in HuH7 cells
HuH7 cells were seeded at 3×106 per flask in 25 mm flasks and incubated at 37°C for 24 hrs before infecting at a MOI of 20 with each of the DENV strains for 1 hr at 37°C/5% CO2, with gentle rocking every 15 min. The cells were then washed thoroughly and replaced with DMEM medium supplemented with 2% FBS and incubated for 20 hrs.
Immunofluorescence assay
Immunofluorescence assay was conducted according to a previously described method [39]. Briefly, the cells from the virus culture were washed once and re-suspended with PBS, and spotted onto a Teflon coated glass slide, air dried and then immersed in 80% acetone for 10 min. The slide was rinsed with PBS and air-dried. 2 µl antibody against prM protein (2H2 monoclonal antibody) was added onto each well, incubated at 37°C for 45 min in a humidified chamber, and washed twice with PBS before drying. FITC-conjugated goat anti-mouse IgG were diluted 1∶30 with 0.1% Evan's Blue and 2 µl was added onto each well. Slides were then incubated at 37°C for 45 min in the humidified chamber and then washed twice with PBS. Slides were dried and mounted with buffered glycerol before imaging under a fluorescent microscope.
Generation of whole-transcriptome cDNA library
Polyadenylated mRNA was isolated from HuH7 cells by three rounds of selection with the Dynabeads mRNA Direct Kit (Invitrogen) and assessed by electrophoresis on the Bioanalyzer 2100 (Agilent) for quality evaluation. For the RNAseq sample preparation, the NEBNext mRNA Sample Prep Master Mix Set 1 was used according to the manufacturer's protocol (NEB). Briefly, 0.5 ug mRNA was used for fragmentation and then subjected to cDNA synthesis using SuperScript III Reverse Transcriptase (Invitrogen) and random primers. The cDNA was further converted into double stranded cDNA and after an end repair process (Klenow fragment, T4 polynucleotide kinase and T4 polymerase), was ligated to Illumina paired end (PE) adaptors. Size selection was performed using a 2% agarose gel, generating cDNA libraries ranging in size from 275–325 bp. Finally, the libraries were enriched using 15 cycles of PCR and purified by the QIAquick PCR purification kit (Qiagen).
Sequence analysis
Libraries were sequenced on an Illumina GAIIx machine at the National Cancer Center, Singapore (Control-NCC, 16007-NCC, PDK13-NCC) or an Illumina HiSeq 2000 machine at the Duke-NUS Genome Biology Facility, Singapore (Control-1, Control-2, 16007-1, 16007-2, PDK13-1, PDK13-2). Resulting reads were mapped to the hg19 build of the human genome using Tophat v1.3.0 (http://tophat.cbcb.umd.edu/index.html) with the coverage-search, microexon-search and butterfly-search options. Differential isoform expression analysis was done using Cufflinks v1.3.0 (http://cufflinks.cbcb.umd.edu/) with the multi-read-correct (Cufflinks), -r and -s (Cuffcompare; using the same annotation gtf and hg19 fasta files respectively as in Tophat) and the mask-file (rRNA), frag-bias-correct (same hg19 fasta file used for Tophat) and multi-read-correct options. Differential splicing analysis was done using MISO v1.0 (http://genes.mit.edu/burgelab/miso/docs/) using the default options. Analysis of RNA sequencing quality was performed with RNAseqC v1.7 (http://www.broadinstitute.org/cancer/cga/rna-seqc) using the default options. Creation of proportional Venn diagrams was done with freeware available at www.venndiagram.tk. Hierarchical clustering of differentially regulated isoforms was done with Partek v6.6 (http://www.partek.com/). Pathway analysis of differentially regulated isoforms was done using Ingenuity Pathway Analysis v9.0 (http://www.ingenuity.com/).
Validation of exon skipping by real-time PCR
Total RNA derived from mock-infected and DENV-infected cells was used to synthesize cDNA using SuperScript III First Strand Synthesis System (Invitrogen) with random hexamers according to manufacturer's instructions. Quantitative real-time PCR was performed using LightCycler 480 Real-Time PCR System (Roche Diagnostics GmbH, Germany) and LightCycler 480 SYBR Green I Master (Roche Diagnostics GmbH, Germany). The reaction was carried out to simultaneously amplify exon-skipped and exon-included isoforms using specific primers complementary to the exons flanking each target exon (Table S13). Percent exon exclusion levels were calculated as the percentage of the isoform excluding an alternative exon divided by the total abundance of the isoforms including and excluding the alternative exon.
Statistical analysis
The statistical analysis employed to analyze biological triplicates for each condition are as previously described using the software Cufflinks and Mixture-of-Isoforms (MISO) [40], [41]. Criteria used to define significance are according to the standard options in the Cufflinks and MISO programs. More detail is provided below.
Results
Experimental design and RNAseq results
To investigate the effect of DENV1-16007 (wild-type strain) and DENV1-PDK13 (attenuated strain) infection on the transcriptome of the human host, we infected human hepatoma cells (HuH7) with each strain for 20 hours at a multiplicity of infection (MOI) of 20 (Figure 1A). The MOI of 20 was chosen so that the subsequent RNAseq profiling would best reflect the infection-induced alterations to the host transcriptome and not be either masked by or derived from a large number of uninfected cells. We also measured viral RNA over the first 30 hours of infection to address the possibility that changes observed might be the result of a delayed replication cycle by one of the viruses. Although the absolute kinetics of the two viruses differ, the 20 hour time point was chosen as it represents the stage in the primary round of replication at which the genome copy numbers are most similar between the two viruses (Figure 1B). Indirect immunofluorescence staining for the pre-membrane protein (prM) production with 2H2 monoclonal antibody also showed similar infection levels for both viruses at this time point (Figure 1C). Infections with wild-type and attenuated strains were performed in three independent biological replicate experiments. Mock-infected HuH7 cells treated in the same way as the infected samples were also done in biological triplicate and served as the control for our experiment. At twenty hours post-infection, mRNA was extracted from all samples independently, and poly-A enriched cDNA libraries were constructed for 75-base, pair-end sequencing on an Illumina GAIIx (one sample for each condition) or Illumina HiSeq2000 machine (two samples for each condition). RNA sequencing was performed independently for each of these replicate experiments.
Figure 1. Experimental design and analysis of infection.

A. Timeline of experimental procedure and data analysis steps. B. Quantification of viral copy number normalized to the cellular B2M gene expression at 0, 6, 12, 20 and 30 hours post infection. Asterisks indicate a p-value of less than or equal to 0.05. C. Immunofluorescence analysis of Huh7 cells either mock-infected (top panel), infected with DENV1-16007 (middle panel) or DENV1- PDK13 (bottom panel) and probed with antibody against prM protein (2H2) followed by FITC-conjugated goat anti-mouse IgG at 20 h post-infection. The cell nuclei were counter-stained with Evan's Blue.
Mapping of reads (Bowtie, Tophat) and analysis of differential transcriptome response (Cufflinks) to infection was performed using the Tuxedo Suite of software [40], [42]. Cufflinks utilizes sequence fragments mapped to the reference genome to estimate the abundance of each isoform arising from the gene. It then tests for differential expression between experimental conditions. Differential expression at the gene level is calculated and is defined as the sum of differential expression of all isoforms at a particular locus. Cufflinks also assesses differential splicing by comparing the relative abundance of isoforms using the same transcriptional start site [40]. As this definition of splicing encompasses all different types of splicing events (see below) [43], systematic analysis of these events for downstream validation work is exceedingly difficult. In order to attain more detail about the types of differential splicing in our samples, we used the MISO software following mapping [41]. MISO utilizes a fixed library of previously characterized splice events to predict alternative splicing and reports the number of occurrences for each type of splicing event: skipped exon (SE), mutually exclusive exons (MXE), alternative 3′ splice site (A3SS), alternative 5′ splice site (A5SS), alternative first exon (AFE), retained intron (RI), tandem untranslated regions (Tandem UTR) [43]. MISO also utilizes a different algorithm than Cufflinks to predict differential splicing between samples which, when compared with the results from Cufflinks, provides an additional level of stringency in selection of candidates for downstream analysis.
To assess the quality of our libraries and sequencing performance and to ensure that any differences observed between our samples was due to the biology and not bias in sequencing, we used the open source program RNAseqC to examine our data [44]. Results indicate that despite using two different Illumina machines to generate the sequences, the individual samples are highly comparable to each other across all the metrics interrogated (Dataset S1).
Wild-type and attenuated DENV-1 strains induce differential regulation of the host transcriptome
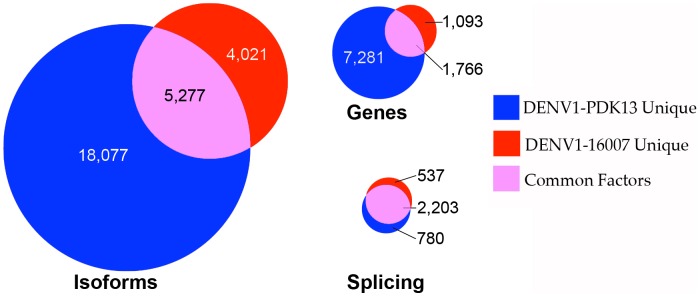
Over 18,000 changes to the host transcriptome were observed in response to infection by the wild-type strain and >41,000 were observed in response to infection with the attenuated strain (Table 1). Differential isoform regulation is the largest category of response due to infection by both strains. Interestingly, there are over two-fold more differentially regulated isoforms following infection with the attenuated than with the wild-type strain. Similarly, infection with the attenuated strain also resulted in three-fold more differentially regulated genes than infection with the wild-type (Figure 2). In order to determine whether this large differential response to infection was specific to these two strains or whether the ‘quieter’ response to the parental DENV1-16007 strain was typical of wild-type DENV1s, we repeated our experiment in HuH7 cells with two low passaged clinical isolates (EDEN3300 and SL107) and compared them to our uninfected control. RNAseq analysis for these isolates indicates even fewer transcriptomic changes (Table 1), suggesting that attenuated virus triggers more host cell response than wild-type viruses. This observation is consistent with what has been reported for yellow fever virus and its attenuated derivative, YF17D [45].
Table 1. Differential regulation of transcripts in response to DENV1 infection.
| Cufflinks | Uninfecteda vs PDK13a | Uninfecteda vs 16007a | Uninfecteda vs EDEN3300b | Uninfecteda vs SL107b |
| Genes exp | 9047 (4610 up/4437 down) | 2859 (1818 up/1041 down) | 5162 (109 up/5053 down) | 5033 (251 up/4782 down) |
| Isoforms exp | 23354 (12386 up/10968 down) | 9298 (5332 up/3966 down) | 6416 (775 up/5641 down) | 7023 (965 up/6058 down) |
| TSS group exp | 3189 | 1194 | 1429 | 1392 |
| CDS exp | 148 | 48 | 190 | 200 |
| CDS | 153 | 132 | 9 | 7 |
| Promoters | 2249 | 2112 | 148 | 184 |
| Splicing | 2983 | 2740 | 159 | 208 |
| Total | 41123 | 18383 | 13513 | 14047 |
Number of events in each category (Rows) predicted by Cuffdiff [40] (Top) and MISO [41] (Bottom) for Uninfected cells vs. DENV1-16007 infected cells (First column), Uninfected cells vs. DENV1-PDK13 infected cells (Second column). The number of up- and down-regulated events is also described for the gene and isoform categories. Superscript “a” indicates biological triplicates while superscript “b” indicates single measurement of the sample. Abbreviations used: differentially expressed genes (Genes exp), differentially expressed isoforms (Isoforms exp), differentially expressed transcriptional start site group (TSS group exp), differentially expressed coding sequence (CDS exp), differential coding output (CDS), differential promoter use (Promoters), differential splicing (Splicing), skipped exon (SE), mutually exclusive exons (MXE), alternative 3′ splice site (A3SS), alternative 5′ splice site (A5SS), alternative first exon (AFE), retained intron (RI) and tandem untranslated region (Tandem UTR).
Figure 2. Comparison of differential regulation of genes, isoforms and splicing in DENV1-16007 and DENV1-PDK13.
Qualitative analysis of the RNAseq data also provides insights into the pathways that are essential to both strains or unique to one strain. Ingenuity Pathway Analysis (IPA) of isoforms indicates that the commonly regulated isoforms are enriched in pathways associated with viral infection and modulation of protein translation. Examples include EIF2 signaling, prolactin signaling, acute phase response signaling, regulation of EIF4 and p7056K and glucocorticoid receptor signaling. Differential expression of pathways associated with cellular growth and proliferation such as mTOR signaling and aryl hydrocarbon receptor signaling are also enriched following infection with both strains (Figure 3A). Conversely, the pathways that are differentially regulated between the wild-type and attenuated strains include the innate immune response and cell cycle control. The top differentially regulated host pathways associated only with wild-type strain infection are involved with immunomodulation and cell cycle arrest, such as PPAR/RXR activation pathway, G2/M DNA damage checkpoint, ATM signaling and the PDGF signaling pathway (Figure 3C). Conversely, infection with the attenuated strain triggered pathways associated with inflammation, induction of apoptosis and stress such as the TNFR1 pathway, TWEAK signaling, NRF2-mediated oxidative stress response, IGF -1 signaling and ERK5 signaling (Figure 3E). IPA also identified molecular and cellular functions associated with both or each of the strains (Figures 3B, 3D and 3F). Taken collectively, the molecular and cellular functions in response to wild-type virus infection are associated with cell signaling and metabolism while those to attenuated virus are associated with transcriptional activation, cell cycle modification and post-translational modification.
Figure 3. Ingenuity pathway analysis.

Pathway (A, C, E) and molecular and cellular function (B, D, F) analysis of common DENV1 response (A, B), unique DENV1-16007 response (C, D) and unique DENV1-PDK13 response (E, F). The numbers within the pie-slices in panels B, D and F are the number of isoforms belonging to the indicated molecular and cellular function.
Differential splicing in the host transcriptome is largely conserved between wild-type and attenuated DENV1 strains
Next, we interrogated the data for alternative splicing within the isoforms sharing the same transcriptional start site. Interestingly, 80% and 74% of all differential splicing following infection with the wild-type strain and attenuated strain, respectively, are common to both viruses (Figure 2). This degree of commonality in the splicing response to infection by each strain is significantly different than what was observed for differential isoform and gene regulation in response to infection. This novel finding suggests that mechanisms responsible for the specific regulation of host splicing may be critical for DENV1 propagation and thus remained relatively unchanged during the attenuation process.
The candidate list from our Cufflinks analysis of alternatively spliced transcripts for both strains of virus was then cross-referenced against the list of skipped exon (SE) events generated by MISO analysis. This cross comparison resulted in 79 total SE events. To assess the accuracy of our alternative splicing predictions, we performed qPCR on each of the predicted SE events. Of the 79 events tested by qPCR, 32 (40%) were differentially expressed in the wild-type strain, the attenuated strain or both in comparison to an uninfected control (Figure 4). The splicing patterns were similar for both viruses across all time points measured, indicating that our observations are not artifacts of temporal sampling bias (Figure 4). Furthermore, the genes involved in these splicing events belong to many of the same pathways shown in Figure 3 suggesting that the virus is, at least in part, exerting its influence on these pathways through alternative splicing (Table S1). If this were true, the SE events should share mechanisms of regulation by utilizing common RNA motifs within and surrounding the identified SE's. Indeed, using the software RegRNA (http://regrna.mbc.nctu.edu.tw) [46] and setting an arbitrary boundary of 250 nucleotides upstream and downstream of the 3′ and 5′ splice-site of the SEs, respectively, we identified 74 predicted RNA regulatory motifs and elements which could be bound by 17 RNA binding proteins. By using GeneCards (http://www.genecards.org) to convert these putative motif binding proteins into HUGO nomenclature, we observed that nearly two thirds (11 of 17) of genes encoding these motif-binding proteins are themselves differentially expressed following infection by one or both of the DENV1 strains (Table S2). These results suggest that DENV1 may be regulating alternative splicing of host mRNA by a hitherto unknown mechanism.
Figure 4. Validation of single exon skipping events predicted by both Cuffdiff and MISO.

Values graphed are the log2 transformed fold change differences between inclusion/skipping of the alternatively spliced exon in DENV1-16007 infected cells (red bars) or DENV1-PDK13 infected cells (blue bars) compared to uninfected control cells for 0, 12, 20 and 30 hours post infection. Asterisks indicate a p-value of less than or equal to 0.05.
Cross-platform, integrative analysis of results
Many differentially processed transcripts found in our study have also been identified in genome-wide RNAi screens for flavivirus host factors, differentially processed in microarray studies of DENV-infected cells and/or shown to interact with DENV gene products [10]–[29], [47]–[51] (Jamison and Garcia-Blanco, unpublished data). To gain additional functional insights into the differentially regulated transcripts identified here, we compiled a cross-platform integrative analysis of our data with other available genomic data on host factors in DENV and other flaviviral infection (Tables S3, S4, S5, S6, S7, S8, S9, S10, S11, S12, S13). Briefly, we examined 11 canonical pathways that were enriched in our study and/or had been previously implicated in the DENV life cycle: apoptosis, autophagy, clathrin-mediated endocytosis, interferon signaling, lipid metabolism, oxidative phosphorylation, regulation of stress granules and P-bodies, splicing-related RNA post-transcriptional modification, ubiquitination, endoplasmic reticulum stress, virus recognition and interferon induction. The proportion of host factors for DENV or other flaviviruses identified through functional genomic studies that were differentially regulated following infection with the wild-type strain or the attenuated strain is indicated in Table 2. Differential isoform regulation of flaviviral host factors ranged from 15.5% to 60.6% and is uniformly higher for the attenuated strain than the pathogenic wild-type strain. Alternative splicing ranged from 7.3% to 24.2% and the rates of these events in the wild-type strain and its attenuated derivative were comparable (Table 2). The intersection of specific host factors in these eleven canonical pathways that are either required for flaviviral propagation or involved in direct interaction with DENV proteins, which are differentially regulated are graphically depicted in Figure 5.
Table 2. Cross-platform analysis of DENV host factors.
| Screen | Reference | Cell Line Used | Identified | Isoform | Isoform | Splicing | Splicing |
| DENV (Drosophila) | Sessions et al 2009 | S2, HuH7 | 56 | 14 (25%) | 24 (42.9%) | 7 (12.5%) | 7 (12.5%) |
| DENV (Human) | Jamison et al Unpublished | HuH7 | 943 | 191 (20.3%) | 360 (38.2%) | 90 (9.5%) | 89 (9.4%) |
| HCV-Replicon | Tai et al 2009 | HuH7/Rep-Feo | 97 | 27 (27.8%) | 43 (44.3%) | 12 (12.4%) | 12 (12.4%) |
| HCV-Virus | Li et al 2009 | HuH7.5.1 | 263 | 68 (25.9%) | 115 (43.7%) | 30 (11.4%) | 29 (11%) |
| WNV | Krishnan et al 2008 | HeLa | 306 | 54 (17.6%) | 105 (34.3%) | 29 (9.5%) | 34 (11.1%) |
| WNV | Krishnan et al 2008 (Unpublished Hits) | HeLa | 1259 | 195 (15.5%) | 369 (29.3%) | 92 (7.3%) | 101 (8%) |
| YFV | Le Sommer et al 2012 | HuH7 | 647 | 116 (17.9%) | 217 (33.5%) | 49 (7.6%) | 54 (8.3%) |
| DENV UTR-BP's | Ward et al 2011 | HuH7 | 62 | 26 (41.9%) | 34 (54.8%) | 14 (22.6%) | 15 (24.2%) |
| DENV-Y2H | Le Breton et al 2011 | S. cerevisae, HEK293T | 104 | 45 (43.3%) | 63 (60.6%) | 16 (15.4%) | 17 (16.3%) |
The number of host factors identified in each of the RNAi screens listed compared to the differentially regulated isoforms and differentially spliced genes for both the wild-type DENV1-16007 and vaccine strain DENV1-PDK13. Percentages of overlap are indicated in parenthesis. The unpublished DENV screen by Jamison SF, Garcia-Blanco, M.A. utilize the same siRNA library, cell line, general methodology and statistical analysis described in Le Sommer et al 2012. The unpublished hits from the WNV screen are the full, unannotated results from the published Krishnan et al 2008 study.
Figure 5. Cross-platform analysis of human-Flaviviridae interactions.

Members of eleven canonical pathways that have been previously implicated to be important for DENV propagation were interrogated for their appearance in an RNAi screen for Flaviviridae host factors, direct interaction with DENV gene products, differential regulation in a microarray study, differential isoform regulation following infection with DENV1-16007 and/or differential isoform regulation following infection with DENV1-PDK13. Host factors identified in published and/or unpublished Flaviviridae RNAi screens [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data) were combined and are indicated by a black bar. Host proteins found to interact DENV gene products [47], [50] were combined and are indicated by a grey bar. Host genes found to be differentially regulated in microarray studies [12]–[29] are indicated by the striped bar. A red bar indicates host isoforms found to be differentially regulated during infection with DENV1-16007. A blue bar indicates host isoforms found to be differentially regulated during infection with DENV1-PDK13.
Discussion
Investigation of host gene expression to date has relied primarily on microarray technology [12]–[29]. This technology is insensitive to the regulation of genes through alternative RNA processing. Thus, detection of differential expression down to the level of alternative isoforms has not been examined and the subtleties of TSS and post-transcriptional modifications that can dramatically alter the function of the derived proteins have been largely ignored. These processes could be a mechanism by which DENV attains specific isoforms of required host factors while suppressing those that act to restrict its replication [27], [52], [53]. Indeed, the need to understand the host response beyond simple gene expression is underscored by the observation that DENV can modify the splicing pattern of an endogenous gene, XBP1, to its advantage [53]. Our findings indicate that there is an extensive amount of previously uncharacterized gene isoforms and alternative processing of host transcripts over a wide range of pathways and host functions in response to DENV infection. Interestingly, the DENV1-16007 and DENV1-PDK13 viruses only differ from each other by 14 nucleotide and 8 amino acids [54], yet the host transcriptional response to these viruses is pronounced. This suggests that infection with different strains of DENV can result in significantly different disease phenotypes despite few nucleotide differences.
We have attempted, in this study, to provide a comprehensive guide to the transcriptomic changes with DENV infection. By analyzing our RNAseq data using two different programs, Cufflinks and MISO [40], [41], maximal information on RNA transcript regulation, could be gleaned. Specific splice events could hence be identified for subsequent mechanistic studies that clarify their role in the host response. In particular, the integrative analysis of this study with existing functional genomics data reveals previously undocumented expression and post-transcriptional regulation of required host factors that should serve as a road map for future mechanistic investigations. A caveat, however, is that our work only profiled the transcriptome at the end of a single round of DENV replication. As hinted by Figure 4, both quantitative and qualitative differences may exist in the transcriptome at different stages of the virus life cycle. Furthermore, the possibility exists that bystander uninfected cells may exert some influence on the observed transcriptional changes although we have attempted to minimize this by using a high MOI in our experiments. Future studies may need to take these possibilities into account.
The large difference in the number of alternative splicing events identified by Cufflinks and MISO also underscores the fledgling nature of RNAseq. While the former identifies all possible splicing events from the data de novo, the latter relies on a pre-defined library to map alternatively spliced transcripts. Additional studies are needed to clarify whether Cufflinks over-estimated the number of splice variants, or there are authentic variants absent from the library used by MISO. Regardless, for transcriptome analysis of host response to infection, RNAseq is superior to microarray in terms of the breadth of information derived.
Our results also indicate that the human transcriptome response to an infection by wild-type DENV or its attenuated derivative differs significantly (Table 1, Figure 2). These differences suggest that alternative processing events may be part of a previously uncharacterized innate immune response to DENV1 infection that is in large part evaded by wild-type strains. This second hypothesis is supported by the greater than two-fold increase in the number of differentially regulated transcripts when infected with the attenuated strain of DENV1 as compared to the parental wild type strain of DENV1, many of which belong to pathways associated with inflammation, induction of apoptosis and stress response (Table 1, Figure 3). This inability to escape the innate immune response achieved by the wild-type virus may explain the lack of reactogenicity. It may also explain why antibody titer engendered by vaccination is not as high as those observed following natural infection unless supplemented with an adjuvant that stimulates this innate immune response. This observation also suggests a mechanism of pathogenicity where DENV regulates host transcriptome changes by interacting with a group of RNA binding proteins to control multiple splicing events.
The development of a live attenuated tetravalent vaccine for DENV1-4 has bedeviled researchers for the past 60 years. The less than optimal efficacy of the leading dengue vaccine candidate makes an improved understanding of the molecular basis of a good vaccine all the more critical [55]. The differential host transcriptome response to infection with DENV1-16007 and DENV1-PDK13 provides an insight into the characteristics of an attenuated virus which, is likely a complex phenotype [45]. A molecular understanding of the basis of attenuation could lead to a quantitative approach to balancing reactogenicity and immunogenicity, which presently remains a hit-or-miss finding made only after lengthy clinical trials.
In conclusion, we provide here a detailed view of the host cell transcriptome response to infection with wild-type DENV-1 and its attenuated derivative that could be useful for future studies on the genetic determinants of viral virulence and attenuation.
Supporting Information
RNASeQC analysis. To assess the quality of our sequencing reactions and mapping, we used the open-source software RNASeQC available at: http://www.broadinstitute.org/cancer/cga/rna-seqc. Information about Total Reads, Mapped Reads, Mate Pairs, Transcript-associated Reads and Strand Specificity are output. Details about each information type are listed below the corresponding table. Additionally, a Spearman Correlation Matrix and Pearson Correlation Matrix were produced for the samples. Tables listing the Coverage Metrics are produced for the Bottom 1000 Expressed Transcripts, the Middle 1000 Expressed Transcripts and the Top 1000 Expressed Transcripts. Graphs depicting the Mean Coverage over the Percentage of Transcript Length (5′ to 3′) and graphs depicting Mean Coverage from the 3′ End for Low Expressed Transcripts, Medium Expressed Transcripts and High Expressed Transcripts are also provided.
(PDF)
Ingenuity pathway analysis of the 33 skipped exon events found to be differentially regulated by qPCR. −log(p-value) reflects the statistical significance of the indicated observation while the ratio refers to the number of genes identified in the indicated pathway divided by the total number of genes belonging to that pathway.
(XLS)
Motif analysis of the RNA surrounding skipped exons. The 500 base areas surrounding the skipped exon described in Figure 4, as well as the exons themselves were interrogated for the presence of RNA regulatory motifs and elements. The predicted motif binding proteins were translated into HUGO nomenclature and checked for differential regulation following infection with DENV1-16007, DENV1-PDK13 or both.
(XLS)
Cross-platform analysis of the apoptosis pathway. Members of the apoptosis pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the autophagy pathway. Members of the autophagy pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the clathrin-mediated endocytosis pathway. Members of the clathrin-mediated endocytosis pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the interferon signaling pathway. Members of the interferon signaling pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the lipid metabolism pathway. Members of the lipid metabolism pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the oxidative phosphorylation pathway. Members of the oxidative phosphorylation pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the regulation of stress granules and p-bodies pathway. Members of the regulation of stress granules and p-bodies pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the splicing related RNA post-transcriptional modification pathway. Members of the splicing related RNA post-transcriptional modification pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the ubiquitination pathway. Members of the ubiquitination pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the endoplasmic reticulum stress pathway. Members of the endoplasmic reticulum stress pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the virus recognition and interferon induction pathway. Members of the virus recognition and interferon induction pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Primers. The primer sequences used to validate the SE exon events are detailed here.
(XLS)
Acknowledgments
The authors would like to thank Drs. D.J. Gubler, M.A. Garcia-Blanco, M. Krishnan, P.F. de Sessions and A. Ward for their critical review of this manuscript. We would also like to thank M. Krishnan, M.A. Garcia-Blanco and S. Jamison for the contribution of their unpublished work, T. Chawla for the DENV1-16007 and DENV1-PDK13 sequencing and the Duke-NUS Genome Biology Facility for next-generation sequencing services.
Funding Statement
This work was supported by the Singapore National Research Foundation under its Clinician-Scientist Award administered by the National Medical Research Council. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Kyle JL, Harris E (2008) Global spread and persistence of dengue. Annu Rev Microbiol 62: 71–92. [DOI] [PubMed] [Google Scholar]
- 2. Wilder-Smith A, Ooi EE, Vasudevan SG, Gubler DJ (2010) Update on dengue: epidemiology, virus evolution, antiviral drugs, and vaccine development. Curr Infect Dis Rep 12: 157–164. [DOI] [PubMed] [Google Scholar]
- 3. Beatty ME, Beutels P, Meltzer MI, Shepard DS, Hombach J, et al. (2011) Health economics of dengue: a systematic literature review and expert panel's assessment. Am J Trop Med Hyg 84: 473–488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Carrasco LR, Lee LK, Lee VJ, Ooi EE, Shepard DS, et al. (2011) Economic impact of dengue illness and the cost-effectiveness of future vaccination programs in Singapore. PLoS Negl Trop Dis 5: e1426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Gubler DJ (2004) The changing epidemiology of yellow fever and dengue, 1900 to 2003: full circle? Comp Immunol Microbiol Infect Dis 27: 319–330. [DOI] [PubMed] [Google Scholar]
- 6. Normile D (2007) Tropical diseases. Hunt for dengue vaccine heats up as the disease burden grows. Science 317: 1494–1495. [DOI] [PubMed] [Google Scholar]
- 7. Shepard DS, Suaya JA, Halstead SB, Nathan MB, Gubler DJ, et al. (2004) Cost-effectiveness of a pediatric dengue vaccine. Vaccine 22: 1275–1280. [DOI] [PubMed] [Google Scholar]
- 8. Suaya JA, Shepard DS, Siqueira JB, Martelli CT, Lum LC, et al. (2009) Cost of dengue cases in eight countries in the Americas and Asia: a prospective study. Am J Trop Med Hyg 80: 846–855. [PubMed] [Google Scholar]
- 9. Fernandez-Garcia MD, Mazzon M, Jacobs M, Amara A (2009) Pathogenesis of flavivirus infections: using and abusing the host cell. Cell Host Microbe 5: 318–328. [DOI] [PubMed] [Google Scholar]
- 10. Krishnan MN, Ng A, Sukumaran B, Gilfoy FD, Uchil PD, et al. (2008) RNA interference screen for human genes associated with West Nile virus infection. Nature 455: 242–245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Sessions OM, Barrows NJ, Souza-Neto JA, Robinson TJ, Hershey CL, et al. (2009) Discovery of insect and human dengue virus host factors. Nature 458: 1047–1050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Balas C, Kennel A, Deauvieau F, Sodoyer R, Arnaud-Barbe N, et al. (2011) Different innate signatures induced in human monocyte-derived dendritic cells by wild-type dengue 3 virus, attenuated but reactogenic dengue 3 vaccine virus, or attenuated nonreactogenic dengue 1–4 vaccine virus strains. J Infect Dis 203: 103–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Becerra A, Warke RV, Martin K, Xhaja K, de Bosch N, et al. (2009) Gene expression profiling of dengue infected human primary cells identifies secreted mediators in vivo. Journal of medical virology 81: 1403–1411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Conceicao TM, El-Bacha T, Villas-Boas CS, Coello G, Ramirez J, et al. (2010) Gene expression analysis during dengue virus infection in HepG2 cells reveals virus control of innate immune response. The Journal of infection 60: 65–75. [DOI] [PubMed] [Google Scholar]
- 15. Fink J, Gu F, Ling L, Tolfvenstam T, Olfat F, et al. (2007) Host gene expression profiling of dengue virus infection in cell lines and patients. PLoS Negl Trop Dis 1: e86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Hoang LT, Lynn DJ, Henn M, Birren BW, Lennon NJ, et al. (2010) The early whole-blood transcriptional signature of dengue virus and features associated with progression to dengue shock syndrome in Vietnamese children and young adults. J Virol 84: 12982–12994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Liew KJ, Chow VT (2006) Microarray and real-time RT-PCR analyses of a novel set of differentially expressed human genes in ECV304 endothelial-like cells infected with dengue virus type 2. Journal of virological methods 131: 47–57. [DOI] [PubMed] [Google Scholar]
- 18. Loke P, Hammond SN, Leung JM, Kim CC, Batra S, et al. (2010) Gene expression patterns of dengue virus-infected children from nicaragua reveal a distinct signature of increased metabolism. PLoS Negl Trop Dis 4: e710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Long HT, Hibberd ML, Hien TT, Dung NM, Van Ngoc T, et al. (2009) Patterns of gene transcript abundance in the blood of children with severe or uncomplicated dengue highlight differences in disease evolution and host response to dengue virus infection. J Infect Dis 199: 537–546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Moreno-Altamirano MM, Romano M, Legorreta-Herrera M, Sanchez-Garcia FJ, Colston MJ (2004) Gene expression in human macrophages infected with dengue virus serotype-2. Scandinavian journal of immunology 60: 631–638. [DOI] [PubMed] [Google Scholar]
- 21. Nascimento EJ, Braga-Neto U, Calzavara-Silva CE, Gomes AL, Abath FG, et al. (2009) Gene expression profiling during early acute febrile stage of dengue infection can predict the disease outcome. PloS one 4: e7892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Nasirudeen AM, Liu DX (2009) Gene expression profiling by microarray analysis reveals an important role for caspase-1 in dengue virus-induced p53-mediated apoptosis. Journal of medical virology 81: 1069–1081. [DOI] [PubMed] [Google Scholar]
- 23. Silveira GF, Meyer F, Delfraro A, Mosimann AL, Coluchi N, et al. (2011) Dengue virus type 3 isolated from a fatal case with visceral complications induces enhanced proinflammatory responses and apoptosis of human dendritic cells. J Virol 85: 5374–5383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Simmons CP, Popper S, Dolocek C, Chau TN, Griffiths M, et al. (2007) Patterns of host genome-wide gene transcript abundance in the peripheral blood of patients with acute dengue hemorrhagic fever. J Infect Dis 195: 1097–1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Tolfvenstam T, Lindblom A, Schreiber MJ, Ling L, Chow A, et al. (2011) Characterization of early host responses in adults with dengue disease. BMC infectious diseases 11: 209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Ubol S, Masrinoul P, Chaijaruwanich J, Kalayanarooj S, Charoensirisuthikul T, et al. (2008) Differences in global gene expression in peripheral blood mononuclear cells indicate a significant role of the innate responses in progression of dengue fever but not dengue hemorrhagic fever. J Infect Dis 197: 1459–1467. [DOI] [PubMed] [Google Scholar]
- 27. Umareddy I, Pluquet O, Wang QY, Vasudevan SG, Chevet E, et al. (2007) Dengue virus serotype infection specifies the activation of the unfolded protein response. Virol J 4: 91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Villas-Boas CS, Conceicao TM, Ramirez J, Santoro AB, Da Poian AT, et al. (2009) Dengue virus-induced regulation of the host cell translational machinery. Brazilian journal of medical and biological research = Revista brasileira de pesquisas medicas e biologicas/Sociedade Brasileira de Biofisica [et al] 42: 1020–1026. [DOI] [PubMed] [Google Scholar]
- 29. Warke RV, Xhaja K, Martin KJ, Fournier MF, Shaw SK, et al. (2003) Dengue virus induces novel changes in gene expression of human umbilical vein endothelial cells. J Virol 77: 11822–11832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Blencowe BJ (2006) Alternative splicing: new insights from global analyses. Cell 126: 37–47. [DOI] [PubMed] [Google Scholar]
- 31. Matlin AJ, Clark F, Smith CW (2005) Understanding alternative splicing: towards a cellular code. Nat Rev Mol Cell Biol 6: 386–398. [DOI] [PubMed] [Google Scholar]
- 32. Marioni JC, Mason CE, Mane SM, Stephens M, Gilad Y (2008) RNA-seq: an assessment of technical reproducibility and comparison with gene expression arrays. Genome Res 18: 1509–1517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Wang Z, Gerstein M, Snyder M (2009) RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet 10: 57–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bhamarapravati N, Yoksan S. (1997) Live attenuated tetravalent dengue vaccine. In: Gubler DJ, Kuno G., editor. Dengue and Dengue Hemorrhagic Fever. Wallingford, UK: CAB International. pp. 367–378.
- 35. Halstead SB, Marchette NJ (2003) Biologic properties of dengue viruses following serial passage in primary dog kidney cells: studies at the University of Hawaii. Am J Trop Med Hyg 69: 5–11. [DOI] [PubMed] [Google Scholar]
- 36. Schreiber MJ, Holmes EC, Ong SH, Soh HS, Liu W, et al. (2009) Genomic epidemiology of a dengue virus epidemic in urban Singapore. J Virol 83: 4163–4173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Tissera HA, Ooi EE, Gubler DJ, Tan Y, Logendra B, et al. (2011) New dengue virus type 1 genotype in Colombo, Sri Lanka. Emerg Infect Dis 17: 2053–2055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Malewicz B, Jenkin HM (1979) Development of dengue virus plaques under serum-free overlay medium. J Clin Microbiol 9: 609–614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Gubler DJ, Kuno G, Sather GE, Velez M, Oliver A (1984) Mosquito cell cultures and specific monoclonal antibodies in surveillance for dengue viruses. Am J Trop Med Hyg 33: 158–165. [DOI] [PubMed] [Google Scholar]
- 40. Trapnell C, Williams BA, Pertea G, Mortazavi A, Kwan G, et al. (2010) Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol 28: 511–515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Katz Y, Wang ET, Airoldi EM, Burge CB (2010) Analysis and design of RNA sequencing experiments for identifying isoform regulation. Nat Methods 7: 1009–1015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Trapnell C, Pachter L, Salzberg SL (2009) TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25: 1105–1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Wang ET, Sandberg R, Luo S, Khrebtukova I, Zhang L, et al. (2008) Alternative isoform regulation in human tissue transcriptomes. Nature 456: 470–476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Deluca DS, Levin JZ, Sivachenko A, Fennell T, Nazaire MD, et al. (2012) RNA-SeQC: RNA-seq metrics for quality control and process optimization. Bioinformatics 28: 1530–1532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Pulendran B, Ahmed R (2011) Immunological mechanisms of vaccination. Nat Immunol 12: 509–517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Huang HY, Chien CH, Jen KH, Huang HD (2006) RegRNA: an integrated web server for identifying regulatory RNA motifs and elements. Nucleic Acids Res 34: W429–434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Le Breton M, Meyniel-Schicklin L, Deloire A, Coutard B, Canard B, et al. (2011) Flavivirus NS3 and NS5 proteins interaction network: a high-throughput yeast two-hybrid screen. BMC Microbiol 11: 234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Li Q, Brass AL, Ng A, Hu Z, Xavier RJ, et al. (2009) A genome-wide genetic screen for host factors required for hepatitis C virus propagation. Proc Natl Acad Sci U S A 106: 16410–16415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Tai AW, Benita Y, Peng LF, Kim SS, Sakamoto N, et al. (2009) A functional genomic screen identifies cellular cofactors of hepatitis C virus replication. Cell Host Microbe 5: 298–307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Ward AM, Bidet K, Yinglin A, Ler SG, Hogue K, et al. (2011) Quantitative mass spectrometry of DENV-2 RNA-interacting proteins reveals that the DEAD-box RNA helicase DDX6 binds the DB1 and DB2 3′ UTR structures. RNA Biol 8: 1173–1186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Le Sommer C, Barrows NJ, Bradrick SS, Pearson JL, Garcia-Blanco MA (2012) G protein-coupled receptor kinase 2 promotes flaviviridae entry and replication. PLoS Negl Trop Dis 6: e1820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Pena J, Harris E (2011) Dengue virus modulates the unfolded protein response in a time-dependent manner. J Biol Chem 286: 14226–14236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Yu CY, Hsu YW, Liao CL, Lin YL (2006) Flavivirus infection activates the XBP1 pathway of the unfolded protein response to cope with endoplasmic reticulum stress. J Virol 80: 11868–11880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Huang CY, Butrapet S, Pierro DJ, Chang GJ, Hunt AR, et al. (2000) Chimeric dengue type 2 (vaccine strain PDK-53)/dengue type 1 virus as a potential candidate dengue type 1 virus vaccine. J Virol 74: 3020–3028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Sabchareon A, Wallace D, Sirivichayakul C, Limkittikul K, Chanthavanich P, et al. (2012) Protective efficacy of the recombinant, live-attenuated, CYD tetravalent dengue vaccine in Thai schoolchildren: a randomised, controlled phase 2b trial. Lancet 380: 1559–1567. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
RNASeQC analysis. To assess the quality of our sequencing reactions and mapping, we used the open-source software RNASeQC available at: http://www.broadinstitute.org/cancer/cga/rna-seqc. Information about Total Reads, Mapped Reads, Mate Pairs, Transcript-associated Reads and Strand Specificity are output. Details about each information type are listed below the corresponding table. Additionally, a Spearman Correlation Matrix and Pearson Correlation Matrix were produced for the samples. Tables listing the Coverage Metrics are produced for the Bottom 1000 Expressed Transcripts, the Middle 1000 Expressed Transcripts and the Top 1000 Expressed Transcripts. Graphs depicting the Mean Coverage over the Percentage of Transcript Length (5′ to 3′) and graphs depicting Mean Coverage from the 3′ End for Low Expressed Transcripts, Medium Expressed Transcripts and High Expressed Transcripts are also provided.
(PDF)
Ingenuity pathway analysis of the 33 skipped exon events found to be differentially regulated by qPCR. −log(p-value) reflects the statistical significance of the indicated observation while the ratio refers to the number of genes identified in the indicated pathway divided by the total number of genes belonging to that pathway.
(XLS)
Motif analysis of the RNA surrounding skipped exons. The 500 base areas surrounding the skipped exon described in Figure 4, as well as the exons themselves were interrogated for the presence of RNA regulatory motifs and elements. The predicted motif binding proteins were translated into HUGO nomenclature and checked for differential regulation following infection with DENV1-16007, DENV1-PDK13 or both.
(XLS)
Cross-platform analysis of the apoptosis pathway. Members of the apoptosis pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the autophagy pathway. Members of the autophagy pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the clathrin-mediated endocytosis pathway. Members of the clathrin-mediated endocytosis pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the interferon signaling pathway. Members of the interferon signaling pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the lipid metabolism pathway. Members of the lipid metabolism pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the oxidative phosphorylation pathway. Members of the oxidative phosphorylation pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the regulation of stress granules and p-bodies pathway. Members of the regulation of stress granules and p-bodies pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the splicing related RNA post-transcriptional modification pathway. Members of the splicing related RNA post-transcriptional modification pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the ubiquitination pathway. Members of the ubiquitination pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the endoplasmic reticulum stress pathway. Members of the endoplasmic reticulum stress pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Cross-platform analysis of the virus recognition and interferon induction pathway. Members of the virus recognition and interferon induction pathway were interrogated for their appearance in an RNAi screen for Flaviviridae host factors [10], [11], [48], [49], [51] (Jamison and Garcia-Blanco, unpublished data), direct interaction with DENV gene products [47], [50] and differential regulation in a microarray study [12]–[29]. Differential regulation following infection with DENV1-16007 and/or DENV1-PDK13 was also interrogated for each member of these pathways with the Cuffdiff values from the current experiment recorded in each of the corresponding columns.
(XLS)
Primers. The primer sequences used to validate the SE exon events are detailed here.
(XLS)