

Abstract
Cognitive control – the ability to exert control over thoughts, attention and behavior in order to achieve a goal – is essential to adaptive functioning and its disruption characterizes various neuropsychiatric and neurodegenerative disorders. In recent years, increased attention has been devoted to investigating the effects of training on performance and brain function, but little is known about whether cognitive control can be improved through training. To fill this gap, we designed a brief training targeting various components of cognitive control, including conflict monitoring and interference resolution. Twenty participants performed a 3-day training protocol, preceded and followed by identical pre- and post-training sessions, respectively, which included event-related potential (ERP) recordings. To detect transfer effects, the training and pre-/post-training sessions employed different tasks hypothesized to rely on similar interference resolution mechanisms. We hypothesized that training would selectively improve performance for high-interference (i.e., incongruent) trials and be associated with reduced amplitudes in the N2 component, a waveform known to index interference. Trial-to-trial behavioral adjustments were also analyzed to assess potential mechanisms of training-induced improvements. Relative to pre-training, participants showed reduced reaction time (RT) and N2 amplitude for incongruent, but not congruent, trials, suggesting improved interference resolution. Critically, participants showing the greatest reductions in interference effects during the course of the training displayed the largest pre- to post-training reductions in N2 amplitudes in a separate task, highlighting transfer effects. Overall, results suggest that a brief training can improve cognitive control, specifically the ability to inhibit task-irrelevant information.
Keywords: Cognitive control, Interference, Training, Plasticity, ERP, Anterior cingulate cortex
Introduction
Although human behavior is regularly executed in a relatively automatic fashion, individuals often have to assert control over their attention, thoughts and behaviors in order to flexibly adapt to current goals — a process known as cognitive control. The effective execution of cognitive control is an essential aspect of adaptive functioning across domains, including social and emotional functioning (Bartholow, 2010; Ochsner and Gross, 2005), self-regulation (Posner and Rothbart, 2000), and mental health (Gross and Muñoz, 1995). Thus, it is not surprising that deficits in cognitive control characterize mental disorders, including schizophrenia (Reichenberg and Harvey, 2007) and major depressive disorder (MDD; Holmes and Pizzagalli, 2008). Given its critical role in both normal and abnormal functioning, investigating whether cognitive control can be improved could have a large impact. The current study sought to understand whether interference resolution, a type of cognitive control, could be improved through training in healthy individuals.
One important function of cognitive control involves the inhibition of conflicting information in order to maximize task-relevant goals. Tasks assessing interference resolution introduce trials with conflicting stimulus dimensions (e.g., target and distractor arrows pointing in opposite directions in incongruent trials in the Flanker task), which are compared to trials without such conflict (e.g., all arrows pointing in the same direction in congruent trials). Two theories have attempted to explain the neural mechanisms underlying this process: the “conflict monitoring” hypothesis (Botvinick et al., 2001, 2004; Carter and van Veen, 2007) and the “cascade-of-control model” (Banich, 2009; Silton et al., 2010). Both theories emphasize frontocingulate pathways encompassing the dorsolateral prefrontal cortex (DLPFC) and dorsal anterior cingulate regions (dACC) in cognitive control. Moreover, although the exact sequence of neural mechanisms is disputed, both theories highlight dACC activity as distinguishing between congruent and incongruent trials (Carter and van Veen, 2007; Silton et al., 2010). This is supported by studies that have examined event-related potentials (ERP) hypothesized to index dACC-related cognitive control processes. Specifically, relative to congruent trials, incongruent Flanker trials have been found to elicit a larger (more negative) N2 component — a negative-going waveform measured over frontocentral electrodes that has been localized to the dACC (Kopp et al., 1996; Silton et al., 2010; van Veen and Carter, 2002; Yeung et al., 2004). Generally, this increased N2 amplitude to incongruent trials is thought to reflect activity “monitoring” for an increase in conflicting information. On trials with little or no conflict, a reduced N2 signal would be expected (Carter and van Veen, 2007; Silton et al., 2010).
In addition to interference effects, cognitive control can be evaluated by considering performance in sequential trials. Speciflcally, several studies have demonstrated that the amount of interference is smaller for incongruent trials following incongruent trials (I-I trials) relative to incongruent trials following congruent trials (C-I trials) — a phenomenon known as the Gratton effect (Gratton et al., 1992; Kerns, 2006; Kerns et al., 2004). It has been hypothesized that participants’ control is diminished following congruent trials, leading to increased interference in the ensuing trial. However, following an incongruent trial, DLPFC-related control processes become engaged and reduce interference on the following trial (Botvinick et al., 1999; Carter and van Veen, 2007; Kerns, 2006; Kerns et al., 2004). Consistent with these hypothesized mechanisms, a recent study demonstrated a reduction in N2 amplitude as the number of consecutive incongruent trials increased (Clayson and Larson, 2011). The authors argued that each successive incongruent trial (up to four) led to increased recruitment of cognitive control resources, resulting in progressively less conflict and, hence, decreased N2 amplitude (Clayson and Larson, 2011). Given this putative role of the N2 in monitoring conflict, improvement in overall interference resolution mechanisms might show a corresponding result — less conflict and reduced N2 amplitude for every incongruent trial.
Recently, substantial interest has been devoted to investigating the effect of cognitive training paradigms on task performance and brain function. Most of these training studies have focused on working memory (Dahlin et al., 2008a,b; Holmes et al., 2009a,b; Jaeggi et al., 2008; Klingberg, 2010; Klingberg and McNab, 2009; Li et al., 2008; McNab et al., 2009; Sayala et al., 2006; Westerberg and Klingberg, 2007). Only one previous study has examined training effects in tasks that require interference resolution (Rueda et al., 2005). Children (4- and 6-year olds) were trained in five sessions over 2–3 weeks on several cognitive tasks, including one interference resolution task. Findings failed to show RT or accuracy differences between trained and control groups from pre- to post-training. However, trained 6-year olds were characterized by a significant Incongruency effect for N2 amplitudes over a central electrode site (channel Cz), a pattern similar to ERPs in adults, whereas control group 6-year olds, as well as trained and control group 4-year olds, showed this effect at a frontal channel (Fz) (Rueda et al., 2005). The authors argued that, despite the lack of group differences in the behavioral data, training was similar to the influence of maturation, with trained participants showing more “adult-like” brain activity. Although limited, these findings provide initial evidence that training can influence brain networks involved in interference resolution (Rueda et al., 2005). Whether training interference resolution in adults shows similar results accompanied by behavioral improvements not found in children has, to our knowledge, not been investigated before.
Although training paradigms typically elicit increased task performance, one major concern is the generalization (“transfer”) of improvements to other tasks within the same domain or more complex processes that may rely on the mechanisms ostensibly improved. Several studies have demonstrated improved performance transfer to working memory tasks not directly trained, including intelligence tests (Dahlin et al., 2008a; Holmes et al., 2009a; Jaeggi et al., 2008; Klingberg, 2010; Klingberg et al., 2002, 2005; Li et al., 2008).
Theoretically, it has been suggested that transfer occurs if both the training and transfer tasks share a common mechanism and activate similar brain regions or networks (Dahlin et al., 2008a). This hypothesis has been supported by studies demonstrating that increased performance in working memory tasks activating frontoparietal network transferred to similar tasks engaging the same network (Olesen et al., 2003; Westerberg and Klingberg, 2007) but not to other tasks recruiting different neural pathways (Dahlin et al., 2008a). However, it should be noted that a recent large study with 11,000 participants across 12 different training tasks, including working memory tasks, found no evidence of transfer (Owen et al., 2010). This study has been, however, criticized due to lack of training quality control (i.e. lack of supervision), short average duration of each training session (10 min), and unstandardized scheduling (i.e., intermittent days without training; Klingberg, 2010).
Most studies examining the effect of training on neural networks have shown training-related increases in activation in some regions and simultaneous decreases in others (Erickson et al., 2006; Milham et al., 2003; Olesen et al., 2003; Sayala et al., 2006; Siegle et al., 2007; Westerberg and Klingberg, 2007). For example, a study training multi-tasking showed increased activation in bilateral DLPFC regions but decreases in dACC (Erickson et al., 2006). Similarly, while examining practice effects on a Stroop task, Milham et al. (2003) found a rapid decrease in dACC activation compared with a slower reduction in DLPFC activation over the same period of time. The authors suggested that this dissociation was consistent with a role of the DLPFC in exerting top-down control and the dACC in detecting response conflict (Milham et al., 2003). In general, these findings suggest that training-related regional changes may be explained by a given region’s role within a larger brain network.
Alternatively, several studies examining changes as a function of motor training have shown reduced activations in motor-related regions, a finding interpreted as indicating increased cortical efficiency (Haslinger et al., 2004; Meister et al., 2005; Poldrack, 2005; Reithler et al., 2010; Wu, 2004). For example, one study showed improved behavior associated with reduced activation in the same motor-related regions over one day of practice and after 3 additional days of training (Reithler et al., 2010). Additionally, after the 3-day training, participants showed a further reduction in brain activation, which was maintained when completing a novel or familiar but untrained motor movement. Along similar lines, cortical efficiency has been invoked after observing reduced practice-related activity in visual and attentional learning (Mukai et al., 2007) as well as working memory (Sayala et al., 2006) tasks. Collectively, these findings indicate that reduced activation may reflect increased efficiency within task-relevant regions.
The overarching goal of the current study was to investigate whether cognitive control, specifically interference resolution, could be improved through training. To accomplish this, we tested whether training-related improvement in a cognitive control task (the Simon task) would transfer to improvement in a different task (the Flanker task). Both tasks activate overlapping frontocingulate pathways and require interference resolution (Fan et al., 2003; Nee et al., 2007), thus meeting hypothesized conditions for the transfer of training-induced improvements (Dahlin et al., 2008a). During a baseline (pre-training) session, we collected behavioral and ERP data on several tasks. A training battery was then administered for 3 consecutive days, followed by a post-training session that was identical to the pre-training session. We hypothesized that the training would lead to increased performance (as manifested by more accurate and faster responses) on trials that include conflicting stimuli dimensions (i.e., incongruent trials) and involve interference resolution. Trials that contain congruent stimuli dimensions introduce little conflict, require little or no interference resolution, and, therefore, should show negligible improvement from improved interference resolution abilities. Furthermore, based on prior studies showing reduced dACC activation through practice (Erickson et al., 2006; Milham et al., 2003), as well as studies and a computational model of conflict monitoring showing reduced N2 amplitude with decreased interference (Clayson and Larson, 2011; Yeung and Cohen, 2006), we expected a reduction in N2 amplitude from pre- to post-training.
It should be noted that tasks assessing emotional interference were also trained and evaluated during the pre- and post-training sessions. To give a complete account of the study, we report the results of the emotional tasks but the main focus is on tasks assessing non-emotional interference resolution.
Material and methods
Participants
Participants were recruited through Craigslist postings. Inclusion criteria included: age between 18 and 35 years, right-handedness, normal or corrected vision, and ability to provide informed consent. Participants were excluded if they: were color-blind, reported current or past neurological or psychiatric illness or learning disability, or were currently using psychotropic medication. Males and females of all ethnic origins were eligible to participate. Twenty-three participants (14 females) were enrolled. One participant withdrew due to an unrelated medical emergency. Two female participants’ data were excluded from all analyses because of near-chance accuracy during training. One of these participants repeatedly misunderstood the instructions and another employed a strategy in which she disregarded accuracy on congruent trials. Mean age of the remaining 20 participants (11 females) was 27 years (SD=4.32), with 16.5 years (SD=1.34) mean years of education (total subjects available for the behavioral analyses, n=20). Simon task data (i.e., mean RT and accuracy for congruent and incongruent trials) for the first training session was lost for one participant. Each missing data point was obtained using the expectation-maximization algorithm (Schafer and Olsen, 1998) in SPSS (version 18). Removing this participant’s data from analyses did not affect results. This participant was excluded from analyses involving change over the course of training because the first training session data were interpolated. Two participants were excluded from all ERP analysis because of excessive artifacts (more than 75% of trials lost; ERP analyses, n=18).
Procedure
Each testing day is referred to as a “session.” The study consisted of 5 consecutive sessions. The first and last sessions – the pre-/post-training sessions – were used to evaluate baseline performance and performance after three training days, respectively. The pre-and post-training sessions were procedurally identical, lasted approximately 2 h, and included electroencephalography (EEG) recordings. In addition, they included a “control task” (see below) to evaluate possible general effects on motor speeding and hypothesized to be unaffected by the cognitive training. Participants were compensated $20 for each pre-/post-training session, and with an additional $15 bonus for completing the study.
During the three intervening training sessions, participants returned to the laboratory and performed a set of tasks different from those administered during the pre-/post-training sessions but hypothesized to train the same underlying mechanisms. Training sessions lasted approximately 45 min and did not include EEG measurements. Subjects were compensated $10 each training day, with the opportunity of earning an additional $5 for performance on the Simon task (see below). All participants completing the study achieved this criterion and received $100 in total.
Task order in training sessions was counterbalanced and pseudo-randomized in pre- and post-training sessions.
Equipment
The pre-/post-training sessions began with the placement of a 128-channel Geodesic Sensor Net (Electrical Geodesic Inc., Eugene, Oregon). Participants performed three computer tasks during collection of ERPs (bandwidth, 0.01–100 Hz; sampling rate: 250 Hz sampling rate; the vertex electrode (Cz) served as recording reference). Impedances were kept below 50 kΩ. Tasks were presented with Eprime software (Psychology Software Tools Inc., Pittsburgh, Pennsylvania) on a 19-inch monitor. Experimenters positioned participants 45 cm from the monitor. Participants’ responses were collected via a button box for the pre-/post-training sessions and via a keyboard for the training sessions.
Tasks
Training session
Simon task
Participants were instructed to make responses to probes presented on either the left or right of a fixation cross with either their left or right index finger. Participants had to press a button located on the same side as the probe if presented with a green circle (congruent probe) but press to the opposite side if presented with a red circle, red square, or green square (incongruent probe). Circle and square probes had a 3 mm diameter and were presented at the midpoint between the edge of the screen and a central fixation cross (9.2 cm from the fixation cross), which was present throughout the task. Congruent trials made up 65% of all trials. Probes were presented for 200 ms followed by a 1500 ms ITI. To incentivize subjects, the task included a $5 bonus, available each day of training, for achieving at least 80% accuracy on incongruent trials. Each of the five blocks included 76 trials. Participants received instructions, both verbally and in writing, to respond as accurately and quickly as possible. The Simon task lasted approximately 17 min, and was adapted from Holmes and Pizzagalli (2007).
Emotional Go/NoGo
In the Emotional Go/NoGo, participants responded in a given block to words within one valence category (i.e., positive, negative or neutral) while withholding their response to a different valence category. Two blocks of each of the following target–distractor pairs were presented: negative targets–positive distractors, negative targets–neutral distractors, positive targets–negative distractors, and neutral targets–negative distractors. The stimuli were taken from the Affective Norms for English Words (ANEW; Bradley and Lang, 1999) and Standard Corpus of Present-Day American English (Francis and Kučera, 1979; Francis et al., 1982). Each word was presented in Arial Narrow, 18-point font for 280 ms, followed by a variable ITI (1000–1200 ms). The task consisted of 8 blocks (90 trials/block), and each block included equal numbers of words from two valence categories. Based on the ANEW database, words from each category did not differ in length (number of letters) or frequency (p<0.1 for all pairwise comparisons) but differed as intended in valence ratings (p<0.05 for all pairwise comparisons). In addition, positive and negative words did not differ on arousal ratings (positive vs. negative, p<0.1), whereas the emotional words had higher arousal ratings relative to neutral words (both ps<0.05). The Emotional Go/NoGo task lasted approximately 15 min, and was adapted from Chiu et al. (2008).
Pre-/post-training sessions
Flanker task
Stimuli consisted of five arrows presented in the center of the screen. The central arrow was the target, while the four non -center arrows were distractors. Trials were either congruent (≪≪< , ≫≫>) or incongruent (≪>≪ , ≫<≫). The trial began with the onset of the four distractor flanker arrows. After 100 ms, the central target arrow appeared for an additional 100 ms. Arrows were 1 cm in length and separated by a space of 0.75 cm. Subjects had 1500 ms to indicate, as fast and accurately as possible, the direction of the target arrow by using the index finger of the corresponding hand. Trials were interspersed with variable (1800–2550 ms) intertrial interval (ITI), during which a fixation cross was presented. To familiarize participants with the task, a practice block of 24 trials was presented (16 congruent, 8 incongruent). Following the practice block, participants completed five experimental blocks, each consisting of 70 trials per block (46 congruent, 24 incongruent). The number of previously congruent trials prior to each incongruent trial was fully randomized using optseq2 (http://surfer.nmr.mgh.harvard.edu/optseq/). The Flanker task lasted approximately 17 min, and was adapted from Holmes et al. (2010).
Emotional Face Stroop task
The Emotional Face Stroop task consisted of 148 presentations of black and white happy or fearful faces derived from the NimStim set of facial expressions (Tottenham et al., 2009). Each face was cropped and presented with the word “FEAR” or “HAPPY” written horizontally in red letters across the middle of the nose. The word and facial expression were either congruent or incongruent. The stimuli, which had a height of 10.5 cm and a width of 7.8 cm, were presented for 1000 ms. A central fixation cross was shown during a variable ITI (3000–5000 ms). In order to familiarize participants with the task, a practice block of 10 trials was presented. Following the practice block, participants completed 148 total trials across 4 blocks, in pseudorandom order. There were equal numbers of incongruent and congruent trials. In addition, to avoid potential confounds of priming effects (Mayr et al., 2003), there were also equal numbers of incongruent–congruent, and incongruent–incongruent, congruent–congruent, and congruent–incongruent trial combinations. To further prevent priming effects, identical faces were not presented consecutively. Participants were instructed to respond as fast and accurately as possible to the facial expression by pressing response buttons corresponding to “fear” or “happy” with their index fingers (within a 1000 ms response window). Buttons corresponding to fear and happy facial expressions were counterbalanced across sessions and participants. The task lasted 13 min, and was adapted from Etkin et al. (2006).
Two-choice Reaction Time task
A Two-choice Reaction Time (RT) task was administered as a motor control task (i.e., performance in this task was expected to be unaffected by the training). The target in the task was a blue circle, which was presented for 200 ms and to which participants had to respond within 1500 ms. The target was followed by a variable ITI (1500–2500 ms). Probes, which had the same dimensions as stimuli in the Simon task (3 mm diameter), were presented on either the left or right of a fixation cross (9.2 cm from the fixation cross) and participants were instructed to make corresponding responses with either their left or their right index finger as quickly and accurately as possible. Throughout the task, a central fixation cross was presented. Participants completed 76 trials in one block. The Two-choice RT task lasted approximately 3 min.
Data reduction
Behavioral performance
For the Flanker, Simon, Emotional Face Stroop, and Two-choice RT tasks, only trials with a response were considered (Flanker: 99.87%; Simon: 99.83%; Emotional Face Stroop: 99.85%; Two-choice RT: 99.80%). Trials with RTs (after log10 transformation) ±3 standard deviations outside an individual’s mean for each condition were also excluded (Flanker: 1.32%; Simon: 0.98%; Emotional Face Stroop: 1.50%; Two-choice RT: 0%). The main analyses for Flanker, Simon and Emotional Face Stroop tasks focused on Incongruency and Gratton effects.1
The Incongruency effect is computed by comparing RT [RTIncongruent–RTCongruent] and accuracy [AccuracyCongruent –AccuracyIncongruent] for congruent versus incongruent trials, with higher scores indicating greater interference effects. The Gratton effect (Gratton et al., 1992) is a measure of behavioral adjustments following trials that introduce conflict. It compares RT and accuracy of incongruent trials preceded by an incongruent (I-I trials) versus congruent trials (C-I trials) [RTIncongruent Trials Following Congruent Trials–RTIncongruent Trials Following Incongruent Trials; AccuracyIncongruent Trials Following Incongruent Trials –AccuracyIncongruent Trials Following Congruent Trials]. Higher scores indicate larger Gratton effects and more adaptive behavioral adjustments following incongruent trials. In order to disentangle interference and error effects, only correct trials were considered in the calculation of the Incongruency and Gratton RT effects (Pizzagalli et al., 2006).
For the Emotional Go/NoGo, RT to targets and d′ were calculated. d′ is a measure of sensitivity to the target and was computed by subtracting the z-score of the false alarm rate from the z-score of the hit rate (Macmillan and Creelman, 2005).
ERP data
Event-related potentials were analyzed using Brain Vision Analyzer (Version 1.05; Brain Products GmbH, Gilching, Germany). After manually inspecting continuous EEG data, periods with large movement-related artifacts were removed. Eye-movement artifacts (e.g., blinks) were corrected using an Independent Component Analysis (ICA). Channels with excessive noise were replaced using a spatially weighted linear interpolation of adjacent channels (Hjorth, 1975). Next, segments were extracted 300 ms before and 924 ms after the onset of the Flanker target, followed by automatic artifact rejection (maximal voltage step/gradient: 50 μV; maximal amplitude: ±175 μV; absolute amplitude difference within-segment: 200 μV). Segments were averaged within each subject, band-pass filtered (no additional high-pass, low-pass 30 Hz, 12 dB/octave), baseline-corrected using a 300–100 ms pre-stimulus window (all findings were confirmed using a 300–200 ms baseline), and re-derived to average reference. Grand mean ERP waveforms were averaged across subjects, conditions, and sessions. Topographic maps were created in Brain Vision Analyzer using a linear interpolation algorithm.
In light of prior findings (Holmes and Pizzagalli, 2008; Kopp et al., 1996; Yeung and Cohen, 2006; Yeung et al., 2004), the N2 component was the main variable of interest to examine training effects. Periods for ERP analyses were empirically defined using a space-oriented bootstrapping segmentation procedure (Koenig and Lehmann, 1996), which defined the onset and end point of stable field configurations thought to index specific brain functions, known as “microstates.” Onset and end point for the N2 component were determined by examining the differences in overall field configuration at two successive time frames, as determined by the Global Map Dissimilarity (GMD) value. GMD values range between 0 (identical maps) and 2 (two maps with identical topography but inversed polarity). Peak GMDs occur at times with the greatest difference in field configuration between two successive maps and are used to establish the boundaries of microstates (Lehmann and Skrandies, 1984). Microstate boundaries were confirmed through visual inspection of ERP waveforms.
Based on prior literature (Holmes and Pizzagalli, 2008) and sensors showing maximal deflections in the current study, amplitudes were extracted from the following fronto-central channels: F1, Fz, F2, FC1, FCz, FC2, C1, Cz, C2, Cp1, Cpz, and Cp2. All analyses were replicated at site Cz, the canonical electrode for N2 (Kopp et al., 1996). N2 amplitude was calculated as the average voltage amplitude 244–332 ms following the onset of the Flanker target. To limit the number of tests and based on prior findings (Kopp et al., 1996), follow-up correlational analyses between behavioral performance and ERP amplitude considered N2 amplitude extracted at Cz. On average, a minimum of 74.50 trials were available for ERP analyses (see Supplemental Materials).
Statistical analyses
Behavioral data
Training sessions
For the Simon task, a 3 (Session; 1, 2, 3)×2 (Condition; incongruent, congruent) repeated measures analysis of variance (ANOVA) was run to examine RT and accuracy Incongruency effects. For the Gratton effects, a similar ANOVA was used except the two levels of Condition were I-I and C-I trials. For the Emotional Go/NoGo, d′ values and RT were entered into separate Session (1, 2, 3)×Emotion (positive, negative, neutral) ANOVAs.
Pre-/post-training
Similar to the Simon task analysis, a repeated measures ANOVA was employed to examine Incongruency and Gratton effects in the Flanker task, with the exception that Session contained two levels (pre-, post-training). Emotional Face Stroop data were entered in similar ANOVAs for Incongruency and Gratton effects with an additional 2-level factor of Emotion (fear, happy facial expression). Because Emotion effects are secondary to study hypotheses, they are reported in the Supplemental Materials. For the Two-choice RT task, separate paired t-tests were conducted to compare pre- vs. post-training performance (RT and accuracy).
For all ANOVAs with a Condition factor, Condition×Session interactions were the main effects of interest. Significant ANOVA effects were followed-up using Newman–Keuls tests. When the assumption of sphericity was violated, a Greenhouse–Geiser correction was used. Partial η2 was reported for all effect sizes. For all ANOVAs examining behavioral data, n=20.
ERP data
N2 mean amplitudes were entered into a 4 (Caudality; F, FC, C, Cp)×3 (Laterality; left, central, right)×2 (Session; pre-, post-training)×2 (Condition; incongruent, congruent) ANOVA (n=18).2
Correlation of change scores
To examine putative transfer effects between tasks, Pearson correlations were computed between variables across sessions. First, to account for the relative difference between incongruent and congruent trials, we calculated Incongruency effects for Flanker RT and Flanker N2 amplitude (at Cz) by subtracting incongruent from congruent trials within each pre- and post-training session separately [e.g., RTPre-training Session Incongruent Trials –RTPre-training Session Congruent Trials]. Then, to capture training-related changes, we subtracted the pre-training session RT Incongruency effect from the same effect in the post-training session [e.g., RTPre-training Incongruency Effect–RTPost-training Session Incongruency Effect]. An identical subtraction was computed for the N2 amplitude. These double subtractions resulted in an Incongruency effect (i.e. incongruent–congruent) change score (i.e. pre-training–post-training session) for both N2 amplitude and RT.
Next, we ran three correlations for the pre-training and post-training sessions (n=18). First, to test for a relationship between ERP and behavioral data within the pre-training session, we correlated pre-training Incongruency effect N2 amplitude and RT. Second, we conducted the same correlation with post-training N2 and RT data. Third, we correlated the Incongruency effect (i.e. incongruent–congruent) change score (i.e. pre-training–post-training session) for Flanker task N2 and Flanker task RT.
In order to better understand transfer effects, we conducted two additional correlations examining pre-training/post-training data and training session RT. We computed a similar Incongruency effect (i.e. incongruent–congruent) change score (i.e. training session 1–training session 3) to capture relative changes in RT from the beginning to the end of training. We then correlated this training session effect with similar Incongruency effect change scores for pre-training/post training Flanker task 1) RT (n=19) and 2) N2 amplitude (n=17; the participant whose first training day data was lost was excluded from this analysis).
Practice effects
To examine potentially confounding practice effects, we compared changes in RT (n=20) and N2 amplitude at site Cz (n=18) over the course of the pre-training session with similar changes between pre-training and post-training days. If participants showed large reductions in RT and N2 amplitude during the pre-training session and smaller reductions between the pre- and post-training, practice effects may account for the reduction from pre- to post-training. Therefore, N2 amplitude and RT for the pre-training and post-training sessions were divided into “early” (Blocks 1 and 2) and “late” (Blocks 4 and 5) blocks. For each variable, a 4 (Time; pre-training early, pre-training late, post-training early, post-training late)×2 (Condition; congruent, incongruent) ANOVA was conducted.
Results
Training sessions
Table 1 summarizes ANOVA effects for the training sessions.
Table 1.
Summary of ANOVA findings for the training tasks.
| Effect | Contrast | df | F | p | Partial η2 |
|---|---|---|---|---|---|
| A) Simon task | |||||
| Incongruency RT | Condition | 1,19 | 18.39 | .001 | .492 |
| Session | 2,38 | 37.17 | .001 | .662 | |
| C×Sa | 2,38 | 0.88 | .423 | .044 | |
| Incongruency accuracy | Condition | 1,19 | 4.78 | .042 | .201 |
| Session | 2,38 | 8.05 | .001 | .298 | |
| C×S | 2,38 | 0.69 | .510 | .035 | |
| Gratton RT | Condition | 1,19 | 3.19 | .090 | .144 |
| Session | 2,38 | 26.27 | .001 | .580 | |
| C×S | 2,38 | 3.85 | .030 | .168 | |
| Gratton accuracy | Condition | 1,19 | 0.11 | .748 | .006 |
| Session | 2,38 | 3.67 | .035 | .162 | |
| C×S | 2,38 | 0.55 | .581 | .028 | |
| B) Emotional Go/NoGob | |||||
| d′ | Session | 2,38 | 6.19 | .005 | .246 |
| RT | Session | 2,38 | 2.91 | .066 | .133 |
C×S: Condition×Session interaction.
Ancillary effects as a function of Emotion are available in the Supplemental Materials.
Simon task
Incongruency effects
The Condition (incongruent, congruent)×Session (1, 2, 3) ANOVA revealed the expected main effect of Condition for both RT and accuracy scores (Table 1A), which were due to slower and less accurate responses to incongruent relative to congruent trials (Table 2). For both RT and accuracy, the main effect of Session was also significant (Table 1A). Post-hoc Newman–Keuls tests demonstrated significant RT decreases throughout the training sessions (Table 3; Fig. 1A). There were significant increases in accuracy from session 1 to sessions 2 and 3, but no difference between sessions 2 and 3 (Table 3; Fig. 1B). For both RT and accuracy, the Condition×Session interaction was not significant (Table 1A).
Table 2.
Mean (SD) RT and accuracy (averaged across sessions) for the Simon task.
| Condition | Mean (SD) |
|---|---|
| Incongruency RT (ms)** | |
| RT incongruent | 557.22 (66.08) |
| RT congruent | 516.42 (80.09) |
| Incongruency effect | 40.80 (42.54) |
| Incongruency accuracy* | |
| Accuracy incongruent | 0.965 (0.036) |
| Accuracy congruent | 0.979 (0.013) |
| Incongruency effect | 0.015 (0.030) |
| Gratton RT (ms) | |
| C-I trialsa | 558.36 (65.52) |
| I-I trialsb | 553.33 (68.15) |
| Gratton effect | 5.02 (12.58) |
| Gratton accuracy | |
| C-I trialsb | 0.965 (0.040) |
| I-I trialsb | 0.968 (0.027) |
| Gratton effect | 0.003 (0.033) |
Significant difference between congruent and incongruent conditions p<0.05.
Significant difference between congruent and incongruent conditions p<0.001.
Incongruent trials following congruent trials.
Incongruent trials following incongruent trials.
Table 3.
Mean (SD) RT and accuracy for Incongruency and Gratton trials across training sessions (averaged across conditions) for the Simon task.
| Session 1 | Session 2 | Session3 | |
|---|---|---|---|
| Incongruency trials | |||
| RT (ms)a | 575.74 (63.40) | 529.99 (77.10) | 504.73 (79.02) |
| Accuracyb | 0.964 (0.022) | 0.974 (0.029) | 0.979 (0.024) |
| Gratton trials | |||
| RT (ms)b | 594.94 (59.99) | 545.90 (73.67) | 526.70 (78.40) |
| Accuracyc | 0.958 (0.028) | 0.965 (0.043) | 0.976 (0.033) |
For Incongruency, significant RT reductions were observed between each successive session (all pairwise p<0.05).
For Incongruency accuracy and Gratton RT, significant differences were observed between sessions 1 and 2, but not between sessions 2 and 3 (all pairwise p<0.05).
For Gratton trials, accuracy was significantly increased from session 1 to 3, but neither session were significantly different from session 2 (p<0.05).
Fig. 1.

Summary of Simon task behavioral findings for the training sessions (n=20). Mean RT (A) and accuracy (B) across training sessions. (C) Significant Condition×Session interaction for Gratton effect RT. Compared with C-I trials, I-I trials showed significantly faster RT in sessions 1 and 2 but not in session 3. (D) Mean accuracy for the Gratton trials. Error bars denote S.E.
Gratton effects
The Condition (C-I, I-I trials)×Session (1, 2, 3) confirmed a main effect of Session for both RT and accuracy (Table 1A). For RT, the effects were due to faster RT from session 1 to sessions 2 and 3 but no difference between sessions 2 and 3 (Table 3; Fig. 1C). For accuracy, participants demonstrated greater accuracy in session 3 relative to session 1 (Table 3; Fig. 1D). For RT, the Session main effect was qualified by a significant Session×Condition interaction (Table 1A), indicating that the Gratton effect differed across sessions. Post-hoc tests indicated that RT was faster to I-I versus C-I trials during session 1 (590.08±60.12 vs. 599.81±61.50 ms, p<.05) and session 2 (541.64±76.01 vs. 550.16±72.12 ms, p>.05), but not in session 3 (528.28±82.86 vs. 525.11±74.87 ms, p>.30; Fig. 1C). Thus, a significant Gratton effect was seen in sessions 1 and 2 but not in session 3.
Collectively, these findings indicate that the Simon training task elicited (1) intended behavioral effects, (2) robust and systematic behavioral improvement over the course of the training sessions, and (3) improved cognitive control at the end of the training, as manifested by the absence of the Gratton effect in session 3.
Emotional Go/NoGo
The Emotion×Session ANOVA for d′ revealed a main effect of Session (Table 1B) due to significant improvement in d′ from training session 1 (2.81±0.48) to session 2 (3.03±0.60; p<.01) but not from session 2 to session 3 (3.05±.47, p>.7). (For more information, as well as effects of Emotion, see the Supplemental Materials).
Pre-/post-training sessions
Table 4 summarizes ANOVA effects for the Flanker task.
Table 4.
Summary of ANOVA findings for the pre-/post-training Flanker task.
| Effect | Contrast | df | F | p | Partial η2 |
|---|---|---|---|---|---|
| A) Behavioral performance | |||||
| Incongruency RT | Condition | 1,19 | 132.27 | .001 | .874 |
| Session | 1,19 | 2.60 | .124 | .120 | |
| C×Sa | 1,19 | 20.79 | .001 | .522 | |
| Incongruency accuracy | Condition | 1,19 | 17.42 | .001 | .478 |
| Session | 1,19 | 1.89 | .185 | .091 | |
| C×S | 1,19 | 2.31 | .145 | .109 | |
| Gratton RT | Condition | 1,19 | 13.54 | .002 | .416 |
| Session | 1,19 | 7.04 | .016 | .270 | |
| C×S | 1,19 | 0.01 | 0.91 | .001 | |
| Gratton accuracy | Condition | 1,19 | 14.80 | .001 | .438 |
| Session | 1,19 | 1.10 | .307 | .055 | |
| C×S | 1,19 | 4.91 | .039 | .205 | |
| B) Event-related potential | |||||
| N2 (244–332 ms) | Condition | 1,17 | 10.91 | .004 | .391 |
| Session | 1,17 | 4.32 | .053 | .203 | |
| C×S | 1,17 | 10.71 | .004 | .387 | |
C×S: Condition×Session interaction.
Flanker task: behavioral data
Incongruency effects
The Session (pre-, post-training)×Condition (incongruent, congruent) ANOVA revealed the expected main effects of Condition for both RT and accuracy (Table 4A). These effects were due to increased RT and lower accuracy for incongruent relative to congruent trials (Table 5A). Critically, for RT, a significant Session×Condition interaction emerged (Table 4A; Fig. 2A). Post-hoc Newman–Keuls tests revealed that RT for incongruent trials decreased significantly from pre-training to post-training (399.23± 38.90 vs. 380.96±43.52 ms, p<.001; Cohen d value: .609), while congruent trials remained unchanged (310.55±43.99 vs. 308.96± 48.95 ms, p>.5; Fig. 2A). No other effects emerged (Table 4A; Fig. 2B).
Table 5.
Mean (SD) RT and accuracy (averaged across sessions) for the Flanker task.
| Flanker task | Mean (SD) |
|---|---|
| A) Behavioral performance | |
| Incongruency RT** | |
| RT incongruent | 390.10 (38.45) |
| RT congruent | 309.75 (44.47) |
| Incongruency effect | 80.34 (31.24) |
| Incongruency accuracy** | |
| Accuracy incongruent | 0.865 (0.138) |
| Accuracy congruent | 0.989 (0.012) |
| Incongruency effect | 0.125 (0.133) |
| Gratton RT* | |
| C-I trialsa | 390.43 (38.02) |
| I-I trialsb | 384.56 (39.95) |
| Gratton effect | 5.88 (7.14) |
| Gratton accuracy* | |
| C-I trialsa | 0.854 (0.145) |
| I-I trialsb | 0.933 (0.073) |
| Gratton effect | 0.080 (0.093) |
| B) Event-related potentials | |
| N2 (244–332 ms)* | |
| Incongruent | −0.14 (2.34) |
| Congruent | 0.58 (1.92) |
Significant difference between conditions p<0.005.
Significant difference between conditions p<0.001.
Incongruent trials following congruent trials.
Incongruent trials following incongruent trials.
Fig. 2.
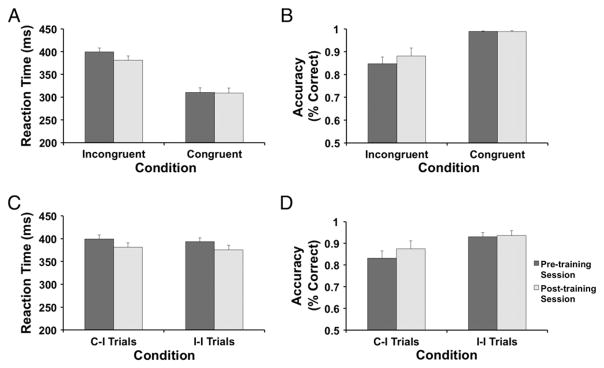
Summary of Flanker task behavioral findings for the pre-/post-training sessions (n=20). Mean RT (A) and accuracy (B) for each session separately. For RT, a significant Condition×Session interaction emerged, due to significantly faster RT for incongruent – but not congruent – trials from pre- to post-training. (C) Mean for Gratton effect RT. (D) A significant Condition×Session interaction for Gratton effect accuracy. Following training, C-I trials showed a greater increase in accuracy than I-I trials. Error bars denote S.E.
Gratton effects
The Session (pre-, post-training)×Condition (C-I, I-I trials) ANOVAs revealed main effects of Condition for both the RT and accuracy (Table 4A) scores, due to the expected decreased RT and higher accuracy for I-I trials versus C-I trials (Table 5A). For RT, a main effect of Session also emerged (Table 4A, Fig. 2C), due to a general decrease in RT from the pre- to post-training session (396.43±38.32 vs. 378.56± 44.73 ms, p<.05). Critically, for accuracy, the main effect of Condition was qualified by a significant Session×Condition interaction (Table 4A), due to significantly improved accuracy pre-to-post for C-I trials (pre: 0.832±0.149, post: 0.875±0.159, p<.001; Cohen d value: −0.404), but not I-I trials (pre: 0.931±0.083, post: 0.936±0.100, p>.50; Fig. 2D).
Collectively, these findings indicate that the Flanker task elicited (1) intended behavioral effects, (2) pre- to post-training behavioral improvements, and (3) improved cognitive control post-training, as manifested by RT speeding for high interference (incongruent) trials and better accuracy for C-I trials after training.
Flanker task: ERP data
ANOVA results for the ERP data are summarized in Table 4B. For brevity, only effects involving Session and Condition will be discussed. For complete ANOVA results, see Table S4 in the Supplemental Materials.
N2
The Caudality (F, FC, C, Cp)×Laterality (left, central, right)×Session (pre-, post-training)×Condition (incongruent, congruent) ANOVA revealed a main effect of Condition (Table 4B); as expected based on prior studies (e.g., Folstein and Van Petten, 2008; Kopp et al., 1996), incongruent trials elicited significantly more negative N2 amplitude compared to congruent trials (Table 5B). The effect of Session reached trend level (Table 4B), with more negative N2 amplitude for pre-training than post-training session (−0.08±2.20 μV vs. 0.52± 2.16 μV, p<.06). Critically, these effects were qualified by a significant Session×Condition interaction (Table 4B). Post-hoc Newman–Keuls tests revealed significantly smaller N2 for incongruent trials in the post-relative to pre-training (0.34±2.50 μV vs. −0.62±2.35 μV; p<.001; Cohen d value: −0.753), whereas there was no change for congruent trials (pre: 0.47±2.14 μV vs. post: 0.69±1.91 μV; p>.15; Figs. 3A, Fig. 4).
Fig. 3.

Summary of ERP findings. (A) Target-locked mean waveforms (averaged across channels F1, Fz, F2, FC1, FCz, FC2, C1, Cz, C2, Cp1, Cpz, Cp2) for incongruent and congruent trials during pre- and post-training sessions. (B) Correlation between change (pre-training session–post-training session) in interference effect (incongruent–congruent) RT and mean N2 amplitude at site Cz in the Flanker task (r=.45, p<.06). (C) Correlation between change (training session 1–training session 3) in interference effect (incongruent–congruent) RT for the Simon task and Incongruency effect change (pre-–post-training) in mean N2 amplitude at site Cz in the Flanker Task (r=.56, p<.05). This latter correlation highlights transfer effects.
Fig. 4.

Topographic map of the N2 component for incongruent and congruent trials during the (A) pre-training session and (B) post-training session. Compared with the pre-training session, post-training session responses to incongruent trials were significantly reduced. Potential response to congruent trials shows no difference from pre- to post-training. Although the map has a more anterior distribution in the post- relative to the pre-training session, the Caudality (F, FC, C, Cp)×Session (pre-, post-training), Caudality×Session×Condition (incongruent, congruent) and Caudality×Laterality (left, central, right)×Session×Condition interactions were not significant (see also Table S4).
The ANOVA also demonstrated a significant Laterality×Condition× Session interaction [F(1,17)=4.70, p<.05; partial η2=.217]. Follow-up Session×Condition ANOVAs revealed a significant Session×Condition interaction at each level of Laterality, but the effect was strongest for central electrodes [Central, F(1,17)=16.51, p=.001; partial η2=.493; Right, F(1,17)=5.01, p=.039; partial η2=.228; Left, F(1,17)=7.48, p=.014; partial η2=.305].
Emotional Face Stroop task
For brevity, only effects involving Condition or Session are discussed. The Emotion×Condition×Session ANOVAs for RT and accuracy for Incongruency and Gratton effects revealed main effects of Condition with congruent and I-I trials eliciting expected increased accuracy and reduced RTs (Table S2 and S3), compared with incongruent and C-I trials, respectively (Fs>11.00, ps<.005). There were no effects of Session or Session×Condition interactions (Fs<1.6, ps>.2). (For the Condition ANOVA results, Condition means and effects of Emotion, see the Supplemental Materials).
Two-choice RT task
For RT, no significant difference emerged between pre- and post-training sessions [t>−2.00, p>.10]. In spite of ceiling effects, accuracy improved from the pre- (0.9921±0.0079) to post- (0.9967± 0.0059) training session [t(19)=−2.34, p<.05, η2=.223].
Correlation between behavioral data and Flanker task ERPs
Correlations between N2 amplitudes at site Cz and RT for Incongruency effects in the Flanker task showed no relationship within either the pre- or post-training session (pre: r=.23; post: r=.16, ps>.30). However, when examining the relationship between change (i.e. pre-–post-training) in Incongruency effects for N2 amplitude and Flanker RT, a trending correlation emerged (r=.45, p<.06; Fig. 3B). Accordingly, participants showing the largest reduction in Incongruency effect for N2 amplitude from pre- to post-training were also characterized by the largest reduction in Flanker RT interference effect.
Particularly important for the study hypotheses, a relationship between pre-to-post changes in Incongruency effect (i.e., congruent–incongruent) for Flanker N2 amplitude and training-induced changes (i.e., Session 1–Session 3) in interference effects (i.e. incongruent–congruent) during the Simon task RT emerged (r=.56, p<.05; Fig. 3C). Thus, participants showing the largest reduction in RT interference in the Simon task from session 1 to session 3 also showed the largest reduction in Flanker N2 amplitude from the pre- to post-training.
Correlations of RT interference changes between the Simon and Flanker Tasks were not significant (r=.13, p>.5).
Practice effects
To examine potential practice effects that would explain improvements from pre- to post-training, a separate 4 (Time; pre-training early, pre-training late, post-training early, post-training late)×2 (Condition; congruent, incongruent) ANOVA was conducted for RT and N2 amplitude at Cz. Both ANOVAs revealed a main effect of Condition [RT: F(1,20)=138.35, p<.001; partial η2=.874; N2: F(1,17)= 11.32, p<.005; partial η2=.400] but no effect of Time (ps>.06). Both ANOVAs revealed significant Time×Condition interactions [RT: F(1,20)=23.57, p<001; partial η2=.541; N2: F(1,17)=4.58, p<.01, partial η2=.212].
Post-hoc Newman–Keuls tests revealed a significant increase in RT for incongruent trials from blocks 1–2 to blocks 4–5 within the pre-training session (p<.05) but no change for congruent trials (p>.2). This indicates that the participants displayed poorer performance for incongruent trials over the course of the pre-training session. However, when comparing the pre- and post-training sessions (i.e. blocks 4–5 of the pre-training session with blocks 1–2 of the post-training), post-hoc tests showed a significant decrease in RT for incongruent trials (p<.001), but no difference for congruent trials (p>.2). Thus, RT only improved for incongruent trials and only from before-to-after the training.
There were similar results for the N2 amplitude. Post-hoc Newman–Keuls tests revealed no differences between blocks 1–2 and blocks 4–5 of the pre-training session for incongruent or congruent trials (p>.2), indicating no changes with the pre-training session. There was, however, a significant decrease in amplitude for incongruent (p<.001), but not congruent (p>.2) trials, when comparing blocks 4–5 of the pre-training session with blocks 1–2 of the post-training session, suggesting that performance enhancements took place during the training days. These findings suggest that pre-to-post-training reductions in RT and N2 amplitude did not occur as the result of practice effects from the pre-training session, but instead due to the three intervening training sessions.
Discussion
The aim of the current study was to investigate whether cognitive control can be trained and whether improvements in cognitive control would transfer to an untrained task assumed to engage similar mechanisms. While prior studies have examined training effects on working memory (Dahlin et al., 2008a; Jaeggi et al., 2008; Klingberg, 2010; McNab et al., 2009; Olesen et al., 2003; Westerberg and Klingberg, 2007) and executive function (Diamond et al., 2007; Moreno et al., 2011; Thorell et al., 2009), no study has shown training and transfer effects with tasks requiring interference resolution, such as the Flanker and Simon tasks, in adults (for initial evidence in children, see Rueda et al., 2005). The current brief training consisted of three consecutive days in which participants performed the Simon task as well as an Emotional Go/NoGo also known to engage frontocingulate pathways (e.g. Chiu et al., 2008; Elliott et al., 2000). We assessed pre- and post-training performance with the Flanker task while collecting ERPs. Several important findings emerged.
First, both behavioral (RT and accuracy) and ERP (N2) data confirmed expected Incongruency effects (e.g., longer RT, lower accuracy and larger N2 for incongruent vs. congruent trials), indicating that tasks elicited the intended effects. Second, a robust and systematic behavioral improvement over the course of the training sessions was evident; this culminated in improved cognitive control at the end of the training, as manifested by the absent Gratton effect in session 3. Third, and most importantly, following training on the Simon and Emotional Go/NoGo task, participants demonstrated improved behavioral performance on the Flanker task, as manifested by faster RTs on trials requiring interference resolution (incongruent trials) while maintaining similar accuracy from pre- to post-training and better accuracy in C-I trials (i.e. trials requiring increased cognitive control). This selective RT speeding was complemented by a hypothesized reduction in N2 amplitude for incongruent, but not congruent, trials. Fourth, control analyses indicated that pre-to-post reductions in both RT and N2 were not the result of practice effects. Finally, participants showing the greatest pre-to-post decrease in Incongruency effects for N2 amplitude displayed the largest reductions in RT interference effects for the Flanker and Simon tasks.
The selective reductions in RT and N2 amplitude for incongruent trials suggest that mechanisms of interference resolution were improved. Support that training led to this enhanced interference resolution stems from the correlation between reduced pre-to-post N2 amplitude and decreased RT interference effects over the course of training. Yeung and Cohen (2006) argued that smaller N2 amplitude should result from decreased processing of the irrelevant stimulus dimension of a trial. Thus, there are at least two different general processes that could explain reduced interference: those that resolve conflict within an incongruent trial and those that monitor and call for increased conflict resolution mechanisms following an incongruent trial. The former category of processes may include enhanced “automatic” attentional processes. Cohen et al. (1992) conceptualized automatic versus controlled processing as a spectrum; practice or training causes activities previously requiring controlled processes to become more automatic. They described several features signifying automaticity, including reduced RT, reduced requirements for attention and release from attentional control following practice (Cohen et al., 1992). Similarly, a study showing reduced dACC and DLPFC activity over the course of a practice session was interpreted as evidence for decreased conflict and a concurrent reduced need for control, respectively (Milham et al., 2003).
In general, automatic attentional processes are relaxed during congruent trials, allowing increased processing of task-irrelevant features, leading to increased conflict on an ensuing incongruent trial (Carter and van Veen, 2007). The automating of these attentional processes due to training may have decreased the extent to which attention was relaxed during congruent trials, resulting in less need for effortful control and less conflict during subsequent incongruent trials. Reduction in N2 and RT from pre- to post-training training provides evidence of less conflict and the possibility that attention became more automated. Additionally, when compared with I-I trials, there was a larger reduction in RT for Simon task C-I trials over the course of training and a larger pre-to-post increase in Flanker task C-I trial accuracy. That is, over the course of training, there was a diminishing influence of a congruent trial on a subsequent incongruent trial, providing evidence that the tendency for attentional processes to relax during congruent trials was reduced after training. Furthermore, increased attention would not necessarily facilitate congruent trials because there is no interference to inhibit or avoid.
Evidence that training improved processes that monitor and call for increased conflict resolution (i.e., trial-to-trial adaptation) was more limited. In both the Flanker and Simon tasks, I-I trials showed significant reductions in RT from pre- to post-training and over the course of training, respectively,3 but this could be explained by the mechanisms discussed previously. Regardless, reduced N2 amplitude could be the result of increased dACC efficiency or connectivity between dACC and DLPFC, including less dACC activity required to call on the DLPFC to exert control. Future studies using similar training procedures in conjunction with functional magnetic resonance imaging (fMRI) and methods to probe functional connectivity between dACC and DLPFC will be needed to assess whether training strengthened connectivity between these regions, which might support improved I-I trial performance.
There were inconsistencies between the behavioral outcomes for the training sessions and the pre-/post-training sessions. First, the Flanker task showed selective RT speeding for incongruent trials from pre- to post-training, whereas the Simon task displayed increased accuracy and decreased RTs for both congruent and incongruent trials over the course of training. Second, compared to I-I trials, participants showed reduced C-I trial RT during the training but higher C-I trial accuracy during the pre-/post-training task. The robust performance enhancement for all trials during the training is not surprising in light of many studies showing that individuals improve on any task that they perform repeatedly (Owen et al., 2010), especially when practiced several days a week (Jaeggi et al., 2008; Olesen et al., 2003). These robust practice effects could have overshadowed subtler performance markers of enhanced cognitive control behavior during the Simon task. For example, improved interference resolution mechanisms might have resulted in a small decrease in incongruent trial RT over the course of training, similar to the Flanker task. But larger practice effects causing both incongruent and congruent trial RT to decrease dramatically may have dwarfed this improvement. The larger RT reduction for high-conflict C-I trials, compared with I-I trials, suggests that, in addition to large practice effects, there were increases in cognitive control.
Incentivizing participants to focus on accuracy during the Simon task may also have contributed to inconsistencies between training and pre-/post-training sessions. We incentivized subjects to perform better than 80% accuracy for incongruent trials, which may have led to slowing on incongruent trials to maintain an acceptable accuracy. Without this modification, participants might have shown even faster RTs for incongruent trials over the course of training. Similar to the previous point, by examining only incongruent trials in the Gratton effect, the larger reduction of C-I trial RT, compared with I-I trials RT, suggests that, when controlling for the accuracy incentives, there was evidence for increased cognitive control. Accuracy incentives may have led participants to increase their focus on incongruent trials, potentially enhancing the training effect. However, the incentives may have also produced altered performance during the training period, making it more difficult to relate behavioral outcomes between the training and pre-/post-training sessions for both Incongruency and Gratton effects.
The Emotional Face Stroop and Emotional Go/NoGo tasks were included to assess potential training-induced improvements in mechanisms involved in emotional conflict. Both tasks have been shown to activate rostral ACC (rACC), an area associated with emotional interference resolution (Chiu et al., 2008; Etkin et al., 2006). Our findings, however, demonstrate a clear lack of training. Although the Emotional Face Stroop robustly elicited expected effects, there was no improvement pre- to post-training. One potential explanation for these findings could be that the Emotional Go/NoGo and Simon training tasks rely on different interference resolution mechanisms than the Emotional Face Stroop task, resulting in a lack of transfer effects. Consistent with this speculation, studies using Emotional Go/NoGo tasks have described rACC activation when comparing emotional to non-emotional targets, but right ventrolateral frontal cortex activation in response to trials requiring inhibitory control (NoGo trials; Chiu et al., 2008; Elliott et al., 2000). This suggests the rACC may not be involved in the mechanisms of interference resolution for the Emotional Go/NoGo. Regardless of the possible mechanisms, training failed to affect emotional interference processes likely localized to the rACC.
Several important limitations of the current study deserve mention. Without a control group, we cannot be certain that the observed improvements were the result of specific changes in cognitive control processes, rather than unspecified, more general mechanisms. The fact that several findings were specific to incongruent versus congruent and C-I (i.e., high-conflict) versus I-I trials (i.e., low-conflict) speaks against global effects. Nevertheless, compared with congruent trials, there was a greater possibility that incongruent trials would show reduced RT from pre- to post-training because incongruent trials elicit slower responses at the outset. In other words, there was more room for improvement for incongruent trials. In contrast, floor effects may have limited the possibility of reduced RT following training for congruent trials. Floor effects also may have limited the usefulness of the Two-choice RT task as a control measure for practice effects because of the minimal cognitive demands the task requires. Second, testing both emotional and cognitive tasks within one group of participants complicated the interpretation of the results. Including both types of tasks may have provided insight into mechanisms underlying emotional conflict tasks, a topic which is currently debated (Etkin et al., 2006; Haas et al., 2006; Mohanty et al., 2007; Ochsner et al., 2009). For example, if improvements in Emotional Face Stroop performance were associated with Simon task training improvements, it would have suggested the presence of overlapping mechanisms responsible for conflict in both emotional and non-emotional conflict tasks. Given the lack of improvement in the Emotional Face Stroop task, this possibility was not examined. A final limitation is that using a button-box during the pre-/post-training sessions and a keyboard during the training session may have created additional noise in RT variables and contributed to the lack of correlation between tasks in the different session types.
In conclusion, the current findings indicate that cognitive control, specifically mechanisms aiding in interference resolution, can be improved by a brief training, and that improvements transfer to a similar but untrained task. In addition to highlighting plasticity in cognitive control processes, the current data showcase promising avenues for populations characterized by impaired cognitive control, including neuropsychiatric and neurodegenerative disorders.
Supplementary Material
Acknowledgments
The authors would like to thank Avram Holmes, Ryan Bogdan, Daniel Dillon, Pia Pechtel, Sunny Dutra, Marie-Anne Vanderhasselt, Richard McNally and Matthew Nock for their assistance or input. This study was supported by a Harvard University Restricted Funds awarded to AJM, and by NIMH (R01MH68376) awarded to DAP. The funding sources had no role in study design; collection, analysis and interpretation of data; preparation of the manuscript; and the decision to submit the manuscript for publication.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.neuroimage.2012.07.032.
Footnotes
Previous presentation. Portions of these data have been presented in preliminary form at the Association for Psychological Science annual convention (2010, May; Boston) and the annual meeting of the Cognitive Neuroscience Society (2011, April; San Francisco).
This article is dedicated to Harish Chamarthi, who prematurely passed away in January, 2011. His courage to re-dedicate his life to learning and working for the social good is a source of inspiration.
The low rate of incorrect responses precluded analyses of post-error behavioral adjustments. Specifically, only four participants averaged more than 3 errors per session over the course of the training period. Furthermore, 10 out of 21 participants had less than 5 errors in either the pre- or post-training session.
Due to artifacts, there were too few I-I trials (pre: 13.22±3.12, post: 15.56±5.14) to conduct an ANOVA for Gratton effects in the ERP data. Although some groups have suggested that 6–8 error trials are sufficient to elicit a stable error-related negativity component (Olvet and Hajcak, 2009), others have argued that 20 target trials are required for other ERP components (Cohen and Polich, 1997). Given the low number of I-I trials and the fact that two subjects had at least one session with less than 6 total I-I trials, we decided to forgo analysis of Gratton effects.
Paired t-tests revealed significantly decreased RT from (1) the pre- to post-training for Flanker I-I trials [t(19)=2.25, p<.05], and (2) session 1 to session 3 for Simon I-I RT [t(19)=5.51, p<.001]. As described in the main text, Flanker I-I accuracy did not change from pre- to post-training. Simon I-I accuracy showed a trend-level improvement from session 1 to session 3 [t(19)=− 1.98, p<.07]. Although processes leading to improved resolution can explain reduced I-I trial RT, I-I performance could have also been enhanced by increased efficiency in mechanisms monitoring and calling for increased cognitive control (i.e. trial-to-trial adaptation).
Disclosures
Dr. Pizzagalli has received consulting fees from AstraZeneca, Ono Pharma USA, Shire and Johnson and Johnson for projects unrelated to the current research. All other authors report no competing interests.
References
- Banich MT. Executive function: the search for an integrated account. Curr Dir Psychol Sci. 2009;18:89–94. [Google Scholar]
- Bartholow BD. On the role of conflict and control in social cognition: event-related brain potential investigations. Psychophysiology. 2010;47:201–212. doi: 10.1111/j.1469-8986.2009.00955.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Botvinick M, Nystrom LE, Fissell K, Carter CS, Cohen JD. Conflict monitoring versus selection-for-action in anterior cingulate cortex. Nature. 1999;402:179–181. doi: 10.1038/46035. [DOI] [PubMed] [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD. Conflict monitoring and cognitive control. Psychol Rev. 2001;108:624–652. doi: 10.1037/0033-295x.108.3.624. [DOI] [PubMed] [Google Scholar]
- Botvinick MM, Cohen JD, Carter CS. Conflict monitoring and anterior cingulate cortex: an update. Trends Cogn Sci. 2004;8:539–546. doi: 10.1016/j.tics.2004.10.003. [DOI] [PubMed] [Google Scholar]
- Bradley MM, Lang PJ. Affective Norms for English Words (ANEW): Stimuli, Instruction Manual and Affective Ratings. The Center for Research in Psychophysiology, University of Florida; Gainesville, FL: 1999. [Google Scholar]
- Carter CS, van Veen V. Anterior cingulate cortex and conflict detection: an update of theory and data. Cogn Affect Behav Neurosci. 2007;7:367–379. doi: 10.3758/cabn.7.4.367. [DOI] [PubMed] [Google Scholar]
- Chiu PH, Holmes AJ, Pizzagalli DA. Dissociable recruitment of rostral anterior cingulate and inferior frontal cortex in emotional response inhibition. Neuroimage. 2008;42:988–997. doi: 10.1016/j.neuroimage.2008.04.248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clayson PE, Larson MJ. Conflict adaptation and sequential trial effects: support for the conflict monitoring theory. Neuropsychologia. 2011;49:1953–1961. doi: 10.1016/j.neuropsychologia.2011.03.023. [DOI] [PubMed] [Google Scholar]
- Cohen J, Polich J. On the number of trials needed for P300. Int J Psychophysiol. 1997;25:249–255. doi: 10.1016/s0167-8760(96)00743-x. [DOI] [PubMed] [Google Scholar]
- Cohen JD, Servan-Schreiber D, McClelland JL. A parallel distributed processing approach to automaticity. Am J Psychol. 1992;105:239–269. [PubMed] [Google Scholar]
- Dahlin E, Neely AS, Larsson A, Backman L, Nyberg L. Transfer of learning after updating training mediated by the striatum. Science. 2008a;320:1510–1512. doi: 10.1126/science.1155466. [DOI] [PubMed] [Google Scholar]
- Dahlin E, Nyberg L, Bäckman L, Neely AS. Plasticity of executive functioning in young and older adults: immediate training gains, transfer, and long-term maintenance. Psychol Aging. 2008b;23:720–730. doi: 10.1037/a0014296. [DOI] [PubMed] [Google Scholar]
- Diamond A, Barnett WS, Thomas J, Munro S. The early years: preschool program improves cognitive control. Science. 2007;318:1387–1388. doi: 10.1126/science.1151148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elliott R, Rubinsztein JS, Sahakian BJ, Dolan RJ. Selective attention to emotional stimuli in a verbal go/no-go task: an fMRI study. Neuroreport. 2000;11:1739–1744. doi: 10.1097/00001756-200006050-00028. [DOI] [PubMed] [Google Scholar]
- Erickson KI, Colcombe SJ, Wadhwa R, Bherer L, Peterson MS, Scalf PE, et al. Training-induced functional activation changes in dual-task processing: an fMRI study. Cereb Cortex. 2006;17:192–204. doi: 10.1093/cercor/bhj137. [DOI] [PubMed] [Google Scholar]
- Etkin A, Egner T, Peraza DM, Kandel ER, Hirsch J. Resolving emotional conflict: a role for the rostral anterior cingulate cortex in modulating activity in the amygdala. Neuron. 2006;51:871–882. doi: 10.1016/j.neuron.2006.07.029. [DOI] [PubMed] [Google Scholar]
- Fan J, Flombaum JI, McCandliss BD, Thomas KM, Posner MI. Cognitive and brain consequences of conflict. Neuroimage. 2003;18:42–57. doi: 10.1006/nimg.2002.1319. [DOI] [PubMed] [Google Scholar]
- Folstein JR, Van Petten C. Influence of cognitive control and mismatch on the N2 component of the ERP: a review. Psychophysiology. 2008;45:152–170. doi: 10.1111/j.1469-8986.2007.00602.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francis WN, Kučera H. A Standard Corpus of Present-day Edited American English. Brown University Department of Linguistics; Providence, Rhode Island: 1979. [Google Scholar]
- Francis WN, Kučera H, Mackie AW. Frequency Analysis of English Usage: Lexicon and Grammar. Houghton Mifflin; Boston: 1982. [Google Scholar]
- Gratton G, Coles MG, Donchin E. Optimizing the use of information: strategic control of activation of responses. J Exp Psychol Gen. 1992;121:480–506. doi: 10.1037//0096-3445.121.4.480. [DOI] [PubMed] [Google Scholar]
- Gross JJ, Muñoz RF. Emotion regulation and mental health. J Clin Sci Pract. 1995;2:151–164. [Google Scholar]
- Haas BW, Omura K, Constable RT, Canli T. Interference produced by emotional conflict associated with anterior cingulate activation. Cogn Affect Behav Neurosci. 2006;6:152–156. doi: 10.3758/cabn.6.2.152. [DOI] [PubMed] [Google Scholar]
- Haslinger B, Erhard P, Altenmuller E, Hennenlotter A, Schwaiger M, Grafin von Einsiedel H, et al. Reduced recruitment of motor association areas during bimanual coordination in concert pianists. Hum Brain Mapp. 2004;22:206–215. doi: 10.1002/hbm.20028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hjorth B. An on-line transformation of EEG scalp potentials into orthogonal source derivations. Electroencephalogr Clin Neurophysiol. 1975;39:526–530. doi: 10.1016/0013-4694(75)90056-5. [DOI] [PubMed] [Google Scholar]
- Holmes AJ, Pizzagalli DA. Task feedback effects on conflict monitoring and executive control: relationship to subclinical measures of depression. Emotion. 2007;7:68–76. doi: 10.1037/1528-3542.7.1.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes AJ, Pizzagalli DA. Response conflict and frontocingulate dysfunction in unmedicated participants with major depression. Neuropsychologia. 2008;46:2904–2913. doi: 10.1016/j.neuropsychologia.2008.05.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes J, Gathercole SE, Dunning DL. Adaptive training leads to sustained enhancement of poor working memory in children. Dev Sci. 2009a;12:F9–F15. doi: 10.1111/j.1467-7687.2009.00848.x. [DOI] [PubMed] [Google Scholar]
- Holmes J, Gathercole SE, Place M, Dunning DL, Hilton KA, Elliott JG. Working memory deficits can be overcome: Impacts of training and medication on working memory in children with ADHD. Appl Cogn Psychol. 2009b;24:827–836. [Google Scholar]
- Holmes AJ, Bogdan R, Pizzagalli DA. Serotonin transporter genotype and action monitoring dysfunction: a possible substrate underlying increased vulnerability to depression. Neuropsychopharmacology. 2010;35:1186–1197. doi: 10.1038/npp.2009.223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaeggi SM, Buschkuehl M, Jonides J, Perrig WJ. Improving fluid intelligence with training on working memory. Proc Natl Acad Sci U S A. 2008;105:6829–6833. doi: 10.1073/pnas.0801268105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kerns JG. Anterior cingulate and prefrontal cortex activity in an fMRI study of trial-to-trial adjustments on the Simon task. Neuroimage. 2006;33:399–405. doi: 10.1016/j.neuroimage.2006.06.012. [DOI] [PubMed] [Google Scholar]
- Kerns JG, Cohen JD, MacDonald AW, Cho RY, Stenger VA, Carter CS. Anterior cingulate conflict monitoring and adjustments in control. Science. 2004;303:1023–1026. doi: 10.1126/science.1089910. [DOI] [PubMed] [Google Scholar]
- Klingberg T. Training and plasticity of working memory. Trends Cogn Sci. 2010;14:317–324. doi: 10.1016/j.tics.2010.05.002. [DOI] [PubMed] [Google Scholar]
- Klingberg T, McNab F. Working memory remediation and the D1 receptor. Am J Psychiatry. 2009;166:515–516. doi: 10.1176/appi.ajp.2009.09030343. [DOI] [PubMed] [Google Scholar]
- Klingberg T, Forssberg H, Westerberg H. Training of working memory in children with ADHD. J Clin Exp Neuropsychol. 2002;24:781–791. doi: 10.1076/jcen.24.6.781.8395. [DOI] [PubMed] [Google Scholar]
- Klingberg T, Fernell E, Olesen PJ, Johnson M, Gustafsson P, Dahlström K, et al. Computerized training of working memory in children with ADHD—a randomized, controlled trial. J Am Acad Child Adolesc Psychiatry. 2005;44:177–186. doi: 10.1097/00004583-200502000-00010. [DOI] [PubMed] [Google Scholar]
- Koenig T, Lehmann D. Microstates in language-related brain potential maps show noun–verb differences. Brain Lang. 1996;53:169–182. doi: 10.1006/brln.1996.0043. [DOI] [PubMed] [Google Scholar]
- Kopp B, Rist F, Mattler U. N200 in the flanker task as a neurobehavioral tool for investigating executive control. Psychophysiology. 1996;33:282–294. doi: 10.1111/j.1469-8986.1996.tb00425.x. [DOI] [PubMed] [Google Scholar]
- Lehmann D, Skrandies W. Spatial analysis of evoked potentials in man—a review. Prog Neurobiol. 1984;23:227–250. doi: 10.1016/0301-0082(84)90003-0. [DOI] [PubMed] [Google Scholar]
- Li SC, Schmiedek F, Huxhold O, Röcke C, Smith J, Lindenberger U. Working memory plasticity in old age: practice gain, transfer, and maintenance. Psychol Aging. 2008;23:731–742. doi: 10.1037/a0014343. [DOI] [PubMed] [Google Scholar]
- Macmillan NA, Creelman CD. Detection Theory: A User’s Guide. 2. Lawrence Erlbaum Associates; Mahwah, N.J: 2005. [Google Scholar]
- Mayr U, Awh E, Laurey P. Conflict adaptation effects in the absence of executive control. Nat Neurosci. 2003;6:450–452. doi: 10.1038/nn1051. [DOI] [PubMed] [Google Scholar]
- McNab F, Varrone A, Farde L, Jucaite A, Bystritsky P, Forssberg H, et al. Changes in cortical dopamine D1 receptor binding associated with cognitive training. Science. 2009;323:800–802. doi: 10.1126/science.1166102. [DOI] [PubMed] [Google Scholar]
- Meister I, Krings T, Foltys H, Boroojerdi B, Müller M, Töpper R, et al. Effects of long-term practice and task complexity in musicians and nonmusicians performing simple and complex motor tasks: implications for cortical motor organization. Hum Brain Mapp. 2005;25:345–352. doi: 10.1002/hbm.20112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milham MP, Banich MT, Claus ED, Cohen NJ. Practice-related effects demonstrate complementary roles of anterior cingulate and prefrontal cortices in attentional control. Neuroimage. 2003;18:483–493. doi: 10.1016/s1053-8119(02)00050-2. [DOI] [PubMed] [Google Scholar]
- Mohanty A, Engels AS, Herrington JD, Heller W, Ho MHR, Banich MT, et al. Differential engagement of anterior cingulate cortex subdivisions for cognitive and emotional function. Psychophysiology. 2007;44:343–351. doi: 10.1111/j.1469-8986.2007.00515.x. [DOI] [PubMed] [Google Scholar]
- Moreno S, Bialystok E, Barac R, Schellenberg EG, Cepeda NJ, Chau T. Short-term music training enhances verbal intelligence and executive function. Psychol Sci. 2011;22:1425–1433. doi: 10.1177/0956797611416999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mukai I, Kim D, Fukunaga M, Japee S, Marrett S, Ungerleider LG. Activations in visual and attention-related areas predict and correlate with the degree of perceptual learning. J Neurosci. 2007;27:11401–11411. doi: 10.1523/JNEUROSCI.3002-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nee DE, Wager TD, Jonides J. Interference resolution: insights from a meta-analysis of neuroimaging tasks. Cogn Affect Behav Neurosci. 2007;7:1–17. doi: 10.3758/cabn.7.1.1. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Gross JJ. The cognitive control of emotion. Trends Cogn Sci. 2005;9:242–249. doi: 10.1016/j.tics.2005.03.010. [DOI] [PubMed] [Google Scholar]
- Ochsner KN, Hughes B, Robertson ER, Cooper JC, Gabrieli JD. Neural systems supporting the control of affective and cognitive conflicts. J Cogn Neurosci. 2009;21:1842–1855. doi: 10.1162/jocn.2009.21129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olesen PJ, Westerberg H, Klingberg T. Increased prefrontal and parietal activity after training of working memory. Nat Neurosci. 2003;7:75–79. doi: 10.1038/nn1165. [DOI] [PubMed] [Google Scholar]
- Olvet DM, Hajcak G. The stability of error-related brain activity with increasing trials. Psychophysiology. 2009;46:957–961. doi: 10.1111/j.1469-8986.2009.00848.x. [DOI] [PubMed] [Google Scholar]
- Owen AM, Hampshire A, Grahn JA, Stenton R, Dajani S, Burns AS, et al. Putting brain training to the test. Nature. 2010;465:775–778. doi: 10.1038/nature09042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pizzagalli DA, Peccoralo LA, Davidson RJ, Cohen JD. Resting anterior cingulate activity and abnormal responses to errors in subjects with elevated depressive symptoms: a 128-channel EEG study. Hum Brain Mapp. 2006;27:185–201. doi: 10.1002/hbm.20172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poldrack RA. The neural correlates of motor skill automaticity. J Neurosci. 2005;25:5356–5364. doi: 10.1523/JNEUROSCI.3880-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Posner MI, Rothbart MK. Developing mechanisms of self-regulation. Dev Psychopathol. 2000;12:427–441. doi: 10.1017/s0954579400003096. [DOI] [PubMed] [Google Scholar]
- Reichenberg A, Harvey PD. Neuropsychological impairments in schizophrenia: integration of performance-based and brain imaging findings. Psychol Bull. 2007;133:833–858. doi: 10.1037/0033-2909.133.5.833. [DOI] [PubMed] [Google Scholar]
- Reithler J, van Mier HI, Goebel R. Continuous motor sequence learning: cortical efficiency gains accompanied by striatal functional reorganization. Neuroimage. 2010;52:263–276. doi: 10.1016/j.neuroimage.2010.03.073. [DOI] [PubMed] [Google Scholar]
- Rueda MR, Rothbart MK, Saccamanno L, Posner MI. Training, maturation and genetic influences on the development of executive attention. Proc Natl Acad Sci U S A. 2005;102:14931–14936. doi: 10.1073/pnas.0506897102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sayala S, Sala JB, Courtney SM. Increased neural efficiency with repeated performance of a working memory task is information-type dependent. Cereb Cortex. 2006;16:609–617. doi: 10.1093/cercor/bhj007. [DOI] [PubMed] [Google Scholar]
- Schafer JL, Olsen MK. Multiple imputation for multivariate missing-data problems: a data analyst’s perspective. Multiv Behav Res. 1998;33:545–571. doi: 10.1207/s15327906mbr3304_5. [DOI] [PubMed] [Google Scholar]
- Siegle GJ, Ghinassi F, Thase ME. Neurobehavioral therapies in the 21st century: summary of an emerging field and an extended example of cognitive control training for depression. Cogn Ther Res. 2007;31:235–262. [Google Scholar]
- Silton RL, Heller W, Towers DN, Engels AS, Spielberg JM, Edgar JC, et al. The time course of activity in dorsolateral prefrontal cortex and anterior cingulate cortex during top-down attentional control. Neuroimage. 2010;50:1292–1302. doi: 10.1016/j.neuroimage.2009.12.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorell LB, Lindqvist S, Bergman Nutley S, Bohlin G, Klingberg T. Training and transfer effects of executive functions in preschool children. Dev Sci. 2009;12:106–113. doi: 10.1111/j.1467-7687.2008.00745.x. [DOI] [PubMed] [Google Scholar]
- Tottenham N, Tanaka JW, Leon AC, McCarry T, Nurse M, Hare TA, et al. The NimStim set of facial expressions: judgments from untrained research participants. Psychiatry Res. 2009;168:242–249. doi: 10.1016/j.psychres.2008.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Veen V, Carter CS. The timing of action-monitoring processes in the anterior cingulate cortex. J Cogn Neurosci. 2002;14:593–602. doi: 10.1162/08989290260045837. [DOI] [PubMed] [Google Scholar]
- Westerberg H, Klingberg T. Changes in cortical activity after training of working memory—a single-subject analysis. Physiol Behav. 2007;92:186–192. doi: 10.1016/j.physbeh.2007.05.041. [DOI] [PubMed] [Google Scholar]
- Wu T. How self-initiated memorized movements become automatic: a functional MRI study. J Neurophysiol. 2004;91:1690–1698. doi: 10.1152/jn.01052.2003. [DOI] [PubMed] [Google Scholar]
- Yeung N, Cohen JD. The impact of cognitive deficits on conflict monitoring. Psychol Sci. 2006;17:164–171. doi: 10.1111/j.1467-9280.2006.01680.x. [DOI] [PubMed] [Google Scholar]
- Yeung N, Botvinick MM, Cohen JD. The neural basis of error detection: conflict monitoring and the error-related negativity. Psychol Rev. 2004;111:931–959. doi: 10.1037/0033-295x.111.4.939. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.