

Abstract
The protein leptin, a pleiotropic hormone regulates appetite and energy balance of the body and plays important roles in controlling linear growth, pubertal development, cardiovascular function, and immunity. Recent findings in the understanding of the structure, functional roles, and clinical significance of conditions with increased and decreased leptin secretion are summarized. Balance between leptin and other hormones is significantly regulated by nutritional status. This balance influences many organ systems, including the brain, liver, and skeletal muscle, to mediate the essential adaptation process. The aim of this review is to summarize the possible physiological functions of leptin and its signaling pathways during childhood and adolescence including control of food intake, energy regulation, growth and puberty, and immunity. Moreover, its secretion and possible roles in the adaptation process during different disease states (obesity, malnutrition, eating disorders, delayed puberty, congenital heart diseases and hepatic disorders) are discussed. The clinical manifestations and the successful management of patients with genetic leptin deficiency and the application of leptin therapy in other diseases including lipodystrophy, states with severe insulin resistance, and diabetes mellitus are discussed.
Keywords: Energy metabolism, growth, immunity, leptin, malnutrition, obesity, overfeeding
INTRODUCTION
Leptin: structure and synthesis
Leptin, the product of the ob gene, is a single-chain proteohormone produced by adipose tissue, but also by placenta (syncytiotrophoblasts), ovaries, skeletal muscle, stomach, mammary epithelial cells, bone marrow, pituitary, and liver, with multiple functions through various receptors located centrally and peripherally.[1,2] Centrally, leptin acts particularly on the hypothalamus to suppress food intake and stimulate energy expenditure.[3] Leptin receptors belong to the cytokine class I receptor family[4] and are found all over the body,[5] indicating a general role of leptin. A circulating form of the leptin receptor exists, which acts as one of several leptin-binding proteins.[6] Several alternatively spliced isoforms of the leptin receptor have been identified (Ob-Ra, Ob-Rb, Ob-Rc, Ob-Rd, and Ob-Re).[7,8]
Functions and control of secretion
Leptin is an important member of several keys that controls food intake and energy metabolism. The feeling of satiety and regulation of food intake is a complex system including several orexigenic and anorexigenic factors that act on the central nervous system and interplay with leptin. Many of them originate from the gastrointestinal track such as cholecystokinin, GLP-1, PYY, ghrelin, insulin, leptin, and adiponectin. In the adipose tissue when secretion of leptin is stimulated, it takes at least 60 min for the cells to respond by increasing levels of leptin synthesis and discharge. Adipocytes also synthesize several of the leptin receptors, the membrane-bound as well as the soluble ones and leptin is always secreted complexed to its receptor. This complexed form increases its half life in circulation. The gastric mucosa contributes for a large part to the levels of circulating leptin, particularly at time of food intake. Gastric leptin secretion is faster when compared to adipocytes secretion. Thus, gastric leptin acts rapidly and transiently to food intake to trigger in the short term the feeling of satiety, whereas adipose tissue leptin may in turn act on the long-term regulating energy expenditure. In addition, gastric leptin exerts a regulatory function on the digestive tract. Intestinal epithelial cells express leptin receptors on their luminal brush border and baso-lateral membranes. Thus, leptin either originating from the gastric juice or from the blood circulation can act on the intestinal cells to regulate digestion of nutrients.[9,10]
Under conditions of weight maintenance, the most important variables that determines circulating leptin concentrations is the body fat mass and body mass index (BMI).[11,12] However, in conditions of negative (weight-loss) and positive (weight-gain) energy balances, the changes in leptin concentrations function as a sensor of energy imbalance. Within 24 h of fasting, leptin concentrations decrease to ~30% of initial basal values. However, massive overfeeding over a 12-h period increases leptin concentrations by ~50% of initial basal values. A few studies have shown a modest increase in leptin secretion at supraphysiologic insulin concentrations 4-6 h after insulin infusion.[13,14]
It appears that leptin acts as a “satiety signal” to prevent obesity in times of energy excess, and as a “starvation signal” to maintain adequate fat stores for survival during times of energy deficit. Data suggest that the leptin system may be more efficient in signaling a decrease in fat mass and lack of nutrients (low leptin state) and triggering a compensatory increase in food intake and a decrease in energy expenditure than as a satiety signal when its serum levels are elevated. In addition, recent evidence suggests that the neurobiology of leptin signaling in obesity appears to involve central leptin insufficiency, as opposed to the previously postulated notion of leptin resistance.[15–19]
The arcuate nucleus (ARC), ventromedial (VMH), dorsomedial (DMH), and lateral (LH) hypothalamic nuclei are important regions regulating food intake and energy expenditure. Disrupting lesions in the ARC, VMH, and DMH of rats resulted in hyperphagia and obesity. Besides, lesions in the LH result in decreased food intake. Binding of leptin to its hypothalamic receptors activates a signaling cascade in the ARC that results in inhibition of orexigenic pathways as indicated by decreased mRNA expression of neuropeptide Y (NPY) and agouti-related peptide (AgRP), and stimulation of anorexigenic pathways as suggested by increases in the mRNA levels of alpha-melanocyte-stimulating hormone (α-MSH) and cocaine and amphetamine regulated transcript (CART). Activation of POMC/CART-expressing neurons by leptin results in release of α-MSH, which subsequently binds to melanocotin receptors (MCRs) and leads to anorexia and increased energy expenditure. At the same time, leptin inhibits NPY/AgRP neurons, which stimulate orexigenic responses and directly inhibits POMC neuron expression as indicated by POMC mRNA expression.[16–19]
Recent data in mice show that leptin acts solely in the hypothalamus to control glucose homeostasis independent of its effects on appetite, by increasing non-thermogenic energy expenditure and glucose disposal in peripheral tissues by action on POMC neurons.[20]
In humans, decreasing leptin concentrations in response to food deprivation are responsible for the starvation-induced suppression of the hypothalamic-pituitary-gonadal axes as well as the malfunction of several other neuroendocrine axes.[19] It seems that leptin may act as the link between adipose tissue, hypothalamic centers regulating energy homeostasis, and the reproductive system.
The human placenta express the leptin and leptin receptor gene implying that it is a site production of the hormone as well as a target of its action (autocrine effect). During pregnancy, leptin production by the fetus and the placenta can signal fetal nutrient status. It may also provide a mechanism whereby maternal fuel reserves are more readily mobilized, favoring utilization by the fetus rather than building maternal reserves. In the case where maternal leptin concentrations are low owing to limiting nutrition, the proportional importance of fetal and placental leptin in mobilizing fuel for use by the fetus may be greater.[21] Neonatal leptin levels are higher in females and are associated with adiposity, and are independently correlationed with newborn length, IGF-I levels and formula feeding. Leptin clearly plays an important role in neonatal energy homeostasis and metabolism.[22]
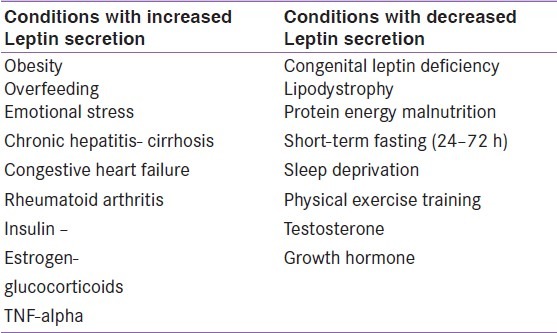
Synthesis of leptin is also modulated by several hormonal variables. Stimulators include insulin, catecholamines, and glucocorticoids.[23] Studies demonstrated that meals and insulin acutely affect leptin concentrations.[24] Suppressors include fasting, cAMP, and β3-adrenoreceptor agonists.[25] It has been demonstrated that leptin production occurs after increases in insulin in response to feeding, and a decrease in leptin concentrations follows decreases in insulin during fasting [Table 1].
Table 1.
Conditions with Changes of Serum Leptin Concentrations
Physiological and developmental changes in leptin during life
Serum leptin concentrations show a moderate circadian variation with a peak during the night at ~0200(~30-100%) higher than the concentrations measured in the morning or early afternoon.[26] This variation, together with the influence of food intake, needs to be taken into account when blood samples are collected. Because body fat mass is the major confounding variable, these ranges should be referred to measures of the percentage body fat such as BMI.[27]
Leptin levels exhibit significant changes during progressive pubertal stages, with a distinct dimorphism between boys and girls. In boys, there is a pre-pubertal peak of serum leptin levels preceding the rise of free testosterone, growth hormone (GH), and insulin-like growth factor (IGF-1). Thereafter, about 3 years after the rise in serum testosterone levels, leptin levels fall to baseline concentrations.[28] Girls have higher serum leptin levels than males and their leptin levels rise throughout puberty, concomitant with the rise in estrogen levels.[29,30] This sexual dimorphism is related higher pulse amplitude of leptin secretion from adipose tissue in females, higher subcutaneous/visceral fat ratio in and lower leptin-binding protein levels and higher free leptin levels in females.[31–34] Reference intervals referring to measures of body fat should therefore be stratified according to gender and pubertal development.[34,35]
Clinical significance of leptin in Obesity, Linear Growth, and Insulin Resistance Syndrome
In the majority of obese individuals, serum leptin concentrations are increased,[36] and leptin administration shows only very limited effects[37] due to leptin resistance. Low soluble receptor concentrations and a high fraction of free to bound leptin are markers of leptin resistance. Leptin resistance is associated with insulin resistance and abdominal obesity and constitutes an additional component of the metabolic syndrome.[38] When free-leptin concentrations are compared between lean and obese individuals, even more pronounced hyperleptinemia in obesity is observed than that reported by measuring total leptin concentrations. Recent evidence suggests that the neurobiology of leptin signaling in obesity appears to involve central leptin insufficiency, as opposed to the previously postulated notion of leptin resistance.[15–19]
Apart from leptin's mandatory regulatory role in energy intake and expenditure, independent participation of leptin in the hypothalamic integration of insulin–glucose homeostasis has been documented. Various lines of evidence show that (a) under the direction of leptin two independent relays emanating from the hypothalamus restrain insulin secretion from the pancreas and mobilize peripheral organs---liver, skeletal muscle and brown adipose tissue---to upregulate glucose disposal, and (b) leptin insufficiency in the hypothalamus produced by either leptinopenia or restriction of leptin transport across the blood brain barrier due to hyperleptinemia of obesity initiates antecedent pathophysiological sequalae of diabetes type 1 and 2. The efficacy and preclinical safety of leptin replenishment in vivo, especially by supplying it to the hypothalamus with the aid of gene therapy, in preventing the antecedent pathophysiological sequalae---hyperinsulinemia, insulin resistance and hyperglycemia---in various animal models and clinical paradigms of diabetes type 1 and 2 with or without attendant obesity have been documented[15,39] [Figure 1].
Figure 1.
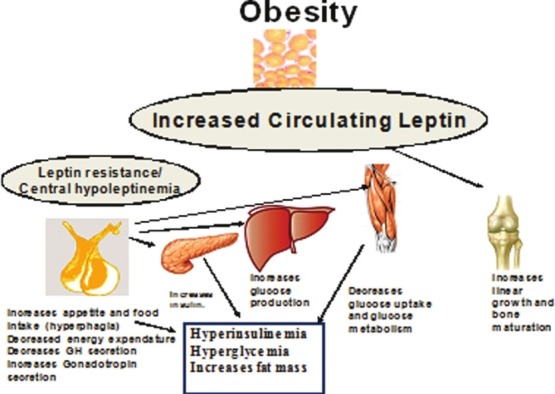
Leptin effects during obesity Central leptin insufficiency in obesity for extended periods of time results in increased fat accrual, decreased energy expenditure, hyperinsulinemia, hyperglycemia. Hyperleptinemia may increase bone growth and accelerate maturation
In support of this view, in leptin-deficient (ob/ob) mice, leptin injections led to dose-dependent reductions in serum glucose levels compared to fed ob/ob controls, before any significant change in body weight occurred.[15,39] Approximately 3-5% of human obese populations can be regarded as “relatively” leptin deficient and could benefit from leptin therapy. Individuals with heterozygous leptin gene mutations have low circulating concentrations of leptin and increased body adiposity.[40]
During the phase of normal or accelerated height velocity in obese children, plasma GH levels remain low and measurements performed during sleep and following pharmacological stimulation as well as spontaneous 24 h of GH secretion are reduced.[41,42] Despite the reduction in GH levels, obese children may have normal, increased or reduced, plasma IGF-I and GH-binding protein levels.[43] The mechanism whereby obese children continue to grow despite the low levels of GH is not known. Several explanations have been postulated.[43–46] Maor et al.[47] reported the presence of leptin receptors in growth plates. Leptin induces both proliferation and differentiation of chondrocytes. It stimulates the width of the proliferative zone of the epiphysial growth plate and increases the expression of chondroitin sulfate within the cartilaginous matrix. Apparently, leptin acts as a direct skeletal growth factor in obese children. It is suggested that in humans, obesity is associated with differential sensitivity to circulating leptin with central resistance and peripheral sensitivity (epiphysial growth plate) to the effect of leptin.
Both leptin and insulin resistance are strongly related to adiposity and other cardiovascular risk factors.[48–50] Studying these relations in childhood may help clarify some aspects in the development of the insulin resistance syndrome. However, conflicting results are published about the dependent versus independent associations between body fatness, leptin levels, and insulin resistance in obese children.[51–53] A clearer understanding of the leptin-obesity–insulin resistance relationship is still required during childhood and adolescence relative to the development of risk factors and type 2 diabetes.
Leptin, Malnutrition, and Adaptation to Prolonged Nutritional Deficiency
In two studies on infants and children with mild and severe forms of protein energy malnutrition (PEM), leptin concentrations are significantly decreased and positively correlated with triceps, scapular, and abdominal fat thickness.[54,55] In severe PEM cases, concentrations of IGF-I are significantly low, whereas basal cortisol and GH concentrations are significantly high versus normal children. The BMI is correlated significantly with leptin, insulin and IGF-I. These findings suggest that during prolonged nutritional deprivation, the decreased energy intake, diminished fat mass, and declining insulin (and possibly IGF-I) concentrations suppress leptin production.
Leptin appears to be an important signal in the process of metabolic/endocrine adaptation to prolonged nutritional deprivation. Low leptin levels decrease leptin inhibition on NPY that affects the regulation of pituitary growth and pituitary adrenal axes. Stimulation of the hypothalamic-pituitary-adrenal (HPA) axis and possibly the hypothalamic-pituitary-GH axis to maintain the high cortisol and GH levels necessary for effective gluconeogenesis and lipolysis to ensure a fuel (glucose and fatty acids) supply for the metabolism of brain and peripheral tissue during nutritional deprivation. The decreased synthesis of IGF-I and the low level of insulin and/or its diminished effect due to an insulin-resistant status in the presence of high circulating GH and cortisol levels ensure substrate diversion away from growth toward metabolic homeostasis[5] [Figure 2].
Figure 2.
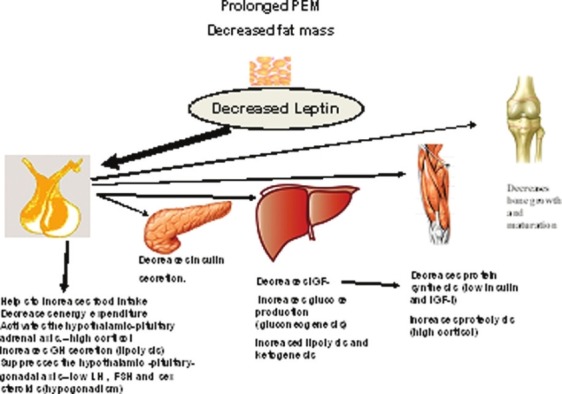
Adaptation to prolonged protein energy malnutrition (PEM). During prolonged malnutrition, low leptin levels, decreases leptin inhibition on NPY that affects the regulation of pituitary growth and pituitary adrenal axes and insulin/glucose homeostasis. Stimulation of the hypothalamicpituitary- adrenal (HPA) axis and possibly the hypothalamic-pituitary-GH axis occurs to maintain the high cortisol and GH levels necessary for effective gluconeogenesis and lipolysis to ensure a fuel (glucose and fatty acids) supply for the metabolism of brain and peripheral tissue during nutritional deprivation. The decreased synthesis of IGF-I and the low level of insulin and/or its diminished effect due to an insulin-resistant status in the presence of high circulating GH and cortisol levels ensure substrate diversion away from growth toward metabolic homeostasis
On the other hand, during recovery from malnutrition, leptin concentrations increase in relation to fat mass. During recovery from severe PEM, an increase in leptin concentration was observed only in children who showed catch-up growth. More interestingly, malnourished children with catch-up growth had higher serum leptin concentrations compared to healthy children. This suggested that leptin affects nutritional status during catch-up growth as a dynamic process, rather than merely being an index of body fat content.[55]
Leptin and eating disorders
In normal-weight subjects with Bulemia nervosa (BN), circulating leptin has been reported to be either decreased, normal, or increased. The reasons for such a discrepancy among the studies may be primarily due to the heterogeneous composition of the patient samples, while patients with anorexia nervosa (AN) have almost uniformly low plasma leptin concentrations, bulimic women exhibit variable levels of plasma leptin with increased, normal, or decreased values are reported. In AN, the level of leptin correlates significantly and positively with the BMI and fat mass of patients. However, although leptin changes in eating disorder patients likely reflect adaptive mechanisms to chronic malnutrition and starvation, it is plausible that, when instated, sustain certain clinical symptoms, since leptin unequivocally acts not only as a hunger suppressant factor but also as a modulator of reproduction, physical activity, immune system and other endocrine functions. Therefore, changes in leptin production, although not directly involved in the etiology of AN and BN, they may have an impact on the course and prognosis of these disorders.[56–59] One of the most exciting research area for the future will be to assess whether pharmacologic and/ or nutritional manipulations of leptin physiology may ameliorate the outcome of AN and BN.
Leptin, Puberty, and Constitutional delay of growth and puberty
Leptin receptors have been identified in the hypothalamus, the gonadotrope cells of the anterior pituitary, and gonads of higher primates and human.[60,61] Leptin administration increased serum concentrations of luteinizing hormone (LH) and growth hormone (GH) in the rodent. Intracerebroventricular (ICV) administration of leptin suppressed feed intake, increased serum GH concentrations, and stimulated hypothalamic gonadotropin releasing hormone (GnRH) release in the rat.[62]
In vitro, leptin has a direct stimulatory effect on the hypothalamic-pituitary-gonadal (HPG) axis by accelerating GnRH secretion through decreasing the expression of NPY in the arcuate hypothalamic neurons.[61–65] Leptin directly stimulates the release of LH and, to a lesser extent, FSH via nitric oxide synthase activation in the gonadotropes.[62–65] Leptin administration has been shown to decrease the expression of NPY in the arcuate nucleus and consequently to remove the inhibitory action of NPY on pulsatile GnRH release.[65] The endocrine and direct paracrine effects of leptin on the gonads are implied by the expression of functional leptin receptors on the surface of ovarian follicular cells, including granulose, theca, and interstitial cells,[64] as well as Leydig cells.
Humans and mice lacking leptin (ob/ob) or leptin receptor (LepR, db/db) are infertile. Leptin administration to leptin-deficient subjects and ob/ob mice induces puberty and restores fertility. Recently, it has been made clear that leptin acts indirectly on gonadotropin-releasing hormone (GnRH)-secreting cells via actions on interneurons.[65–69] Data from two longitudinal cohorts suggest a role for leptin in the normal regulation of childhood weight gain, maturation, and the development of secondary sexual features and body composition.[67]
Serum leptin concentrations are higher in early adolescence compared with childhood and may play a facilitator role in pubertal development. Constitutional delay in growth and puberty (CDGP) is a disorder of the tempo of physical maturation and may be associated with relative hypoleptinemia. In adolescent boys, after controlling for pubertal status, age and BMI, leptin concentrations are higher in early puberty compared with pre-puberty in normal boys but are not significantly elevated in pubertal boys with CDGP. In boys with CDGP, leptin SDS is negatively correlated with both chronological age and bone age, indicating that older boys with CDGP had lower leptin levels than expected given their age and BMI-SDS. Therefore, absence of physiological increases of leptin concentrations may be associated with delayed puberty in boys.[70]
Leptin and bone growth
Leptin possibly signals to bone on multiple levels, with local and systemic effects. Leptin after crossing the blood-brain barrier and binding to its receptors in the brain can act through the sympathetic nervous system to regulate bone metabolism. It is also possible that, in addition to its effects through the brain, leptin may act directly on cells in the bone to regulate bone metabolism. Leptin acts to reduce cancellous bone but conversely increases cortical bone. A recent theory suggests that increased leptin during obesity may represent a mechanism for enlarging cortical bone size to cope with increased body weight, and decreased leptin during malnutrition may contribute to decreased bone growth.[71–73]
Leptin in thyroid diseases and during glucocorticoid excess
In hypothyroid patients treated with L-T4 and hyperthyroid patients with methimazole, no correlation has been detected between serum leptin and thyroid hormone (free thyroxine, thyrotrophin and T3) levels at any stage.[74,75]
In humans, glucocorticoids stimulate leptin gene expression and secretion independently of effects on food intake.[76,77] Leptin infusion (0.5 mg/h) significantly decreased food intake and body weight in adrenalectomized mice.[78] Glucocorticoid replacement increased food intake without reversing leptin inhibition of hypothalamic neuropeptide Y (NPY) mRNA levels. These investigators indicate that glucocorticoid levels within the physiological range interfere with leptin action and that glucocorticoid effects are at least partly independent of NPY. In obese adults and children, dexamethasone administration (10 mg/day) caused increases in leptin levels after 5 days of treatment.[76,79] The mechanism(s) of glucocorticoid stimulation of plasma leptin is still unknown.
Leptin and immunity
The overall leptin action in the immune system is a proinflammatory effect, activating proinflammatory cells, promoting T-helper 1 responses, and mediating the production of the other proinflammatory cytokines. Leptin receptor is also upregulated by proinflammatory signals. Human leptin was found to stimulate proliferation and activation of human circulating monocytes in vitro, promoting the expression of activation markers: CD69, CD25, CD38, and CD71, in addition to increasing the expression of monocytes surface markers, such as HLA- DR, CD11b, and CD11c.[80] Leptin dose dependently stimulates the production of proinflammatory cytokines by monocytes, that is, TNF-α and IL-6.[81] Moreover, leptin promotes neutrophils chemotaxis and has a stimulating effect on intracellular hydrogen peroxide production in neutrophils and monocytes.[82,83] It enhances both the development and the activation of NK cells, increasing IL-12 and reducing the expression of IL-15. Besides, leptin mediates the activation of NK cells indirectly by modulation of IL-1β, IL-6, and TNF-α by monocytes and macrophages.[84] On the other hand, leptin promotes T-cell survival[85] by modulating the expression of antiapoptotic proteins, such as Bcl-xL in stress-induced apoptosis.[86]
In human obesity due to congenital leptin deficiency, there is a T-cell hypo-responsiveness and not only leptin treatment in these patients is effective lowering body weight but it can also revert T-cell response to mitogen activation in vitro.[87] Acute starvation reduces serum leptin levels, resulting in an impaired cellular immune response. The pro-inflammatory immune response, mediated by the low leptin levels, is also impaired during starvation.
Leptin has also been shown to enhance immune reactions in autoimmune diseases that are commonly associated with inflammatory responses. Recent evidence indicates that increased leptin is involved in the dysregulated balance between Th1 and Th2 cytokines and contributes to the pathogenesis of rheumatoid arthritis (RA).[88] In contrast, leptin deficiency has a protective effect on autoimmune diseases by altering the balance of Th1:Th2 cytokine production and promoting a Th2 response. This is shown in fasting patients with RA exhibiting significantly improved clinical disease activity correlated with a marked reduction in serum leptin and a shift toward Th2 cytokine production.[89] The application of leptin antagonist therapy for prevention and treatment of immunity-related disorders is in progress.[88]
Leptin in chronic liver diseases: Possible carcinogenesis potential
The data on leptin in human studies show elevated peripheral leptin with hepatitis, liver cirrhosis and fibrosis. In patients with chronic hepatitis C, the severity of liver fibrosis is associated with higher serum leptin.[90,38]
Angiogenesis has been shown to play an important role in the progression of chronic liver disease. Animal studies illustrate that leptin-mediated neovascularization in the liver, which takes place only in the presence of vascular endothelial growth factor (VEGF), significantly increases in parallel with the progression of non-alcoholic steatohepatitis (NASH). These results suggest that leptin-mediated neovascularization, which coordinates with VEGF, plays a pivotal role in the progression of liver fibrosis and hepatocellular carcinoma (HCC) in NASH.[91,92] The carcinogenic potential of leptin in the development prostate and breast cancer is not in the scope of this review.
Leptin and cardiovascular system
Leptin possesses potent vascular effects and participates in the regulation of sympathetic tone and arterial blood pressure through central as well as peripheral actions.[93]
Intra-cerebroventricular administration of leptin elicits an increase in arterial blood pressure consistent with its sympatho-excitatory property[94] attributed to a central mechanism.[95] Locally, leptin is capable of regulating cardiac and vascular contractility through a local nitric oxide-dependent mechanism. These findings have prompted the speculation that leptin in the physiological range may serve as a physiological regulator of cardiovascular function whereas elevated plasma leptin levels due to tissue leptin resistance may act as a patho-physiological trigger and/or marker for cardiovascular diseases.[90,96,97]
Significantly, higher serum levels of leptin are detected in children with congenital heart disease (CHD), particularly in patients with cyanotic CHD. CHD patients with SpO2<90%, pulmonary hypertension (PH), severe pulmonary stenosis (PS), detectable collaterals, cardiomegaly and/or heart failure showed significantly higher levels of leptin than those with higher SpO2 or those without these findings. Elevated plasma leptin levels and its soluble receptor in these patients with CHD suggest that leptin may participate in the catabolic cardiac cachexia and failure to thrive in these patients.[90] It is possible that this catabolic state, affecting body fat and muscle, may be useful to secure calories to spare the mechanically compromised heart. Moreover, elevated leptin may modify the process of angiogenesis that could be essential to enhance renal perfusion in some cases of CHD.[98]
Congenital leptin deficiency (homozygous for the Δ133G mutation)
The absence of serum leptin is caused by homozygous frameshift or missense mutations in the ob gene (7q31.3) and is inherited as an autosomal recessive trait. However, a similar phenotype has been observed in some individuals from one consanguineous family who carried homozygous mutations in the gene encoding the leptin receptor (LEPR, 1p31).[99–101] Congenital leptin deficiency is a form of monogenic obesity characterized by severe early-onset obesity and marked hyperphagia. Patients with congenital leptin deficiency are severely hyperphagic from early infancy and, although birth weight is normal, they rapidly become obese during early childhood. An increased susceptibility to infections has also been reported in these infants and appears to be associated with reduced numbers of circulating CD4+ T cells, and impaired T cell proliferation and cytokine release. Other features of the disorder include hyper-insulinaemia, advanced bone age, hypothalamic hypothyroidism and hypogonadotropic hypogonadism leading to a failure to undergo puberty.
Congenital leptin deficiency can be successfully treated with daily subcutaneous injections of recombinant human leptin. Leptin replacement therapy is undertaken at low physiological doses, starting at 0.02-0.04 mg/kg/day given subcutaneously at 6 pm. The child's dose is adjusted to increase the peak serum leptin to 70 ng/mL. Treatment results in sustained weight loss, and reduces appetite, hyper-insulinaemia, and hyper-lipidemia, and attains appropriate pubertal development. In addition, leptin administration corrects abnormal thyroid biochemistry and allows the withdrawal of T4 treatment. The white blood cell count (lymphocytes, neutrophils, and monocytes) all increase and remain elevated for the first 3 months. There is no evidence of concomitant infection at the time of therapy.[102,103]
Leptin and lipodystrophy
Generalized lipodystrophy is a rare disorder characterized by a profound deficiency of adipose tissue with resultant loss of triglyceride storage capacity and reduced adipokines, including leptin. Subjects with generalized lipodystrophy and reduced leptin levels often have an increased appetite leading to hyperphagia. Excess fuel consumption, coupled with a lack of adipose tissue, contributes to the significant ectopic triglyceride accumulation in the muscle and liver seen in these subjects. Typical daily replacement doses for metreleptin of 0.06 to 0.08 mg/kg for female patients and 0.04 mg/kg for male patients are administered by subcutaneous injection twice daily. Leptin replacement in these subjects treated for 12 months showed significant reductions in serum fasting glucose, HbA1c, triglycerides, and LDL. Liver volumes were significantly reduced (loss of steatosis) with decreases in total body weight and resting energy. Therefore, leptin represents the first novel, effective, long-term treatment for severe forms of lipodystrophy.[104,105]
Potential therapeutic uses of leptin
Apart from the successful use of leptin therapy in congenital leptin deficiency and generalized forms of lipodystrophy, animal research suggested potential beneficial effect of leptin in 1 diabetes mellitus. In non-obese diabetic mice with uncontrolled type 1 diabetes, leptin therapy alone or combined with low-dose insulin reverses the catabolic state through suppression of hyper-glucagonemia. Additionally, leptin therapy normalizes the levels of a wide array of hepatic intermediary metabolites in multiple chemical classes, including acylcarnitines, organic acids (tricarboxylic acid cycle intermediates), amino acids, and acyl CoAs. In contrast to insulin monotherapy, however, leptin lowers both lipogenic and cholesterologenic transcription factors and enzymes and reduces plasma and tissue lipids. The results imply that leptin administration may have multiple short- and long-term advantages over insulin monotherapy for type 1-diabetes.[106] Leptin administration in rats, which develop adult-onset polygenic obesity and type 2 diabetes, normalized fasting plasma glucose and was accompanied by lowered HbA1c, plasma glucagon, and triglyceride concentrations and expression of hepatic gluconeogenic enzymes.[107] Additionally, supplying leptin to the hypothalamus with the aid of gene therapy successfully prevents the antecedent pathophysiological sequalae--hyperinsulinemia, insulin resistance and hyperglycemia, and type 2 diabetes mellitus in various animal models.[39]
In human, however, short-term treatment with either low-dose or high-dose r-Met hu leptin does not improve liver, skeletal muscle, or adipose tissue insulin sensitivity in weight stable, obese subjects with type 2 diabetes. This suggests that increasing circulating leptin above normal plasma concentrations does not have significant effect, because it does not correct the central hypoleptinemia.[15–19,108]
Leptin levels have been found to be low in subjects with mutations in the insulin receptor, despite having the most severe degree of insulin resistance (e.g. Donohue syndrome, Rabson Mendenhall syndrome, and type-A insulin resistance). These findings propose that leptin therapy may be potentially useful in these patients. Up till now, no therapeutic trial has been published in these syndromes.[109]
SUMMARY
Leptin is an adipocyte-secreted hormone that regulates appetite and energy balance of the body and has an important role in adaptation during underfeeding and overfeeding. It takes a significant task in controlling hypothalamic-pituitary growth, adrenal and gonadal functions. In humans, its roles in controlling linear growth, bone metabolism, pubertal development, cardiovascular function, and immunity are still evolving. During malnutrition (underfeeding), eating disorders and other chronic diseases (congenital heart diseases, hepatic diseases and autoimmune disorders), it seems that it controls adaptation to spare essential functions and systems. It is used successfully to treat obese children with congenital leptin deficiency and those with insulin resistance and generalized lipodystrophy. Leptin ameliorates obesity, insulin resistance, hyperglycemia, hyperinsulinemia, dyslipidemia, hepatic steatosis and restores gonadotropin secretion.
In mice, gene therapy to increase hypothalamic leptin proves successful in preventing and treating diabetes and represents a potential therapy for human.[110]
Footnotes
Source of Support: Nil
Conflict of Interest: None declared
REFERENCES
- 1.Baratta M. Leptin—from a signal of adiposity to a hormone mediator in peripheral tissues. Med Sci Monit. 2002;8:RA282–92. [PubMed] [Google Scholar]
- 2.Friedman JM, Halaas JL. Leptin and the regulation of body weight in mammals. Nature. 1998;22:763–70. doi: 10.1038/27376. [DOI] [PubMed] [Google Scholar]
- 3.Webber J. Energy balance in obesity. Proc Nutr Soc. 2003;62:539–43. doi: 10.1079/pns2003256. [DOI] [PubMed] [Google Scholar]
- 4.Tsiotra PC, Pappa V, Raphis SA, Tsigos C. Expression of the long and short leptin receptor isoforms in peripheral blood mononuclear cells: Implications for leptin's actions. Metabolism. 2000;49:1537–41. doi: 10.1053/meta.2000.18519. [DOI] [PubMed] [Google Scholar]
- 5.Trayhurn P, Hoggard N, Mercer JG, Rayner DV. Leptin: Fundamental aspects. Int J Obes Relat Metab Disord. 1999;Suppl 1:S22–8. doi: 10.1038/sj.ijo.0800791. [DOI] [PubMed] [Google Scholar]
- 6.Lammert A, Kiess W, Bottner A, Glasow A, Kratzsch J. Soluble leptin receptor represents the main leptin binding activity in human blood. Biochem Biophys Res Commun. 2001;283:982–8. doi: 10.1006/bbrc.2001.4885. [DOI] [PubMed] [Google Scholar]
- 7.Wallace AM. Measurement of leptin and leptin binding in the human circulation. Ann Clin Biochem. 2000;37:244–52. doi: 10.1258/0004563001899311. [DOI] [PubMed] [Google Scholar]
- 8.Funahashi H, Yada T, Suzuki R, Shioda S. Distribution, function, and properties of leptin receptors in the brain. Int Rev Cytol. 2003;224:1–27. doi: 10.1016/s0074-7696(05)24001-9. [DOI] [PubMed] [Google Scholar]
- 9.Oswal A, Yeo G. Leptin and the Control of Body Weight: A Review of Its Diverse Central Targets, Signaling Mechanisms, and Role in the Pathogenesis of Obesity. Obesity (Silver spring) 2010;18:221–9. doi: 10.1038/oby.2009.228. [DOI] [PubMed] [Google Scholar]
- 10.Cammisotto P, Bendayan M. A review on gastric leptin: The exocrine secretion of a gastric hormone. Anat Cell Biol. 2012;45:1–15. doi: 10.5115/acb.2012.45.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Margetic S, Gazzola C, Pegg GG, Hill RA. Leptin: A review of its peripheral actions and interactions. Int J Obes Relat Metab Disord. 2002;46:1407–33. doi: 10.1038/sj.ijo.0802142. [DOI] [PubMed] [Google Scholar]
- 12.Frayn KN, Karpe F, Fielding BA, Macdonald IA, Coppack SW. Integrative physiology of human adipose tissue. Int J Obes Relat Metab Disord. 2003;27:875–88. doi: 10.1038/sj.ijo.0802326. [DOI] [PubMed] [Google Scholar]
- 13.Radic R, Nikolic V, Karner I, Kosovic P, Kurbel S, Selthofer R, et al. Circadian rhythm of blood leptin in obese and non-obese people. Coll Antropol. 2003;27:555–61. [PubMed] [Google Scholar]
- 14.Wong ML, Licinio J, Yildiz BO, Mantzoros CS, Prolo P, Kling M, et al. Simultaneous and continuous 24-hour plasma and cerebrospinal fluid leptin measurements: Dissociation of concentrations in central and peripheral compartments. J Clin Endocrinol Metab. 2004;89:258–65. doi: 10.1210/jc.2003-031275. [DOI] [PubMed] [Google Scholar]
- 15.Engineer DR, Garcia JM. Leptin in anorexia and cachexia syndrome. Int J Pept. 2012;2012:287457. doi: 10.1155/2012/287457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Simpson KA, Niamh M. Hypothalamic regulation of food intake and clinical therapeutic application. Arq Bras Endocrinol Metabol. 2009;53:120–8. doi: 10.1590/s0004-27302009000200002. [DOI] [PubMed] [Google Scholar]
- 17.Broberge C. Brain regulation of food intake and appetite: Molecules and networks. J Intern Med. 2005;258:301–27. doi: 10.1111/j.1365-2796.2005.01553.x. [DOI] [PubMed] [Google Scholar]
- 18.Horvath TL, Andrews ZB, Diano S. “Fuel utilization by hypothalamic neurons: Roles for ROS”. Trends Endocrinol Metab. 2009;20:78–87. doi: 10.1016/j.tem.2008.10.003. [DOI] [PubMed] [Google Scholar]
- 19.Sainsbury A, Cooney GJ, Herzog H. Hypothalamic regulation of energy homeostasis. Best Pract Res Clin Endocrinol Metab. 2002;16:623–7. doi: 10.1053/beem.2002.0230. [DOI] [PubMed] [Google Scholar]
- 20.Berglund ED, Vianna CR, Donato J, Jr, Kim MH, Chuang JC, Lee CE, et al. Direct leptin action on POMC neurons regulates glucose homeostasis and hepatic insulin sensitivity in mice. J Clin Invest. 2012;122:1000–9. doi: 10.1172/JCI59816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cameo P, Bischof P, Calvo JC. Effect of Leptin on Progesterone, Human Chorionic Gonadotropin, and Interleukin-6 Secretion by Human Term Trophoblast Cells in Culture. Biol Reprod. 2003;68:472–7. doi: 10.1095/biolreprod.102.006122. [DOI] [PubMed] [Google Scholar]
- 22.Petridou E, Mantzoros CS, Belechri M, Skalkidou A, Dessypris N, Papathoma E, et al. Neonatal leptin levels are strongly associated with female gender, birth length, IGF-I levels and formula feeding. Clin Endocrinol (Oxf) 2005;62:366–71. doi: 10.1111/j.1365-2265.2005.02225.x. [DOI] [PubMed] [Google Scholar]
- 23.Koutkia P, Canavan B, Johnson ML, DePaoli A, Grinspoon S. Characterization of leptin pulse dynamics and relationship to fat mass, growth hormone, cortisol, and insulin. Am J Physiol Endocrinol Metab. 2003;285:E372–9. doi: 10.1152/ajpendo.00097.2003. [DOI] [PubMed] [Google Scholar]
- 24.Lee MJ, Fried SK. Integration of hormonal and nutrient signals that regulate leptin synthesis and secretion. Am J Physiol Endocrinol Metab. 2009;296:E1230–8. doi: 10.1152/ajpendo.90927.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Slieker LJ, Sloop KW, Surface PL, Kriauciunas A, LaQuier F, Manetta J, et al. Regulation of expression of ob mRNA and protein by glucocorticoids and cAMP. J Biol Chem. 1996;271:5301–4. doi: 10.1074/jbc.271.10.5301. [DOI] [PubMed] [Google Scholar]
- 26.Sinha MK, Ohannesian JP, Heiman ML, Kriauciunas A, Stephens TW, Magosin S, et al. Nocturnal rise of leptin in lean, obese, and non-insulin dependent diabetes mellitus subjects. J Clin Invest. 1996;97:1344–7. doi: 10.1172/JCI118551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Haluzik M, Fiedler J, Nedvidkova J, Ceska R. Serum leptin levels in patients with hyperlipidemias. Nutrition. 2000;16:429–33. doi: 10.1016/s0899-9007(00)00288-4. [DOI] [PubMed] [Google Scholar]
- 28.Clayton PE, Gill MS, Hall CM, Tillmann V, Whatmore AJ, Price DA. Serum leptin through childhood and adolescence. Clin Endocrinol (Oxf) 1997;46:727–33. doi: 10.1046/j.1365-2265.1997.2081026.x. [DOI] [PubMed] [Google Scholar]
- 29.Saad MF, Damani S, Gingerich RL, Riad-Gabriel MG, Khan A, Boyadjian R, et al. Sexual dimorphism in plasma leptin concentration. J Clin Endocrinol Metab. 1997;82:579–84. doi: 10.1210/jcem.82.2.3739. [DOI] [PubMed] [Google Scholar]
- 30.Licinio J, Negrao AB, Mantzoros C, Kaklamani V, Wong ML, Bongiorno PB, et al. Sex differences in circulating human leptin pulse amplitude: Clinical implications. J Clin Endocrinol Metab. 1998;83:4140–7. doi: 10.1210/jcem.83.11.5291. [DOI] [PubMed] [Google Scholar]
- 31.Montague CT, Prins JB, Sanders L, Digby JE, O’Rahilly S. Depotand sex-specific differences in human leptin mRNA expression: Implications for the control of regional fat distribution. Diabetes. 1997;46:342–7. doi: 10.2337/diab.46.3.342. [DOI] [PubMed] [Google Scholar]
- 32.McConway MG, Johnson D, Kelly A, Griffin D, Smith J, Wallace AM. Difference in circulating concentrations of total, free and bound leptin relate to gender and body composition in adult humans. Ann Clin Biochem. 2000;37(Pt 5):717–23. doi: 10.1258/0004563001899771. [DOI] [PubMed] [Google Scholar]
- 33.Halleux CM, Servais I, Reul BA, Detry R, Brichard SM. Multihormonal control of ob gene expression and leptin secretion from cultured human visceral adipose tissue: Increased responsiveness to glucocorticoids in obesity. J Clin Endocrinol Metab. 1998;83:902–10. doi: 10.1210/jcem.83.3.4644. [DOI] [PubMed] [Google Scholar]
- 34.Blum WF, Juul A. Reference ranges of serum leptin. In: Blum WF, et al., editors. Leptin—The voice of adipose tissue. Verlag Heidelberg: Johann Ambrosius; 1997. pp. 318–26. [Google Scholar]
- 35.Li HJ, Ji CY, Wang W, Hu YH. A twin study for serum leptin, soluble leptin receptor, and free insulin-like growth factor-I in pubertal females. J Clin Endocrinol Metab. 2005;90:3659–64. doi: 10.1210/jc.2004-2079. [DOI] [PubMed] [Google Scholar]
- 36.Rosicka M, Krsek M, Matoulek M, Jarkovska Z, Marek J, Justova V, et al. Serum ghrelin levels in obese patients: The relationship to serum leptin levels and soluble leptin receptor levels. Physiol Res. 2003;52:61–6. [PubMed] [Google Scholar]
- 37.Heymsfield SB, Greenberg AS, Fujioka K, Dixon RM, Kushner R, Hunt T, et al. Recombinant leptin for weight loss in obese and lean adults: A randomized, controlled, dose-escalation trial. JAMA. 1999;282:1568–75. doi: 10.1001/jama.282.16.1568. [DOI] [PubMed] [Google Scholar]
- 38.Schulze PC, Kratzsch J, Linke A, Schoene N, Adams V, Gielen S, et al. Elevated serum levels of leptin and leptin receptor in patients with advanced chronic heart failure. Eur J Heart Fail. 2003;5:33–40. doi: 10.1016/s1388-9842(02)00177-0. [DOI] [PubMed] [Google Scholar]
- 39.Kalra SP. Central leptin gene therapy ameliorates diabetes type 1 and 2 through two independent hypothalamic relays; a benefit beyond weight and appetite regulation. Peptides. 2009;30:1957–63. doi: 10.1016/j.peptides.2009.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Farooqi IS, O’Rahilly S. Monogenic Obesity in Humans. Annu Rev Med. 2005;56:443–58. doi: 10.1146/annurev.med.56.062904.144924. [DOI] [PubMed] [Google Scholar]
- 41.Scacchi M, Pincelli AI, Cavagnini F. Growth hormone in obesity. Int J Obes Relat Metab Disord. 1999;23:260–71. doi: 10.1038/sj.ijo.0800807. [DOI] [PubMed] [Google Scholar]
- 42.Kopelman PG, Noonan K, Goulton R, Forrest AJ. Impaired growth hormone response to growth hormone releasing factor and insulin-hypoglycemia in obesity. Clin Endocrinol (Oxf) 1985;23:87–94. doi: 10.1111/j.1365-2265.1985.tb00187.x. [DOI] [PubMed] [Google Scholar]
- 43.Park MJ, Kim HS, Kang JH, Kim DH, Chung CY. Serum levels of insulin-like growth factor (IGF-1), free IGF-1, IGF-binding protein (IGFBP-1), and IGFBP-3 and insulin in obese children. J Pediatr Endocrinol Metab. 1999;12:139–44. doi: 10.1515/JPEM.1999.12.2.139. [DOI] [PubMed] [Google Scholar]
- 44.Bucher H, Zapf J, Torresani T, Prader A, Froesch ER, Illig R. Insulin-like growth factors I and II, prolactin and insulin in 19 growth hormone-deficient children with excessive, normal or decreased longitudinal growth after operation for craniopharyngioma. N Engl J Med. 1983;309:1142–6. doi: 10.1056/NEJM198311103091902. [DOI] [PubMed] [Google Scholar]
- 45.Attia N, Tamborlane WV, Heptulla R, Maggs D, Grozman A, Sherwin RS, et al. The metabolic syndrome and insulin-like growth factor I regulation in adolescent obesity. J Clin Endocrinol Metab. 1998;83:1467–71. doi: 10.1210/jcem.83.5.4827. [DOI] [PubMed] [Google Scholar]
- 46.Bistritzer T, Lovchik JC, Chalew SA, Kowarski AA. Growth without growth hormone: The ‘invisible’ GH syndrome. Lancet. 1988;1:321–3. doi: 10.1016/s0140-6736(88)91118-x. [DOI] [PubMed] [Google Scholar]
- 47.Maor G, Rochwerger M, Segev Y, Phillip M. Leptin acts as a growth factor on the chondrocytes of skeletal growth centers. J Bone Mineral Res. 2002;17:1034–43. doi: 10.1359/jbmr.2002.17.6.1034. [DOI] [PubMed] [Google Scholar]
- 48.Sinaiko AR, Jacobs DR, Jr, Steinberger J, Moran A, Luepker R, Rocchini AP, et al. Insulin resistance syndrome in childhood: Association of the euglycemic insulin clamp and fasting insulin with fatness and other risk factors. J Pediatr. 2001;139:700–7. doi: 10.1067/mpd.2001.118535. [DOI] [PubMed] [Google Scholar]
- 49.Steinberger J, Moran A, Hong CP, Jacobs DR, Jr, Sinaiko AR. Adiposity in childhood predicts insulin resistance and obesity in young adulthood. J Pediatr. 2001;138:469–73. doi: 10.1067/mpd.2001.112658. [DOI] [PubMed] [Google Scholar]
- 50.Bao W, Srinivasan SR, Berenson GS. Persistent elevation of plasma insulin levels is associated with increased cardiovascular risk in children and young adults. The Bogalusa Heart Study. Circulation. 1996;93:54–9. doi: 10.1161/01.cir.93.1.54. [DOI] [PubMed] [Google Scholar]
- 51.Chu NF, Wang DJ, Shieh SM, Rimm EB. Plasma leptin concentrations and obesity in relation to insulin resistance syndrome components among school children in Taiwan-The Taipei Children Heart Study. Int J Obes Relat Metab Disord. 2000;24:1265–71. doi: 10.1038/sj.ijo.0801404. [DOI] [PubMed] [Google Scholar]
- 52.Kamoda T, Saitoh H, Nakahara S, Izumi I, Hirano T, Matsui A. Serum leptin and insulin concentrations in prepubertal lean, obese and insulin-dependent diabetes mellitus children. Clin Endocrinol(Oxf) 1998;49:385–9. doi: 10.1046/j.1365-2265.1998.00538.x. [DOI] [PubMed] [Google Scholar]
- 53.Sudi K, Gallistl S, Trobinger M, Reiterer E, Payerl D, Aigner R, et al. Insulin and insulin resistance index are not independent determinants for the variation in leptin in obese children and adolescents. J Pediatr Endocrinol Metab. 2000;13:923–32. doi: 10.1515/JPEM.2000.13.7.923. [DOI] [PubMed] [Google Scholar]
- 54.Soliman AT, ElZalabany MM, Salama M, Ansari BM. Serum leptin concentrations during severe protein-energy malnutrition: Correlation with growth parameters and endocrine function. Metabolism. 2000;49:819–25. doi: 10.1053/meta.2000.6745. [DOI] [PubMed] [Google Scholar]
- 55.Büyükgebiz B, Oztürk Y, Yilmaz S, Arslan N. Serum leptin concentrations in children with mild protein-energy malnutrition and catch-up growth. Pediatr Int. 2004;46:534–8. doi: 10.1111/j.1442-200x.2004.01951.x. [DOI] [PubMed] [Google Scholar]
- 56.Monteleone P, DiLieto A, Castaldo E, Maj M. Leptin Functioning in Eating Disorders. CNS Spectr. 2004;9:523–9. doi: 10.1017/s1092852900009615. [DOI] [PubMed] [Google Scholar]
- 57.Holtkamp K, Herpertz-Dahlmann B, Mika C, Heer M, Heussen N, Fichter M. Elevated physical activity and low leptin levels co-occur in patients with anorexia nervosa. J Clin Endocrinol Metab. 2003;88:5169–74. doi: 10.1210/jc.2003-030569. [DOI] [PubMed] [Google Scholar]
- 58.Hebebrand J, Blum WF, Barth N, Coners H, Englaro P, Juul A, et al. Leptin levels in patients with anorexia nervosa are reduced in the acute stage and elevated upon short-term weight restoration. Mol Psychiatry. 1997;2:330–4. doi: 10.1038/sj.mp.4000282. [DOI] [PubMed] [Google Scholar]
- 59.Holtkamp K, Hebebrand J, Mika C, Grzella I, Heer M, Heussen N, et al. The effect of therapeutically induced weight gain on plasma leptin levels in patients with anorexia nervosa. J Psychiatr Res. 2003;37:165–9. doi: 10.1016/s0022-3956(02)00100-0. [DOI] [PubMed] [Google Scholar]
- 60.Plant TM, Shahab M. Neuroendocrine mechanisms that delay and initiate puberty in higher primates. Physiol Behav. 2002;77:717–22. doi: 10.1016/s0031-9384(02)00924-1. [DOI] [PubMed] [Google Scholar]
- 61.Yu WH, Kimura M, Walczewska A, Karanth S, McCann SM. Role of leptin in hypothalamic-pituitary function. Proc Natl Acad Sci USA. 1997;94:1023–8. doi: 10.1073/pnas.94.3.1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yokosuka M, Xu B, Pu S, Kalra PS, Kalra SP. Neural substrates for leptin and neuropeptide Y (NPY) interaction: Hypothalamic sites associated with inhibition of NPY-induced food intake. Physiol Behav. 1998;64:331–8. doi: 10.1016/s0031-9384(98)00065-1. [DOI] [PubMed] [Google Scholar]
- 63.Lebrethon MC, Vandersmissen E, Gerard A, Parent AS, Junien JL, Bourguignon JP. In vitro stimulation of the prepubertal rat gonadotropin-releasing-hormone pulse generator by leptin and neuropeptide Y through distinct mechanisms. Endocrinology. 2000;141:1464–9. doi: 10.1210/endo.141.4.7432. [DOI] [PubMed] [Google Scholar]
- 64.Donato J, Jr, Cravo RM, Frazão R, Elias CF. Hypothalamic sites of leptin action linking metabolism and reproduction. Neuroendocrinology. 2011;93:9–18. doi: 10.1159/000322472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Karlsson C, Lindell K, Svensson E, Bergh C, Lind P, Billig H, et al. Expression of functional leptin receptors in the human ovary. J Clin Endocrinol Metab. 1997;82:4144–8. doi: 10.1210/jcem.82.12.4446. [DOI] [PubMed] [Google Scholar]
- 66.Strobel A, Issad T, Camoin L, Ozata M, Strosberg AD. A leptin missense mutation associated with hypogonadism and morbid obesity. Nat Genet. 1998;18:213–5. doi: 10.1038/ng0398-213. [DOI] [PubMed] [Google Scholar]
- 67.Ong KK, Ahmed KL, Dunger DB. The role of leptin in human growth and puberty. Acta Paediatr Suppl. 1999;88:95–8. doi: 10.1111/j.1651-2227.1999.tb14413.x. [DOI] [PubMed] [Google Scholar]
- 68.Farooqi IS, Jebb SA, Langmack G, Lawrence E, Cheetham CH, Prentice AM, et al. Effect of recombinant leptin therapy in a child with congenital leptin deficiency. N Engl J Med. 1999;341:879–84. doi: 10.1056/NEJM199909163411204. [DOI] [PubMed] [Google Scholar]
- 69.Farooqi IS, Matarese G, Lord GM, Keogh JM, Lawrence E, Agwu C, et al. Beneficial effects of leptin on obesity, T cell hyporesponsiveness, and neuroendocrine/metabolic dysfunction of human congenital leptin deficiency. J Clin Invest. 2002;110:1093–103. doi: 10.1172/JCI15693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Gill MS, Hall CM, Tillmann V, Clayton PE. Constitutional delay in growth and puberty (CDGP) is associated with hypoleptinaemia. Clin Endocrinol (Oxf) 1999;50:721–6. doi: 10.1046/j.1365-2265.1999.00736.x. [DOI] [PubMed] [Google Scholar]
- 71.Hamrick MW, Ferrari SL. Leptin and the sympathetic connection of fat to bone. Osteoporos Int. 2008;19:905–12. doi: 10.1007/s00198-007-0487-9. [DOI] [PubMed] [Google Scholar]
- 72.Gordeladze JO, Reseland JE. A unified model for the action of leptin on bone turnover. J Cell Biochem. 2003;88:706–12. doi: 10.1002/jcb.10385. [DOI] [PubMed] [Google Scholar]
- 73.Takeda S, Elefteriou F, Levasseur R, Liu X, Zhao L, Parker KL, et al. Leptin regulates bone formation via the sympathetic nervous system. Cell. 2002;111:305–17. doi: 10.1016/s0092-8674(02)01049-8. [DOI] [PubMed] [Google Scholar]
- 74.Mantzoros CS, Rosen HN, Greenspan SL, Flier JS, Moses AC. Short-term hyperthyroidism has no effect on leptin levels in man. J Clin Endocrinol Metab. 1997;82:497–9. doi: 10.1210/jcem.82.2.3741. [DOI] [PubMed] [Google Scholar]
- 75.Sesmilo G, Casamitjana R, Halperin I, Gomis R, Vilardell E. Role of thyroid hormones on serum leptin levels. Eur J Endocrinol. 1998;139:428–30. doi: 10.1530/eje.0.1390428. [DOI] [PubMed] [Google Scholar]
- 76.Dagogo-Jack S, Selke G, Melson AK, Newcomer JW. Robust leptin secretory responses to dexamethasone in obese subjects. J Clin Endocrinol Metab. 1997;82:3230–3. doi: 10.1210/jcem.82.10.4154. [DOI] [PubMed] [Google Scholar]
- 77.Miell JP, Englaro P, Blum WF. Dexamethasone induces an acute and sustained rise in circulating leptin levels in normal human subjects. Horm Metab Res. 1996;28:704–7. doi: 10.1055/s-2007-979882. [DOI] [PubMed] [Google Scholar]
- 78.Solano JM, Jacobson L. Glucocorticoids reverse leptin effects on food intake and body fat in mice without increasing NPY mRNA. Am J Physiol. 1999;277(4 Pt 1):E708–16. doi: 10.1152/ajpendo.1999.277.4.E708. [DOI] [PubMed] [Google Scholar]
- 79.Kiess W, Englaro P, Hanitsch S, Rascher W, Attanasio A, Blum WF. High leptin concentrations in serum of very obese children are further stimulated by dexamethasone. Horm Metab Res. 1996;28:708–10. doi: 10.1055/s-2007-979883. [DOI] [PubMed] [Google Scholar]
- 80.Santos-Alvarez J, Goberna R, Sanchez-Margalet V. Human leptin stimulates proliferation and activation of human circulating monocytes. Cell Immunol. 1999;194:6–11. doi: 10.1006/cimm.1999.1490. [DOI] [PubMed] [Google Scholar]
- 81.Kiguchi N, Maeda T, Kobayashi Y, Fukazawa Y, Kishioka S. Leptin enhances CC-chemokine ligand expression in cultured murine macrophage. Biochem Biophys Res Commun. 2009;384:311–5. doi: 10.1016/j.bbrc.2009.04.121. [DOI] [PubMed] [Google Scholar]
- 82.Bruno A, Conus S, Schmid I, Simon HU. Apoptotic pathways are inhibited by leptin receptor activation in neutrophils. J Immunol. 2005;174:8090–6. doi: 10.4049/jimmunol.174.12.8090. [DOI] [PubMed] [Google Scholar]
- 83.Moore SI, Huffnagle GB, Chen GH, White ES, Mancuso P. Leptin modulates neutrophil phagocytosis of Klebsiella pneumoniae. Infect Immun. 2003;71:4182–5. doi: 10.1128/IAI.71.7.4182-4185.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Napoleone E, Di Santo A, Amore C, Baccante G, di Febbo C, Porreca E, et al. Leptin induces tissue factor expression in human peripheral blood mononuclear cells: A possible link between obesity and cardiovascular risk? J Thromb Haemost. 2007;5:1462–8. doi: 10.1111/j.1538-7836.2007.02578.x. [DOI] [PubMed] [Google Scholar]
- 85.Fernández-Riejos P, Goberna R, Sanchez-Margalet V. Leptin promotes cell survival and activates Jurkat T lymphocytes by stimulation of mitogen-activated protein kinase. Clin Exp Immunol. 2008;151:505–18. doi: 10.1111/j.1365-2249.2007.03563.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Fujita Y, Murakami M, Ogawa Y, Masuzaki H, Tanaka M, Ozaki S, et al. Leptin inhibits stress-induced apoptosis of T lymphocytes. Clin Exp Immunol. 2002;128:21–6. doi: 10.1046/j.1365-2249.2002.01797.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Farooqi IS, Matarese G, Lord GM, Keogh JM, Lawrence E, Agwu C, et al. Beneficial effects of leptin on obesity, T cell hyporesponsiveness, and neuroendocrine/metabolic dysfunction of human congenital leptin deficiency. J Clin Invest. 2002;110:1093–103. doi: 10.1172/JCI15693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Babaei A, Zarkesh-Esfahani SH, Bahrami E, Ross RJ. Restricted leptin antagonism as a therapeutic approach to treatment of autoimmune diseases. Hormones (Athens) 2011;10:16–26. doi: 10.14310/horm.2002.1289. [DOI] [PubMed] [Google Scholar]
- 89.Fraser DA, Thoen J, Reseland JE, Forre O, Kjeldsen-Kragh J. Decreased CD4+ lymphocyte activation and increased interleukin-4 production in peripheral blood of rheumatoid arthritis patients after acute starvation. Clin Rheumatol. 1999;18:394–401. doi: 10.1007/s100670050125. [DOI] [PubMed] [Google Scholar]
- 90.Otte C, Otte JM, Strodthoff D, Bornstein SR, Folsch UR, Monig H, et al. Expression of leptin and leptin receptor during the development of liver fibrosis and cirrhosis. Exp Clin Endocrinol Diabetes. 2004;112:10–7. doi: 10.1055/s-2004-815720. [DOI] [PubMed] [Google Scholar]
- 91.Piche T, Vandenbos F, Abakar-Mahamat A, Vanbiervliet G, Barjoan EM, Calle G, et al. The severity of liver fibrosis is associated with high leptin levels in chronic hepatitis C. J Viral Hepat. 2004;11:91–6. doi: 10.1046/j.1365-2893.2003.00483.x. [DOI] [PubMed] [Google Scholar]
- 92.Kitade M, Yoshiji H, Kojima H, Ikenaka Y, Noguchi R, Kaji K, et al. Leptin-Mediated Neovascularization Is a Prerequisite for Progression of Nonalcoholic Steatohepatitis in Rats. Hepatology. 2006;44:983–91. doi: 10.1002/hep.21338. [DOI] [PubMed] [Google Scholar]
- 93.Haynes WG, Morgan DA, Walsh SA, Mark AL, Sivitz WI. Receptor mediated regional sympathetic nerve activation by leptin. J Clin Invest. 1997;100:270–8. doi: 10.1172/JCI119532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Matsumura K, Abe I, Tsuchihashi T, Fujishima M. Central effects of leptin on cardiovascular and neurohormonal responses in conscious rabbits. Am J Physiol Regul Integr Comp Physiol. 2000;278:R1314–20. doi: 10.1152/ajpregu.2000.278.5.R1314. [DOI] [PubMed] [Google Scholar]
- 95.Shirasaka T, Takasaki M, Kannan H. Cardiovascular effects of leptin and orexins. American Journal of Physiology. Regulatory. Am J Physiol Regul Integr Comp Physiol. 2003;284:R369–51. doi: 10.1152/ajpregu.00359.2002. [DOI] [PubMed] [Google Scholar]
- 96.Ren J. Leptin and hyperleptinemia – from friend to foe for cardiovascular function. J Endocrinol. 2004;181:1–10. doi: 10.1677/joe.0.1810001. [DOI] [PubMed] [Google Scholar]
- 97.Fujimaki S, Kanda T, Fujita K, Tamura J, Kobayashi I. The significance of measuring plasma leptin in acute myocardial infarction. J Int Med Res. 2001;29:108–13. doi: 10.1177/147323000102900207. [DOI] [PubMed] [Google Scholar]
- 98.El-Melegy NT, Mohamed NA. Angiogenic biomarkers in children with congenital heart disease: Possible implications. Ital J Pediatr. 2010;36:32. doi: 10.1186/1824-7288-36-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Gibson WT, Farooqi IS, Moreau M, DePaoli AM, Lawrence E, O’Rahilly S, et al. Congenital Leptin Deficiency Due to Homozygosity for the Δ133G Mutation: Report of Another Case and Evaluation of Response to Four Years of Leptin Therapy. J Clin Endocrinol Metab. 2004;89:4281–6. doi: 10.1210/jc.2004-0376. [DOI] [PubMed] [Google Scholar]
- 100.Montague CT, Farooqi IS, Whitehead JP, Soos MA, Rau H, Wareham NJ, et al. Congenital leptin deficiency is associated with severe early-onset obesity in humans. Nature. 1997;387:903–8. doi: 10.1038/43185. [DOI] [PubMed] [Google Scholar]
- 101.Strobel A, Issad T, Camoin L, Ozata M, Strosberg AD. A leptin missense mutation associated with hypogonadism and morbid obesity. Nat Genet. 1998;18:213–5. doi: 10.1038/ng0398-213. [DOI] [PubMed] [Google Scholar]
- 102.Farooqi IS, Matarese G, Lord GM, Keogh JM, Lawrence E, Agwu C, et al. Beneficial effects of leptin on obesity, T cell hyporesponsiveness, and neuroendocrine/metabolic dysfunction of human congenital leptin deficiency. J Clin Invest. 2002;110:1093–103. doi: 10.1172/JCI15693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Licinio J, Caglayan S, Ozata M, Yildiz BO, de Miranda PB, O’Kirwan F, et al. Phenotypic effects of leptin replacement on morbid obesity, diabetes mellitus, hypogonadism, and behavior in leptin-deficient adults. Proc Natl Acad Sci USA. 2004;101:4531–6. doi: 10.1073/pnas.0308767101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Javor ED, Cochran EK, Musso C, Young JR, Depaoli AM, Gorden P. Long-term efficacy of leptin replacement in patients with generalized lipodystrophy. Diabetes. 2005;54:1994–2002. doi: 10.2337/diabetes.54.7.1994. [DOI] [PubMed] [Google Scholar]
- 105.Chong AY, Lupsa BC, Cochran EK, Gorden P. Efficacy of leptin therapy in the different forms of human lipodystrophy. Diabetologia. 2010;53:27–35. doi: 10.1007/s00125-009-1502-9. [DOI] [PubMed] [Google Scholar]
- 106.Wanga MY, Chena L, Clarka GO, Leea Y, Stevensb RD, Ilkayevab OR, et al. Leptin therapy in insulin-deficient type I diabetes. Proc Natl Acad Sci U S A. 2010;107:4813–19. doi: 10.1073/pnas.0909422107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Cummings BP, Bettaie A, Graham JL, Stanhope KL, Dill R, Morton GJ, et al. Subcutaneous administration of leptin normalizes fasting plasma glucose in obese type 2 diabetic UCD-T2DM rats. Proc Natl Acad Sci U S A. 2011;108:14670–5. doi: 10.1073/pnas.1107163108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Mittendorfer B, Horowitz JF, DePaoli AM, McCamish MA, Patterson BW, Klein S. Recombinant Human Leptin Treatment Does Not Improve Insulin Action in Obese Subjects With Type 2 Diabetes. Diabetes. 2011;60:1474–7. doi: 10.2337/db10-1302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Semple RK, Soos MA, Luan J, Mitchell CS, Wilson JC, Gurnell M, et al. Elevated Plasma Adiponectin in Humans with Genetically Defective Insulin Receptors. J Clin Endocrinol Metab. 2006;91:3219–23. doi: 10.1210/jc.2006-0166. [DOI] [PubMed] [Google Scholar]
- 110.Moran A, Jacobs DR, Jr, Steinberger J, Hong CP, Prineas R, Luepker R, et al. Insulin resistance during puberty: Results from insulin clamp studies in 356 children. Diabetes. 1999;48:2038–44. doi: 10.2337/diabetes.48.10.2039. [DOI] [PubMed] [Google Scholar]