

Abstract
In human, mutation of the transcription factor SATB2 causes severe defects to the palate and jaw. The expression and sequence of SATB2 is highly conserved across vertebrate species, including zebrafish. We sought to understand the regulation of satb2 using the zebrafish model system. Due to the normal expression domains of satb2, we analyzed satb2 expression in mutants with disrupted Hh signaling or defective ventral patterning. While satb2 expression appears independent of Edn1 signaling, appropriate expression requires Shha, Smo, Smad5 and Hand2 function. Transplantation experiments show that neural crest cells receive both Bmp and Hh signaling to induce satb2 expression. Dorsomorphin- and cyclopamine-mediated inhibition of Bmp and Hh signaling, respectively, suggests that proper satb2 expression requires a relatively earlier Bmp signal and a later Hh signal. We propose that Bmp signaling establishes competence for the neural crest to respond to Hh signaling, thus inducing satb2 expression.
Introduction
Cranial neural crest cells are a highly specialized vertebrate-specific cell type which give rise to diverse structures including most of the bone and cartilage of the head, face and palate. In all vertebrates, this pluripotent cell population migrates to the periphery in three streams and condenses within transient structures known as pharyngeal arches [1], [2]. In amniotes, neural crest cells of the anterior-most stream contribute to the jaw and palatal skeleton [3], [4]. The palatal precursors reside in the frontonasal prominence to generate the primary palate, and the maxillary prominence of the first arch to generate the secondary palate and upper jaw [5]. The mandibular prominence of the first arch contributes to the lower jaw. The second crest stream migrates to the second arch and contributes skeletal elements such as the hyoid bone 6,7. The third stream populates the more posterior arches. Zebrafish have similar neural crest cell populations, with frontonasal and maxillary crest populations generating distinct regions of the anterior neurocranium, or zebrafish palate [8], [9], [10], [11]. As in amniotes, maxillary and mandibular neural crest give rise to the upper and lower jaws, respectively and neural crest cells of the second arch form the dorsal and ventral jaw support elements [8], [12].
Pharyngeal arches consist of a mesodermal core surrounded by neural crest cell-derived mesenchyme that is encapsulated by epithelia (the ectoderm and the endoderm). Reciprocal interactions between these different cell types are critical to establish the signaling networks required to generate the multitude of craniofacial elements of the appropriate size and shape [5], [9], [13]. Craniofacial growth and specification is controlled by the interaction of numerous signaling pathways, including those mediated via Sonic hedgehog (Shh), Endothelin-1 (Edn1) and Bone morphogenic protein (Bmp) [5]. Across vertebrates, the Bmp and Edn1 pathways pattern of the jaw. Mice and zebrafish carrying mutations of Edn1 show reduced expression of genes known to be essential for mandibular development such as Hand2 [14], [15], [16], [17]. Bmp also functions in ventral patterning of the pharyngeal arches with activities both upstream and independent of Edn1 [18], [19], [20], [21]. Notch signaling acts antagonistically to these ventral signals to establish dorsal identity [22]. In addition to its role in ventral arch patterning, Bmp signals play crucial roles in the development of the palatal skeleton in amniotes and zebrafish [12], [23], [24], [25]. In its role during palatogenesis, Bmp signaling induces the expression of another critically important signaling molecule, Shh 25,26.
Shh is a member of the secreted family of Hedgehog (Hh) proteins and all hedgehog signaling is transduced through Smoothened (Smo). Shh is crucial for regulating patterning and outgrowth of the face and palate across vertebrate species [8], [10], [11], [27], [28], [29], [30]. Mutation of human SHH or its downstream effectors result in a variety of severe developmental disorders; those affecting the head and face include holoprosencephaly, cyclopia and hypotelorism (Belloni et al., 1996; Dubourg et al., 2007; Nanni et al., 1999; Roessler et al., 1996). Neural crest cells must receive Hh signaling for the appropriate expression of patterning genes in the developing craniofacial skeleton [11], [29]. In the developing palatal skeleton, Shh signaling plays a crucial role in outgrowth of the palatal shelves and in maintaining proliferation of the palatal mesenchyme [5], [31].
Considering the numerous transcription factors and signaling molecules involved in regulating craniofacial development it is not surprising that craniofacial defects are common among congenital birth defects. The most common craniofacial defects are cleft lip and/or palate and mandibular dysmorphogenesis [32]. The use of model organisms in dissecting the underlying molecular pathways regulating facial development and palatogenesis is imperative to our understanding of these processes in humans. Such work has been carried out to understand the role of the transcription factor SATB2. Mutation of this gene in human and mouse results in strikingly similar craniofacial defects including cleft palate and jaw abnormalities [33], [34], [35], [36], [37]. We have previously shown that the sequence identity of the SATB2 protein and expression of the transcript is highly conserved across vertebrate species including zebrafish [38].
Here, we use zebrafish to gain insight into how Satb2 integrates into the signaling cascades involved in craniofacial development. We show that satb2 is a target of Bmp and Hh signaling. Using genetic mosaics, we find that neural crest cells must receive both signals for proper satb2 expression. Using pharmacologic inhibition of each pathway, we find that Bmp signaling is required temporally before Hh signaling. Collectively, we provide some of the first evidence regarding the regulation of satb2.
Materials and Methods
Zebrafish Lines
The study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Institutional Animal Care and Use Committee at the University of Texas at Austin (AUP-2012-00053). Zebrafish (Danio rerio) embryos were raised at 28.5°C and staged as described previously [39]. Some embryos used for in situ hybridization analysis were incubated in embryo media containing PTU to prevent pigmentation. Cyclopamine experiments [8] were performed in the AB background or the transgenic tg(fli1a;EGFP)y1 line, designated fli1:EGFP for clarity [40]. Dorsomorphin (DM – Cat. #3093; Tocris Bioscience, Ellisville, MO) was dissolved in dimethyl sulfoxide (DMSO) to a concentration of 10 mM and aliquoted for long-term storage at −20°C. Zebrafish embryos were treated with 10 µM DM or with 10 µM DMSO alone as a control. Dorsomorphin experiments were carried out in the transgenic tg(BRE-AAVmlp:dmKO2)mw40 line, designated BRE:mKO2 [41]. The inhibitory effects of dorsomorphin was verified by the loss of BRE:mKO2 expression at the time embryos were fixed for in situ hybridization. The exact same settings were used for the collection of all BRE:mKO2 images. shhat4 [42], smob577 [43], edn1tf216b [16], hand2s6 [44] and sox32ta56 [45] mutant embryos were identified by morphology or genotyping as described previously. The edn1 and hand2 mutant embryos were kind gifts from The University of Oregon aquarium. Heterozygous carriers of the smoothened null allele, smo, and sox32 allele [43], [46] were maintained on the fli1:EGFP background and are referred to as smo- and sox32− in the text for clarity. The smad5b1100 [12] allele is a cysteine to thymidine mutation at nt 733 of the first coding exon of smad5. This mutation is predicted to generate a Proline to Serine missense mutation in the DNA binding MH1 domain of Smad5. PCR with primers jaa1 (5′ aagggcctcccacacgtcatct 3′) and yj32 (5′ ctggactttcaactcgtttgtagtgccatg 3′), followed by digestion with NcoI was used for genotyping. The mutant allele is 160 bp and the wt allele is 129 bp.
Skeletal Analysis and in situ Hybridization
Zebrafish larvae were stained with Alcian Blue and Alizarin Red [47] and the cartilages flat-mounted at 5 days post-fertilization (dpf). The satb2 probe used for in situ hybridization is described elsewhere [38].
Cell Transplantations
Transplantation experiments were performed as described elsewhere [8]. Briefly, donor embryos were injected with Alexa-568 dextran at the one to two cell stage. At shield stage, donor and host embryos were placed on a depression slide in filter sterilized Ringers solution containing Penicillin and Streptomycin, immobilized in 4% methylcellulose. Using a pulled micropipette, approximately 20–30 cells were removed from donor embryos and transplanted to the neural crest domain [48] of host embryos. Transplanted embryos were allowed to recover for 15–20 minutes and then placed in filter sterilized embryo media containing Penicillin and Streptomycin.
Results
Multiple Signaling Pathways Regulate satb2 Expression
Despite its functional involvement in growth and development, the regulation of Satb2 is largely unknown. Because it plays a central role in gene regulation [49], [50], we hypothesized that Satb2 would be an effector of some of the major signaling pathways crucial for development of the face and palate. Because of the restriction of satb2 in the ventral arch we focused our analyses on the Edn1, Bmp and Hh pathways. Expression analyses were performed at 36 and 48 hpf corresponding with the peak of satb2 expression in crest-derived mesenchyme (Fig. 1). In zebrafish, and other vertebrates, satb2 is expressed by neural crest cell-derived mesenchyme in both the maxillary and frontonasal prominences [33], [35], [38]. For simplicity, we refer to this expression domain collectively as palatal precursors.
Figure 1. Expression of satb2 requires Hh and Bmp signaling.

(A–L) Lateral images of satb2 expression at 36 or 48 hpf. (A–D) Compared to wild-type embryos, in 36 and 48 hpf shha− embryos, satb2 expression is reduced in the palatal precursors (arrows in C and D) and throughout the pharyngeal arches. (E,F) Nearly all neural crest cell expression of satb2 appears to be absent in smo− embryos. The prominent expression in panel F, is in neural tissue. (G,H) Weak to no expression of satb2 was observed in smad5 mutant embryos at 36 and 48 hpf. (I,J) satb2 expression in edn1 mutants was similar wild-type embryos at both time points analyzed. (K,L) While expression of satb2 was absent throughout the posterior arches of hand2 mutants, the ventral first arch and palatal precursors maintained satb2 expression. p, palatal precursors; e, eye; The arches are numbered in A & C for clarity.
Our results show that multiple signaling pathways are essential for appropriate expression of satb2. Hh signaling is necessary for the condensation of palatal precursors [8], [10], precluding analysis of this population. Ventral pharyngeal arch expression of satb2 is reduced in the absence of Shha (Fig. 1 A–D) and almost completely lost in smo mutants, which lack all Hh signaling (Fig. 1 E,F), even though the ventral arch is present and maintains expression of other markers [11]. Bmp signaling is essential for palatogenesis and dorsal/ventral patterning of the arches in zebrafish [18], [19]. In smad5 mutants, in which Bmp signaling is disrupted, palatal precursors have been shown to condense [12] here we show that they fail to express satb2 (Fig. 1 G,H). Furthermore, expression of satb2 in the ventral pharyngeal arch was greatly reduced or absent (Fig. 1 G,H). Disruption of Bmp signaling has been shown to expand dorsal arch fates at the expense of ventral fates [18], [19] strongly supporting an interpretation that these results are due to a loss of satb2 expression as opposed to the loss of territory normally expressing satb2. While Edn1 signaling is also indispensible for dorsal/ventral patterning of the zebrafish pharyngeal arches [16], [51], [52], satb2 expression appeared normal in edn1 mutants (Fig. 1 I,J). hand2 is a target of both Bmp and Edn1 signaling [15], [18], [19] and from our data the expression domain of hand2 appears to overlap extensively with satb2 [38]. Only palatal crest and a population of crest in the ventral first arch maintained satb2 expression in hand2 mutants (Fig. 1 K,L).
Neural Crest Cells Must Receive both Hedgehog and Bmp Signaling for Proper satb2 Expression
Reception of Hh signaling by neural crest cells is necessary for the proper expression of Fox genes and several markers of dorsal/ventral patterning [11], [29]. To determine if neural crest cells require the reception of Hh signaling for satb2 expression, we generated genetic mosaics to restore the reception of signaling in neural crest cells. We have previously shown that an early Hh signal from the ventral brain to the oral ectoderm is required for neural crest condensation in the maxillary region [8] and thus crest transplants fail to rescue the condensation defect. Therefore, for these analyses we focused on satb2 expression in the ventral arches. We analyzed only embryos with extensive contribution of donor neural crest cells to the ventral arches and found that transplantation of smo+/+ neural crest cells into smo mutants rescued satb2 expression (Fig. 2 A, B; n = 7/7). These data indicate that Hh signaling to the neural crest is necessary for satb2 expression in the ventral arch.
Figure 2. Neural crest cells require the reception of Bmp and Hh signaling to express satb2.

(A, C) Lateral views at 36 hpf; (B, D) ventral views at 36 hpf. Both donor and host embryos were fli1:EFGP transgenics to allow for visualization of neural crest cells. (A) Wild-type crest transplanted into the arches of smo mutant embryos are shown in red in the inset. (B) smo+ crest restores the ventral arch expression of satb2 on the side of the transplant (t) as compared to the mutant side of the embryo (m). (C) Lateral views of a 36 hpf smad5 mutant that received a transplant of smad5+ neural crest. The distribution of smad5+ cells (red in inset) is shown. (D) Expression of satb2 is restored in the ventral pharyngeal arch and in the palatal precursors (arrow) on the transplanted side of the embryo. Arches 1 & 2 are numbered in each panel.
The zebrafish ventral arch also responds to Bmp signaling [18], [19]. We used a similar genetic mosaic strategy to determine if neural crest cells must receive Bmp signaling to express satb2. Transplantation of smad5+/+ neural crest cells into smad5 mutants rescued the expression of satb2 in the ventral pharyngeal arch (Fig. 2 C, D; n = 7/7). This transplant also sparsely populated the region of the maxillary domain expressing satb2. As such, a very small population of satb2 expressing cells are evident (Fig. 2, arrow). Collectively, these results demonstrate that neural crest cells must receive, at a minimum, both Hh and Bmp signaling in order to express satb2.
Endoderm is a Critical Source of Signals Necessary for satb2 Expression
In zebrafish, satb2 is most strongly expressed in neural crest cells in the medial region of the arch that are in close proximity to the pharyngeal endoderm, a source of both Bmp and Hh signaling [8], [12], [53], [54]. To determine if the endoderm is necessary for satb2 expression, we analyzed sox32 mutants, which lack endoderm [46]. At 38 hpf wild-type embryos expressed satb2 in the ventral arch neural crest as well as palatal precursor cells (Fig. 3 A). While the expression of satb2 was retained in the palatal precursors and the ventral first arch in sox32 mutants, satb2 expression was lost in the more posterior pharyngeal arches (Fig. 3 B). Collectively, these results suggest that signals from the endoderm are essential to induce satb2 in the ventral region of the second and more posterior arches, while Bmp and Shh signaling from other tissues, such as facial ectoderm [8], [12], [53], are important to induce satb2 in the ventral first arch and palatal precursors.
Figure 3. Endodermal signals are required for satb2 expression in ventral arches 2–7.
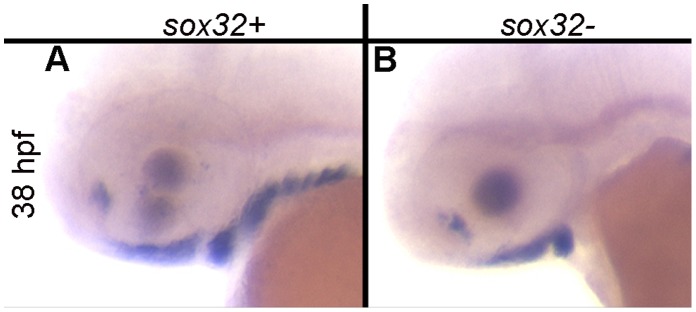
(A) At 38 hpf, satb2 is expressed robustly in the ventral pharyngeal arches and palatal precursors of wild-type embryos. (B) Expression of satb2 in the palatal precursors and ventral arch 1 is retained in sox32 mutants; however, satb2 expression is ablated in the posterior arches.
While Bmps are expressed much earlier in the endoderm [12], [54] relative to the timing of satb2 expression in the ventral arch, the endoderm begins to express shha at 30 hpf (Fig. 4 A). By 32 hpf satb2 expression is initiated in the adjacent arches 1–3 (Fig. 4 C, D). These observations suggest a model in which Bmp and Shh signaling are sequential events both necessary for the induction of satb2 in the ventral arch. To test this model we used pharmacological inhibition of the two pathways to determine when Bmp and Shh signaling are required for satb2 expression.
Figure 4. Expression of shh and satb2 correlate spatially during pharyngeal arch development.

(A–D) Dorsal-lateral views of 30 hpf and 32 hpf wild-type embryos. (A) shha expression initiates in the pharyngeal endoderm at 30 hpf while (B) satb2 is not expressed in the craniofacial region at this time. (C, arrows) By 32 hpf shh expression has intensified and can be seen to arc around the presumptive arches. (D, arrows) satb2 expression is initiated in the ventral arch by 32 hpf and lies in close proximity to the presumptive pharyngeal endoderm.
Sequential Bmp and Shh Signaling Regulates satb2 Expression
Ventral arch neural crest cells respond to Bmp signaling beginning by at least 24 hpf [18]. Therefore, we first tested if this initial Bmp response was essential for satb2 expression. Embryos treated with dorsomorphin from 20 to 36 hpf showed a dramatic reduction in the level of satb2 expression relative to untreated control embryos (Fig. 5 A–D). Compared to controls, expression of satb2 in the palatal precursors was undetectable (Fig. 5 A & C, arrowhead) and only a small region of ventral medial second arch crest maintained a low level of satb2 expression (Fig. 5 B & D, arrows). Even when dorsomorphin was washed off at 30 hpf and the embryos were allowed to further develop to 36 hpf, the expression of satb2 expression was still dramatically reduced (Fig. 5 E & F), although not to the extent seen with Bmp blockage from 20–36 hpf. These analyses were performed in BRE:mKO2 transgenics, allowing us to verify that Bmp signaling had been partially, albeit not completely, restored in the ventral neural crest (Figure S1). While the expression of satb2 in the palatal precursors was still absent (Fig. 5 E, arrowhead), a small number of ventral medial crest cells within the first and second arch expressed satb2 weakly (Fig. 5 F, arrows). While the partial recovery of satb2 expression and the BRE response correlate with one another, blocking Bmp signaling from 30 to 42 hpf caused no gross alteration to satb2 expression (Fig. 5 G–J). Collectively, these results demonstrate that the Bmp signal necessary for satb2 expression is required predominantly prior to the initiation of satb2 and that, following this initial signaling event, canonical Bmp signaling is dispensable for satb2 expression.
Figure 5. Bmp signaling is required during early arch patterning for the appropriate expression of satb2.

(A, C, E) Lateral and (B, D, F) ventral images of 36 hpf embryos labelled with satb2 riboprobe. (A) The ventral region of the pharyngeal arches expresses satb2 robustly. (B) In the ventral arches, satb2 expression is strongest medially. (C, D) Inhibiting Bmp signaling via dorsomorphin from 20 to 36 hpf eliminates the majority of satb2 expression including that in the palatal precursors (arrowhead in C). Only a small population of crest in ventromedial arch 2 expresses satb2 following this treatment (arrows in C & D). (E, F) When Bmp signaling is blocked from 20 to 30 hpf and allowed to recover until 36 hpf, satb2 expression is still greatly reduced. Palatal precursors fail to express satb2 (arrowhead in E) and only ventromedial crest in arches 1 & 2 express satb2 (arrows in E & F). Later inhibition of Bmp signaling does not alter satb2 expression. (G–J) 42 hpf embryos labelled with riboprobe to satb2 in lateral (G, I) and ventral (H, J) views. In both control embryos (G, H) and embryos treated with dorsomorphin from 30 to 42 hpf (I, J), satb2 is strongly expressed by the palatal precursors and in the ventral pharyngeal arches. l, lateral; m, medial; p, palatal precursors.
Initiation of satb2 expression in the pharyngeal arches shortly follows the initiation of shha expression in the pharyngeal endoderm, suggesting that Hh signaling may be more proximal to the initiation of satb2 than Bmp signaling. Overall, we found that cyclopamine treatments that started at 30 hpf, just prior to the initiation of satb2 expression, caused dramatic loss in satb2 expression (Fig. 6 A–L). By blocking Hh between 30–60 hpf, corresponding with the apparent peak of satb2 expression, we see the most dramatic reduction of satb2 (Fig. 6 A–D). In these embryos only ventral arch 1 expressed satb2 strongly, with weaker expression seen in the palatal precursors and ventral arch 2 (Fig. 6 C & D). Cyclopamine treatment from 30–36 hpf gave similar results (Fig. 6 E–H). If, however, embryos were treated between 30 to 36 hpf, rinsed and then reared for another 10 hours, partial recovery of satb2 expression in the arches was observed (Fig. 6 I–L), suggesting that Hh signaling continues to be important for satb2 expression. We performed further cyclopamine treatments to investigate if satb2 requires a continuous Hh signal to maintain expression.
Figure 6. Continuous Hh signaling is required for satb2 expression.

Ventral (A, B, I, J, M, N, Q, R), lateral (C, D, E, F, K, L, O, P, S, T, U, V) and dorsal-lateral (G, H) images of satb2 expression by in situ hybridization are shown in control and cyclopmaine-treated (CYA) embryos. (A, C) In controls at 60 hpf, satb2 is expressed in palatal precursors and in the ventral region of each arch. (B, D) Embryos treated with cyclopamine from 30–60 hpf show a dramatic reduction of satb2 expression in the palatal precursors, pharyngeal arch 1 and 2 as well as complete loss of expression in the posterior arches. (E–H) Compared to controls, embryos treated with cyclopamine from 30 to 36 hpf also show reduction of satb2 expression in the palatal precursors and arches 1 and 2, with a more complete loss of expression in the more posterior arches. (I–L) If, however, embryos are removed from cyclopamine at 36 hpf and allowed to develop for 10 hours, there is a partial recovery of satb2 expression. (M–T) Cyclopamine treatment either from 38 to 42 hpf (M–P) or 44 to 50 hpf (Q–T) results in mild reduction of satb2 expression in the palatal precursors and pharyngeal arches. (U, V) While the maxillary domain is lost in embryos treated with cyclopamine from 6–30 hpf the expression in the ventral arches appears largely intact. CYA, cyclopamine; e, eye; b, brain; p, palatal precursors; pharyngeal arches are numbered in some panels for clarity.
We used either a 4 or 6-hour treatment of cyclopamine to determine the temporal requirements for Hh signaling in satb2 expression. Embryos were treated with cyclopamine between 38–42 hpf and 44–50 hpf and satb2 expression assessed immediately following treatment (Fig. 6 M–P and Q–T, respectively). At both time points the cyclopamine-treated embryos showed reduction of satb2 expression in the arches and subtle reduction in the palatal precursors. These observations suggest that continued Hh signaling is necessary to maintain an appropriate level of satb2 expression.
During zebrafish palatogenesis, an early Hh signal from the presumptive ventral brain signals to the oral ectoderm 12 hours prior to the arrival of CNCC in the maxillary domain [8]. This early signal is required for CNCC condensation and proper palatogenesis to occur [8]. Thus, we asked whether or not an early Hh signal was necessary to ‘prime’ the ventral arch CNCC to express satb2 later in development. We blocked Hh with cyclopamine between 6 hpf and 30 hpf then analyzed satb2 expression at 42 hpf, once Hh signaling had recovered. Inhibiting Hh signaling during this time window disrupts the condensation of palatal precursors [8], preventing an analysis of this cell population. However, the level of satb2 expression in the arches of treated embryos appeared similar to non-treated controls suggesting that an early Hh signal is not required to pre-pattern the ventral arch to express satb2 (Fig. 6 U, V). Collectively, our inhibitor studies strongly suggest that Bmp signaling to the ventral pharyngeal arch mesenchyme is required prior to Hh signaling and that Hh signaling continues to be important for satb2 throughout arch development.
Discussion/Conclusions
satb2 Expression Depends upon the Integration of Bmp and Hh Signaling
We show that Hh and Bmp signaling are required for satb2 expression in the developing pharyngeal arches (Fig. 7). The expression of satb2 in both the maxillary domain and ventral pharyngeal arches require Bmp and Hh signaling. Blocking Bmp signaling from 20–36 hpf vastly decreases satb2 expression in both the maxillary domain and ventral pharyngeal arches. Likewise, both maxillary domain and ventral arch expression of satb2 is greatly reduced by blocking Hh signaling from 30–36 hpf. While these results show that the timing of Bmp and Hh signaling important for satb2 expression is similar, it is possible that the precise mechanisms by which Bmp and Hh signals regulate satb2 expression across neural crest domains differs. Future experiments aimed at characterizing the induction of satb2 expression will be necessary to fully understand these potential differences.
Figure 7. Model.
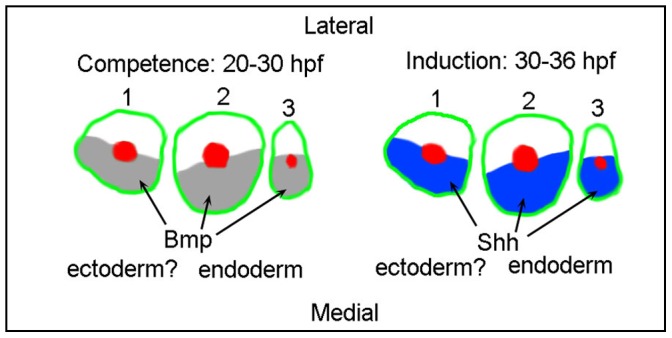
Bmp signaling and Shh signaling are each necessary for proper satb2 expression. Bmp signaling is necessary prior to both the induction of satb2 and the time window when Hh signaling is necessary for satb2 expression. Shh signaling is required immediately prior to and overlapping with the induction of satb2 expression in the pharyngeal arches. Additionally, the induction of satb2 occurs in crest in close proximity to shha-expressing endoderm. These findings support a model in which the early Bmp signal establishes competence of the neural crest to respond to the later Shh signal. The first three arches are shown in schematic and numbered.
The frequent coincident expression of Hh and Bmp ligands has long suggested functional interactions between these pathways [55]. During the development of numerous craniofacial elements these two signaling pathways have been shown to interact. Typically, these signaling pathways regulate one another through epithelial-mesenchymal interactions, such as during development of the palatal skeleton [26], [56], tooth germ [57], [58], frontonasal ectodermal zone [59] and scleral ossicles [60]. Additionally, ligands for both pathways have clear roles in the facial epithelia [56], [61]. While our data do not fully determine the sources of Shh and Bmp necessary for the induction of satb2 expression, our analyses of sox32 mutants show that the pharyngeal endoderm is clearly important for ventral expression in arches 2–7. It is likely that ectodermal expression of these ligands [8], [12], 53 is important for satb2 expression in the ventral first arch.
It has been shown in zebrafish that ventral specification of the pharyngeal arches requires the Bmp and Edn1 pathways acting in opposition to dorsal Jagged/Notch signaling [16], [18], [19], [22]. Due to their known roles in other vertebrate species, these interactions are likely to be conserved in amniotes [13], [20], [62], [63], [64], [65]. The reception of these signals is necessary in the neural crest for proper specification [19], [22], [52], [66], [67], although Edn1 signaling to the facial epithelia is also important [68]. Appropriate specification of the pharyngeal arches also requires the reception of Hh signaling by the neural crest [11], [29]. Here, we show that neural crest cells must receive both Bmp and Hh signaling for the ventral expression of satb2. Hand2 is downstream of Edn1 and Bmp signaling during craniofacial development [14], [16], [18], [19] and is also necessary for satb2 expression, at least in the second and more posterior arches. Our previous analyses have shown that hand2 expression is retained in zebrafish smo mutants [11], therefore, we can conclude that Hand2 is necessary but not sufficient for appropriate satb2 expression.
Because hand2 is an Edn1 target [14], [16], it was surprising to find that satb2 expression was normal in edn1 mutants. However, hand2 expression is not completely lost in zebrafish edn1 mutants [16]. Thus, this residual hand2 expression may activate satb2 expression. Alternatively, expression of satb2 may be influenced by an Edn1-independent effecter such as Mef2ca [66]. While mef2ca expression in the zebrafish pharyngeal arches does not require Edn1 signaling, the expression of Edn1 target genes requires Mef2ca function [66]. Differential regulation of satb2 expression by Edn1 and Hand2 would not be completely surprising as it has been suggested that Hand2 maybe a branch point of Edn1 regulation during arch development [13].
While Bmp and Hh signaling are crucial for satb2 expression, the timing of these signaling events would appear to be somewhat, although not completely, distinct. Even though the ventral pharyngeal arch continues to respond to Bmp signaling until at least 48 hpf [18], our dorsomorphin analysis suggests that signaling past 30 hpf is dispensable for satb2 expression. In contrast, our cyclopamine data suggest that Hh signaling from 30 hpf onward is essential for appropriate satb2 expression. In our cyclopamine treatments we never eliminated satb2 expression, suggesting an incomplete inhibition of the Hh pathway or that some earlier Hh signaling is important in the expression of satb2 in the ventral first and second arch. Even though the necessary Bmp signaling event precedes the initiation of satb2, it has recently been shown that Smad1/5 binds chromatin approximately 1 kb 5′ of Satb2 in mouse mandibles [69]. While similar analyses of Gli binding regions near Satb2 in neural crest have not been performed, a strong Gli3 binding region is located just over 100 kb 5′ of Satb2 in neural tissue [70]. Because both Bmp and Hh signaling are necessary for satb2 expression, we propose a model to be tested in which the earlier Bmp signal establishes competence for neural crest cells to respond to a later Hh signal (Fig. 7).
Supporting Information
Wash out of Dorsomorphin causes a partial restoration of Bmp signaling. (A) Untreated 30 hpf BRE:mKO2 embryos have rhobust transgene expression in the ventral pharyngeal arches (arrows). (B) Nearly all expression, except for some in the dorsal retina is lost following dorsomorphin treatment from 20–30 hpf. (C) At 36 hpf the expression of the BRE:mKO2 transgene closely resembles that observed at 30 hpf. (D) Embryos treated with dorsomorphin from 20–30 hpf and then washed out of the drug show a partial recovery of BRE:mKO2 expression in the ventral pharyngeal arches (arrows).
(TIF)
Acknowledgments
We would like to thank the staff of the Fish Facilities at the University of Manchester, University of Texas at Austin and the University of Oregon for their time and help in rearing and providing fish and embryo stocks. Thanks to Young-Jun Jeon and Joshua Atkinson for help in genotyping the smad5 mutations. Many thanks to Dr. Charles Kimmel for providing his lab staff and facilities to help train K. S-R. Thanks to Dr. Jeffrey Gross and his lab for use of their microscope.
Funding Statement
This work was supported by United States National Institutes of Health grants DE018088 and DE020884 to J.K.E. and MRC grant (G1001601) and support from the Healing Foundation to M.J.D. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Noden DM (1991) Cell movements and control of patterned tissue assembly during craniofacial development. J Craniofac Genet Dev Biol 11: 192–213. [PubMed] [Google Scholar]
- 2. Kuratani S (2005) Cephalic neural crest cells and the evolution of craniofacial structures in vertebrates: morphological and embryological significance of the premandibular-mandibular boundary. Zoology (Jena) 108: 13–25. [DOI] [PubMed] [Google Scholar]
- 3. Chai Y, Maxson RE Jr (2006) Recent advances in craniofacial morphogenesis. Dev Dyn 235: 2353–2375. [DOI] [PubMed] [Google Scholar]
- 4. Trainor PA, Melton KR, Manzanares M (2003) Origins and plasticity of neural crest cells and their roles in jaw and craniofacial evolution. Int J Dev Biol 47: 541–553. [PubMed] [Google Scholar]
- 5. Bush JO, Jiang R (2012) Palatogenesis: morphogenetic and molecular mechanisms of secondary palate development. Development 139: 231–243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Kontges G, Lumsden A (1996) Rhombencephalic neural crest segmentation is preserved throughout craniofacial ontogeny. Development 122: 3229–3242. [DOI] [PubMed] [Google Scholar]
- 7. Gross JB, Hanken J (2008) Review of fate-mapping studies of osteogenic cranial neural crest in vertebrates. Dev Biol 317: 389–400. [DOI] [PubMed] [Google Scholar]
- 8. Eberhart JK, Swartz ME, Crump JG, Kimmel CB (2006) Early Hedgehog signaling from neural to oral epithelium organizes anterior craniofacial development. Development 133: 1069–1077. [DOI] [PubMed] [Google Scholar]
- 9. Kimmel CB, Eberhart JK (2008) The midline, oral ectoderm, and the arch-0 problem. Integr Comp Biol 48: 668–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Wada N, Javidan Y, Nelson S, Carney TJ, Kelsh RN, et al. (2005) Hedgehog signaling is required for cranial neural crest morphogenesis and chondrogenesis at the midline in the zebrafish skull. Development 132: 3977–3988. [DOI] [PubMed] [Google Scholar]
- 11. Swartz ME, Nguyen V, McCarthy NQ, Eberhart JK (2012) Hh signaling regulates patterning and morphogenesis of the pharyngeal arch-derived skeleton. Dev Biol 369: 65–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Swartz ME, Sheehan-Rooney K, Dixon MJ, Eberhart JK (2011) Examination of a palatogenic gene program in zebrafish. Developmental Dynamics 240: 2204–2220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Clouthier DE, Schilling TF (2004) Understanding endothelin-1 function during craniofacial development in the mouse and zebrafish. Birth Defects Res C Embryo Today 72: 190–199. [DOI] [PubMed] [Google Scholar]
- 14. Thomas T, Kurihara H, Yamagishi H, Kurihara Y, Yazaki Y, et al. (1998) A signaling cascade involving endothelin-1, dHAND and msx1 regulates development of neural-crest-derived branchial arch mesenchyme. Development 125: 3005–3014. [DOI] [PubMed] [Google Scholar]
- 15. Miller CT, Yelon D, Stainier DY, Kimmel CB (2003) Two endothelin 1 effectors, hand2 and bapx1, pattern ventral pharyngeal cartilage and the jaw joint. Development 130: 1353–1365. [DOI] [PubMed] [Google Scholar]
- 16. Miller CT, Schilling TF, Lee K-H, Parker J, Kimmel CB (2000) sucker encodes a zebrafish endothelin-1 required for ventral pharyngeal arch development. Development 127: 3815–3828. [DOI] [PubMed] [Google Scholar]
- 17. Ozeki H, Kurihara Y, Tonami K, Watatani S, Kurihara H (2004) Endothelin-1 regulates the dorsoventral branchial arch patterning in mice. Mech Dev 121: 387–395. [DOI] [PubMed] [Google Scholar]
- 18. Alexander C, Zuniga E, Blitz IL, Wada N, Le Pabic P, et al. (2011) Combinatorial roles for BMPs and Endothelin 1 in patterning the dorsal-ventral axis of the craniofacial skeleton. Development 138: 5135–5146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Zuniga E, Rippen M, Alexander C, Schilling TF, Crump JG (2011) Gremlin 2 regulates distinct roles of BMP and Endothelin 1 signaling in dorsoventral patterning of the facial skeleton. Development 138: 5147–5156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wilson J, Tucker AS (2004) Fgf and Bmp signals repress the expression of Bapx1 in the mandibular mesenchyme and control the position of the developing jaw joint. Dev Biol 266: 138–150. [DOI] [PubMed] [Google Scholar]
- 21. Tucker AS, Al Khamis A, Sharpe PT (1998) Interactions between Bmp-4 and Msx-1 act to restrict gene expression to odontogenic mesenchyme. Dev Dyn 212: 533–539. [DOI] [PubMed] [Google Scholar]
- 22. Zuniga E, Stellabotte F, Crump JG (2010) Jagged-Notch signaling ensures dorsal skeletal identity in the vertebrate face. Development 137: 1843–1852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Dudas M, Sridurongrit S, Nagy A, Okazaki K, Kaartinen V (2004) Craniofacial defects in mice lacking BMP type I receptor Alk2 in neural crest cells. Mech Dev 121: 173–182. [DOI] [PubMed] [Google Scholar]
- 24. Liu W, Sun X, Braut A, Mishina Y, Behringer RR, et al. (2005) Distinct functions for Bmp signaling in lip and palate fusion in mice. Development 132: 1453–1461. [DOI] [PubMed] [Google Scholar]
- 25. Ko SO, Chung IH, Xu X, Oka S, Zhao H, et al. (2007) Smad4 is required to regulate the fate of cranial neural crest cells. Dev Biol 312: 435–447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Baek JA, Lan Y, Liu H, Maltby KM, Mishina Y, et al. (2011) Bmpr1a signaling plays critical roles in palatal shelf growth and palatal bone formation. Dev Biol 350: 520–531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Hu D, Marcucio RS (2009) A SHH-responsive signaling center in the forebrain regulates craniofacial morphogenesis via the facial ectoderm. Development 136: 107–116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Chiang C, Litingtun Y, Lee E, Young KE, Corden JL, et al. (1996) Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature 383: 407–413. [DOI] [PubMed] [Google Scholar]
- 29. Jeong J, Mao J, Tenzen T, Kottmann AH, McMahon AP (2004) Hedgehog signaling in the neural crest cells regulates the patterning and growth of facial primordia. Genes and Development 18: 937–951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Abzhanov A, Tabin CJ (2004) Shh and Fgf8 act synergistically to drive cartilage outgrowth during cranial development. Developmental Biology 273: 134–148. [DOI] [PubMed] [Google Scholar]
- 31. Lan Y, Jiang R (2009) Sonic hedgehog signaling regulates reciprocal epithelial-mesenchymal interactions controlling palatal outgrowth. Development 136: 1387–1396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Stanier P, Moore GE (2004) Genetics of cleft lip and palate: syndromic genes contribute to the incidence of non-syndromic clefts. Hum Mol Genet 13 Spec No 1: R73–81. [DOI] [PubMed] [Google Scholar]
- 33. Britanova O, Depew MJ, Schwark M, Thomas BL, Miletich I, et al. (2006) Satb2 haploinsufficiency phenocopies 2q32-q33 deletions, whereas loss suggests a fundamental role in the coordination of jaw development. Am J Hum Genet 79: 668–678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. FitzPatrick DR, Carr IM, McLaren L, Leek JP, Wightman P, et al. (2003) Identification of SATB2 as the cleft palate gene on 2q32-q33. Hum Mol Genet 12: 2491–2501. [DOI] [PubMed] [Google Scholar]
- 35. Dobreva G, Chahrour M, Dautzenberg M, Chirivella L, Kanzler B, et al. (2006) SATB2 is a multifunctional determinant of craniofacial patterning and osteoblast differentiation. Cell 125: 971–986. [DOI] [PubMed] [Google Scholar]
- 36. Leoyklang P, Suphapeetiporn K, Siriwan P, Desudchit T, Chaowanapanja P, et al. (2007) Heterozygous nonsense mutation SATB2 associated with cleft palate, osteoporosis, and cognitive defects. Hum Mutat 28: 732–738. [DOI] [PubMed] [Google Scholar]
- 37. Fish JL, Villmoare B, Kobernick K, Compagnucci C, Britanova O, et al. (2011) Satb2, modularity, and the evolvability of the vertebrate jaw. Evol Dev 13: 549–564. [DOI] [PubMed] [Google Scholar]
- 38. Sheehan-Rooney K, Palinkasova B, Eberhart JK, Dixon MJ (2010) A cross-species analysis of Satb2 expression suggests deep conservation across vertebrate lineages. Dev Dyn 239: 3481–3491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Westerfield M (1993) The Zebrafish Book; A guide for the laboratory use of zebrafish (Brachydanio rerio).
- 40. Lawson N, Weinstein BM (2002) In vivo imiagining of embryonic vascular development using transgenic zebrafish. Developmental Biology 248: 307–318. [DOI] [PubMed] [Google Scholar]
- 41. Collery RF, Link BA (2011) Dynamic smad-mediated BMP signaling revealed through transgenic zebrafish. Dev Dyn 240: 712–722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Schauerte HE, Eeden FJMv, Fricke C, Odenthal J, Strahle U, et al. (1998) Sonic hedgehog is not required for the induction of medial floor plate cells in the zebrafish. Development 125: 2983–2993. [DOI] [PubMed] [Google Scholar]
- 43. Varga ZM, Amores A, Lewis KE, Yan YL, Postlethwait JH, et al. (2001) Zebrafish smoothened functions in ventral neural tube specification and axon tract formation. Development 128: 3497–3509. [DOI] [PubMed] [Google Scholar]
- 44. Yelon D, Ticho B, Halpern ME, Ruvinsky I, Ho RK, et al. (2000) The bHLH transcription factor hand2 plays parallel roles in zebrafish heart and pectoral fin development. Development 127: 2573–2582. [DOI] [PubMed] [Google Scholar]
- 45. Alexander J, Rothenberg M, Henry GL, Stainier DY (1999) casanova plays an early and essential role in endoderm formation in zebrafish. Dev Biol 215: 343–357. [DOI] [PubMed] [Google Scholar]
- 46. Kikuchi Y, Agathon A, Alexander J, Thisse C, Waldron S, et al. (2001) casanova encodes a novel Sox-related protein necessary and sufficient for early endoderm formation in zebrafish. Genes Dev 15: 1493–1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Walker MB, Kimmel CB (2007) A two-color acid-free cartilage and bone stain for zebrafish larvae. Biotech Histochem 82: 23–28. [DOI] [PubMed] [Google Scholar]
- 48. Woo K, Fraser SE (1995) Order and coherence in the fate map of the zebrafish nervous system. Development 121: 2595–2609. [DOI] [PubMed] [Google Scholar]
- 49. Dobreva G, Dambacher J, Grosschedl R (2003) SUMO modification of a novel MAR-binding protein, SATB2, modulates immunoglobulin mu gene expression. Genes Dev 17: 3048–3061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Britanova O, Akopov S, Lukyanov S, Gruss P, Tarabykin V (2005) Novel transcription factor Satb2 interacts with matrix attachment region DNA elements in a tissue-specific manner and demonstrates cell-type-dependent expression in the developing mouse CNS. Eur J Neurosci 21: 658–668. [DOI] [PubMed] [Google Scholar]
- 51. Walker MB, Miller CT, Coffin Talbot J, Stock DW, Kimmel CB (2006) Zebrafish furin mutants reveal intricacies in regulating Endothelin1 signaling in craniofacial patterning. Dev Biol 295: 194–205. [DOI] [PubMed] [Google Scholar]
- 52. Walker MB, Miller CT, Swartz ME, Eberhart JK, Kimmel CB (2007) phospholipase C, beta 3 is required for Endothelin1 regulation of pharyngeal arch patterning in zebrafish. Dev Biol 304: 194–207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Balczerski B, Matsutani M, Castillo P, Osborne N, Stainier DY, et al.. (2011) Analysis of Sphingosine-1-phosphate signaling mutants reveals endodermal requirements for the growth but not dorsoventral patterning of jaw skeletal precursors. Dev Biol. [DOI] [PMC free article] [PubMed]
- 54. Holzschuh J, Wada N, Wada C, Schaffer A, Javidan Y, et al. (2005) Requirements for endoderm and BMP signaling in sensory neurogenesis in zebrafish. Development 132: 3731–3742. [DOI] [PubMed] [Google Scholar]
- 55. Bitgood MJ, McMahon AP (1995) Hedgehog and Bmp genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev Biol 172: 126–138. [DOI] [PubMed] [Google Scholar]
- 56. Zhang Z, Song Y, Zhao X, Zhang X, Fermin C, et al. (2002) Rescue of cleft palate in Msx1-deficient mice by transgenic Bmp4 reveals a network of BMP and Shh signaling in the regulation of mammalian palatogenesis. Development 129: 4135–4146. [DOI] [PubMed] [Google Scholar]
- 57. Zhang Y, Zhang Z, Zhao X, Yu X, Hu Y, et al. (2000) A new function of BMP4: dual role for BMP4 in regulation of Sonic hedgehog expression in the mouse tooth germ. Development 127: 1431–1443. [DOI] [PubMed] [Google Scholar]
- 58. Huang X, Xu X, Bringas P Jr, Hung YP, Chai Y (2010) Smad4-Shh-Nfic signaling cascade-mediated epithelial-mesenchymal interaction is crucial in regulating tooth root development. J Bone Miner Res 25: 1167–1178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Foppiano S, Hu D, Marcucio RS (2007) Signaling by Bone Morphogenetic Proteins directs formation of an ectodermal signaling center that regulates craniofacial development. Developmental Biology in press. [DOI] [PMC free article] [PubMed]
- 60. Duench K, Franz-Odendaal TA (2012) BMP and Hedgehog signaling during the development of scleral ossicles. Dev Biol 365: 251–258. [DOI] [PubMed] [Google Scholar]
- 61. Dassule HR, McMahon AP (1998) Analysis of epithelial-mesenchymal interactions in the initial morphogenesis of the mammalian tooth. Dev Biol 202: 215–227. [DOI] [PubMed] [Google Scholar]
- 62. Clouthier DE, Hosoda K, Richardson JA, Williams SC, Yanagisawa H, et al. (1998) Cranial and cardiac neural crest defects in endothelin-A receptor-deficient mice. Development 125: 813–824. [DOI] [PubMed] [Google Scholar]
- 63. Yanagisawa H, Clouthier DE, Richardson JA, Charite J, Olson EN (2003) Targeted deletion of a branchial arch-specific enhancer reveals a role of dHAND in craniofacial development. Development 130: 1069–1078. [DOI] [PubMed] [Google Scholar]
- 64. Vieux-Rochas M, Mantero S, Heude E, Barbieri O, Astigiano S, et al. (2010) Spatio-temporal dynamics of gene expression of the Edn1-Dlx5/6 pathway during development of the lower jaw. Genesis 48: 262–373. [DOI] [PubMed] [Google Scholar]
- 65. Sato T, Kurihara Y, Asai R, Kawamura Y, Tonami K, et al. (2008) An endothelin-1 switch specifies maxillomandibular identity. Proc Natl Acad Sci U S A 105: 18806–18811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Miller CT, Swartz ME, Khuu PA, Walker MB, Eberhart JK, et al. (2007) mef2ca is required in cranial neural crest to effect Endothelin1 signaling in zebrafish. Dev Biol 308: 144–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Abe M, Ruest LB, Clouthier DE (2007) Fate of cranial neural crest cells during craniofacial development in endothelin-A receptor-deficient mice. Int J Dev Biol 51: 97–105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Nair S, Li W, Cornell R, Schilling TF (2007) Requirements for Endothelin type-A receptors and Endothelin-1 signaling in the facial ectoderm for the patterning of skeletogenic neural crest cells in zebrafish. Development 134: 335–345. [DOI] [PubMed] [Google Scholar]
- 69. Bonilla-Claudio M, Wang J, Bai Y, Klysik E, Selever J, et al. (2012) Bmp signaling regulates a dose-dependent transcriptional program to control facial skeletal development. Development 139: 709–719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Vokes SA, Ji H, McCuine S, Tenzen T, Giles S, et al. (2007) Genomic characterization of Gli-activator targets in sonic hedgehog-mediated neural patterning. Development 134: 1977–1989. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Wash out of Dorsomorphin causes a partial restoration of Bmp signaling. (A) Untreated 30 hpf BRE:mKO2 embryos have rhobust transgene expression in the ventral pharyngeal arches (arrows). (B) Nearly all expression, except for some in the dorsal retina is lost following dorsomorphin treatment from 20–30 hpf. (C) At 36 hpf the expression of the BRE:mKO2 transgene closely resembles that observed at 30 hpf. (D) Embryos treated with dorsomorphin from 20–30 hpf and then washed out of the drug show a partial recovery of BRE:mKO2 expression in the ventral pharyngeal arches (arrows).
(TIF)