

Abstract
Aims
The role of kidney infiltrating T cells in the pathology of lupus nephritis is unclear. This study was undertaken to investigate whether CD4+ T cell responses to a surrogate mesangial antigen can initiate glomerulonephritis.
Methods
Ovalbumin was deposited in the glomerular mesangium of C57BL/6 (B6) mice using anti-α8 integrin immunoliposomes (α8ILs). This was followed by injection of activated ovalbumin-reactive CD4+ transgenic OT2 T cells. Trafficking of antigen-specific OT2 T cells to kidneys and lymph nodes was studied by flow cytometry. Glomerular pathology and immune cell infiltration was characterized by immunostaining. Role of CCR2 deficiency on T cell mediated glomerulonephritis was investigated using B6.ccr2−/− mice.
Results
α8ILs delivered ovalbumin specifically to the renal glomeruli. Adoptively transferred OT2 T cells preferentially accumulated in renal lymph nodes and in the renal cortex. Kidneys showed glomerular inflammation with recruitment of endogenous T cells, dendritic cells and macrophages. T cell mediated inflammation induced mesangial cell activation and an increase in glomerular MCP1 and fibronectin. The formation of inflammatory foci was driven by Ly6C monocytes and was CCR2 dependent.
Conclusions
The findings from this study show that T cells reactive with antigens in the mesangium are sufficient to initiate glomerular pathology. Antigen-specific CD4 T cells act by inducing glomerular MCP1 production which mediates recruitment of inflammatory monocytes resulting in glomerulonephritis. Thus, down modulation of T cell responses within the kidneys of lupus patients will be a beneficial therapeutic approach.
INTRODUCTION
CD4+T cells are important mediators of systemic lupus erythematosus (SLE) [1]. In addition to helping autoreactive B cells to produce autoantibodies, CD4+T cells are directly involved in the renal pathology of lupus. Our studies in New Zealand Mixed 2328 mice, a spontaneous mouse model of SLE, showed that mesangial immune complex deposition was accompanied by CD4+ T cell activation in the renal lymph nodes and glomerular T cell infiltration [2]. T cell receptor analyses indicated a local antigen-driven T cell response. Direct role for T cells in pathogenesis of lupus nephritis is also supported by the finding that inhibiting T cell co-stimulation increases the life span of nephritic mice. [3]. Yet, whether CD4+T cells mediate the initiation of renal injury is unclear.
In this study we tested the hypothesis that T cells responding to antigens in the mesangium can induce glomerulonephritis. Since the identity of antigens recognized by pathogenic T cells in lupus is not known, we used ovalbumin as a surrogate mesangial antigen. We have previously described anti-α8 integrin immunoliposomes (α8ILs), a unique methodology for targeted delivery to the glomerular mesangium [4, 5]. Intra venous injection of liposomes sized to ~100 nm in diameter enter the mesangial space through the fenestrated glomerular endothelium. The anti-α8-integrin antibody conjugated to the liposomes reacts with α8-integrin on the mesangial cell surface. This allows accumulation of liposomes in the glomerular mesangium and leads to delivery of the liposomal contents at the site. Thus, in the present study, ovalbumin was deposited in the mesangium using α8ILs and ability of anti-ovalbumin CD4+T cells responses to initiate glomerulonephritis was investigated.
METHODS
Preparation of liposomes
Anti-α8 integrin immunoliposomes were prepared as previously described [4]. To incorporate ovalbumin, the lipid mixture was hydrated either with chicken ovalbumin (Sigma Aldrich Corporation, St. Louis, MO) or fluorescein isothiocyanate conjugated ovalbumin (FITC-OVA) (Invitrogen, Carlsbad, CA) in water, lyophilized and rehydrated in 0.025M Tris, pH8.0. After repeated washing, the liposomal ovalbumin (OVA) content was estimated by modified BCA assay (Peirce Biotechnology, Rockford, IL). Liposomes were then conjugated to either a rabbit anti-α8 integrin antibody (OVA-α8ILs) or to rabbit IgG (control OVA-ILs).
Mice
Mice were maintained in a specific pathogen free mouse facility at the University of Virginia. All experimental procedures were approved by the Institutional Animal Care and Use Committee. Breeders for B6.Cg-Tg(TcrαTcrβ) 425Cbn/J (OT2) and B6.129S4-Ccr2tm1Ifc/J (B6.ccr2−/−) mice were obtained from Jackson Laboratories, Bar Harbor, ME. C57BL/6 (B6) mice were purchased from National Cancer Institute, Frederick, MD. OT2 transgenic mice express CD4+ T cell receptor transgene (Vα2, Vβ5) reactive with ovalbumin323–339 peptide and were used as donors for antigen specific T cells [6]. B6 and B6.ccr2−/− female mice were used as recipients.
Cell transfers
OT2 T cells were activated with ovalbumin323–339 peptide (Ana Spec, Fremont CA) for 72 hours using standard protocols [7], live cells were harvested over a Ficoll gradient, and injected i.v. into recipient mice. For some experiments, CD4+T cells were purified by negative selection (Miltenyi Biotec, Auburn, CA), activated with ovalbumin323–339 peptide and irradiated syngenic spleen cells. This method of stimulation yielded T helper 1 (Th1) deviated OT2 cells, and this was confirmed by detection of IFNγ in the culture supernatants at 24 and 48 hours after stimulation (data not shown). To investigate activation of OT2 cells in vivo, naïve OT2 T cells were isolated and injected i.v. into recipient mice. In all experiments, mice received 12 to14 million OT2 T cells.
Flow cytometry
Cell suspensions from spleens, lymph nodes, and kidneys were studied by flow cytometry using standard methods [2]. Antibodies conjugated to fluorochromes were obtained from eBiosciences, San Deigo, CA or BD Biosciences, San Jose, CA except as indicated. Cells were incubated with anti-CD16/32 (Clone 93) and OT2 T cell frequencies were analyzed by 4-color staining for CD45 (30-F11), CD4 (L3T4), T cell receptor Vα2 (B20.1) and Vβ5 (MR9-4). Kidney infiltrating macrophages and dendritic cells were studied by 6 color flow cytometry using antibodies to CD45 (30-F11), F4/80 (BM8), CD11b (M1/70), CD11c (HL3) and Ly6C (HK1.4, Abcam). Bromodeoxyuridine (BrdU) incorporation was measured by intracellular staining using anti-BrdU antibody (BU20A) followed by FITC conjugated F(ab)2anti-mouse IgG. Stained cells were acquired on 8-color BD FACS Calibur using Cytek dxP8 software (Cytek Technologies, Freemont, CA). Data were analyzed using FlowJo Software 9.1 (Tree Star Inc., Ashland, OR).
Immunofluorescence staining of isolated glomeruli and kidney
For detection of FITC-OVA, glomeruli were isolated, and stained as previously described [4]. Non-specific staining was blocked with normal rabbit serum, and incubated with goat anti-FITC antibody (Vector laboratories, Burlingame, CA). Bound antibody was detected with fluoresceinated rabbit anti-goat IgG (Vector laboratories, Burlingame, CA).
In the kidney, FITC-OVA was detected in frozen sections, fixed in cold acetone and observed under a fluorescence microscope. Infiltrating cells were characterized by immunostaining with antibodies to CD4 (GK1.5), CD11c (HL3), CD68 (AbD serotec, Raleigh, NC) and MHC II (M5/114.15.2) as previously described [2, 8]. MCP1 was detected using rabbit anti-MCP1 antibody (Peprotech, Rocky Hill, NJ) followed by PE-conjugated anti-rabbit IgG (Jackson ImmunoResearch Laboratories Inc, West Grove, PA). Glomerular fibronectin was detected with rabbit anti-fibronectin antibody (Millipore Corporation, Sunnyvale CA) [8]. All fluorescence images were captured on a Zeiss Confocal Microscope or Zeiss Apotome fluorescence microscope, and the images were analyzed using LSM5 and Axiovision 4.7 software (Carl Zeiss MicroImaging LLC, Thornwood, NY) respectively.
Renal histopathology
Kidneys were collected in formalin and processed for paraffin embedding. Sections (3μM thickness) were stained with hematoxylin and eosin and evaluated for pathologic changes as previously described [2].
Statistical analysis
Statistical analyses were done by Student’s t test or ANOVA with Bonferroni post test using GraphPad Prism 5.0 (GraphPad Software Inc., La Jolla, CA).
RESULTS
Deposition of ovalbumin as a surrogate self antigen in the glomerular mesangium
To study the kinetics of glomerular delivery, FITC-OVA loaded α8ILs were injected intravenously into B6 mice. FITC-OVA was rapidly deposited in the glomeruli and could be detected by indirect immunofluorescence 6 hrs after injection (Figure 1A). The amount of FITC-OVA in the glomeruli increased by day 5 (Figure 1B). In the kidney, FITC-OVA was restricted to glomeruli and showed a typical mesangial pattern of distribution (Figure 1C). FITC-OVA loaded liposomes conjugated to rabbit IgG (control OVA-ILs) did not traffic to the renal glomeruli.
FIGURE 1. Accumulation of ovalbumin (OVA) in the kidney is rapid, specific to the glomeruli, and has a mesangial distribution.

FITC-OVA-α8ILs or control FITC-OVA-ILs were injected into B6 mice and delivery of FITC-OVA was studied at different time points. A: Representative photomicrographs show accumulation of FITC-OVA (arrows) in glomerular isolates at 6 hrs after injection. B: Frozen sections of the kidney on day 5 after transfer showed FITC-OVA restricted to the glomeruli within the kidney. (g). C: Higher magnification showed a typical mesangial distribution of FITC-OVA. Scale bar = 50 microns.
Activated OT2 T cells target mesangial ovalbumin and induce glomerulonephritis
Mice were given 2 intravenous injections of OVA-α8ILs or control liposomes, 12 hours apart followed by a single injection of activated OT2 splenocytes as shown in figure 2. Kidneys studied on day 8 after transfer showed inflammation in the renal cortex predominantly in glomeruli and peri-glomerular regions (Figure 3A). These results were further confirmed by adoptive transfer of purified and activated OT2 T cells (94% purity, 99% Vα2 Vβ5 positive). Kidneys were harvested on days 3, 8 and 14 after cell transfer and infiltrating cells analyzed by flow cytometry showed an increased frequency of OT2 cells on days 8 and 14 (Figure 3B). This was accompanied by an increase in the CD4+T cells infiltrating the kidney. Interestingly, OT2 cells were only 3 to 8% of the infiltrating CD4+ cells. Thus, trafficking of OT2 cells into the kidney led to the recruitment of endogenous, non-OT2, CD4+T cells.
FIGURE 2. Experimental design.
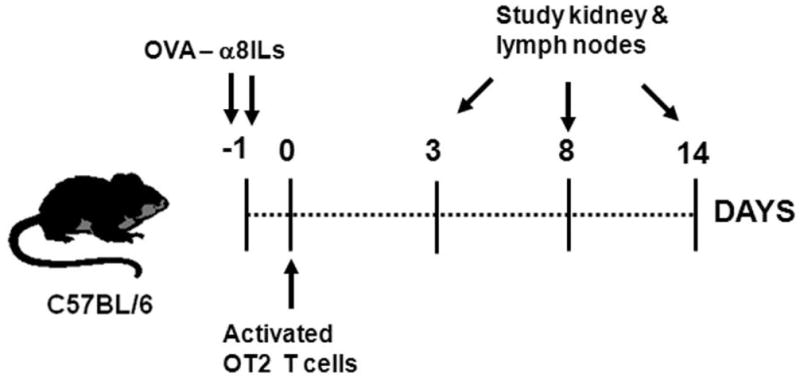
Female mice were injected with OVA-α8ILs in the tail vein on day -1. The OVA-α8ILs traffic to the renal glomeruli and deposit OVA in the glomerular mesangium. This was followed by i.v. injection of activated OT2 cells on day 0. Mice were euthanized on days 3, 8, or 14 after OT2 cell injection and kidneys and lymph nodes were studied. Mice injected with only α8ILs (lacking OVA), only OVA-α8ILs, or only activated OT2 T cells were used as controls as indicated in each experiment.
FIGURE 3. Ovalbumin delivered to glomeruli by OVA-α8ILs is a target for activated antigen specific T cells.

A: OVA-α8ILs injection followed by transfer of OT2 cells induced glomerulonephritis with inflammatory cell infiltrates (arrows) on day 8 after cell transfer. This was not seen in control mice injected with control OVA-ILs and OT2 cells. B: Frequencies of OT2 cells (of CD4 gate) and total CD4+ T cells (of CD45 gate) infiltrating the kidneys were analyzed by flow cytometry on days 3, 8, and 14 and show increased accumulation in mice injected with OVA-α8ILs and OT2 T cells (●) compared to OT2 injected control mice (○). C: Mice injected with activated OT2 cells developed infiltration of CD4+ T cells and inflammation following glomerular ovalbumin deposition. Representative photomicrographs of kidneys from B6 mice, injected with OVA-α8ILs followed by OT2 cells and sacrificed 3 days or 14 days later are shown. Sections were stained for CD4+ T cells (red) and MHCII antigen presenting cells (green). Inflammatory foci (arrows) with clusters of CD4+ T cell and MHCII+ antigen presenting cells were seen in the renal cortex on day 3 with peri-glomerular inflammation on day 14. A higher magnification photomicrograph of CD4+ T cells infiltrating into the glomeruli is also shown. Scale bar = 50 microns. Control mice injected with OT2 cells alone did not develop inflammatory infiltrates.
Immunostaining showed aggregates of CD4+T cells and MHCII antigen presenting cells in the renal cortex on day 3; with significant intra-glomerular and peri-glomerular infiltrates on days 8 and 14. The CD11c+ dendritic cells (Figure 4A) and CD68 and F4/80 positive macrophages (Figure 4B) in the infiltrates were the predominant MHC II+ cells, suggesting that they are the antigen-presenting cells (insets 4A and 4B respectively). Inflammatory foci were not seen in control mice.
FIGURE 4. Inflammatory cells and glomerular responses in OT2 T cell-mediated glomerulonephritis.

Representative photomicrographs of renal cortex from mice treated with OT2 T cells or OVA-α8ILs and OT2 T cells on day 8 after cell transfer stained for A: dendritic cells, B: macrophages, and C: fibronectin. Arrows indicate clusters of CD11c (red) cells co-expressing MHCII (green) (A). Inset shows intra-glomerular infiltration of dendritic cells. Macrophages detected by CD68 (red) and F4/80 (green) were increased and are indicated with arrows (B). Inset B shows that intraglomerular CD68 (red) macrophages infiltrating the glomeruli were activated and express MHC II (green). Representative photomicrographs from control mice injected with OT2 T cells alone did not show inflammatory infiltrates. Results are representative of 2 independent experiments. Kidney sections stained for fibronectin and images captured from four different areas of the renal cortex in each mouse at 20x magnification and the intensity of fluorescent staining in glomeruli was evaluated (C). Representative images show that fibronectin deposition in glomeruli (arrows) was increased in mice injected with OVA-α8ILs and OT2 T cells compared to controls given OT2 cells alone. Scale bars = 50 microns.
To evaluate the pathogenic effect of glomerular inflammation, kidneys were stained for fibronectin, a component of the extracellular matrix, and an early marker for mesangial cell activation [9]. On day 8, glomerular fibronectin was increased in mice given OVA-α8ILs and OT2 cells (Figure 4C). At these time points, the kidney function was normal. Thus, activated CD4+T cells reacting to mesangial antigens can initiate mesangial injury and glomerulonephritis.
Mesangial antigens are presented to activated T cells in renal lymph nodes
Lymph nodes and spleens were harvested from mice to study trafficking of OT2 T cells (Figure 5). On day 8 after transfer, mice with mesangial ovalbumin consistently showed an increased frequency of OT2 cells in the renal lymph nodes (p<0.01) compared to controls (figure 5B). Furthermore, the enrichment of OT2 cells was specific to renal lymph nodes compared to the non-renal lymph nodes (p<0.001). This enrichment was not seen in control mice given activated OT2 cells alone. In the spleen, frequencies of OT2 cells in mice injected with OVAα8ILs and activated OT2 cells were 11.6±1.7 (absolute numbers 1.06±0.15×106; mean±SEM) and were not significantly higher than mice injected with activated OT2 cells alone with an average frequency of 8.3±0.5 (absolute numbers 0.85±0.15 ×106). These data suggest that the presentation of ovalbumin was occurring predominantly in the kidney and local lymph nodes and provides additional evidence that delivery by OVA-α8ILs was specific to the kidney.
FIGURE 5. OT2 T cells traffic preferentially to renal lymph nodes in mice with mesangial ovalbumin.

A: Trafficking of OT2 T cells was studied by flow cytometry in renal and non-renal lymph nodes. Gates for CD4 and OT2 T cells (Vα2 Vβ5) are shown. B: OT2 frequencies in renal and non-renal lymph nodes from mice injected with OVA -α8ILs and OT2 cells or with OT2 cells alone. Data are from one of five different experiments. Each data point indicates one mouse. p values calculated by ANOVA with Bonferroni’s post test.
Recruitment of Ly6C+ monocytes is critical for glomerular inflammation
Monocyte Chemoattractant Protein 1 (MCP1) in the glomerulus is an early indicator of inflammation [10]. MCP1 mediates macrophage recruitment through its receptor, chemokine CC motif receptor 2 (CCR2) [11]. To study the role of macrophages and the MCP1-CCR2 axis, OVA-α8ILs were injected into ccr2+/+ sufficient (wild type) and ccr2−/− mice, and followed by adoptive transfer of activated OT2 cells. MCP1 expression measured by qPCR was increased in the kidneys from both ccr2+/+ (2.8±1.0 fold; mean ± SD, n=5) and ccr2−/− mice (6.0±2.5 fold; mean ± SD, n=5) over controls injected with activated OT2 cells alone. These results were confirmed by immunostaining for MCP1 protein in the glomeruli (Figure 6). Control mice did not show glomerular MCP1 expression. Flow cytometry analyses showed that CD4+T cell infiltration was comparable between WT and ccr2−/− mice (Figure 7A left panel) and there was no significant difference in the recruitment of antigen specific OT2 cells (data not shown) into the kidney. Further, the frequencies of CD11b macrophages and CD11c dendritic cells were comparable (data not shown). However, there was a small though consistent, statistically significant reduction in the frequency of Ly6C inflammatory monocytes in ccr2−/− mice (n=5/grp; p=0.0096) (figure 7A right panel). This was reproduced in an additional experiment p= 0.019; n=4/grp, data not shown). Representative gates used for analyses of Ly6C cells are shown (Figure 7B). Surprisingly, despite comparable OT2 and endogenous CD4+T cell recruitment, ccr2−/− mouse kidney showed a complete lack of inflammatory foci (Figure 7C). Thus, trafficking of antigen specific and endogenous T cells into the kidney was not affected by CCR2 deficiency. However, recruitment of Ly6C cells was important for the development of glomerulonephritis.
FIGURE 6. Activated OT2 cells induce MCP1 expression in glomeruli of mice injected with OVA-α8ILs.
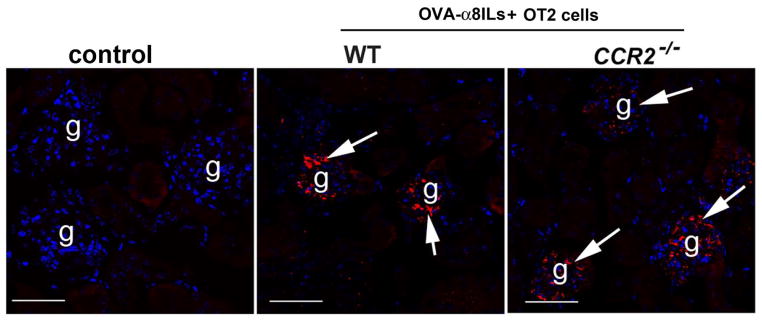
Increased expression of glomerular MCP1 (red) was seen in B6. ccr2+/+ (WT) and ccr2−/−. MCP1 was not detected in glomeruli of wild type mice given OT2 cells alone. Note that the MCP1 expression is predominantly in the glomeruli.
FIGURE 7. Ly6C+ monocyte recruitment is required for glomerular inflammation.

B6. ccr2+/+ (WT) and ccr2−/− mice were injected with OVA-α8ILs followed by activated OT2 cells and kidneys were studied on day 8 after cell transfer. A: In the CD45 gates, frequencies of CD4+ T cells (left panel) were comparable between the two groups. However, Ly6C+ cells (right panel) were reduced in ccr2−/− mice. B: Representative gates for analysis of CD45+ and Ly6C+ cells infiltrating the kidney are shown. C: Immunostaining for CD4 (red) and MHC II (green) cells in the kidney in showed inflammatory foci (arrows) in B6 ccr2+/+ mice. These foci were absent in ccr2−/− mice, although CD4+T cells (red) were seen scattered in the renal cortex. Scale bar = 50 microns
Mesangial antigens traffic to the renal lymph nodes and activate antigen specific T cells
The results presented above show that mesangial antigens are presented to activated CD4+ T cells in the kidney leading to recruitment of bystander T cells and inflammatory macrophages inducing glomerular inflammation. This is associated with activation of antigen specific T cells in the renal lymph nodes. To investigate whether mesangial antigens drain to the renal lymph nodes, naïve OT2 cells were transferred into mice injected with OVA-α8ILs. Control mice received only naïve OT2 cells. The mice were given a single injection of BrdU i.p (2mg/mouse), 12 hours prior to sacrifice on day 3 after transfer (Figure 8A). Accumulation of naïve OT2 cells in the renal lymph nodes was accompanied by increased proliferation in mice injected with OVA-α8ILs compared to controls (Figure 8B). At this point, few OT2 cells were detected in the kidneys of either group and none of them showed BrdU uptake (data not shown). These results suggest the glomerular antigens drain into the renal lymph nodes or are carried there by antigen presenting cells and are presented to antigen specific CD4 T cells.
FIGURE 8. Proliferation of naive OT2 cells in renal lymph nodes in response to mesangial antigens.
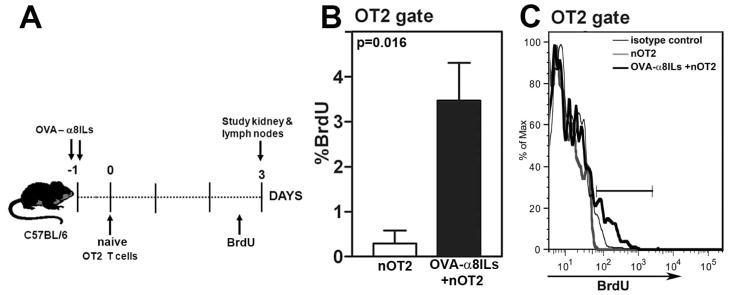
A: Female mice were injected with OVA-α8ILs in the tail vein followed by i.v. injection of naive OT2 (nOT2) cells on day 0. Mice injected with only naive OT2 T cells were used as controls. Mice were injected with BrdU 12 hours before sacrifice on day 3 and kidneys and lymph nodes were studied. B: Frequencies of OT2 cells incorporating BrdU in kidney lymph nodes from mice injected with nOT2 cells alone or OVA-α8ILs and nOT2 cells (n=5 mice/group; p=0.016). Data are from one of two independent experiments. C: Histogram showing BrdU incorporation in OT2 T cells in one representative mouse from each group. Pooled lymph node cells stained with isotype control antibody for BrdU staining is also shown.
DISCUSSION
Patient and experimental mouse studies suggest that pathology in lupus glomerulonephritis initiates in the mesangium [12, 13] and CD4+ T cells participate in the pathogenesis of lupus nephritis [2, 3]. However, the lack of information on target antigens in lupus nephritis is a major barrier in investigating mechanisms of T cells in initiating disease. In this study we demonstrate that activated antigen-specific CD4+T cells, directed against a surrogate mesangial antigen, are sufficient to initiate the typical inflammatory lesions consisting of peri-glomerular macrophage, dendritic cell and T cell infiltrates. This induces mesangial cell activation and extracellular matrix production. These changes represent early events and at this time point, are not associated with loss of kidney function. This pattern of glomerulonephritis mimics the patterns reported in some lupus patients with CD4+T cell infiltrates forming focal or circumferential caps around glomeruli in the renal cortex [14]. The novelty of this study lies in being able to target antigens that are in the mesangium with antigen specific T cells in immune sufficient mice, mimicking the location and pathology of spontaneous glomerulonephritis. This will allow us to dissect the individual contributions of inflammatory mediators in mesangio-proliferative glomerulonephritis.
Use of antigen-specific T cells in the induction of glomerulonephritis have been previously described. Mesangial deposition of ovalbumin polymers followed by injection of ovalbumin reactive T cell lines in SCID mice showed that Th1 T cells could traffic into the kidney leading to glomerulonephritis [15]. Transgenic mice expressing ovalbumin and hen egg lysozyme under the nephrin promoter in podocytes (NOH) mice have been recently used to investigate glomerulonephritis [16]. Repeated transfers of CD8+ OT1 T cells co-injected with activated CD4+ OT2 T cells in NOH mice resulted in glomerulonephritis. However, injection of either cell type alone failed to cause disease. These results are different than our study, where activated CD4+ OT2 cells were sufficient for glomerulonephritis. Other studies have proposed glomerular basement membrane (GBM) proteins like collagen [17] as potential targets for autoreactive T cells using rats primed with pertussis toxin to facilitate trafficking of T cells into non-lymphoid tissues; or antigens trapped on the GBM using Rag-deficient recipient mice facilitating homeostatic expansion of autoreactive T cells [18]. In our experiments, glomerular inflammation was comparable in mice with or without priming with pertussis toxin (data not shown) and was seen in normal B6 recipient mice. Although the role of podocyte antigens in spontaneous glomerulonephritis is unclear [19], together these studies show that the glomerular location of the target antigen clearly influences CD4 T cell mediated pathology.
This study focuses on the early events in initiation of glomerulonephritis. Whether this sequence of events occurs with each renal flare would be of clinical relevance and would provide the rationale for preventing local T cell responses and macrophage trafficking into the kidney as important strategies for therapeutic intervention.
Acknowledgments
The authors would like to thank Mr. Saleh Mohammad for outstanding technical assistance. This work was supported by grants from National Institutes of Health R01DK069769 (H.B.), ARRA supplement to R01DK069769 (H.B.), R01AI079621 (USD), DK76095 (W.K.B.) and from Alliance Lupus for Research #113300 (H.B).
Footnotes
Disclosure: The authors have no financial conflicts of interest to disclose.
References
- 1.Wofsy D. Treatment of murine lupus with anti-CD4 monoclonal antibodies. Immunol Ser. 1993;59:221–236. [PubMed] [Google Scholar]
- 2.Bagavant H, Deshmukh US, Wang H, Ly T, Fu SM. Role for nephritogenic T cells in lupus glomerulonephritis: progression to renal failure is accompanied by T cell activation and expansion in regional lymph nodes. J Immunol. 2006;177:8258–8265. doi: 10.4049/jimmunol.177.11.8258. [DOI] [PubMed] [Google Scholar]
- 3.Schiffer L, Sinha J, Wang X, Huang W, von Gersdorff G, Schiffer M, Madaio MP, Davidson A. Short term administration of costimulatory blockade and cyclophosphamide induces remission of systemic lupus erythematosus nephritis in NZB/W F1 mice by a mechanism downstream of renal immune complex deposition. J Immunol. 2003;171:489–497. doi: 10.4049/jimmunol.171.1.489. [DOI] [PubMed] [Google Scholar]
- 4.Scindia Y, Deshmukh U, Thimmalapura PR, Bagavant H. Anti-alpha8 integrin immunoliposomes in glomeruli of lupus-susceptible mice: a novel system for delivery of therapeutic agents to the renal glomerulus in systemic lupus erythematosus. Arthritis Rheum. 2008;58:3884–3891. doi: 10.1002/art.24026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Scindia YM, Deshmukh US, Bagavant H. Mesangial pathology in glomerular disease: targets for therapeutic intervention. Adv Drug Deliv Rev. 2010;62:1337–1343. doi: 10.1016/j.addr.2010.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Barnden MJ, Allison J, Heath WR, Carbone FR. Defective TCR expression in transgenic mice constructed using cDNA-based alpha- and beta-chain genes under the control of heterologous regulatory elements. Immunol Cell Biol. 1998;76:34–40. doi: 10.1046/j.1440-1711.1998.00709.x. [DOI] [PubMed] [Google Scholar]
- 7.Bagavant H, Adams S, Terranova P, Chang A, Kraemer FW, Lou Y, Kasai K, Luo AM, Tung KSK. Autoimmune ovarian inflammation triggered by proinflammatory (Th1) Tcells is compatible with normal ovarian function in mice. Biol Reprod. 1999;61:635–642. doi: 10.1095/biolreprod61.3.635. [DOI] [PubMed] [Google Scholar]
- 8.Bagavant H, Scindia YM, Nackiewicz D, Nandula SR, Doran A, Cutchins A, Oldham S, Deshmukh U, McNamara C. Deficiency of a transcriptional regulator, Inhibitor of differentiation 3 (Id3), induces glomerulonephritis in Apolipoprotein E deficient mice: a model linking hyperlipidemia and renal disease. Am J Pathol. 2011;107:651–660. doi: 10.1016/j.ajpath.2011.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Couchman JR, Beavan LA, McCarthy KJ. Glomerular matrix: synthesis, turnover and role in mesangial expansion. Kidney Int. 1994;45:328–335. doi: 10.1038/ki.1994.42. [DOI] [PubMed] [Google Scholar]
- 10.Pérez de Lema G, Maier H, Nieto E, Vielhauer V, Luckow B, Mampaso F, Schlöndorff D. Chemokine expression precedes inflammatory cell infiltration and chemokine receptor and cytokine expression during the initiation of murine lupus nephritis. J Am Soc Nephrol. 2001;12:1369–1382. doi: 10.1681/ASN.V1271369. [DOI] [PubMed] [Google Scholar]
- 11.Charo IF, Myers SJ, Herman A, Franci C, Connolly AJ, Coughlin SR. Molecular cloning and functional expression of two monocyte chemoattractant protein 1 receptors reveals alternative splicing of the carboxyl-terminal tails. Proc Nat Acad Sci USA. 1994;91:2752–2756. doi: 10.1073/pnas.91.7.2752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Pollak VE, Pirani CL, Schwartz FD. The natural history of the renal manifestations of systemic lupus erythematosus. J Lab Clin Med. 1964;63:537–550. [PubMed] [Google Scholar]
- 13.Weening JJ, D’Agati VD, Schwartz MM, Seshan SV, Alpers CE, Appel GB, Balow JE, Bruijn JA, Cook T, Ferrario F, Fogo AB, Ginzler EM, Hebert L, Hill G, Hill P, Jennette JC, Kong NC, Lesavre P, Lockshin M, Looi LM, Makino H, Moura LA, Nagata M International Society of Nephrology Working Group on the Classification of Lupus Nephritis; Renal Pathology Society Working Group on the Classification of Lupus Nephritis. The classification of glomerulonephritis in systemic lupus erythematosus revisited. Kidney Int. 2004;65:521–530. doi: 10.1111/j.1523-1755.2004.00443.x. [DOI] [PubMed] [Google Scholar]
- 14.Winchester R, Wiesendanger M, Zhang HZ, Steshenko V, Peterson K, Geraldino-Pardilla L, Ruiz-Vazquez E, D’Agati V. Immunologic characteristics of intrarenal T cells: Trafficking of expanded CD8 T cell b-chain clonotypes in progressive lupus nephritis. Arthritis Rheum. 2011;64:1589–1600. doi: 10.1002/art.33488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Radeke HH, Tschernig T, Karulin A, Schumm G, Emancipator SN, Resch K, Tary-Lehmann M. CD4+ T cells recognizing specific antigen deposited in glomeruli cause glomerulonephritis-like kidney injury. Clin Immunol. 2002;104:161–173. doi: 10.1006/clim.2002.5246. [DOI] [PubMed] [Google Scholar]
- 16.Heymann F, Meyer-Schwesinger C, Hamilton-Williams EE, Hammerich L, Panzer U, Kaden S, Quaggin SE, Floege J, Gröne HJ, Kurts C. Kidney dendritic cell activation is required for progression of renal disease in a mouse model of glomerular injury. J Clin Invest. 2009;119:1286–1297. doi: 10.1172/JCI38399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wu J, Hicks J, Borillo J, Glass WF, 2nd, Lou YH. CD4(+) T cells specific to a glomerular basement membrane antigen mediate glomerulonephritis. J Clin Invest. 2002;109(4):517–524. doi: 10.1172/JCI13876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Summers SA, Steinmetz OM, Li M, Kausman JY, Semple T, Edgtton KL, Borza H, Holdworth SR, Kitching AR. Th1 and Th17 cells induce proliferative glomerulonephritis. J Am Soc Nephrol. 2009;20:2518–2524. doi: 10.1681/ASN.2009030337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kim AH, Markiewicz MA, Shaw AS. New roles revealed for T cells and DCs in glomerulonephritis. J Clin Invest. 2009;119:1074–1076. doi: 10.1172/JCI39071. [DOI] [PMC free article] [PubMed] [Google Scholar]